COMMENTS: De kontroversjepartij fan dit papier befettet de folgjende:
Der binne in oantal elektrophysiologysk basearre stúdzjes, troch de groep fan Prause dy't inkele bewiis leverje dat seksueel begearte, net hyperseksualiteit, selsregulearje fan seksueel oprop [92]. Dizze ûndersikers jouwe yn oare wurken dy't subjects rapporteare problemen regelje fan harren besjen fan fisuele seksuele stimulaasjes (VSS) dy't ek hegere seksueel winsken litte sjen litte leechtere posityf potinsjes (LPP) yn antwurd op VSS. De auteurs suggerearje dat dit patroan ferskynt ferskilt fan substansykmodellen [93].
De twa EEG-papieren fan 'e groep fan Prause binne wiidweidich bekritiseare yn peer-reviewed tydskriften en troch oaren (ynklusyf YBOP). Yn 'e realiteit binne de befiningen yn beide papieren konsekwint mei it ferslavingsmodel, nettsjinsteande de dubieuze konklúzjes fan har auteurs. Foar de 2013 EEG-stúdzje sjoch:
- Neat korreleart mei neat yn 'e nije porno-stúdzje fan SPAN Lab (2013)
- Peer-oersjoch krityk: 'Heulewrâld', of 'gewoan' in taflecht? In reaksje nei Steele et al. (2014) troch Donald L. Hilton, Jr., MD,
- Peer-oersjoch krityk: "Neurowittenskip fan ferslaving op ynternetpornografy: in resinsje en fernijing" (2015)
- Peer-revidearre analyze: "Neurale korrelaten fan seksuele reaktyfens yn yndividuen mei en sûnder twangmjittige seksuele gedrach" (2014)
Foar it 2015 EEG stúdzje sjoch:
- Krityk op "Modulaasje fan lette positive potensjes troch seksuele bylden yn brûkers fan problemen en kontrôles yn striid mei 'Pornoferslaving' (2015)"
- Peer-oersjoch krityk: Neurology fan 'e Internet Pornografy Addiction: In Review en Update (2015)
Soe it net geweldich wêze as ûndersikers de stúdzjes eins lêze dy't se sitearje yn stee fan gewoan de kopteksten fan 'e haad auteur te papegaaien?
LINK TO PUBLIKATION
Published: Oktober 12, 2015 (sjoch histoarje)
DOI: 10.7759 / cureus.348
Boarne bewurkje Blum K, Badgaiyan RD, Gold MS (oktober 12, 2015) Hyperseksualiteit Addiction en weromfal: Phenomenology, Neurogenetics and Epigenetics. Cureus 7 (10): e348. doi: 10.7759 / cureus.348
Abstract
Hyperseksualiteit is definieare as abnormaal ferhege seksuele aktiviteit. Epidemiologyske en klinyske stúdzjes hawwe oantoand dat dizze net-parafile tastân bestiet út "oermjittige" seksuele gedrach en steurnissen begelaat troch persoanlike need en sosjale en medyske syklike. It is in heul kontroversjeel en polityk ûnderwerp yn termen fan hoe it it bêste te kategorisearjen is as ferlykber as net ferlykber mei ferslavend gedrach ynklusyf substansmisbrûk. Hyperseksuele steuring wurdt konseptualisearre as in net-parafile seksuele winsksteuring mei ympulsiviteit. Pathophysiologyske perspektiven omfetsje dysregulaasje fan seksuele opwekking en winsk, seksuele ympulsiviteit, en seksuele compulsiviteit. De kearn accumbens, leit binnen it ventral striatum, bemiddelt de fersterkende effekten fan drugs fan misbrûk, lykas kokaïne, alkohol, nikotine, en iten as muzyk. Eins wurdt leaud dat dizze struktuer gedrach ferplichtet troch stimulearjende stimulâns. Dit gedrach omfettet natuerlike beleanningen lykas iten, drinken, seksueel gedrach, en ferkennende beweging. In essensjele regel fan positive fersterking is dat motorreaksjes tanimme yn grutte en krêft as folge troch in beleanjend barren. Hjir hypotezearje wy dat d'r in mienskiplik meganisme fan aksje (MOA) is foar de krêftige effekten dy't medisinen, muzyk, iten en seks hawwe op minsklike motivaasje. De minsklike driuw foar de trije needsaaklike motivaasjegedrach "honger, toarst en seks" kin allegear mienskiplike molekulêre genetyske antykedinten hawwe dy't, as beheind, liede ta ôfwikende gedragingen. Wy hypoteze dat basearre op in oerfloed fan wittenskiplike stipe hyperseksuele aktiviteit yndie is lykas drugs, iten en muzyk dy't mesolimbyske belestingsskeakelingen aktivearje. Boppedat hawwe dopaminergyske genen en mooglik oare kandidaat neurotransmitter-relatearre genpolymorfismen ynfloed op sawol hedonyske as anhedonyske gedrachsresultaten. D'r is net folle bekend oer sawol de genetika as epigenetika fan hyperseksualiteit yn 'e hjoeddeistige literatuer. Wy ferwachtsje lykwols dat takomstige stúdzjes basearre op beoardielingen mei klinyske ynstruminten yn kombinaasje mei genotyping fan seksferslaafden bewiis sille leverje foar spesifike klustering fan seksuele typologyen mei polymorfe ferienings. D'r binne wat stúdzjes west mei elektrofysiologyske techniken dy't de opfetting net stypje dat hyperseksualiteit yndie fergelykber is mei substansmisbrûk en oare gedrachsferslavingen. De auteurs stimulearje ek sawol klinyske as akademyske wittenskippers om ûndersyk te begjinnen mei neuroimaging-ark om natuerlike dopaminergyske agonistyske aginten te ûndersiikjen op spesifike genpolymorfismen om hyperseksueel gedrach te "normalisearjen".
Yntroduksje en eftergrûn
Geweldich is hyperseksuele gedrach yn 'e klinyske en ûndersyksynstellingen yn' e ôfrûne tsien jier dokumentearre [1]. Benjamin Rush, in dokter ien fan 'e oprjochte heit fan' e Feriene Steaten, klonklik dokumentearre oerfallende seksuele gedrach [2] Richard von Krafft-Ebing, in 19e-ieuske westerske Europeesk pionier-seksolooch, en Hirshfeld yn 1948, beide stipe it wurk [3-4]. De basishâldende fan dizze ûndersikers hat besocht dat hyperseksualiteit persistente sosjale ôfwiking fan seksueel gedrach (s) yn beide manlju en froulju mei in heulende seksueel appetitens is maladaptive. Yn 1975 karakterisearre de Stroller de betingst as Don Juanisme [5]. Yn 1969 suggerearre Allen satyriasis foar manlju en nymphomania yn froulju dy't stipe wurde troch Ellis en Sagarin [6-7], Ek al is hyperseksualiteit net opnommen as in psychiatryske diagnoaze yn DSM, kin wurk fan in oantal hjoeddeistige ûndersikers, lykas Kafka, Reid, Bancroft, har kollega's en de oanstriid fan 'e World Health organisaasje liede ta opnimmen fan dizze tastân as in aparte diagnostyske entiteit [8-11].
Resinsje
Literatuermetoade
De Medline-database, mei yngong fan 12 july 2015, waard brûkt om in ynternetbasearre literatueresykje út te fieren. De folgjende termen waarden opnommen: hyperseksueel (170), hyperseksualiteit (479), seksuele ferslaving (1,652), seksferslaafde (1,842), seksuele ympulsiviteit (989), twangmjittige seksuele (946), twangmjittige seks (1,512), seksuele twang (782) ), parafilia-relateare steuring (234), en oermjittige seksuele (857). Om't dit artikel in koarte resinsje is ynstee fan in meta-analyze, is it basearre op in represintative seleksje fan dizze stúdzjes dy't relevant binne foar de ûnderwerpen dy't behannele binne. It net opnimmen fan in bepaalde stúdzje negeart it belang foar it fjild net. Begryplik binne d'r dejingen dy't it net iens binne mei it konsept dat seksferslaving yndie in echte oandwaning is en sels bewiis kin hawwe om oan te toanen dat se op fêste grûn binne. Wy stelle lykwols dat d'r genôch bewiis is om foar te stellen dat seksuele ferslaving bestiet en stúdzjes dy't neuroimaging, neurogenetika en sels epigenetika oandogge, stypje it begryp dat twangmjittige seksferslaving as hyperseksualiteit kinne wurde beskôge as in ferslavende oandwaning. Wylst wy bewust binne fan dizze ferskilens, leverje wy respektivelik ús stânpunt yn om fierdere wittenskiplike kontrôle te generearjen en net wittenskiplike retoryk te ferdriuwen oer dit wichtige ûnderwerp. Om ekstra ljocht te werpen op it ûnderwerp hawwe wy de literatuer socht nei wat konsensus. Op 17 augustus 2015 sochten wy PUBMED Central mei de folgjende term - "Is seks in echte ferslaving?" en 46 artikels ophelle.
Definysje fan seksuele ferslavingen
Seksuele ferslaving wurdt definieare as elk twangmjittich seksueel gedrach dat it normale libben bemuoit en swiere stress feroarsaket op 'e famylje, freonen, dierberen, en ien syn wurkomjouwing. Seksuele ferslaving is seksuele ôfhinklikheid, hyperseksualiteit, en seksuele twangmjittigens neamd. Mei elke namme is it in twangmjittich gedrach dat it libben fan 'e ferslaafde folslein beheart. Seksuele ferslaafden meitsje seks in prioriteit wichtiger dan famylje, freonen en wurk. Seks wurdt it organisearjende prinsipe fan it libben fan ferslaafden. Se binne ree om te offerjen wat se it meast koesterje om har ûngesund gedrach te behâlden en troch te setten [12]. Hyperseksueel winsk is bepaald as winsk basearre op in libben-beoardieling fan 'e frekwinsje fan seksueel gedrach en tiid dy't yn' e assosjearre seksueel fantasyën brocht wurdt. Yn manlju is in evaluaasje fan hyperseksuele winsken definiearre troch Kafka en Hannen as de heechste ûnderstreke perioade (op syn minst seis moanne) fan persoanlik opnommen seksueel gedrach (totale seksuele rinte / wike nei leeftiid 15). In feite, in longitudinale skiednis fan hyperseksuele winsk, operatyf as definiearre definiearre, waard identifisearre yn 72-80% fan manlju dy't behanneling behannele wurde foar paraphilia's en paraphilia-ferbûne feroaringen [13].
Hyperseksualiteit en taalfersifers
It is goed fêstlein dat yn 'e minsklike seksuele mienskip en literatuer seksueel begearte kapitulearre wurdt as it oanwêzigens fan seksueel fantasy's, aktiviteiten of opropt, en motivaasje troch de minske om yn seksuele gedrach te beheljen. Der binne sawol ynterne en eksterne relevante toanen [14]. Evolúsjonêre teoryfoarming hawwe bewiisd dat manlju en froulju ferskillende agindas hawwe as it giet om seksyske aktiviteit [15]. In soad ûndersiken ferskine ferskate ferskillen tusken manlju en froulju. Minsken hawwe seksueel fantasy ferhege [16], ferhege frekwinsje fan masturbaasje [17], fergrutte oanpast foar ekstreem generearre fisuele seksuele opheffing [18] permissive hâldingen foar it slimste seks [19], fasiliteit fan oprop [20], en yntinsive motivaasje [21]. Yn tsjinstelling ta sjogge wyfkes in oare seksueel lânskip mei seksueel motivaasje, seksueel opwaaks, en seksueel gedrach binne foarmje troch evolúsjonêre faktoaren [22] en gruttere biologyske, emosjonele en tydlike ynvestearring yn reproduksje en berne-oplieding [23]. Femalen binne minder kwetsber foar hyperseksualiteit [24] en oanpast oan affichearjende relaasjes en langere termyn fan partner oan te fangen [25]. Wylst seksueel-ferslacht wurde beskôge om oant 3% nei 6% fan 'e befolking te ferstean, is it dúdlike ynsjoch fan' e neurobiologyske foarhannen beheind [26] lykas klinyske beoardielen [27]. Wy stimulearje fierder lêze op seksuele twang, oanhing en seksuele oriïntaasje [28], en gersdifers yn responsen op seksuele stimulearrings [29-30].
It is dúdlik dat Kafka en Hennen binne [13], fûn dat de gemiddelde leeftyd fan begjin fan persistente hyperseksuele gedrach wie 18.7 ± 7.2 jier yn seksueel aktive manlju en it leeftydskommunikaasje fan hyperseksuele gedrach wie âld jier 7-46. De gemiddelde tiid fan dizze heechste konsekwint bewarre frekwinsje fan seksueel appetitive gedrach wie 12.3 ± 10.1 jier. It betsjuttende leeftyd fan dizze aktive seksuele manlju is lykwols hyperseksual dat sykjen behannele is 37 ± 9 jier. Hanson, et al. ek evaluearre hyperseksualiteit yn ferrifelingen en fûn dat leechferbrûkers legere residivisme tariven hiene as heech risikoofenders [31].
Hyperseksualiteit en ko-moarige substans misbrûk
Der is in hege ko-morbiditeit tusken hyperseksuele stompe en oare socht, lykas stoflik gebrûk fan disorder [32-33]. Gewoanlik stelde Garcia en Thibaut dat de fenomenology fan in oerfloedige non-parafilike seksueel ûngelok as klassike gedrach klassiearre wurde moat, anterste as in obsessive-compulsive, of in ymposysjebehearsking [34]. Se sjogge goed út dat de kritearia krekt tawiis is oan dy fan sêftmoedigensstelders, lykas troch oaren trochsteld [35], Dizze ûndersikers hawwe de ympuls levere foar fuortset ûndersyk op dit gebiet en mooglike takomstige ynklúzje fan hyperseksuele steuring yn 'e DSM-6. Drugs fan misbrûk, rock 'n' roll en seks komme tagelyk foar, en hiele festivals binne boud om dizze kombinaasjes fan Woodstock oant hjoed de dei.
De literatuer lit sjen dat brûkers fan methamphetamine rapporteare dat dit stimulearjende medisyks seksueel begearte ferheget, benammen risiko-gedrach. Mar amphetamine is lykwols oanwiisd om de seksuele aktiviteit fan froulike ratten te ferleegjen. Mei dit yn 'e tinken, Holder, et al. evaluearre de rol fan methamphetamin yn froulike ratels [36]. Se fûnen dat yn it tsjinoerstelde methamphetamine it froulike seksueel gedrach fasilitearre en dizze effekt is troch it fersterkjen fan dopaminergyske transmission en sels mooglik neurotransmission trochwege de kombinaasje fan eierhoarnen en methamphetamine. Spesifyk fûnen se in ferbettering fan seksueel motivaasje oanbean mei aktivaasje fan neuroanale aktiviteit yn 'e mediale amigodala en ventromediale nucleus fan' e hypothalamus.
Boppedat hawwe wittenskippers fan Nederlân de ko-morbiditeit fan substans misbrûk yn self-identifisearre swingers studearre [37]. Yn dit stúdzje, Spauwen, et al. It konkludearret dat 79% fan swingers rekreaasje-rekreaasje brûkt (lykas alkohol en gebrûk fan ektectile dysfunksjegroepen); 46% fan harren melde meardere drugsgebou. In feitlik rekreaasje-drug use (eksklusyf alkohol en erektile dysfunksjegedokter) is mei-inoar ferbûn mei heule risiko-seksuele gedrach yn manlju en froulju. Ek drugsgebrûk wie selsstannich ferbûn mei seksueel trochferwiske ynfeksjes (STI) yn froulike swingers, benammen dy dy't meidogge yn groepseeks.
Castelo-Branco, et al. rapporteare dat jonge froulike froulju fermearje dat seksualiteit in wichtich part fan har libben is, mar gjin primêre soarch (77.6%) [38]. Hja rapporteare ek dat alkohol de barriensken foarkomt om seks (62.3%) te meitsjen. Wichtich binne se ek fûn dat alkoholmisbrûk in foarsizzende fariant wie foar it fersterkjen fan risiko 's gedrach ûnôfhinklik fan' e leeftiid fan 'e froulik.
It is dúdlik dat Jia, et al. rapporteare gefaarlike seksuele gedrach tusken psychostimulant- en heroine-abusers, wêrûnder meardere seksuele kontakten, akkuraaze seks partners, homoseksuele partners, en nea of foarkommen praktyske seks [39].
Us haadhanneler is dat medisinen, lykas methamphetamine, kokaïne, heldewyn en alkohol, kinne seksueel langstme stimulearje yn net-bedriuwen. Yn fersen is it hiel oars; De selde medisinen kinne feroarsaakje oan in chronike basis. Dochs binne post-ferslaving yn 'e rekreaasjefaze yn in protte gefallen in aphrodyske-like gedrach behannele.
Hyperseksualiteit en weromlûken
In PubMed-sykaksje (7-19-15) mei de term "hyperseksualiteit en weromlûksymptomen" resultearre yn mar fiif artikels, wêrfan gjinien beskreau "weromlûksymptomatology." In alternatyf sykjen mei de termen "hege symptomen foar weromlûking fan seksuele aktiviteit" resultearre lykwols yn 25 artikels neamd.
Addiken yn rekreatyf rapport fergruttet yn iten en in appetitriid foar bepaalde iten en misbrûk fan sigaretten yn 'e lange termyn. Gewichtsferwidering is ek beweard en dokumintearre yn koartlyn en langstige ûnthâldde dieren en minsken [39]. Bruijnzeel makke de nijsgjirrige observaasje dat acute opiate weromlûking kin yn spontane orgasmen [40]. Belangryk, yn it artikel brocht Bruijnzeel út dat de sykte fan sykte fan medisinen en eventueel chronike yntinsive seksuele aktiviteit kin wêze troch de unopposed funksje fan kappa opioid receptor-sinjalearing dy't de release fan dopamine ferhellet, wylst it norepinefine yn 'e harsens beliedsirkels groeit.
De papieren yn 'e literatuer leinen fan in tekoart oan strangens oangeande akuere en langstige ôfwiking en ynstinsintaasje befêstige troch urineproblemen. Klinyske behannelingprogramma's wurde hyltyd ferpleatst fan behanneling fan beide geslachtingen om de beide seksjes te isolearjen, foar it jaan fan geskikte spesjale behannelingprogramma's. Se biede ek ûnderwiis om de frate en lange tiid te ferheegjen yn seksueel belang en aktiviteiten, en de relaasje fan ferwikseling foar iten en oerwinning.
Hyperseksualiteit-oanwakke tekoartsymptomen binne troch in oantal ûndersikers rapportearre mei ferskate degradaasje fan hurdens en moarnsidige substans misbrûk [41-45]. As gefolch fan dizze syktocht hawwe wy gjin inkeld papier fûn om aktuele weromdrawingsymptomen te beskermjen dy't ferbân hâlde mei ynstinking fan aktive seksuele bewenners. De measten fan 'e papieren befette de effekten fan ôfwaging fan drugs fan misbrûk, lykas opioïden, nicotine, amphetamine en kokaïne, dy't seksuele aktiviteit feroarsaakje kinne.
Hyperseksualiteit en neurogenetik
In PubMed-sykopdracht (7-19-15) ûntdekte mar seis notearre papieren mei de term "genen en hyperseksualiteit", meast rjochte op artikels relatearre oan Kleine-Levin syndroom (KLS), in heul seldsume sykte wêrby hyperseksualiteit oant 27 jier koe duorje. Yn ien stúdzje waard fûn dat in immuunresponsive HLA-DQBl, DQBl * 0602 waard ûntdutsen yn wichtige hoemannichten by pasjinten mei KLS en koe it risiko fan KLS ferheegje [46-47].
Doe't wy lykwols de termen "seksuele aktiviteit en genen" brûkten, waarden 2,826 artikels neamd, en wy leverje in koarte synopsis fan in pear wichtige neurogenetyske aspekten. It is ús hypoteze dat sawol hedoanyske as anhedonyske gedrach resultaten binne foar in part fan 'e risiko-allelen fan in yndividu foar dizze gedrach en dat behanneling bestiet út passend op dizze identifisearre polymorfismen. Boppedat is behannelingreaksje ek ôfhinklik fan dizze risiko-allelen en leveret in wichtige reden foar pharmakogenetyske testen en pharmakogenomyske / nutrigenomyske oplossingen.
Nei it kontroversjele begjinfûns troch Blum, et al. yn 1990 fan 'e earste bewiis foar in feriening tusken it DRD2 Al allele en it swiere alkoholisme, binne der 3,938 artikels yn PubMed (7-19-15) [48]. De stúdzjes befetsje it psychiatryske gene polymorphisme, de DRD2 Al allele, en dêrmei ferbûne gedrach en fysiology. Der is lykwols in lyts gebrûk fan gegevens dy't seksueel aktiviteit oanbelangje oan dizze en oare relategenen, nettsjinsteande de oerweldige bewiis foar mesolimbyske aktivearring, benammen yn dopaminergyske paadwizen en neuronale lokaasjes dy't relatearre oan seksuele stimulearrings en aktiviteiten. It is dúdlik dat Blum en Noble de DRD2-gene justifisearre hawwe as in generalisearre belesjen fan fertsjintwurdigers foar alle gedrach fan 'e belestingsmindsyndroam (RDS). It feit dat gebrûk fan 'e Bayesian-teorie-analyse-trailers fan' e Taq Al allele hawwe, oer har libben, in 74% kâns, dat se rendezvous hawwe mei ien of mear bedrachmiddels (RDS) gedrach [49].
De earste ferieniging fan elk gene polymorphisme en seksuële aktiviteit foel net oant 1999 doe't Miller, et al. bepaalde dopaminergyske genen [50]. De basisfeardigens is dat it dopaminergyske systeem yn it hynste in wichtige rol spilet yn 'e regeling fan seksueel gedrach. De relaasje tusken genes foar de Dl, D2, en D4 dopamynfektoaren en leeftiid op it earste seksuele ferbân (AFSI) waard ûndersocht yn in foarbyld fan 414 non-Hispanisch, Europeesk-Amerikaanske manlju en froulju. In wichtige feriening waard observearre tusken in DRD2 allele en AFSI en in noch sterker feriening doe't it DRD2 allele mei in DRDl allele ynteraktyf waard. In yntegrearre regressionmodel waard foarsjoen fan AFSI te brûken troch seks en in groep fan njoggen psychososjale fariabelen as foarbylders. It tafoegjen fan 'e DRD2 en de DRD2-by-DRD1-prediktors nei dit model fergrutsje de fergrutte ferzje fan 23% en 55% respektivelik. It feit dat dizze befiningen in sterker ferieniging tusken mantsjes sjogge as ûnder de froulju is yn oerienkomst mei it resinte wurk fan oaren dy't hegere seksuele stimuaasjemiddels sjen litte yn mantsjes dan yn 't froulju [51], Dat miskien "manlju binne fan Mars en froulju fan Venus" en dit kin sels wier wêze foar kokaïne misbrûk [52].
Presiids hawwe beide preklinike en klinyske stúdzjes seksueel dimorfyske patroanen yn 'e gedrach fan' e gedrach fan 'e kokaïne yn alle fazen fan' e koalysje-ferslavingproses (yndeksje, ûnderhâld, en weromkearing) oanjûn. Sa is in dúdlike byld ûntstien, wat bepaald dat der in biologyske basis is fan seksueel spesifike ferskillen yn 'e kokaïne-ferslaving. Dizze ferskillen foarmje út 'e ûnderskate regeljouwing fan' e CNS troch manlike en froulike gonadale hormonen en kin presiisd wurde troch de oanwêzigens fan DRD2-gene polymorphisme [53]. Boppedat is it bekend dat genetyske ferienings tusken COMT en ferskate psychiatryske fenotypen faak ferskillen tusken manlju en froulju sjen litte. Dizze binne ûnder oaren it funksjonele Val (158) Met polymorphisme yn COMT dy't ferbûn is mei obsessive-compulsive disorder yn manlju en mei angst fenotypen yn froulju. Dêrnjonken hat de Val (158) Met polymorphisme yn COMT in grutter ynfloed op kognitive funksje yn jonges as famkes [54].
Miller, et al. wie gjin feriening fan 'e polymorphisme dy't yn ferbân brocht mei it DRD4-gen en leeftiid fan earste seksuele oarloch [50], Oaren fûnen lykwols in wichtige feriening yn bepaalde etnyske groepen. Spesifyk jouwt har analyze fan 'e polymorfismen yn DRD4 oan dat dyjingen mei elk - 3R genotype in risiko hawwe ûnderfûn fan earste geslachtsferkear heger dan dy mei oare (as hokker - 4R) genotype yn' e all-etnyske groepen (n = 2,552). Opfallend is dat it risiko fan earste seks net ferskilt tusken de twa genotypen yn 'e Afro-Amerikaanske stekproef, wat de fraach opropt oer kulturele opfieding [55].
De seksuele ûnderfining, lykas werhelle medisyngebrûk, produktt lange termyn feroarje, ynklusyf sensibilisaasje yn 'e nucleus accumbens (NAc) en dorsale striatum. Bradley, et al. mei help fan microarray-analyse om ûndersikers te studearjen foar it earste kear dat de seksueel ûnderfining yn sawol manlike of froulike bisten differinsjaal of opheind regulearret mRNA ekspresje fan in searje genen yn 'e NAc [56]. Se fûnen dat yn ferliking mei seksuele naïve dieren seksueel ûnderfûnen hamsters krigen in stimulus manlik op Week 7 eksposearre in groei fan in grut tal genen. Oarsom hawwe seksueel ûnderfining fan froulike hamsters gjin stimulearre mens op Week 7 eksposearre in minderheid yn 'e ekspresje fan in protte genen. Neffens de auteurs kin dizze earste ea-profeaasje yn froulike hamsters in ynsjoch yn 'e meganismen leverje wêrby't beide gedrach (seks) en drugs fan misbrûk langstme feroaringen yn' e mesolimbyske en nigrostriatale dopamine-paden ynfiere.
Bipolarelektroden, dy't bilateraal yn 'e laterale hypothalamus en substansjele nigraventraal tegmentale gebiet ymplant binne, waarden stereotaksysk brûkt om kwantike selsstimulearjende belibjende ûnderfiningen oan te meitsjen dy't ferlykber binne mei seksueel gedrach. Dizze soarte stimulearjen is fûn om in signifikant ferheging fan it oantal synapses yn 'e CA3-regio fan' e hippocampus en de molekulêre lagen fan 'e motor cortex yn rat te stimulearjen. Yn essinsje is de chronike hynstestimulaasje langere termyn (LTP), dy't bekend is om nije synaptyske ferbiningen te ferheegjen [57]. In ienige eksposysje foar kokaïne yn naïvige bisten is genôch om langstige feroaringen yn 't ventral tegmentale gebiet (VTA) glutamatergyske synapses te trigearjen dy't likernôch aktiviteiten ôfhinklik LTP yn oare harsensregio's binne. Dizze kokaïne-induzearre LTP ferskynt omtinken te meitsjen fia dopamine D5-receptor-aktivearring fan N-methyl-D-aspartate (NMDA) -receptors en om de proteinsynsystik te freegjen [58], it opnimmen fan ús foardering opnij opnij dat drugs en seks meast mienskiplike neurochemyske substraten hawwe.
Empirysk ûndersyk hat in positive relaasje west tusken it tal seksy partners en belutsenheid yn antisosjale gedrach [59]. De measte problemen om dizze ferieniging te ferklearjen hawwe in evolúsjonêre perspektyf nommen. Fanút it evolúsjonearjende perspektyf binne deselde trekken, bygelyks ympulsivo, skitterjendeens en agressiviteit, dy't relatearre binne oan in grut oantal seks partners, binne ek ferbûn mei kriminele belutsenens. Lykwols is der ek reden om te leauwen dat de gearfetting tusken seks partners en feroardering kin diels ferklearre wurde troch in mienskiplike genetyske wei, dêr't genes dy't relatearre binne mei seks partnerpartners binne ek relatearre oan antisosjale gedrach. Mei it brûken fan de hjirboppe beskreaune begraffenis, Beaver et al. In sterke positive feriening fûn tusken seks partners en antisosjale gedrach en polymorphisme fan 'e dopamine transportergen (DAT1) ferklearret ferdieling yn beide getallen fan seksuele partners en kriminaliteit foar manlju [59]. De polymorfyske ynfloed fan 'e DAT-l-gen en it tal seksypartners kinne in feriening wêze dat fûn binne tusken beskate polymorphisme en moarns ierde pensile ejaculation. Carriers fan 'e genotyp 1OR / 1OR hiene punten dy't in legere drompel oanjûn hawwe om op elk fan' e yndikaasjes te ferjitten yn fergeliking mei de kombinearre 9R9R / 9R10R (9R hegere aktiviteit legere dopamin beskikber) [60]. Polymorphisme fan it DATl-gen, benammen de genotyp 10R / 10R, binne fûn yn juvenile delinquents dy't oan 'e Brown School (San Marcos, Texas) oanwêzich binne foar psykologyske agressyf gedrach, ynklusyf anty-sosjale gedrach [61]. In positive korrelaasje fan beide DRD2 en de DATL-polymorphisme wiene bepaald mei pseologyske geweld yn adolesinten yn in bliuwende klinyske proef. Boppedat wurdt, lykwols, yn begjinsel begrepen as it resultaat fan 'e peer-ymotaasje fan' e bern-ynset of libbens-ferrassende jeugd, is it beweechzjen fan twillingûndersiken dat adolesinte-ynset of adolescent-beheinde antisosjale gedrach ek genetysk beynfloede wurde kin. Burt en Mikolajewski befestigje net allinich dizze fynsten mei de DATl-gene, mar ferwidere dizze befinings om de syn452Tyr fariant fan 'e gene te ferwiderjen, dy't de 5-HT2A-receptor ek kodearret [62], Mear resint, Jozkow et al. rapporteare in feriening tusken de seksuele dimensje fan symptomen fan fergrizing fan manlju (AMS) en genetyske farianten fan 5-HTRlB G861C [63]. Boppedat is Sales, et al. Fûn troch multivariable logistyske regression analzje, in ynteraksje tusken misbrûk en 5-HTTLPR-groep wêrby't in wichtige feriening is mei net-wizigingstatus, tegearre mei partnerkommunikaasjefrekwinsjes by folgjen [64] In misbrûk fan misbrûk wie wichtich ferbûn mei gruttere kandidaten fan net-wiziging yn kondom brûke post-yntervinsje foar allinich dejingen mei de s allele.
It is bekend dat polymorphisme yn net-kodearjende regio's fan 'e vasopressin la receptor-gen (Avpr la) ferbûn binne mei sosjaal-emosjoneel eigenskippen yn minske, skimpâns, en foles, en kinne troch in side-spesifike fariant yn' e gene ekspresje. Neffens Barrett, et al., Biedt de sosjaal monogamous prairiefoll in unike kâns om de neurobiology fan monogamy te studearjen [65]. Yn feite is it signalisearjen fan vasopressin la receptor (VlaR) needsaaklik foar de formaasje fan 'e paadbining yn mantsjes. Ynteressant binne de sosjale prairievelen in gruttere VlaR-ferbining yn 'e belibjende ferwurking fan ventral pallidum as asosiale folen fan deselde genus. Barrett, et al. fûn dat ferdieling fan pallidale VlaR-tichtens in wichtige beoardieling yn 'e foarkar foar in mienskiplike frouljuspartner en in ferleegjen fan anxiety-like gedrach yn folwoeksenen [65], Oare wurk fan Garcia, et al. die bliken dat persoanen mei teminsten ien 7-repeat allele (7R +) fan 'e DRD4 rapportearje in grutter kategoarysk taryf fan promiskuos seksueel gedrach, ynklusyf it hawwen fan in "one-night stand", en rapportearje in mear as 50% ferheging yn gefallen fan seksuele ûntrou [66].
Wichtich is dat Daw en Guo rapporteare dat persoanen dy't de genotypen drage, DRD2 * Al / A2, DRD2 * A2 / A2, DATL * 9R / 10R, en MAOA * 2R / binne ferbûn mei hegere kâns fan ûnbeskikte geslachtsferkear as oare genotypes op dizze loci [67], De DRD2-ferienings binne fan tapassing op sawol manlju as froulju, wylst de oare keppelings allinich jilde foar froulju. Uteinlik Emanuele, et al. rapporteare in wichtige assosjaasje tusken de DRD2 TaqI A genotypen en "Eros · (in leafdefolle styl karakterisearre troch in oanstriid om yntinsive emosjonele ûnderfiningen te ûntwikkeljen basearre op 'e fysike oanlûkingskrêft foar de partner), lykas tusken it C516T 5HT2A polymorfisme en" mania "( in besitlike en ôfhinklike romantyske oanhing, karakterisearre troch emoasjes dy't sels ferslaan) [68].
Epigenetik en seksuele aktiviteit
In oersicht fan 'e literatuer lit sjen dat in oantal resinte artikels it belang fan epigeetyske effekten op seksuele aktiviteit sjen litte. Bygelyks hat Matsuda de epigeetyske feroaringen fan 'e estrogenreptor a (ERalpha) en ynfloed op sosjaoseksuele gedrach beoardiele [69], Eins feroaret feroaring fan ER-alfa-genaktiviteit bemiddele troch epigenetyske meganismen, lykas histoanmodifikaasjes en DNA-methylaasje, it seksuele gedrach fan ien. Yn termen fan homoseksualiteit, Rice, et al. ûntwikkele in model dat de kanalisaasje (bekearing) homoseksualiteit seksuele ûntwikkeling kin ferklearje [70]. Hja ferklearje dat dit model basearre is op epigenetyske marken dy't yn 'e antwurd op' e XX-tsjin-XY-karyotype yn embryonale stammeltsjes lizze. Dęrnei stimulearje dizze marken sensibiliteit oan testosteron yn XY-fetussen en legeren it yn XX fetus, sadat de seksuele ûntwikkeling kanalisearret. It is postúmearre dat in subset fan dizze konvertearjende epigenetyske marken kwantitatyf oer trans-genetysk oer drage kin en kin ta mosaisisme foar seksylike ûntwikkeling yn tsjinoer-seksjebewiis bringe - de homoseksuele fenotype.
Yn 'e sosjale monogamous prairie vole (Microtus ochrogaster), bringt mating enduerende paadbondels dy't ynspireare binne troch partnerfoarmingfoarming en regeleare troch in ferskaat oan neurotransmitters, wêrûnder oxytocine, vasopressine, en dopamine. Wurk fan Gundersen [71], en Wang, et al. [72] Hy suggerearret dat histone-deacetylase de partnerfoarming fasilitearje kin yn froulike prairievels dy't relevânsje hawwe foar minsken. Oanwêzich, Wang, et al. fûnen dat histonedeacetylase-inhibitoren-natriumbutyrate en trichostatin A (TSA) fergrutte partnerfoarmingfoarming yn froulike prairievels [72]. Dizze partnerfoarming waard ferbûn mei in oprregulaasje fan oxytocine-receptor (OTR, oxtr) en vasopressin V la receptor (VlaR, avprla) yn 'e NAc, troch in tanimming fan histone acetylaasje oan har ûnderskate promotoren.
Der is belangstelling foar groei fan bewiis dat oantsjutte dat froulju aktyf yngripe yn polyearen, om genetyske ynkompatibiliteit te foarkommen, of om paternity yn foardiel te meitsjen foar genetysk superioren. Der is de mooglikheid dat seleksje fan in heule manljusfetsje kin efterynteigene effekten binne. Neffens Zeh en Zeh, yn tsjinstelling ta DNA-sequinsbasearre fariaasje, kin epigeetyske farizje sterk beynfloede wurde troch miljeu-en stochastyske effekten dy't ûnder it libben fan in yndividu binne [73]. Se suggerearje dat epigeetyske farianting wichtich wêze kin foar de post-copulatory seksuële seleksje en kin rekkenskip wurde foar befiningen dy't keppeljen sperma konkurse kapasiteitsje oan fekânsje fitness.
Genetyske en meme evolúsje: Human procreation
Eysenck stelde in positive korrelaasje foar tusken ekstraversje en yntinsiver seksueel gedrach en tusken neurotykisme en problemen yn seksueel gedrach (anty-sosjaal gedrach). In eardere stúdzje mei troude minsken liet gjin fan dizze korrelaasjes sjen. It waard hypoteze dat dizze ferbining allinich bestiet foar net-troude persoanen dy't net dwaande binne mei langduorjende relaasjes, om't de kwaliteit fan 'e relaasje de seksuele ynteraksje bepaalt. Binnen in stekproef fan jonge net-troude manlju wie d'r in positive korrelaasje tusken ekstraversje en items wêryn't de persoan earder seksuele aktiviteit beskreau mei mear yndividuen en yn hegere frekwinsje. Gjin korrelaasje waard fûn mei neurotyk. D'r wiene ek lichte korrelaasjes mei oare skalen foar persoanlikheid en sosjale hâlding. Fanwegen de korrelaasje mei in aktearjende persoanlikheidsskaal waarden de befiningen ynterpretearre út in sosjaal-psychologysk perspektyf. Yn 'e hjoeddeistige maatskippij wurdt fan' e jonge man ferwachte dat se it inisjatyf nimme yn in seksuele ynteraksje dy't in ekstraverteare jonge man better kin realisearje dan ien dy't yntrovert is [74], Dit perspektyf is yn direkte oerienkomst mei Richard Brodie's idee oer egoïstyske genen fan 'e geast [75], Fanút it eachpunt fan it DNA soene antropologen it fansels iens wêze "wy binne hjir noch mar om ien reden; útgean en fermannichfâldigje. ” Wylst evolúsjonêre foarútgong stadich is, ien stap om 'e 20 jier of sa, yn ferliking mei "meme-evolúsje, muteart in idee yn' e tiid dat it nimt om in sin te lêzen." Us harsens hawwe neat te meitsjen mei genetyske evolúsje, útsein om't it relatearre is oan tûke minsken dy't minder poppen hawwe. Eins, as d'r genen binne dy't minsken de oanstriid jouwe memes oan te nimmen dy't har oantal neiteam beheine, sille se yn in pear generaasjes útstjerre yn it foardiel fan genen dy't minsken in oanstriid jouwe bern te krijen. Hoewol wat kontroversjeel, spitigernôch suggerearje in oantal ûndersiken dat Homo sapiens oer de lêste 42,000 jier har IQ's hawwe ferlege fanwegen selektyf parjen [76].
Utseinoarmich stelt it út dat der ekstraferzje ferbûn is mei ferhege seksyske aktiviteit benammen yn manlju, de kwantitative genetysk priizget de heriteberens fan 'e útwreide persoanlikheid om 40-60% te wêzen. Smillie en assoziaten studearre en fûnen dat ien kopy fan 'e DRD2-gen Al allele assosjearre is mei signifikant sterker ekstraversion [77], Dizze feriening ropt in nijsgjirrige fraach op yn termen fan minsklike fuortplanting. Comings suggerearde dat fanwegen har markante effekt op reproduktyf gedrach, learsteurnissen en oare ympulsive, twangmjittige, agressive en ferslaavjende steuringen dy dragers fan 'e DRD2 Al it potensjeel hawwe om progressive en permaninte feroaringen te feroarsaakjen yn' e frekwinsje fan 'e DRD2Al allele "dy't liedt ta de genetyske meltdown fan 'e soarte " [78].
Yn syn boek jout Comings bewize dat minsken mei ferdivedigjende gedrachsjes earder bern hawwe, en dit beynfloedet de seleksje fan genôchgenoaten lykas de DRD2 Al allele [79], Hy suggereart dat persoanen dy't dit fersteurende risiko-allel drage bern krije sille litte we sizze op 20-jierrige leeftyd en yndividuen sûnder dit allel sille bern hawwe op 25 jier. As resultaat sil it mutante gen rapper reprodusearje, nammentlik elke 20 jier, wylst de normale foarm fan it gen elke 25 jier sil reprodusearje. De ferhâlding fan 25/20 is 1.25. Sadwaande sil de snelheid wêrmei't in gen dat in 1.25-fold selektyf foardiel hat, tanimme yn frekwinsje fan generaasje op generaasje. In ferskil fan fiif jier yn 'e leeftyd fan memmen of heiten as se har earste bern hawwe is genôch om te resultearjen yn in wichtige en relatyf rappe seleksje foar genen dy't wurde droegen troch groep dy't op in eardere leeftyd in bernefeardigens begjint. Ferheegingen yn guon RDS-gedrach binne dokuminteare fan 1955 oant hjoed de dei. Dizze ferhegingen omfetsje adolesinsgedrachsyndroam (drugs, seks, jongereinswangerskippen, en misdiedich gedrach, smoken), gedragsstoornis, misdied, drugsmisbrûk, alkoholisme, ûnbeskermd seksueel gedrach, net-winske memmen, wolwêzen, skoalle útset, en skoalútfal, lykas in tagelyk fermindering fan it IQ [80]. Dizze resultaten binne basearre op it Berkeley Studie dat gebrûk makket fan longitudinale gegevens fan 'e Child Health and Developmental Studies en de National Longitudinal Surveys of Youth of NLYS [81]. Utlis fan dizze ynformaasje befettet Comings dat fan 1955 nei 2015 in dûbeling fan de frekwinsje fan bygelyks de DRD2 Al allele is, wêrtroch de prevalens fan RDS-gedrach ferheget, wêrûnder prekeardich geslachtsferkear [50]. Wy stimulearje in folchoarder fan dizze nijsgjirrige predikaasje.
Nettsjinsteande in geweldig misdiedigens dogge wy foar it hyperseksualiteitskrêft as subtype fan RDS te dielen fan skaaimerken mei substans en non-substansjoneel ferrifeljend gedrach mei har klinyske ekspresje dy't diels beynfloede wurdt troch sawol genetyske en epigenetik. Hoewol dit net tefreden is, hawwe wy ek koarte termyn FDA-goedkard medisitaasje-behannele behannelingen (MAT) foarsafier't it foarkommen fan it blokkearjen fan dopaminefunksje folge troch sanlike activaasje fan dopaminergyske weeën dy't ta langstme dopamine homeostasis leverje. De lêste kin begelaat wurde troch guon modaliteiten dy't helpe kinne yn rekreaasje.
Hoewol potinsjele foaroardielen, se binne dopamine agonist therapy-nutraceuticals (KB220), it programma 12 Step en tradysje, holistyske behanneling, cognitive behavioral therapy (CBT), trauma-relieftriary (TRT), en dopamine-stimulearjende aktiviteiten en iten 1) [82].
De kontroversje
Wylst wy festigje dat hyperseksualiteitskrêft yn 'e takomstige edysjes fan DSM opnaam wurde, binne wy wat ferwûnen dat sa lyts is bekend oer dizze ûngelok yn termen fan neurogenetics en epigenetik en sels ôfwaging symptomatology en folsleine fenomenology [83]. It primêre take-home-berjocht is dat wy de wittenskiplike mienskip stimulearje om eksperiminten út te fieren, benammen yn it ryk fan neuroimaging en neurogenetics, ynklusyf epigenetika spesifyk foar genes, lykas oxytocin-vasopressin-orexin-dopamine as ek oare liengenen. Mooglik kin dizze betingens foardielje fan behanneling dy't tariedings gene polymorphisme oanbelanget om te helpen by it befoarderjen fan dopamine-homoostasis [84-89]. In oantal resinsjes fan Joranby, et al. en Edge en Gold stypje mienskiplike behannele mooglikheden yn ferbân mei dielde neurochemyske meganismen yn 'e harsensbeleidingsrjochting as eardere yn it RDS-konsept [90-91].
Histoarysk waard "seksferslaving" opnommen yn DSM - III, lykwols waard it fuortsmiten fan DSM-1V om't de konsensus fan 'e auteurs fan DSM-1V leaude dat d'r net genôch bewiis wie om syn konklúzje te fertsjinjen. Dit beslút wie fol mei enoarme emoasje troch lieders yn it fjild. Nei dit barren besleaten in oantal wittenskippers, lykas Kafka, Reid, Prause, en oaren om "Hyperseksualiteit" net te ramt as in seksferslaving, mar in selsstannige mentale oandwaning en net as in ferslaving per se. Wylst har eardere wurk yn 2010 suggereare as hjiryn ferwiisd dat "hyperseksualiteit" fergelykber wie mei seksferslaving en mooglik oare ferslavingen, ynklusyf substansmisbrûk, stipet har nijere wurk fan dizze striid. Resint wurk yn dit gebiet ferriedt kontroversje. D'r binne in oantal elektrophysiologysk basearre stúdzjes, troch de groep fan Prause dy't wat bewiis leverje dat seksueel langstme, net hyperseksualiteit, selsregulaasje fan seksuele opwekking foarsjocht. [92]. Dizze ûndersikers jouwe yn oare wurken dy't subjects rapporteare problemen regelje fan harren besjen fan fisuele seksuele stimulaasjes (VSS) dy't ek hegere seksueel winsken litte sjen litte leechtere posityf potinsjes (LPP) yn antwurd op VSS. De auteurs suggerearje dat dit patroan ferskynt ferskilt fan substansykmodellen [93]. Dochs wylst net ûnderwerpen mei hyperseksuele ûngelokken wurkje, wurk fan Voon's groep hat sjen litten dat yn komplekse-seksueel-gedrach ûnderwerpen, eksposearje oan seksueel eksportearre fideos, in gruttere aktiviteit yn it neuronale netwurk, likegoed as dy observearre yn drug-cue-reactivity studies [94]. Greate begearte of wolle jo ynteressearje net fierder ferbûn mei aktiviteit yn dit neuronale netwurk. Dit wurk ferdylt mei teoryen fan stimulearring [95].
Wy, de auteurs fan it hjoeddeistige artikel, jouwe ta dat wy net yn 'e kunde west hawwe mei alle wichtige ynteraksjes dy't binne foarkommen tusken de foarstanners fan "Hyperseksualiteitsteuring" en har earlike bedoeling om dizze steuring yn' e hjoeddeiske DSM-5 op te nimmen. Hoewol it de saneamde "acid-test" mislearre, is d'r alle reden om te leauwen dat it sil wurde opnommen yn takomstige edysjes fan 'e DSM. It is opmerklik dat Steven Hyman, de hjoeddeiske NIH-direkteur, korrekt bewearde dat “de DSM in minne spegel is fan klinyske en biologyske realiteiten; in fûneminteel nije oanpak fan diagnostyske klassifikaasje is nedich as ûndersikers nije manieren ûntdekke om mentale sykte te bestudearjen en te begripen ” [96]. Boppedat is Casey, et al. It presmente dat de DSM beskôgje as ferskate ûngelokken as ûnderskate entiteiten, "grinzen tusken diskriminaasjes binne faak net sa strang as de DSM suggerearret" [97].
Yn 2014, Karila, et al. joech suggereard dat seksuele ferslaving, ek wol hyperseksuele striid neamd, is troch guon psychiatristen negearre, nettsjinsteande de betingst dy't serieuze psychososjale problemen feroarsaak. Neffens dizze auteurs sizze se dat seksuech-fersiken of hyperseksuele ûngelok ferskate termen foar itselde probleem fertsjintwurdigje. Se sjogge dat prevalenske tariven fan seksueu-ferslavingstreurs rûnom fan 3% nei 6%. Dêrtroch is it konstrukt fan 'e seksuele tafoeging / hyperseksual ûntefreging problematyske gedrachoanwizers, ûnder oaren: oertsjûge masturbaasje, cyberseks, pornografyske gebrûk, ferwiderjende seksueel gedrach mei ynkommens folwoeksenen, tillefoanseks, stripferieningsbeskikkings, en oare ferslavingsgedrach [98]. Geweldich geane wy oerien dat der ferskillende ferskillen tusken seksueel-ferslaving en hyperseksualiteit kin wurde lykas Carvalho, et al. [99], Rettenberger, et al. [100], Kor, et al. [1], Reid, et al. [9], Kafka en Hennen [13], en Prause, et al. [93-94] ûnder oaren.
Yn gearfetting hawwe wy foarsteld dat, wylst der ferskate ferskillen tusken hyperseksualiteit en seksukaasje binne, mear ûndersyk is nedich om dizze tige wichtige betingsten passend te kategorisearjen. Wy dogge it iens mei it wurk fan Walters, et al. [101] dat bepaalde dat de yndividuele ferskillen yn hyperseksualiteit kwantitatyf binne as kwalitatyf yn 'e natuer. Sy suggerje ek dat hypersekualiteit organisearre wurdt troch in kontinuïteums dy't falt op it heule ein fan 'e kontrôtomint (ôfbylding 1).

Ofbylding 1: In beskriuwende kaart fan hyperseksualiteitstart as subtype fan RDS
De figuer yllustreart ynteraktive neurogenetyske en epigenetyske effekten. Sawol koarte termyn dopamine-blokkearjende as lange-termyn "dopaminergyske-homeostasis" -basearre behannelingen en dopamine stimulearjende terapyen en deistige aktiviteiten wurde neamd. Sirkels jouwe RDS-skaaimerken oan en fakjes jouwe RDS-gedrach oan.
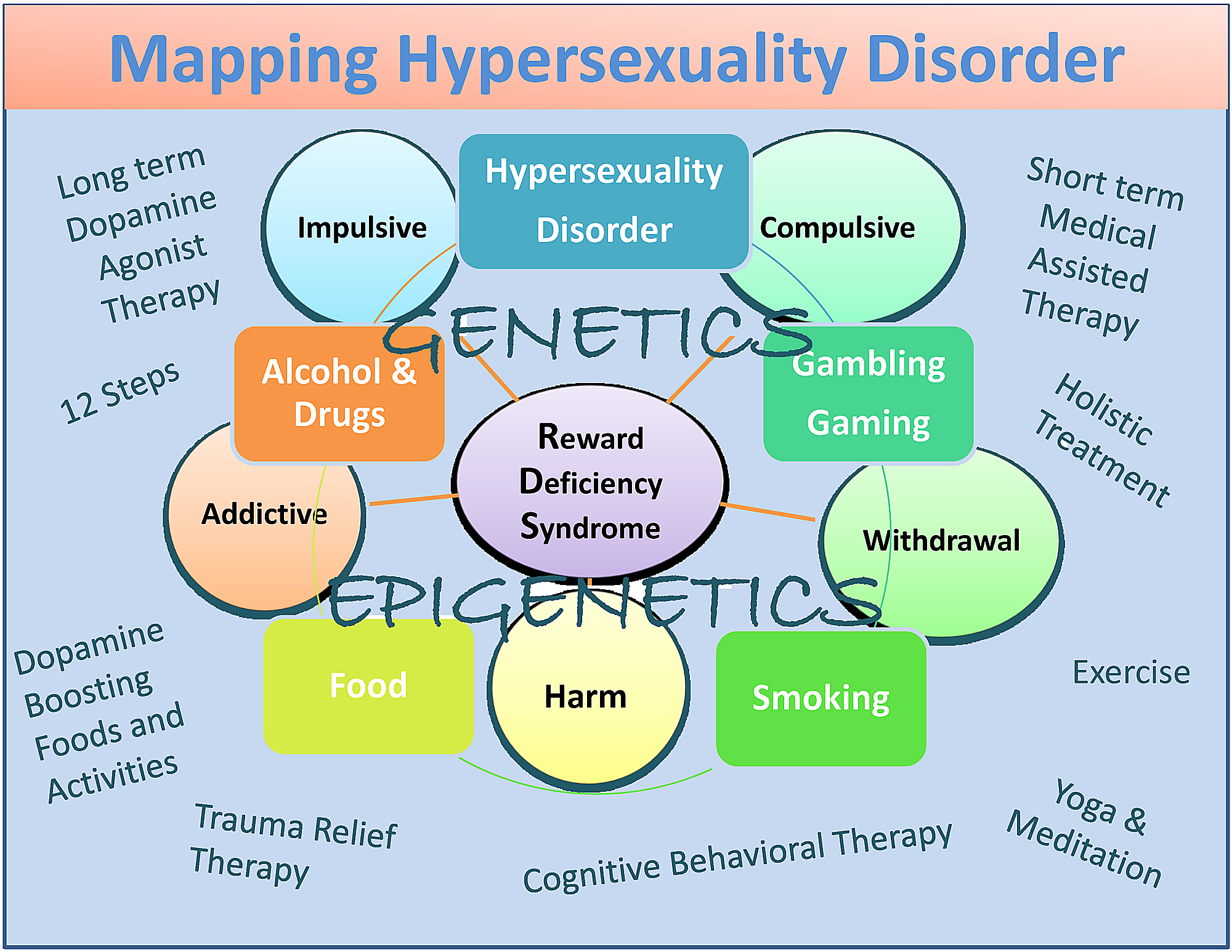{kind=link}
konklúzjes
Wylst wy de kontroversje werkenne, stelle wy foar dat mooglike ferskillen en oerienkomsten tusken hyperseksualiteitsteuring en seksferslaving adekwaat moatte wurde ûndersocht mei neuroimaging (fMRI, PET, SPECT), optogenetika, kandidaat- en mikroarray-analyse, en epigenetyske techniken. Wy leauwe dat dizze ûndersiken de basis sille leverje foar opnimmen fan hyperseksualiteit as steuring yn takomstige edysjes fan 'e DSM.
Referinsjes
- Kor A, Fogel Y, Reid RC, Potenza MN: Is it hyperseksuele disorder omskreaun as in bedriging?. Sex Addict Compulsivity. 2013, 20:http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3836191/. 10.1080/10720162.2013.768132
- Rush B: Medyske oanfragen en observaasjes oer de sykte fan 'e minske. Rush B (ed): Gryphon Editions Ltd., Birmingham, AL; (Oarspronklik wurk publisearre 1812) wurde yn 1979 hifke.
- von Krafft-Ebing R: Psychopathia Sexualis. Klaf FS (ed): Stein en dag, New York; (oarspronklik wurk 1886 publisearre) wurde yn 1965 evaluearre.
- Hirshfeld M: Seksuele anomalies: De oarsprong, natuer en behanneling fan seksueel ûnrjocht. Hirshfeld M (ed): Emerson Books, New York; 1948.
- Stroller RJ: Perversion: De eroatyske foarm fan haat. Pantheon Books, New York; 1975.
- Allen CA: In learboek fan psychoseksualen. Oxford University Press, Londen; 1962.
- Ellis A, Sagarin E: Nymphomania: In stúdzje fan 'e fergese fraude. Gilbert Press, New York; 1964.
- Kafka MP: Wat barde mei hyperseksuele ûngelok?. Arch Sex Behav. 2014, 43: 1259-1261. 10.1007 / s10508-014-0326-y
- Reid RC, Temko J, Moghaddam JF, Fong TW: Smaak, ruminaasje en selsmeilijen yn manlju beoardiele foar hyperseksuele ûngelok. J Psychiatriepraktyk. 2014, 20: 260-268. 10.1097 / 01.pra.0000452562.98286.c5
- Bancroft J: Human Sexuality en har problemen. Tredde edysje. Elsevier, Oxford, Ingelân; 2009.
- Ynternasjonale statistyske klassifikaasje fan sykte en relatearre sûnensproblemen, 10e Revision . (2007). Accessed: July 23, 2015: http://apps.who.int/classifications/apps/icd/icd10online2007/.
- Singer B, Toates FM: Seksuele motivaasje. J Seksûndersyk. 1987, 23: 481-501. 10.1080/00224498709551386
- Kafka MP, Hennen J: Hyperseksual winsk yn manlju: binne mantsjes mei paraphilia's ferskillend fan manlju mei paraphilia-ferbûne feroaringen?. Sex Abuse. 2003, 15: 307-321. 10.1023 / A: 1025000227956
- Buss DM, Schmitt DP: Seksuele strategystheorie: in evolúsjonêre perspektyf op 'e mantelsoarch. Psychol Rev. 1993, 100: 204-232. 10.1037 / 0033-295X.100.2.204
- Corbett-Detig RB, Hartl DL, Sackton TB: Natuerlike seleksje beskermje neutraal ferskaat oer in breed oanbod fan soarten, Psychol Bull. 1995, 13: e1002112.- 469-496. doi: 10.1371 / journal.pbio.1002112
- Laumann EO, Michael RT, Gagnon JH: In politike skiednis fan 'e nasjonale seksûndersyk fan folwoeksenen. Fam Plann Perspect. 1994, 26: 34-38. 10.2307/2136095
- Jones JC, Barlow DH: Self-rapporteare frekwinsje fan seksueel rinnende, fantasyen en masturbatorjende fantasys yn heteroseksuele mantsjes en froulju. Arch Sex Behav. 1990, 19: 269-79. 10.1007 / BF01541552
- Oliver MB, Hyde JS: Gender-ferskillen yn seksualiteit: in meta-analyze. Psychol Bull. 1993, 114: 29-51. 10.1037 / 0033-2909.114.1.29
- Okami P, Shackelford TK: Human sekondêre ferskillen yn seksuele psychology en gedrach, Annu Rev Seks Res. 2001, 12: 186-241. 10.1080/10532528.2001.10559798
- Leitenberg H, Detzer MJ, Srebnik D: Gleone ferskillen yn masturbaasje en de relaasje fan masturbaasje ûnderfining yn preadoleszenz en / of frjemde adolesinsje foar seksueel gedrach en seksueel oanpassing yn jonge folwoeksenens. Arch Sex Behav. 1993, 22: 87-98. 10.1007 / BF01542359
- Fetterman AK, Kruger NN, Robinson MD: Seksje-keppele matingstrategyen ûnderskiede mei in manipulaasje fan genitaltealens. Motyf Emot. 2015, 39: 99-103. 10.1007/s11031-014-9420-7
- Basson R: Mei in oar model foar seksueel antwurd fan froulju om problemen problematysk leech seksueel begearte oan te pakken, J Seks Marital Ther. 2001, 27: 395-403. 10.1080/713846827
- Andersen BL, Cyranowski JM, Aarestad S: Beyond artificial, sex-linked links to designate female sexuality: comment on Baumeister. Psychol Bull. 2000, 126: 380-389. 10.1037 // 0033-2909.126.3.380
- Ridder RA, Sims-Knight JE: De ûntwikkeling fan prestaasjes fan seksuele oandwaning tsjin froulju: testen alternatyf hypoteesjes mei struktureel equaasjemodeling. Ann NY Acad Sci. 2003, 989: 72-85. 10.1111 / j.1749-6632.2003.tb07294.x
- Blum K, Werner T, Carnes S, Carnes P, Bowirrat A, Giordano J, Oscar-Berman M, Goud M: Seks, drugs, en rock 'n' roll: hypoteze fan mienskiplike mesolimbyske aktivearring as in funksje fan beleanninggenpolymorfismen. J Psychoaktyske drugs. 2012, 44: 38-55. 10.1080/02791072.2012.662112
- Carnes PJ, Green BA, Merlo LJ, Polles A, Carnes S, Gold MS: PATHOS: in koarte screeningapplikaasje foar beoardieling fan seksuele ferslaving. J Addict Med. 2012, 6: 29-34. 10.1097/ADM.0b013e3182251a28
- Hanson RK: Static-99 fertelt residivisme ûnder âldere seksuiteurs?, Seks misbrûk. 2006, 18: 343–355. 10.1007 / s11194-006-9027-y
- Weinstein A, Katz L, Eberhardt H, Cohen K, Lejoyeux M: Seksuele twang - Ferhâlding mei seks, oanhing en seksuele oriïntaasje. J Behav Addict. 2015, 4: 22-26. 10.1556 / JBA.4.2015.1.6
- Chung WS, Lim SM, Yoo JH, Yoon H: Geschneld ferskil yn 'e harsetaktivaasje nei audio-fisuele seksuele stimulearring; Do frije froulju en manlju itselde nivo fan oprop as reaksje op deselde fideoklip?. Int J Impot Res. 2013, 25: 138-142. 10.1038 / ijir.2012.47
- Rupp HA, Wallen K: Seks ferskillen yn antwurd op fisuele siken stimulearje: in resinsje. Arch Sex Behav. 2008, 37: 206-218. 10.1007/s10508-007-9217-9
- Hanson RK, Harris AJ, Helmus L, Thornton D: Hetero-risiko-seksoerders meie net heech risiko wêze. J Interpers geweld. 2014, 29: 2792-813. 10.1177/0886260514526062
- Reid RC, Cyders MA, Moghaddam JF Fong TW: Psychometryske eigenskippen fan 'e Barratt Impulsiveness Skala yn pasjinten mei spielers, hyperseksualiteit, en methamphetamineôfhinklikens. Addict Behav. 2014, 39: 1640-1645. 10.1016 / j.addbeh.2013.11.008
- Reid RC, Carpenter BN, Hook JN, Garos S, Manning JC, Gilliland R, Cooper EB, McKittrick H, Davtian M, Fong T: Rapportearje fan befinings yn in DSM-5-ferdrach foar hyperseksuele ûngelok. J Sex Med. 2012, 9: 2868-2877. 10.1111 / j.1743-6109.2012.02936.x
- Garcia FD, Thibaut F: Seksuele fersiken. Am J Drug Alcohol Abuse. 2010, 36: 254-260. 10.3109/00952990.2010.503823
- Reid RC, Bramen JE, Anderson A, Cohen MS: Mempealiteit, emosjonele dysregulaasje, ympulsiviteit, en stressfeardigens tusken hyperseksuele pasjinten. J Clin Psychol. 2014, 70: 313-321. 10.1002 / jclp.22027
- Holder MK, Hadjimarkou MM, Zup SL, Blutstein T, Benham RS, McCarthy MM, Mong JA: Methamphetamine makket it froulike seksueel hâlden en fasilitearret neuroanale aktivearring yn 'e mediale amigdala en ventromediale nucleus fan' e hypothalamus. Psychoneuroendokrinology. 2010, 35: 197-208. 10.1016 / j.psyneuen.2009.06.005
- Spauwen LW, Niekamp AM, Hoebe CJ, Dukers-Muijrers NH: Drogene gebrûk, seksueel risiko-gedrach en seksueel trochferwiske ynfeksjes ûnder swingers: in kwaleksjeare stúdzje yn Nederlân. Sex Transm Infect. 2015, 91: 31-36. 10.1136 / sextrans-2014-051626
- Castelo-Branco C, Parera N, Mendoza N, Pérez-Campos E, Lete I, CEA groep: Alkohol- en drugsmisbrûk en risikoare seksuele gedrach yn jonge folwoeksen froulju. Gynecol Endocrinol. 2014, 30: 581-586. 10.3109/09513590.2014.910190
- Jia ZJ, Yan SY, Bao YP, Lian Z, Zhang HR, Liu ZM: Seksuele gedrachsdifferens tusken amphetamine-type stimulâns brûkers en heroine-brûkers. J Addict Med. 2013, 7: 422-427. 10.1097/ADM.0b013e3182a952b2
- Bruijnzeel AW: kappa-opioid receptor-symboalearjen en brain-belibjenfunksje. Brain Res Rev. 2009, 62: 127-146. 10.1016 / j.brainresrev.2009.09.008
- Orsini CA, Ginton G, Shimp KG, Avena NM, Gold MS, Setlow B: De konsumpsje fan iten en gewichtswinning nei bepaling fan 'e chronike amfetamine administraasje. Honger. 2014, 78: 76-80. 10.1016 / j.appet.2014.03.013
- Ibrahim C, Reynaert C: Hyperseksualiteit yn neurokognitive steuringen by âldere minsken - in wiidweidige oersjoch fan 'e literatuer en saakstúdzje. Psychiatrus Danub. 2014, 26: 36-40.
- Vallejo-Medina P, Sierra JC: Effekt fan drugsgebou en ynfloed fan ôfstimming op seksueel funksjonearjen yn in Spaanske manlike drug-ôfhinklike probleem: in multisite-stúdzje. J Sex Med. 2013, 10: 333-341. 10.1111 / j.1743-6109.2012.02977.x
- Baskerville TA, Douglas AJ: Dopamine en oxytocine-ynteraksjes dy't ûnderdielen behannele binne: potensjele bydragen foar gedrachsleuringen, CNS Neurosci Ther. 2010, 16: e92-123. 10.1111 / j.1755-5949.2010.00154.x
- Riebe CJ, Lee TT, Hill MN, Gorzalka BBB: Precipitated withdrawal counters the adverse effects of subchronic cannabinoid administration on male sex sexual behavior. Neurosci Lett. 2010, 472: 171-174. 10.1016 / j.neulet.2010.01.079
- Korda JB, Pfaus JG, Kellner CH, Goldstein I: Persistente genital oprop fan symptomen (PGAD): saakrapport fan lange termyn symptomatyk bestjoer mei electroconvulsive therapy. J Sex Med. 2009, 6: 2901-2909. 10.1111 / j.1743-6109.2009.01421.x
- Huang CJ, Liao HT, Yeh GC, Hung KL: Ferdieling fan HLA-DQB1 allele yn pasjinten mei Lytse-Levin-syndroam. J Clin Neurosci. 2012, 19: 628-630. 10.1016 / j.jocn.2011.08.020
- Blum K, Noble EP, Sheridan PJ, Montgomery A, Ritchie T, Jagadeeswaran P, Nogami H, Briggs AH, Cohn JB: Algemien feriening fan human dopamine D2 receptor-gene yn alkoholisme. JAMA. 1990, 263: 2055-2060. 10.1001 / jama.263.15.2055
- Blum K, Sheridan PJ, Hout RC, Braverman ER, Chen TJ, Cull JG, Comings DE: De D2 dopamine-receptor-gene as in determinant fan belestingsyndromsyndroam. JR Soc Med. 1996, 89: 396-400. 10.1177/014107689608900711
- Miller WB, Pasta DJ, MacMurray J, Chiu C, Wu H, Comings DE: Dopamine-receptor-genen binne ferbûn mei leeftiid op it earste geslachtsferkear. J Biosoc Sci. 1999, 31: 43-54. 10.1017 / S0021932099000437
- Hamann S, Herman RA, Nolan CL, Wallen K: Manlju en froulju ferskille yn amygdala antwurd op visuele seksuele stimulearingen. Nat Neurosci. 2004, 7: 411-416. 10.1038 / nn1208
- Quiñones-Jenab V: Wêrom binne froulju fan Venus en manlju fan Mars as se mekoar brûke?. Brain Res. 2006, 1126: 200-203. 10.1016 / j.brainres.2006.08.109
- Noble EP, Blum K, Khalsa ME, Ritchie T, Montgomery, A Wood RC, Fitch RJ, Ozkaragoz T, Sheridan PJ, Anglin MD, Paredes A, Treiman LJ, Sparkes RS: Algemien feriening fan it DXAMX Dopamine Receptor-gen mei kokaïneôfhinklikens. Drug Alcohol hinget ôf. 1993, 33: 271-285. 10.1016/0376-8716(93)90113-5
- Harrison PJ, Tunbridge EM: Catechol-O-methyltransferase (COMT): in geneus bydrage oan seksfersifers yn 'e harsensfunksje, en foar seksuele dimorphisme yn' e predisposysje nei psychiatryske störings. Neuropsychopharmakology. 2008, 33: 3037-3045. 10.1038 / sj.npp.1301543
- Guo G, Tong Y: Leeftiid yn it earste geslachtsferkear, genen, en sosjale kontekst: bewiis fan twilling en dopamine D4 receptor-gen. Demografy. 2006, 43: 747-769. 10.1353 / dem.2006.0029
- Bradley KC, Boulware MB, Jiang H, Doerge RW, Meisel RL, Mermelstein PG: Feroaringen yn 'e gene ekspresje binnen de nucleus accumbens en striatum folgjende seksuele ûnderfining. Genes Brain Behav. 2005, 4: 31-44. 10.1111 / j.1601-183X.2004.00093.x
- Rao BS, Raju TR, Meti BL: Fergrutte numerike tinens fan synapses yn 'e CA3-regio fan hippocampus en molekulêre lagen fan motor cortex nei selsstimulearring fan lokkich ûnderfining. Neuroscience. 1999, 91: 799-803. 10.1016/S0306-4522(99)00083-4
- Heshmati M: Kokakanaal-induktive LTP yn it ventral tegmentale gebiet: nije ynsjoggen yn meganisme en tiidskursus ljochtje de selleare substraten fan soarch. J neurophysiol. 2009, 101: 2735-2737. 10.1152 / jn.00127.2009
- Beaver KM, Wright JP, Walsh A: In gene-basearre evolúsjonêre ferklearring foar de feriening tusken misdiedigens en tal fan seks partners. Biodemografy Soc Biol. 2008, 54: 47-55. 10.1080/19485565.2008.9989131
- Santtila P, Jern P, Westberg L, Walum H, Pedersen CT, Eriksson E, Sandnabba N: It dopamintransportergen (DAT1) polymorfisme is assosjeare mei foartidige ejaculation. J Sex Med. 2010, 7: 1538-1546. 10.1111 / j.1743-6109.2009.01696.x
- Chen TJH, Blum K, Mathews D, Fisher L, Schnautz N, Braverman Er, Schoolfield J, Downs W, Blum SH, Mengucci J, Meshkin B, Arcuri V, Bajaj A, Waite RL, Foarige feriening fan sawol de Dopamine D2 Receptor (DRD2) en de Dopamine Transporter (DAT1) [1 bp Allele] genen mei pasykologysk agressyf gedrach, in klinyske subtype Reward Deficiency Syndrome (RDS) yn adolesinten. Gene Ther Mol Biol. 2007, 1: 93-112. Accessed: July 23, 2015: http://gtmb.org/pages/Vol11A/HTML/11._Chen_et_al,_93-102.htm.
- Burt SA, Mikolajewski AJ: Foarige bewiis dat spesifike kandidaatgenoaten ferbûn binne mei adolesinte-opset antisosjale gedrach. Agressearje Behav. 2008, 34: 437-445. 10.1002 / ab.20251
- Jóźków P, Słowińska-Lisowska M, Łaczmański Ł, Mędraś M: Polymorfe farianten fan neurotransmitterreceptorgenen kinne ynfloed ha op seksuele funksje by âldere manlju: gegevens út 'e HALS-stúdzje. Neuroendokrinology. 2013, 98: 51-59. 10.1159/000350324
- Ferkeap JM, DiClemente RJ, Brody GH, Philibert RA, Rose E: Ynteraksje tusken 5-HTTLPR polymorphisme en misbrûkhistoarje oer adolesinte Afro-Amerikaanske wyfkes 'gedrach fan gebrûk nei oanlieding fan in yntervinsje foar previnsje fan HIV. Prev Sci. 2014, 15: 257-267. 10.1007/s11121-013-0378-6
- Barrett CE, Keebaugh AC, Ahern TH, Bass CE, Terwilliger EF, Young LJ: Variaasje yn vasopressine-receptor (Avpr1a) makket it ekspresje ferskaat yn gedrach yn ferbân mei monogamy yn prairievels. Horm Behav. 2013, 63: 518-526. 10.1016 / j.yhbeh.2013.01.005
- Garcia JR, MacKillop J, Aller EL, Merriwether AM, Wilson DS, Lum JK: Ferienings tusken dopamine D4-receptor-genferskieding mei sawol misdiedigens en seksuele promoasje. PLoS One. 2010, 5: e14162. 10.1371 / journal.pone.0014162
- Daw J, Guo G: De ynfloed fan trije genen oer oft adolesinten antykonsepsje brûke, USA 1994-2002. Popul Stud (Camb). 2011, 65: 253-271. 10.1080/00324728.2011.598942
- Emanuele E, Brondino N, Pesenti S, Re S, Geroldi D: Genetyske oanbod op humoristyske stilen. Neuro Endokrinol Lett. 2007, 28: 815-821.
- Matsuda KI: Epigenetyske feroarings yn 'e estrogen-receptor α-gen promotor: gefolgen yn sosjeksoalyske gedrach. Front Neurosci. 2014, 8: 344. 10.3389 / fnins.2014.00344
- Rice WR, Friberg U, Gavrilets S: Homoseksualiteit fia kanalisearre seksuele ûntwikkeling: in testprotokol foar in nij epigenetysk model. Bioessays. 2013, 35: 764-770. 10.1002 / bies.201300033
- Gundersen B: Pair-bonding fia epigenetika. Nat Neurosci. 2013, 16: 779. 10.1038 / nn0713-779
- Wang H, Duclot F, Liu Y, Wang Z, Kabbaj M: Histone deacetylase ynhibitoren fasilitearje partnerfoarmingfoarming yn froulike prairiefollen. Nat Neurosci. 2013, 16: 919-924. 10.1038 / nn.3420
- Zeh JA, Zeh DW: Maternale erfenis, epigenetika en de evolúsje fan polyandry. Genetica. 2008, 134: 45-54. 10.1007 / s10709-007-9192-z
- Addad M, Lesiau A: Extraversion, neurotisme, ûngemurken en kriminaliteit. Med Law. 1989, 8: 611-622.
- Brodie R: Viro fan it tinken: De nije wittenskip fan 'e Mem. Hay House, Inc, New York, NY; 1996, pp 66.
- Hernnstein R, Murray C: De Bellkurve: yntelliginsje en klassenstruktuer yn it Amerikaanske libben. De Free Press, New York, NY; 1994.
- Smillie LD, Cooper AJ, Proitsi P, Powell JF, Pickering AD: Variation in DRD2 dopamine-gene prognose ekstravertieke persoanlikheid. Neurosci Lett. 2010, 468: 234-327. 10.1016 / j.neulet.2009.10.095
- Comings DE: The Gene Bomb. Does Higher Education and Advanced Technology Accelerate Selection of Genes for Learning Disorders, ADHD, Addictive and Disruptive Behavior?. Hope Press, Duarte CA; 1996.
- Comings DE: It Adolescent Problem Behavior Syndrome. The Gene Bomb. Does Higher Education and Advanced Technology Accelerate Selection of Genes for Learning Disorders, ADHD, Addictive and Disruptive Behaviors ?. Hope Press, Duarte CA; 1996. pp 91-94.
- Comings DE: Gene seleksje. The Gene Bomb. Does Higher Education and Advanced Technology Accelerate Selection of Genes for Learning Disorders, ADHD, Addictive and Disruptive Behaviors ?. Hope Press, Duarte CA; 1996. pp 89-90.
- Comings DE: De Berkeley Studie. The Gene Bomb. Does Higher Education and Advanced Technology Accelerate Selection of Genes for Learning Disorders, ADHD, Addictive and Disruptive Behaviors ?. Hope Press, Duarte Kalifornje; 1996. pp 105-210.
- Borsten J: Malibu Beach Recovery Diet Cookbook. Vidov Publishing Inc, Malibu, CA; 2015.
- Derbyshire KL, Grant JE: Begrippe seksuele gedrach: In oersicht fan 'e literatuer. J Behav Addict. 2015, 4: 37-43. 10.1556/2006.4.2015.003
- Nirenberg MJ: Dopamine agonist withdrawal syndroam: gefolgen foar pasjinten. Drugs Aging. 2013, 30: 587-592. 10.1007 / s40266-013-0090-z
- Grant JE, Brouwer JA, Potenza MN: De neurobiology fan substansje en gedrachsdoeken. CNS Spectr. 2006, 11: 924-930.
- Sakata K, Duke SM: Tekoart oan BDNF-ekspresje fia promotor IV fersteurt ekspresje fan monoamine-genen yn 'e frontale cortex en hippocampus. Neuroscience. 2014, 260: 265-75. 10.1016 / j.neuroscience.2013.12.013
- Blom K, Liu Y, Wang W, Wang Y, Zhang Y, Oscar-Berman M, Smolen A, Febo M, Han D, Simpatico T, Cronjé FJ, Demetrovics Z, Gold MS: rsfMRI-effekten fan KB220Z ™ op neurologyske paadpunten yn betrekkingstekstriis fan ynstinkende genotypearre heroine-ferwûnen. Postgrad Med. 2015, 127: 232-241.
- McLaughlin T, Blum K, Oscar-Berman M, Febo M, Demetrovics Z, Agan G, Fratantonio J, Gold MS: Mei it brûken fan de Neuroadaptagen KB200z ™ om ferskriklike, lichtige nachtmerken yn RDS-pasjinten te ferbetterjen: de rol fan fersterke, brain-reward, funksjonele ferbinings en dopaminergyske homeostasis. J Reward Defic Syndr. 2015, 1: 24-35. 10.17756 / jrds.2015-006
- Blum K, Thanos PK, Badgaiyan RD, Febo M, Oscar-Berman M, Fratantonio J, Demotrovics Z, Gold MS: Neurogenetik en genetheary foar belestingsyndiksysyndroom: geane wy nei it Promised Land?. Expert Opin Biol Ther. 2015, 5: 973-985. 10.1517/14712598.2015.1045871
- Joranby L, Pineda-Frost KY, Gouden MS: Oanfreegje oan iten en hinnebesjildsystemen, Seksuele ferslaving en kompulsiviteit. 2005, 12: 201–217. 10.1080/10720160500203765
- Râne PJ, Gouden MS: Drugsopname en hyperphagia: lessen fan tabak en oare drugs. Curr Pharm Des. 2011, 17: 1173-1179. 10.2174/138161211795656738
- Moholy M, Prause N, Proudfit, GH, Rahman A, Fong T: Seksuele winsken, net hyperseksualiteit, prestiizje selsregulearring fan seksueel oprop. Cogn Emot. 2015, 6: 1012.
- Praude N, Steele, VR, Staley C, Sabatinelli, D, Hajcak G: Modulaasje fan letpotse potensjele posysjes troch seksuele bylden yn problemen brûkers en kontrôles dy't ynstimd binne mei 'pornodigheid'. Biol Psychol. 2015, 109: 192-199. 10.1016 / j.biopsycho.2015.06.005
- Mechelmans DJ, Irvine M, Banca P, et al: Ferhege omtinken foar foardielen nei seksueel útwreide tekeningen yn yndividuen mei en sûnder ferrifeljende seksuele gedrach. PLoS One. 2014, 25, 9 (8): e105476. 10.1371 / journal.pone.0105476
- Blum K, Gardner E, Oscar-Berman M, goud M: "Like" en "wolle" keppele oan Reward Deficiency Syndrome (RDS): hypoteze fan differinsjale responsiviteit yn sirkwy fan harsensbelesting. Curr Pharm Des. 2012, 18 (1): 113-118.
- Hyman SE: Diagnostearjen fan 'e DSM: Diagnostykklassifikaasje needsaaklike fundamentale reformearring. Cerebrum. 2011, 2011: 6. Tagong: 2011 Apr 26: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3574782/.
- Casey BJ, Craddock N, Cuthbert BN, Hyman SE, Lee FS, Ressler KJ: DSM-5 en RDoC: fuortgong yn psychiatrysk ûndersyk?. Nat Rev Neurosci. 2013, 14: 810-14. 10.1038 / nrn3621
- Karila L, Wéry A, Weinstein A, Cottencin O, Petit A, Reynaud M, Billieux J: Seksuele ferslavings of hyperseksuele striid: ferskillende terminen foar deselde probleem? In oersicht fan 'e literatuer. Curr Pharm Des. 2014, 20: 4012-20. 10.2174/13816128113199990619
- Carvalho J, Štulhofer A, Vieira AL, Jurin T: Hyperseksualiteit en hege seksueel langstme: ûndersiikje de struktuer fan problematyske seksualiteit. J Sex Med. 2015, 12: 1356-67. Access: 2015 Mar 23: http://onlinelibrary.wiley.com/doi/10.1111/jsm.12865/abstract;jsessionid=6F416CCBB66B7F0EA28E428D4993EBD5.f01t04. 10.1111 / jsm.12865
- Rettenberger M, Klein V, Briken P: De relaasje tusken hyperseksuele gedrach, seksueel eksitatisaasje, seksualisaasje en persoanlikse teksten. Arch Sex Behav. 2015, Jan 6: (Epub foar print). Opfrege: 2015 Jan 6: 10.1007/s10508-014-0399-7
- Walters GD, Ridder RA, Långström N: Is hyperseksualiteit dimensjoneel? Bewissing foar DSM-5 fan algemiene befolking en klinyske problemen. Arch Sex Behav. 2011, 40: 1309-21. Tagong: 2011 Feb 3: 10.1007/s10508-010-9719-8