COMMENTI SU YBOP: l'effetto Coolidge è alla base del potere del porno su Internet. L'effetto Coolidge è un fenomeno osservato nelle specie di mammiferi in cui i maschi (e in misura minore le femmine) mostrano rinnovato interesse sessuale se introdotti a nuovi partner sessuali ricettivi, anche dopo aver rifiutato il sesso da partner sessuali precedenti ma ancora disponibili. Le novità sessuali prevalgono su questa abitudine con rinnovata eccitazione causata da una maggiore dopamina. Un flusso continuo di novità è ciò che rende il porno su Internet così diverso dal porno del passato.
Articolo originale, con grafici
- Dennis F. Fiorino,
- Ariane Coury e
- Anthony G. Phillips
- Il Journal of Neuroscience, 15 giugno 1997, 17 (12): 4849-4855;
Astratto
L'effetto Coolidge descrive la reiniziazione del comportamento sessuale in un animale "sessualmente saziato" in risposta a un nuovo compagno ricettivo. Dato il ruolo del sistema mesolimbico della dopamina (DA) nell'iniziazione e nel mantenimento del comportamento motivato, la microdialisi è stata utilizzata per monitorare la trasmissione DA del nucleo accumbens (NAC) durante la copulazione, la sazietà sessuale e la reiniziazione del comportamento sessuale. In accordo con le relazioni precedenti, la presentazione di una femmina estra dietro uno schermo e la copulazione sono stati associati ad aumenti significativi dell'efflusso di DA NAC. Il ritorno delle concentrazioni di DA della NAC ai valori basali coincide con un periodo di sazietà sessuale, sebbene le concentrazioni dei metaboliti DA, acido diidrossifenilacetico e acido omovanillico siano rimaste elevate. La presentazione di una nuova femmina recettiva dietro uno schermo ha comportato un leggero aumento del DA della NAC, che è stato aumentato significativamente durante la copulazione rinnovata con la nuova femmina. I dati attuali suggeriscono che le proprietà di stimolo di una nuova femmina ricettiva possono servire ad aumentare la trasmissione di DA NAC in un ratto maschio sessualmente sazio, e questo, a sua volta, può essere correlato alla reiniziazione del comportamento sessuale.
Introduzione
Un topo maschio che ha copulato con sazietà può essere indotto ad accoppiarsi di nuovo se la femmina iniziale viene sostituita con una nuova femmina ricettiva. Questo è diventato noto come effetto Coolidge ed è stato osservato in un certo numero di specie di mammiferi (Wilson et al., 1963). Fattori generali come la stanchezza o la depressione motorio non sono sufficienti a spiegare l'apparente stato di sazietà sessuale, perché gli stimoli di una nuova femmina possono ancora indurre copulazione. La sazietà sessuale può anche essere "invertita" farmacologicamente, in misura significativa, dalla somministrazione di una varietà di farmaci che possono agire su diversi sistemi di neurotrasmettitori. Questi farmaci comprendono la yohimbina, 8-OH-DPAT (Rodriguez-Manzo e Fernandez-Guasti, 1994, 1995a), nalaxone (Pfaus e Gorzalka, 1987; Rodriguez-Manzo e Fernandez-Guasti, 1995a,b) e apomorfina (Mas et al., 1995c). Sebbene l'azione periferica di questi farmaci non possa essere esclusa (ad esempio, effetti adrenergici sulla funzione erettile), gli effetti sui meccanismi centrali alla base della sazietà sessuale sono stati proposti sulla base di esperimenti di lesioni noradrenergiche centrali selettive (Rodriguez-Manzo e Fernandez-Guasti, 1995a) e gli esperimenti di microdialisi che monitoravano il metabolismo dopaminergico nell'area preottica mediale (Mas et al., 1995a,b).
Dato che i meccanismi centrali possono mediare il reintegro del comportamento sessuale caratteristico dell'effetto Coolidge, un probabile candidato è il sistema mesolimbico della dopamina (DA), proiettato dall'area tegmentale ventrale al NAC. Il DA mesolimbico sembra agire come un modulatore primario in processi integrativi complessi che implicano la valutazione di stimoli ambientali, come segnali di una femmina sessualmente recettiva, e l'organizzazione di comportamenti diretti all'obiettivo, inclusa la copulazione (Fibiger and Phillips, 1986; Blackburn et al., 1992; Phillips et al., 1992; LeMoal, 1995; Salamone, 1996).
Sebbene i neuroni DA mesencefalo rispondano a ricompense primarie e segnali predittivi di ricompensa, stimoli ambientali nuovi o imprevedibili inducono l'attivazione neuronale in modo più efficace rispetto a sessioni di allenamento ripetute (Fabre et al., 1983; Schultz, 1992; Mirenowicz e Schultz, 1994). Vi è una grande quantità di prove che supportano un ruolo facilitatore importante per la DA mesolimbica nell'iniziazione e nel mantenimento del comportamento sessuale dei ratti (Pfaus e Everitt, 1995) e numerosi studi di microdialisi riportano aumenti dell'efflusso di DA NAC durante le fasi appetitive e di consumo del comportamento sessuale maschile (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995b). Ci sono, tuttavia, relativamente pochi dati sui correlati neurochimici della sazietà sessuale e la reiniziazione del comportamento sessuale. L'applicazione di in vivo La microdialisi per monitorare la neurotrasmissione di DA mesolimbica durante l'effetto Coolidge offre un'opportunità unica per esaminare il ruolo di NAC DA nella copulazione, la sazietà sessuale e la reiniziazione della copulazione.
È stato condotto un esperimento di microdialisi per determinare quanto segue: (1) se l'insorgenza della sazietà sessuale è accompagnata dal ritorno di concentrazioni extracellulari di DA nel NAC ai valori di precopolazione o inferiori e (2) se la reintegrazione del comportamento copulatorio in un " "ratto maschio sessualmente sazio con una nuova femmina ricettiva è correlato con aumenti nell'efflusso di DA NAC.
MATERIALI E METODI
Soggetti. Ratti maschi Sprague Dawley, ottenuti dal Centro per la cura degli animali (presso l'Università della British Columbia), e ratti femmina Long-Evans, ottenuti da Charles River Canada (St. Constant, Quebec, Canada), sono stati alloggiati in gabbie di rete metallica (18 × 25 × 65 cm; cinque per gabbia) in stanze separate per colonia. Le stanze della colonia sono state mantenute a una temperatura di ~20 ° C su un ciclo 12 hr luce / buio invertito. I ratti avevano accesso illimitato al cibo (Purina Rat Chow) e all'acqua.
Chirurgia e test comportamentali prima della microdialisi cerebrale.I ratti femmina sono stati ovariectomizzati bilateralmente in anestesia con gas alotano (Fluothane, Ayerst Laboratories) almeno 4 settimane prima del test. La recettività sessuale nelle femmine di stimolo è stata indotta da iniezioni sottocutanee di estradiolo benzoato (10 μg) e progesterone (500 μg), 48 e 4 hr, rispettivamente, prima di ogni sessione di test. Ratti maschi sono stati sottoposti a screening per comportamento sessuale in due occasioni, 4 a parte, in camere di plexiglas (35 × 35 × 40 cm) con pavimenti in rete metallica. Solo ratti maschi che hanno raggiunto un criterio di prestazione, che includevano l'intromissione entro 5 min della presentazione della femmina e l'eiaculazione entro 15 min della prima intromissione, durante i due test di screening sono stati impiantati con cannule guida per sonda microdialitica.
Ratti maschi (n = 5) sono stati anestetizzati con ketamina cloridrato (100 mg / kg, ip) e xilazina (10 mg / kg, ip) prima dell'intervento stereotassico. Le cannule guida-sonda per microdialisi (misuratore 19) sono state impiantate bilateralmente sul NAC (coordinate da bregma: anteriore, + 1.7 mm, mediale, ± 1.1 mm, ventrale, -1.0 mm, cranio piatto) e sono state fissate al cranio con acrilico dentale e viti del gioielliere. Sono stati utilizzati impianti di cannule bilaterali di guida per massimizzare l'opportunità di un esperimento di microdialisi di successo. Fortunatamente, nel presente esperimento, era necessaria una sola cannula per ogni topo. Ratti maschi sono stati alloggiati individualmente in grandi gabbie di plastica con lettiera di pannocchia per il resto dell'esperimento. Una settimana dopo l'intervento, i ratti sono stati testati per comportamento sessuale. Durante questa parte dell'allenamento, la camera di prova era dotata di uno schermo scorrevole in plexiglas che divideva la camera in vani grandi e piccoli. Ratti maschi sono stati introdotti nel grande compartimento e 15 minuti dopo, una femmina è stata posizionata dietro lo schermo. Dopo un periodo di preparazione min di 15, lo schermo è stato rimosso e ai ratti è stato permesso di copulare per 30 min. Sono state condotte tre sessioni di formazione, una per ogni 4 d. Tutti i ratti hanno raggiunto il criterio delle prestazioni durante ogni sessione.
Esperimento effetto Coolidge. I ratti sono stati impiantati unilateralmente con le sonde per microdialisi 12-18 hr prima dell'esperimento dell'effetto Coolidge e collocati nell'ampio comparto della camera di test con libero accesso a cibo e acqua. La mattina dell'esperimento, sono stati raccolti campioni di microdialisi ogni 15 min. L'esperimento consisteva nelle seguenti sette fasi consecutive: (1) baseline (almeno 60 min); (2) 1 femmina dietro lo schermo (15 min); (3) copulazione con 1 femmina fino a quando un periodo 30 min è passato senza montatura; (4) reintroduzione della femmina 1 dietro lo schermo (15 min); (5) l'accesso alla femmina 1 per un periodo di 15 min a condizione che non ci fosse il montaggio (se si fosse verificato il montaggio, questa fase era trattata come fase 3); (6) introduzione della femmina 2 dietro lo schermo (15 min); 7) copulazione con 2 femmina per 60 min.
Il comportamento è stato filmato in condizioni di scarsa illuminazione utilizzando un sistema video JVC e osservato su un monitor video situato all'esterno della sala prove. Le misure standard del comportamento sessuale sono state registrate utilizzando un computer e un software appropriato (Holmes et al., 1987).
Dopo l'esperimento di microdialisi, agli animali è stato somministrato un sovradosaggio di idrato di cloralio e perfusi intracardialmente con soluzione salina e formalina (4%). I cervelli venivano tagliati e congelati e, successivamente, le sezioni coronali venivano colorate con il violetto cresilico per determinare il posizionamento delle sonde per microdialisi. Solo ratti con posizionamenti sonda all'interno del NAC sono stati utilizzati per analisi comportamentali e neurochimiche.
Microdialisi e rilevamento elettrochimico HPLC. Le sonde per microdialisi erano di concezione concentrica con una membrana a fibra cava semipermeabile (2 mm membrana esposta, 340 μm diametro esterno, 65000 taglio di peso molecolare, Filtral 12, Hospal) all'estremità distale. Le sonde sono state perfuse a 1.0 μl / min con una soluzione modificata di Ringer (0.01 m tampone fosfato di sodio, pH 7.4, 1.3 mmCaCl2, 3.0 mm KCl, 1.0 mmMgCl2, 147 mm NaCl) utilizzando una siringa a tenuta di gas (Hamilton, Reno, NV) e una pompa a siringa (modello 22, Harvard Apparatus, South Natick, MA). Per fissare la sonda per microdialisi all'interno della cannula di guida è stato utilizzato un collare di guida per sonda in microdialisi. Per proteggere il tubo della sonda è stata utilizzata una bobina di acciaio, fissata a un raccordo girevole liquido (Instech 375s) montato sulla parte superiore della camera di prova (Fiorino et al., 1993).
Gli analiti del microdializzato, che includevano DA e i suoi metaboliti, acido diidrossifililacetico (DOPAC) e acido omovanillico (HVA), sono stati separati mediante cromatografia in fase inversa (colonna Ultrasfera, Beckman, Fullerton, CA, ODS 5 μm, 15 cm, 4.6 mm, diametro interno ) utilizzando un tampone 0.083m per sodio acetato, pH 3.5 (5% metanolo). Le concentrazioni dell'analita sono state quantificate mediante rilevazione elettrochimica (CE). L'apparecchio consisteva in una pompa Bio-Rad (Richmond, CA), un iniettore a due posizioni EC10W Valco Instruments (Houston, TX), un rivelatore EC Coulochem II ESA (Bedford, MA) e un registratore grafico a doppio canale (Kipp e Zonen, Bohemia, NY). I parametri del rivelatore elettrochimico erano i seguenti: elettrodo 1, + 450 mV; elettrodo 2, -300 mV; e cella di guardia, -450 mV. Recuperi tipici della sonda, condotti in vitro ea temperatura ambiente, erano 22% per DA, 18% per DOPAC e 18% per HVA.
RISULTATI
Comportamento
Le misure comportamentali dall'esperimento dell'effetto Coolidge sono presentate in Tabella 1. Le latenze per il montaggio, l'intromissione e l'eiaculazione, nonché l'intervallo post-operatorio dopo la prima eiaculazione erano simili a quelli della sessione di allenamento precedente (dati non mostrati). Ciò suggerisce che la procedura di microdialisi non ha alterato il normale comportamento sessuale. Lo sviluppo della sazietà sessuale, come indicato dal numero medio di eiaculazioni prima che il criterio fosse soddisfatto (7.8 ± 0.5), una diminuzione progressiva del numero di intromissioni precedenti ogni eiaculazione e un progressivo aumento dell'intervallo post-operatorio (dati non mostrati) , era simile a quello riportato negli studi precedenti (Spiaggia e Giordania, 1956; Fowler e Whalen, 1961; Fisher, 1962; Bermant et al., 1966; Rodriguez-Manzo e Fernandez-Guasti, 1994; Mas et al., 1995d). È stata osservata una variabilità individuale rispetto al numero di eiaculazioni ottenute con 1 femminile, il tempo impiegato per copulare con 1 femmina e il numero di presentazioni di 1 femminile necessarie per raggiungere il criterio di sazietà (Tabella 1, parte inferiore). Alcuni ratti hanno richiesto numerose reintroduzioni di 1 femminile fino al completamento della fase 5 (n = 3). Gli atti di collocare 1 femmina dietro lo schermo e la rimozione della partizione potrebbero essere serviti da segnali appetitivi primari che hanno portato alla copulazione. Va anche notato che un criterio di saturazione di 30 min senza montatura, sebbene usato precedentemente (Beach e Jordan, 1965; Mas et al., 1995b), è arbitrario e non garantisce che un topo non avrebbe montato dato più tempo. Anche così, ritardi o procedure di rimozione e sostituzione non sono risultati affidabili in caso di copulazione rinnovata con 1 femmina (ad es. Fasi 4 e 5).

Comportamento durante l'esperimento con effetto Coolidge
Tutti i ratti hanno mostrato l'effetto Coolidge. L'attività associata alla collocazione di 2 femminile dietro lo schermo e, in particolare, la rimozione della partizione potrebbe aver contribuito a questo risultato, ma, ancora una volta, questi eventi non erano di per sé sufficienti per rinnovare la copulazione in precedenza nell'esperimento. I confronti tra le misure del comportamento sessuale con la femmina 1 e la femmina 2 sono stati fatti usando t prova con una correzione Bonferroni. Sebbene le latenze di montatura e di intromissione in risposta alla femmina 2 non differissero significativamente da quelle del primo episodio copulatorio con 1 femminile, in generale, il comportamento sessuale con 2 femminile era meno robusto, come indicato da un numero significativamente inferiore di eiaculazioni (media, 0.6 vs 4.2; F = 49.86;p <0.01) e intromissioni (media = 11.2 vs 37.0;F = 20.17; p <0.05) durante la prima ora. Il numero di cavalcature nella prima ora con le femmine 1 e 2 non era significativamente diverso.
È importante notare che le femmine utilizzate durante la porzione di sazietà dell'esperimento (cioè 1 femminile) mostravano ancora un forte comportamento procreativo (cioè saltellante e dardeggiante) e ricettivo (cioè lordosi) per tutta la durata del loro contatto con il maschio .
neurochimica
Le concentrazioni nanomolari basali di DA e dei suoi metaboliti in microdializzati, presentate come media ± SEM, dei primi tre campioni di riferimento erano: DA, 3.0 ± 0.7; DOPAC, 619.1 ± 77.7; e HVA, 234.2 ± 49.0 (non corretto per il recupero della sonda;n = 5). Questi valori rappresentano i punteggi di base 100%.
Per le analisi neurochimiche sono stati utilizzati punti dati comportamentalmente definiti, corrispondenti a ciascuna fase dell'esperimento e comuni a tutti i ratti. Questi hanno incluso i seguenti: (1) sette campioni dopo la prima introduzione di 1 femmina, (2) quattro campioni che accompagnavano l'assenza di comportamento copulatorio con 1 femmina e (3) cinque campioni dopo la presentazione di 2 femminile. figura 1 illustra i cambiamenti nelle concentrazioni di DA (grafico lineare, mezzo) e metaboliti della DA (grafico lineare, top) parallelamente al comportamento copulatorio (grafico a barre, fondo) durante il test per l'effetto Coolidge.
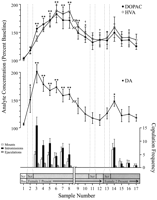
Visualizza la versione più grande:
Fig. 1.
Nucleo accumbens correlazioni neurochimiche del comportamento sessuale durante l'effetto Coolidge. I primi otto campioni rappresentano punti dati cronologicamente continui dalle fasi 1 a 3. Il campione 1 è il quarto e ultimo campione di riferimento di precopolazione (Basso). Esempio 2 rappresenta l'introduzione della femmina 1 dietro lo schermo (Scr). Dopo 15 min, lo schermo è stato rimosso ei ratti sono stati autorizzati a copulare (campioni 3-8). Ilrompere sul x-axis corrisponde all'esclusione dei dati da tre ratti che hanno copulato per lunghi periodi con la femmina iniziale. Gli ultimi nove campioni erano anche in ordine cronologico. I campioni 9 e 10 corrispondono al periodo di sazietà della fase 3 (cioè, 30 min senza montatura). 1 femmina è stata quindi reinserita dietro lo schermo (esempio 11) e, 15 min dopo, lo schermo è stato rimosso (esempio 12). Dopo 15 min privo di copulazione, 2 femmina è stata posizionata dietro lo schermo (esempio 13). I campioni 14-17 corrispondono alla copula con 2 femmina. Il numero di montature, intromissioni o eiaculazioni associate a ciascun campione di microdialisi 15 min è mostrato nel grafico a barre in basso. I dati neurochimici sono espressi in termini di percentuale delle concentrazioni basali. Cambiamenti in NAC DA (quadrati chiusi), DOPAC (cerchi chiusi) e HVA (cerchi aperti) efflux sono presentati come grafici a linee. Sono stati effettuati i seguenti confronti: campione di riferimento 1 rispetto a campioni 2-10; nuovo campione di riferimento 10 rispetto ai campioni 11 e 12; nuovo campione di riferimento 12 rispetto a campioni 13-17 (*p <0.05; ** p <0.01). Indipendente t sono stati fatti test tra valori di riferimento (campioni 1, 10 e 12). Per differenze significative rispetto al primo riferimento (campione 1), †p <0.05.
Sono stati eseguiti ANOVA a una via ripetuta, a misure ripetute su dati neurochimici associati a 1 femmina (campioni 1-12) e 2 femmina (campioni 12-17). A priori i confronti sono stati effettuati utilizzando il test di confronto multiplo di Dunn (Bonferroni t). Sono stati fatti i seguenti tre principali confronti: (1) linea di base iniziale (campione 1) rispetto ai campioni 2-10 (prima esposizione a 1 femminile), (2) secondo baseline (campione 10) rispetto ai campioni 11 e 12 (riesposizione a 1 femminile) e (3) terza baseline (campione 12) rispetto ai campioni 13-17 (esposizione a 2 femminile).
C'è stato un cambiamento generale significativo nell'efflux DA in risposta alla femmina 1 [F (11,44) = 8.48; p <0.001] e femmina 2 [F (5,20) = 2.83;p <0.05]. Un aumento significativo dell'efflusso di DA è stato riscontrato quando la femmina 1 era presente dietro lo schermo (+ 44%,p <0.05; campione 2). Durante la copulazione, le concentrazioni di DA aumentano ulteriormente, raggiungendo un valore massimo (+ 95%;p <0.01) durante il primo periodo di copulazione (campione 3). DA è rimasto elevato durante la copulazione ed è tornato alle concentrazioni basali solo nel periodo di 30 minuti in cui non si è verificato alcun montaggio (campioni 9 e 10). Né la reintroduzione della femmina 1 dietro lo schermo (campione 11) né l'opportunità di interagire fisicamente, ma senza il montaggio (campione 12), concentrazioni di DA elevate rispetto al secondo valore di base (campione 10). La presenza della femmina 2 dietro lo schermo (campione 13) ha determinato un piccolo aumento dell'efflusso di DA (12%) dal terzo valore di base (campione 12) che non ha raggiunto la significatività statistica. La copulazione rinnovata con la femmina 2 ha determinato un aumento significativo (34%) (p <0.05) nell'efflusso di DA durante il primo campione di copulazione (campione 14). Sebbene il comportamento copulatorio debole sia continuato nei successivi tre campioni, le concentrazioni di DA sono diminuite ai valori basali (campioni 15-17). Indipendente t test condotti su campioni "di riferimento" (es. 1, 10 e 12) hanno dimostrato che questi valori non erano significativamente differenti.
Nei tre ratti che hanno ripreso la copulazione quando è stata reintrodotta la femmina 1, le concentrazioni di DA NAC aumentavano quando 1 femmina era presente dietro lo schermo (intervallo, 25-47%) e durante la copulazione (intervallo, 13-37%), rispetto al campione appena prima della reintroduzione della femmina. Questi aumenti, tuttavia, si sono verificati solo quando il comportamento sessuale è stato vigoroso e ha portato all'eiaculazione.
Cambiamenti globali significativi in DOPAC [F (11,44) = 9.57; p <0.001] e HVA [F (11,44) = 12.47; p <0. 001] concentrazioni sono state trovate in risposta alla femmina 1, ma non alla femmina 2. Le concentrazioni dei metaboliti sono aumentate leggermente (+ 15% in entrambi i casi) durante la presentazione della femmina 1 dietro lo schermo (campione 2), ma questo non era significativo statisticamente. Ci sono stati, tuttavia, aumenti significativi nelle concentrazioni di DOPAC e HVA durante la copulazione (campioni 3–8), raggiungendo valori massimi (+80 e + 86%, rispettivamente; p <0.01) dopo 60 min (campione 6 in entrambi i casi). Sebbene le concentrazioni dei metaboliti diminuissero durante il periodo di inattività sessuale alla fine del contatto con la femmina 1 (campioni 9 e 10), le concentrazioni rimasero comunque elevate rispetto al basale (p <0.05 in entrambi i casi). La reintroduzione della femmina 1 dietro lo schermo (campione 11), l'accesso alla femmina 1 dopo la rimozione dello schermo (campione 12) e l'introduzione della femmina 2 (campione 13) non hanno comportato alcun cambiamento nelle concentrazioni dei metaboliti. Leggeri, ma statisticamente insignificanti, aumenti delle concentrazioni di DOPAC e HVA (+ 23% in entrambi i casi) rispetto al basale (campione 12) corrispondevano al primo periodo di copulazione con la femmina 2 (campione 14). Tuttavia, questo aumento è stato di breve durata ed è sceso ai valori basali per i restanti tre campioni (15-17). Indipendente ttest condotti su campioni "di riferimento" (cioè 1, 10 e 12) hanno indicato che il secondo e il terzo valore di riferimento (campioni 10 e 12, rispettivamente), sebbene non diversi tra loro, rimanevano elevati rispetto al primo campione di riferimento per DOPAC e HVA (p <0.05 in entrambi i casi).
Istologia
Le sonde per microdialisi erano situate nel NAC (Fig.2) in un intervallo che va da + 1.20 a + 1.70 mm da bregma (cranio piatto). C'era variabilità anche nel piano mediolaterale; i dati riflettono il campionamento dalla shell e dalle sottoregioni core del NAC.
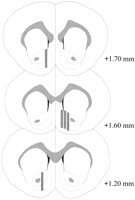
Visualizza la versione più grande:
Fig. 2.
Posizione delle sonde di microdialisi all'interno del NAC dei ratti maschi utilizzati nell'esperimento dell'effetto Coolidge. Rettangoli ombreggiati corrispondono all'area della membrana esposta delle sonde di microdialisi. Le sezioni del cervello coronale seriale sono state ridisegnate daPaxinos e Watson (1986).
DISCUSSIONE
In accordo con le relazioni precedenti, i risultati attuali dimostrano una trasmissione DA potenziata mesolimbica associata a componenti appetitive e di consumo del comportamento sessuale dei ratti maschi, valutata dain vivo microdialisi (Mas et al., 1990; Pfaus et al., 1990;Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995a,b,d). Inoltre, questi risultati forniscono un correlato neurochimico per la sazietà sessuale e la successiva reintegrazione della copulazione in risposta a una nuova femmina ricettiva (l'effetto Coolidge). I dati attuali suggeriscono che le proprietà di stimolo di una nuova femmina ricettiva possono servire ad aumentare la trasmissione della NAC DA in un ratto maschio sessualmente sazio che può, a sua volta, essere correlato alla reiniziazione del comportamento sessuale. Questo è evidente innanzitutto nel leggero aumento di NAC DA durante la presentazione di una nuova donna dietro lo schermo e si verifica in modo più convincente come un aumento più pronunciato durante la copulazione rinnovata con 2 femminile (Fig.1).
La presenza della prima femmina recettiva dietro lo schermo ha comportato un robusto aumento appetitivo nell'efflusso di NAC DA (44% dalla linea di base) simile in grandezza a quanto riportato in esperimenti precedenti usando un disegno simile (30%, Pfaus et al., 1990; 35%,Damsma et al., 1992). Anche in accordo con questi studi c'era l'osservazione che l'efflusso di NAC DA è stato ulteriormente migliorato durante la copulazione (a> 95% sopra la linea di base nel presente esperimento). Sebbene possiamo considerare i comportamenti consumistici come associati a una versione avanzata di NAC DA (Wenkstern et al., 1993; Wilson et al., 1995), è importante esaminare i termini "appetitivo" e "consumazione" nel contesto del comportamento sessuale. Mentre la fase in cui la femmina è presente dietro lo schermo è esclusivamente appetitiva o preparatoria, il comportamento durante la fase di copulazione non può essere considerato puramente consumativo. Poiché "appetitivo" può essere usato per descrivere tutti i comportamenti che portano al compimento di un comportamento motivato (copulazione), il comportamento primario che il maschio esibisce mentre è attivo nella fase "consummativa" è meglio descritto come appetitivo; il maschio passa la maggior parte del suo tempo e fatica a perseguire la femmina per copulare. A questo proposito, possiamo correlare la massima trasmissione DA di NAC con il consumo così come componenti appetitivi intensi del comportamento sessuale dei ratti maschi.
L'accesso alla seconda, nuova donna ha portato a una copulazione rinnovata in ogni materia. Precedenti studi hanno dimostrato che la maggior parte dei ratti autorizzati a copulare fino alla sazietà, utilizzando un protocollo comportamentale simile a quello usato nell'esperimento attuale, non ha ripreso l'accoppiamento quando testò 24 più tardi (Spiaggia e Giordania, 1956). È probabile che la presenza delle nuove proprietà di stimolo della femmina 2, che potrebbe aver incluso segnali olfattivi e visivi e uditivi, abbia portato a una copulazione rinnovata. Una domanda interessante, a cui si deve ancora rispondere, è in base a quale meccanismo un topo maschio distingue una femmina novella da una femmina con cui si è recentemente accoppiata. Un sito per quel meccanismo potrebbe trovarsi nel sistema olfattivo principale. È stato riportato che l'integrità di questo sistema è essenziale per l'effetto Coolidge nei criceti (Johnston e Rasmussen, 1984). Il sistema olfattivo accessorio vomeronasale, tuttavia, in cui un processo di memoria feromonale è stato descritto di recente nei topi (Kaba et al., 1994), è anche un candidato principale. A questo proposito, è interessante notare che gli aumenti della trasmissione NAC DA sono stati misurati usando in vivo voltammetria in ratti maschi presentati con lettiera che è stata esposta a ratti femmina in estro (Louillot et al., 1991; Mitchell e Gratton, 1992). Inoltre, l'applicazione di K+ direttamente allo strato nervoso vomeronasale del bulbo olfattivo accessorio, nonché al bulbo olfattivo accessorio stesso, era sufficiente per aumentare la trasmissione NAC DA (Mitchell e Gratton, 1992).
Il primo periodo minimo di copulazione 15 con la femmina 2 è stato associato ad un significativo aumento del DA della NAC. Al contrario della femmina 1, l'interazione con la femmina 2 non ha prodotto aumenti di NAC DA della stessa entità durante le fasi appetitive (12%) o consumatorie (34%). Questi piccoli aumenti di NAC DA, tuttavia, si correlano bene con il ridotto livello di comportamento sessuale visualizzato con 2 femminile rispetto a 1 femminile. Le concentrazioni di metaboliti sono rimaste elevate durante la fase di sazietà, risultando in nuove concentrazioni basali (campioni 10 e 12) che erano significativamente aumentati rispetto al valore iniziale della linea di base (campione 1).
Il ritardo temporale nell'aumento delle concentrazioni di DOPAC e HVA durante la copulazione è coerente con la loro formazione come metaboliti del composto progenitore, DA. È stato suggerito che le concentrazioni di metaboliti in microdialisi, almeno durante un comportamento naturale non guidato farmacologicamente, forniscono un utile indice dell'attività neuronale (Damsma et al., 1992; Fumero et al., 1994). Il fatto che le concentrazioni di metaboliti siano rimaste elevate anche durante i periodi di inattività sessuale in questo esperimento, quando le concentrazioni di DA erano tornate a valori di base pretest, getta dubbi su questo suggerimento.
L'aumento persistente delle concentrazioni di metaboliti DA osservate in questo esperimento rispecchia il profilo dell'area preoptica mediale (mPOA) dei metaboliti DA osservati nei ratti il primo giorno dopo che avevano copulato in sazietà (Mas et al., 1995a,b). Elevazioni sostenute di concentrazioni di DOPAC e HVA nel NAC o mPOA non sono sempre osservate quando il periodo dell'accoppiamento è di una durata fissa, molto più breve del tempo richiesto per raggiungere la sazietà. Ad esempio, molti studi hanno dimostrato che le concentrazioni di DOPAC sono aumentate e sono rimaste elevate durante la copulazione, ma sono diminuite ai valori di riferimento subito dopo la rimozione della femmina (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992;Hull et al., 1993; Wenkstern et al., 1993; Hull et al., 1995). Nello studio di Mas et al. (1995b)concentrazioni extracellulari basali di DOPAC e HVA nel mPOA sono rimaste elevate su 4 giorni consecutivi corrispondenti ad un periodo di inattività sessuale. Al quarto giorno, appena prima che gli animali riprendessero la copulazione, le concentrazioni basali dei metaboliti erano vicine ai valori di presatiazione. Gli autori hanno paragonato il modello dei cambiamenti neurochimici a quelli osservati dopo la somministrazione dei bloccanti dei recettori DA (Zetterström et al., 1984; Imperato e DiChiara, 1985) e hanno suggerito che lo stato di inattività sessuale può essere mediato dal rilascio di prolattina, che può agire come un "neurolettico endogeno" (Mas et al., 1995a,b,d). È chiaro che la somministrazione neurolettica è accompagnata da aumenti delle concentrazioni del metabolita extracellulare e dell'efflusso DA (Zetterström et al., 1984; Imperato e DiChiara, 1985). Sfortunatamente, Mas et al. (1995a,b) non sono stati in grado di rilevare le concentrazioni di mPOA DA. Nel presente studio, le concentrazioni di DA nel NAC sono tornate ai valori di precopolazione, mentre le concentrazioni di DOPAC e HVA sono rimaste elevate. Questo modello è incoerente con il ruolo di un'azione neurolettica endogena nel NAC per indurre sazietà sessuale.
Dato il coinvolgimento dei neuroni DA mesolimbici nel comportamento motivato (Fibiger and Phillips, 1986; Blackburn et al., 1992; Kalivas et al., 1993; LeMoal, 1995) e la loro sensibilità a nuovi stimoli ambientali (Fabre et al., 1983; Schultz, 1992; Mirenowicz e Schultz, 1994), gli aumenti osservati delle concentrazioni extracellulari di NAC DA in risposta alla nuova femmina sono coerenti con l'ipotesi che l'attività in questo sistema DA sia importante per la reiniziazione del comportamento sessuale. Inoltre, segnalazioni di aumenti appetitosi e di consumo nella trasmissione DA (Hull et al., 1993, 1995;Mas et al., 1995b; Sato et al., 1995) e attività neuronale (Shimura et al., 1994) nel mPOA di ratti maschi durante il comportamento sessuale suggeriscono che questa struttura può anche contribuire a una rinnovata caratteristica di copulazione dell'effetto Coolidge.
In linea con un ruolo generale del sistema mesolimbico di DA nel comportamento motivato, è ben noto che anche le concentrazioni extracellulari di DA sono elevate prima, durante e immediatamente dopo il consumo di un pasto, con un ritorno ai valori basali ~30 min dopo (Wilson et al., 1995). È noto che la sazietà indotta dal cibo è influenzata dalle sue proprietà sensoriali. Gli esseri umani e gli animali rifiutano il cibo sul quale sono stati nutriti per la sazietà e ingeriscono altri alimenti che non sono stati mangiati (Rotoli, 1986). Ciò solleva la questione se l'efflusso extracellulare di DA nel NAC sarebbe aumentato in modo selettivo dalla presentazione di un nuovo tipo di cibo, ma non dal cibo consumato di recente alla sazietà in un modo analogo a quello riportato nel presente studio nel contesto di motivazione sessuale Se confermata, questa relazione generale tra le proprietà sensoriali delle ricompense naturali, la sazietà e la trasmissione mesolimbica di DA implicherebbe un ruolo critico per questo sistema neurale nella regolazione dei processi motivazionali, la cui interruzione potrebbe portare a gravi disturbi del cibo e della funzione sessuale .
BIBLIOGRAFIA
- ↵
- Beach FA,
- Jordan L
(1956) Esaurimento sessuale e recupero nel ratto maschio. QJ Exp Psychol 8: 121-133.
- ↵
- Bermant G,
- Lott DF,
- Anderson L
(1966) Caratteristiche temporali dell'effetto Coolidge nel comportamento copulatorio del ratto maschio. J Comp Physiol Psychiatry 65: 447-452.
- ↵
- Blackburn JR,
- Pfaus JG,
- Phillips AG
(1992) Le funzioni della dopamina nei comportamenti appetitivi e difensivi. Prog Neurobiol 39: 247-279.
- ↵
- Damsma G,
- Pfaus JG,
- Wenkstern D,
- Phillips AG,
- Fibiger HC
(1992) Il comportamento sessuale aumenta la trasmissione di dopamina nel nucleo accumbens e nello striato dei ratti maschi: confronto con novità e locomozione. Behav Neurosci 106: 181-191.
- ↵
- Fabre M,
- Rolls ET,
- Ashton JP,
- Williams G
(1983) Attività dei neuroni nella regione tegmentale ventrale della scimmia che si comporta. Behav Brain Res 9: 213-235.
- ↵
- Fibiger HC,
- Phillips AG
(1986) Ricompensa, motivazione, cognizione: psicobiologia dei sistemi di dopamina mesotelencefalici. in Manuale di fisiologia: il sistema nervoso IV, edito Bloom FE, Geiger SD (American Physiology Society, Bethesda, MD), pp 647-675.
- ↵
- Fiorino DF,
- Coury AG,
- Fibiger HC,
- Phillips AG
(1993) La stimolazione elettrica dei siti di ricompensa nell'area tegmentale ventrale aumenta la trasmissione di dopamina nel nucleo accumbens del ratto. Behav Brain Res 55: 131-141.
- ↵
- Fisher A
(1962) Effetti della variazione dello stimolo sulla sazietà sessuale nel ratto maschio. J Comp Physiol Psychiatry 55: 614-620.
- ↵
- Fowler H,
- Whalen RE
(1961) Variazione dello stimolo incentivante e del comportamento sessuale nel ratto maschio. J Comp Physiol Psychiatry 54: 68-71.
- ↵
- Fumero B,
- Fernendez-Vera JR,
- Gonzalez-Mora JL,
- Mas M
(1994) Cambiamenti nel turnover delle monoamine nelle aree del proencefalo associate al comportamento sessuale maschile: uno studio di microdialisi. brain Res 662: 233-239.
- ↵
- Holmes GM,
- Holmes DG,
- Sachs BD
(1987) Un sistema di raccolta dati basato su IBM-PC per registrare il comportamento sessuale dei roditori e per la registrazione di eventi generali. Physiol Behav 44: 825-828.
- ↵
- Scafo EM,
- Eaton RC,
- Moses J,
- Lorrain DS
(1993) La copulazione aumenta l'attività della dopamina nella zona preottica mediale dei ratti maschi. Scienza della vita 52: 935-940.
- ↵
- Scafo EM,
- Jianfang D,
- Lorrain DS,
- Matuszewich L
(1995) La dopamina extracellulare nella zona preottica mediale: implicazioni per la motivazione sessuale e il controllo ormonale della copulazione. J Neurosci 15: 7465-7471.
- ↵
- Imperato A,
- DiChiara G
(1985) Rilascio e metabolismo della dopamina in ratti svegli dopo neurolettici sistemici come studiato dalla dialisi trans-striatale. J Neurosci 5: 297-306.
- ↵
- Johnston RE,
- Rasmussen K
(1984) Riconoscimento individuale di criceti femminili da parte dei maschi: ruolo dei segnali chimici e dei sistemi olfattivo e vomeronasale. Physiol Behav 33: 95-104.
- ↵
- Kaba H,
- Hayashi Y,
- Higuchi T,
- Nakanishi S
(1994) Induzione di una memoria olfattiva mediante l'attivazione di un recettore del glutammato metabotropico. Scienze 265: 262-264.
- ↵
- Kalivas PW,
- Sorg BA,
- Ganci MS
(1993) La farmacologia e il circuito neurale della sensibilizzazione agli psicostimolanti. Behav Pharmacol 4: 315-334.
- ↵
- LeMoal M
(1995) I neuroni dopaminergici mesocorticolimbici. Ruoli funzionali e normativi. in Psychopharmacology: la quarta generazione di progressi, edizioni Bloom FE, Kupfer DJ (Raven, New York), pp 283-294.
- ↵
- Louillot A,
- Gonzalez-Mora JL,
- Guadalupe T,
- Mas M
(1991) Gli stimoli olfattivi legati al sesso inducono un aumento selettivo del rilascio di dopamina nel nucleo accumbens dei ratti maschi. brain Res 553: 313-317.
- ↵
- Mas M,
- Gonzalez-Mora JL,
- Louillot A,
- Sole C,
- Guadalupe T
(1990) Aumento del rilascio di dopamina nel nucleo accumbens di copulanti ratti maschi come evidenziato dalla voltammetria in vivo. Neurosci Lett 110: 303-308.
- ↵
- Mas M,
- Fumero B,
- Fernandez-Vera JR,
- Gonzalez-Mora JL
(1995a) Correlati neurochimici dell'esaurimento e del recupero sessuale, valutati mediante microdialisi in vivo. brain Res 675: 13-19.
- ↵
- Mas M,
- Fumero B,
- Gonzalez-Mora JL
(1995b) Monitoraggio voltammetrico e in microdialisi del rilascio di neurotrasmettitore di monoamina cerebrale durante interazioni sociosessuali. Behav Brain Res 71: 69-79.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I
(1995c) Induzione del comportamento di accoppiamento da apomorfina in ratti sessualmente sazi. Eur J Pharmacol 280: 331-334.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I,
- Gonzalez-Mora JL
(1995d) La neurochimica della sazietà sessuale. Un modello sperimentale di desiderio inibito. in La farmacologia della funzione sessuale e disfunzione, e Bancroft J (Raven, New York), pp 115-126.
- ↵
- Mirenowicz J,
- Schultz W
(1994) Importanza dell'imprevedibilità per le risposte ricompensa nei neuroni dopaminergici primati. J Neurophysiol 72: 1024-1027.
- ↵
- Mitchell JB,
- Gratton A
(1992) Rilascio di dopamina mesolimbico provocato dall'attivazione del sistema olfattivo accessorio: uno studio cronoamperometrico ad alta velocità. Neurosci Lett 140: 81-84.
- ↵
- Paxinos G,
- Watson C
(1986) Il cervello di ratto in coordinate stereotassiche (2nd ndr). (Accademico, San Diego).
- ↵
- Pfaus JG,
- Damsma G,
- Nomikos GG,
- Wenkstern D,
- Blaha CD,
- Phillips AG,
- Fibiger HC
(1990) Il comportamento sessuale migliora la trasmissione centrale della dopamina nel ratto maschio. brain Res 530: 345-348.
- ↵
- Pfaus JG,
- Everitt BJ
(1995) La psicofarmacologia del comportamento sessuale. in Psychopharmacology: la quarta generazione di progressi, edizioni Bloom FE, Kupfer DJ (Raven, New York), pp 743-758.
- ↵
- Pfaus JG,
- Gorzalka BB
(1987) Oppioidi e comportamento sessuale. Neurosci Biobehav Rev 11: 1-34.
- ↵
- Phillips AG,
- Blaha CD,
- Pfaus JG,
- Blackburn JR
(1992) Correlati neurobiologici di stati emotivi positivi: dopamina, anticipazione e ricompensa. nella rivista internazionale di studi sull'emozione, a cura di Strongman (Wiley, New York), pp 31-50.
- ↵
- Pleim ET,
- Matochik JA,
- Barfield RJ,
- Auerbach SB
(1990) Correlazione del rilascio di dopamina nel nucleo accumbens con comportamento sessuale maschile nei ratti. brain Res 524: 160-163.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1994) Inversione dell'esaurimento sessuale da parte di agenti serotoninergici e noradrenergici. Behav Brain Res 62: 127-134.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995a) Partecipazione del sistema centrale noradrenergico al ristabilimento del comportamento copulatorio di ratti sessualmente esausti da parte di yohimbina, naloxone e 8-OH-DPAT. Cervello Res Bull 38: 399-404.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995b) Antagonisti degli oppioidi e il fenomeno della sazietà sessuale. Psychopharmacol 122: 131-136.
- ↵
- Rotoli BJ
(1986) Sazietà sensoriale specifica. Nutr Rev 44: 93-101.
- ↵
- Salamone JD
(1996) La neurochimica comportamentale della motivazione: aspetti metodologici e concettuali negli studi sull'attività dinamica del nucleo accumbens della dopamina. Metodi di J Neurosci 64: 137-149.
- ↵
- Sato Y,
- Wada H,
- Horita H,
- Suzuki N,
- Shibuya A,
- Adachi H,
- Kato R,
- Tsukamoto T,
- Kumamoto Y
(1995) Rilascio della dopamina nella zona preottica mediale durante il comportamento copulatorio maschile nei ratti. brain Res 692: 66-70.
- ↵
- Schultz W
(1992) Attività dei neuroni della dopamina nel primate che si comporta. Semin Neurosci 4: 129-138.
- ↵
- Shimura T,
- Yamamoto T,
- Shimokochi M
(1994) L'area preottica mediale è coinvolta sia nell'eccitazione sessuale che nelle prestazioni nei ratti maschi: ri-valutazione dell'attività dei neuroni negli animali che si muovono liberamente. brain Res 640: 215-222.
- ↵
- Wenkstern D,
- Pfaus JG,
- Fibiger HC
(1993) La trasmissione della dopamina aumenta nel nucleo accumbens dei ratti maschi durante la loro prima esposizione a ratti femmina sessualmente ricettivi. brain Res 618: 41-46.
- ↵
- Wilson C,
- Nomikos GG,
- Collu M,
- Fibiger HC
(1995) Correlati dopaminergici del comportamento motivato: importanza della pulsione. J Neurosci 15: 5169-5178.
- ↵
- Wilson JR,
- Kahn RE,
- Beach FA
(1963) Modifica nel comportamento sessuale dei ratti maschi prodotti cambiando lo stimolo femminile. J Comp Physiol Psychiatry 56: 636-644.
- ↵
- Zetterström T,
- Sharp T,
- Ungertedt U
(1984) Effetto dei farmaci neurolettici sul rilascio della dopamina striatale e sul metabolismo nel ratto sveglio studiato per dialisi intracerebrale. Eur J Pharmacol 106: 27-37.
Articoli che citano questo articolo
- Neuroplasticità Endogena Indotta Da Oppioidi Dei Neuroni Dopaminergici Nell'area Tegmentale Ventrale Influenza Ricompensa Naturale e Oppiacei Journal of Neuroscience, 25 June 2014, 34 (26): 8825-8836
- Squilibrio nella sensibilità a diversi tipi di ricompense nel gioco d'azzardo patologico Cervello, 1 August 2013, 136 (8): 2527-2538
- La legge sui premi naturali e sui farmaci sui meccanismi comuni di plasticità neurale con {Delta} FosB come mediatore chiave Journal of Neuroscience, 20 February 2013, 33 (8): 3434-3442
- Risposte neuronali nel guscio di Nucleus Accumbens durante il comportamento sessuale nei ratti maschi Journal of Neuroscience, 1 February 2012, 32 (5): 1672-1686
- Amore romantico: un sistema di cervello di mammifero per scelta del compagno Transazioni filosofiche della Royal Society B: Scienze biologiche, 29 December 2006, 361 (1476): 2173-2186
- Hot spot Hedonic nel cervello The Neuroscientist, 1 December 2006, 12 (6): 500-511
- Modulazione da nuclei amigdalari centrali e basolaterali di correlazioni dopaminergiche di alimentazione alla sazietà nel nucleo di ratto Accumbens e corteccia prefrontale mediale Journal of Neuroscience, 15 December 2002, 22 (24): 10958-10965
- Frequenza dei transienti di concentrazione della dopamina aumenta nello striato dorsale e ventrale dei ratti maschi durante l'introduzione dei conspecifici Journal of Neuroscience, 1 December 2002, 22 (23): 10477-10486
- Revisione del libro: Dinamica della dopamina extracellulare nelle azioni acute e croniche della cocaina Il neuroscienziato, 1 August 2002, 8 (4): 315-322
- Coinvolgimento differenziale di recettori NMDA, AMPA / Kainate e dopamina nel nucleo di Nucleus Accumbens nell'acquisizione e nelle prestazioni del comportamento di approccio pavloviano Journal of Neuroscience, 1 December 2001, 21 (23): 9471-9477
- Controllo del comportamento alla ricerca di cocaina da stimoli associati alla droga nei ratti: effetti sul recupero dei livelli di dopamina estinti a risposta operante e extracellulare nell'amigdala e nel nucleo accumbens PNAS, 11 April 2000, 97 (8): 4321-4326
- Facilitazione del comportamento sessuale e aumento dell'efflusso di dopamina nel nucleus accumbens dei ratti maschi dopo sensibilizzazione comportamentale indotta da D-anfetamina Journal of Neuroscience, 1 gennaio 1999, 19 (1): 456-463
- NEUROSCIENZE: Ottenere l'attenzione del cervello Science, 3 October 1997, 278 (5335): 35-37
- Correlati dopaminergici di sazietà sensoriale-specifica nella corteccia prefrontale mediale e nel nucleolo accumbens del ratto Journal of Neuroscience, 1 October 1999, 19 (19): RC29
