 ម៉ាទីនហ្សកក។
ម៉ាទីនហ្សកក។- 1Cognitive Psychopharmacology Laboratory, Neuroscience Department, Centre for Addiction and Mental Health, Toronto, ON, Canada
- 2Translational Neuroscience Program, Department of Psychiatry, School of Medicine, University of Pennsylvania, Philadelphia, PA, USA
- 3Biopsychology Section, Neuroscience Department, Centre for Addiction and Mental Health, Toronto, ON, Canada
Addiction is considered to be a brain disease caused by chronic exposure to drugs. Sensitization of brain dopamine (DA) systems partly mediates this effect. Pathological gambling (PG) is considered to be a behavioral addiction. Therefore, PG may be caused by chronic exposure to gambling. Identifying a gambling-induced sensitization of DA systems would support this possibility. Gambling rewards evoke DA release. One episode of slot machine play shifts the DA response from reward delivery to onset of cues (spinning reels) for reward, in line with temporal difference learning principles. Thus, conditioned stimuli (CS) play a key role in DA responses to gambling. In primates, DA response to a CS is strongest when reward probability is 50%. Under this schedule the CS elicits an expectancy of reward but provides no information about whether it will occur on a given trial. During gambling, a 50% schedule should elicit maximal DA release. This closely matches reward frequency (46%) on a commercial slot machine. DA release can contribute to sensitization, especially for amphetamine. Chronic exposure to a CS that predicts reward 50% of the time could mimic this effect. We tested this hypothesis in three studies with rats. Animals received 15 × 45-min exposures to a CS that predicted reward with a probability of 0, 25, 50, 75, or 100%. The CS was a light; the reward was a 10% sucrose solution. After training, rats received a sensitizing regimen of five separate doses (1 mg/kg) of d-amphetamine. Lastly they received a 0.5 or 1 mg/kg amphetamine challenge prior to a 90-min locomotor activity test. In all three studies the 50% group displayed greater activity than the other groups in response to both challenge doses. Effect sizes were modest but consistent, as reflected by a significant group × rank association (ϕ = 0.986, p = 0.025). Chronic exposure to a gambling-like schedule of reward predictive stimuli can promote sensitization to amphetamine much like exposure to amphetamine itself.
សេចក្តីផ្តើម
Addiction has been characterized as a brain disease caused by chronic exposure to drugs of abuse (Leshner, 1997 ។). Neuroplasticity is thought to mediate the effects of such exposure (Nestler, 2001). Sensitization of brain dopamine (DA) systems is a form of neuroplasticity implicated in hyper-reactivity to conditioned stimuli (CS) for drugs, and compulsive drug seeking (Robinson និង Berridge, 2001). Sensitization has been operationally defined by increased DA release in response to a CS for reward and by increased locomotor response to pharmacological DA challenge (Robinson និង Berridge, 1993; Pierce និង Kalivas, 1997; Vanderschuren និង Kalivas, 2000). Although sensitization is only one of many brain changes linked with addiction (cf. Robbins និង Everitt, 1999; Koob និង Le Moal, 2008), changes in presynaptic dopamine release have been suggested to represent common neuroadaptations involved in addiction-based drug-seeking (e.g., relapse), in that drugs that induce locomotor sensitization to opiate (e.g., morphine) or stimulant challenge (e.g., amphetamine), also cause reinstatement of extinguished operant responses for heroin or cocaine self-administration—an animal model of relapse (Vanderschuren et al ។ , 1999). Evidence that incentive sensitization (increased value of drug reward) is most pronounced after initial exposure to addictive drugs further suggests that sensitization may be involved in the early stages of addiction as well (Vanderschuren and Pierce, 2010).
Pathological gambling (PG) has been described as a behavioral addiction and recently reclassified to the same category as substance dependence disorders in the 5th edition of the Diagnostic and Statistical Manual of Mental Disorders (Frascella et al ។ , 2010; A.P.A., 2013). This implies that PG may be caused by chronic exposure to gambling-like activity, that common mechanisms may mediate the effects of gambling and drug exposure (Zack និង Poulos, 2009; Leeman និង Potenza, 2012); ហើយការប្រែលប្រួលផ្លូវខួរក្បាល DA អាចជាធាតុសំខាន់មួយនៃដំណើរការនេះ។
Clinical evidence indirectly supports this possibility: Using positron emission tomography (PET) Boileau and colleagues found that male PG subjects exhibit significantly greater striatal DA release in response to amphetamine (0.4 mg/kg) than healthy male controls (Boileau et al ។ , 2013). Overall group differences were significant in the associative and somatosensory striatum. In the limbic striatum, which includes the nucleus accumbens, the groups did not differ. However, in PG subjects, DA release in the limbic striatum correlated directly with the severity of PG symptoms. These findings are consistent with sensitization of brain DA pathways in PG, but also suggest some important differences with human substance dependent individuals and with the classic animal model of amphetamine sensitization. Unlike PG subjects and animals exposed to low doses of amphetamine (cf. Robinson et al ។ , 1982), humans with substance dependence consistently exhibit decreased DA release to a stimulant challenge (Volkow et al ។ , 1997; Martinez et al ។ , 2007), and evidence from animals suggests that this may reflect deficits in DA function during the initial stages of abstinence following binge patterns of substance abuse (Mateo et al ។ , 2005). In studies where stimulant sensitization is demonstrated in animals, enhanced DA release is usually observed in the limbic striatum rather than the dorsal (associative, somatosensory) striatum (Vezina, 2004). However, cue-induced (i.e., conditioned) drug-seeking in animals repeatedly exposed to cocaine has been linked with enhanced DA release in the dorsal striatum, a result thought to indicate a more habitual form of motivated behavior (Ito et al ។ , 2002). Thus, the overall elevation in DA release in dorsal regions in PG subjects may be related to habit-based (inflexible, routinized) reward seeking involving “a progression from ventral to more dorsal domains of the striatum” (Everitt និង Robbins, 2005, ទំ។ 1481) ចំណែកឯការចេញផ្សាយ DA ដែលពឹងផ្អែកលើភាពធ្ងន់ធ្ងរនៅក្នុងអវយវៈភេទនៅក្នុងមុខវិជ្ជាទាំងនេះអាចទាក់ទងកាន់តែជិតស្និទ្ធទៅនឹងការលើកទឹកចិត្តដែលមានលក្ខណៈជាគំរូនៅក្នុងសត្វ។ ការរកឃើញរបស់ PET មិនអាចបង្ហាញថាតើសកម្មភាពប្រតិកម្មខ្ពស់របស់ឌីអេសអេចគឺជាលក្ខណៈពិសេសដែលមានស្រាប់នៃមុខវិជ្ជាភី។ អេ។ ភីលទ្ធផលនៃការប៉ះពាល់នឹងល្បែងឬលទ្ធផលនៃដំណើរការផ្សេងទៀតមួយចំនួន។ ដើម្បីឆ្លើយសំនួរនេះវាចាំបាច់ត្រូវបង្ហាញពីការទាក់ទាញអារម្មណ៍ដោយការលេងល្បែងស៊ីសងរ៉ាំរ៉ៃលើមុខវិជ្ជាដែលមានលក្ខណៈធម្មតាមុនពេលប៉ះពាល់។ នេះធ្វើឱ្យមានសំណួរថាតើលក្ខណៈពិសេសនៃការលេងល្បែងអ្វីខ្លះដែលទំនងជាធ្វើឱ្យមានការចាប់អារម្មណ៍។
Skinner noted that the variable schedule of reinforcement was fundamental to gambling’s allure (or at least its persistence) (Skinner, 1953). Betting behavior in a slot machine game conforms well to the basic principles of instrumental conditioning, as reflected by a prospective correlation between monetary payoff and bet size on consecutive spins (Tremblay et al ។ , 2011 ។) ។ ដូច្នេះប្រតិបត្ដិសមាមាត្រសមាមាត្រឆ្លើយតបហាក់ដូចជាផ្តល់នូវគំរូដែលមានសុពលភាពខាងក្រៅនៃការលេងល្បែងម៉ាស៊ីនរន្ធដោត។
Recent research with animals provides strong initial support for a causal effect of gambling exposure on sensitization. Singer and colleagues examined the effects of 55 1–h daily sessions of fixed (FR20) or variable (VR20) saccharin reinforcement in an operant lever-press paradigm on subsequent locomotor response to low dose (0.5 mg/kg) amphetamine in healthy male (Sprague Dawley) rats (អ្នកចម្រៀងនិងអ្នកផ្សេងទៀត, 2012) ។ ពួកគេបានសន្មតថាប្រសិនបើការលេងល្បែងនាំឱ្យមានការចាប់អារម្មណ៍សត្វកណ្តុរដែលប៉ះពាល់នឹងកាលវិភាគប្រែប្រួលដែលធ្វើត្រាប់តាមការលេងល្បែងស៊ីសងគួរតែបង្ហាញនូវការឆ្លើយតបកាន់តែច្រើនទៅនឹងថ្នាំអំហ្វេតាមីនជាងកណ្តុរដែលទាក់ទងនឹងកាលវិភាគថេរ។ ដូចដែលបានព្យាករណ៍ក្រុម VR20 បង្ហាញការឆ្លើយតបក្បាលរថភ្លើងកាន់តែច្រើន 50% ទៅនឹងថ្នាំអំហ្វេតាមីនជាងក្រុម FR20 ។ ផ្ទុយទៅវិញក្រុមទាំងនោះបានបង្ហាញក្បាលម៉ាស៊ីនស្មើគ្នាបន្ទាប់ពីចាក់បញ្ចូលជាតិប្រៃ។ ការរកឃើញទាំងនេះបញ្ជាក់ថាការប៉ះពាល់រ៉ាំរ៉ៃទៅនឹងការពង្រឹងអថេរគឺគ្រប់គ្រាន់ដើម្បីជំរុញឱ្យមានប្រតិកម្មខ្ពស់ចំពោះបញ្ហាប្រឈមរបស់ឌីអេក្នុងសត្វដែលមានសុខភាពល្អចៃដន្យទៅតាមកាលវិភាគរៀងៗខ្លួន។
A number of questions arise from this result: First, to what extent does the perceived contingency—or lack thereof—between the operant response and its outcome mediate these effects? In learning terms, does this effect involve a “response-outcome expectancy,” or might a similar effect be seen in the absence of an operant response, i.e., “a stimulus-outcome expectancy” in a Pavlovian paradigm (cf. Bolles, ឆ្នាំ ១៩៧២)? ទីពីរតើកម្រិតនៃភាពជាប់លាប់រវាងព្រឹត្តិការណ៍ចាស់ (ការឆ្លើយតបឬការរំញោច) និងលទ្ធផលរបស់វាជះឥទ្ធិពលដល់កម្រិតនៃការប្រែលប្រួល?
សំណួរទីពីរទាក់ទងនឹងតួនាទីនៃភាពមិនច្បាស់លាស់នៅក្នុងការប្រែលប្រួល។ ឧទាហរណ៍តើហ្គេមដែលមានលទ្ធផលចៃដន្យ - មិនអាចទាយទុកជាមុនបាន - មានសក្តានុពលក្នុងការទាក់ទាញអារម្មណ៍ជាងហ្គេមដែលហាងឆេងនៃការឈ្នះត្រូវបានកំណត់យ៉ាងច្បាស់ប៉ុន្តែមិនចៃដន្យទោះបីជាអត្រានៃរង្វាន់គឺទាបក៏ដោយ? ការស្រាវជ្រាវបច្ចុប្បន្នបានឆ្លើយសំណួរទាំងនេះ។
The experimental design was informed by a seminal study on reward expectancy and DA neuron response in monkeys (Fiorillo et al ។ , 2003) ។ សត្វនៅក្នុងការសិក្សានោះទទួលបានរង្វាន់ទឹក (អាមេរិក) ក្រោម 0, 25, 50, 75, ឬតារាងសមាមាត្រអថេរ 100% ។ កាលវិភាគត្រូវបានកំណត់ដោយ 1 នៃ 4 ផ្សេងគ្នា CS (រូបតំណាង) ។ កាលវិភាគ 0% បានផ្តល់រង្វាន់ជាញឹកញាប់ដូចកាលវិភាគ 100% ប៉ុន្តែបានលុប CS ។ អត្រានៃការបាញ់ថ្នាំណឺរ៉ូនក្នុងចន្លោះពេលរវាងការចាប់ផ្តើមស៊ីអេសអេសនិងការបញ្ជូនឬការលុបចោលរបស់សហរដ្ឋអាមេរិកគឺជាវិធានការណ៍ពឹងផ្អែកសំខាន់។ ការសិក្សាបានរកឃើញថាការឆ្លើយតបរបស់ឌីអេអេបានកើនឡើងជាមុខងារនៃភាពមិនប្រាកដប្រជានៃការផ្តល់រង្វាន់។ ដូច្នេះក្រោមកាលវិភាគ 100% CS បានធ្វើឱ្យសកម្មភាពតិចតួចកើតឡើងក្រោមកាលវិភាគ 25 និង 75% ស៊ីអេសអេសបានធ្វើសកម្មភាពកម្រិតមធ្យមនិងប្រហាក់ប្រហែលហើយក្រោមកាលវិភាគ 50% សកម្មភាពស៊ីអេសអេសបានធ្វើសកម្មភាពអតិបរមា។ ក្នុងករណីនិមួយៗអត្រានៃការបាញ់បានកើនឡើងក្នុងចន្លោះពេលនៃស៊ីអេស - អាមេរិកពោលគឺនៅពេលដែលការរំពឹងទុកឈានដល់លទ្ធផល។
ការរកឃើញទាំងនេះបង្ហាញថាសកម្មភាពរបស់ឌីអេសមិនត្រឹមតែមានភាពខុសប្លែកគ្នាថាតើរង្វាន់ជាក់លាក់ (សមាមាត្រថេរ) ឬមិនប្រាកដប្រជាទេ (សមាមាត្រអថេរ) ប៉ុន្តែក៏មានសមាមាត្រផ្ទុយទៅនឹងបរិមាណព័ត៌មានអំពីការផ្តល់រង្វាន់ដែលបានប្រគល់ជូនដោយអេស។ នៅក្នុងលក្ខខណ្ឌ 100%, ស៊ីអេសអេចគេចផុតពីការរំពឹងទុករង្វាន់ហើយព្យាករណ៍ថាការបញ្ជូនរបស់វានឹងឥតខ្ចោះ។ នៅក្នុងលក្ខខណ្ឌ 25 និង 75%, ស៊ីអេសអេសបណ្តាលឱ្យមានអាយុកាលរំពឹងទុកនិងព្យាករណ៍ពីការផ្តល់រង្វាន់ចំនួនបីក្នុងចំណោមបួនដង។ នៅក្នុងលក្ខខណ្ឌ 50% ស៊ីអេសបានធ្វើឱ្យមានការរំពឹងទុកប៉ុន្តែមិនមានព័ត៌មានអំពីការផ្តល់រង្វាន់លើសពីឱកាសតែមួយគត់។ ផ្អែកលើការរកឃើញរបស់ពួកគេ Fiorillo et al ។ បានសន្និដ្ឋានថា“ ការកើនឡើង dopamine ដែលបណ្តាលមកពីភាពមិនច្បាស់លាស់អាចជួយឱ្យមានលក្ខណៈសម្បត្តិល្អនៃការលេងល្បែង” (ទំព័រ 1901) ។
The effects of 50% variable reward in a single session should not change over the course of multiple sessions because the likelihood of reward remains entirely unpredictable on every trial. Thus, when considering the conditions that would maximize chronic activation of DA neurons over repeated episodes of gambling the 50% schedule should engender the most enduring as well as the most robust effect. This is noteworthy given that the long run rate of reward (payoff > 0) observed over thousands of spins on a commercial slot machine was 45.8% (Tremblay et al ។ , 2011 ។) ។ ដូច្នេះរង្វាន់អថេរ 50% ហាក់ដូចជាឆ្លុះបញ្ចាំងពីកាលវិភាគទូទាត់ដែលគ្រប់គ្រងដោយឧបករណ៍លេងល្បែងជាក់ស្តែង។
ការសិក្សានាពេលបច្ចុប្បន្ននេះបានប្រើកាលៈទេសៈដូចគ្នានឹង Fiorillo et al ។ នៅក្នុងការប៉ះពាល់រ៉ាំរ៉ៃការរចនារវាងក្រុមនិងកណ្តុរ។ សត្វបានឆ្លងកាត់ ~ ៣ សប្តាហ៍នៃវគ្គម៉ាស៊ីនត្រជាក់ប្រចាំថ្ងៃដែលស៊ីអេស (ពន្លឺ) ត្រូវបានផ្គូរផ្គងជាមួយសហរដ្ឋអាមេរិក (ចំនួនទឹកក្រូចតិច) ។ បន្ទាប់ពីដំណាក់កាលបណ្តុះបណ្តាលសត្វបានសម្រាកមុនពេលការវាយតម្លៃនៃការប្រែលប្រួលសន្ទស្សន៍ដោយការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹងថ្នាំអំហ្វេតាមីន។ ផ្អែកលើអក្សរសិល្ប៍វាត្រូវបានគេព្យាករណ៍ថាកណ្តុរដែលត្រូវបានប៉ះពាល់នឹងកាលវិភាគរង្វាន់ផ្សេងៗនឹងមិនខុសគ្នានៅក្នុងឥរិយាបថក្បាលម៉ាស៊ីនសេរីរបស់ថ្នាំទេប៉ុន្តែនឹងបង្ហាញកម្រិតខុសគ្នានៃក្បាលរថភ្លើងបន្ទាប់ពីថ្នាំអំហ្វេតាមីនជាមួយនឹងក្រុម ៥០ ភាគរយបង្ហាញការឆ្លើយតបជាយថាហេតុកាន់តែច្រើនចំពោះថ្នាំដែលទាក់ទងទៅនឹងថ្នាំ។ ក្រុមផ្សេងទៀតក្នុងរយៈពេលដូសនៃថ្នាំដែលជាគំរូមួយដែលនឹងត្រូវបានគេរំពឹងទុកប្រសិនបើសត្វ ៥០ ភាគរយធ្លាប់ត្រូវបានប៉ះពាល់ទៅនឹងកម្រិតនៃថ្នាំអំហ្វេតាមីនដដែល (ឧទាហរណ៍ការឆ្លាស់គ្នា) ។
ពិសោធន៍ 1
សំភារៈនិងវិធីសាស្រ្ត
ប្រធានបទ
បួនក្រុម (n = សត្វកណ្តុរមនុស្សពេញវ័យ (8) 300 ក្រាម) នៃសត្វកណ្តុរ Sprague-Dawley (ទន្លេ Charles, St. Constant, Quebec, Canada) ត្រូវបានដាក់ជាលក្ខណៈបុគ្គលនៅក្នុងប្រអប់ប៉ូលីកាបូណាតច្បាស់ (350 × 20 × 43 សង់ទីម៉ែត្រ) ក្រោម 22 បញ្ច្រាស៖ វដ្តពន្លឺងងឹត 12 ។ ពួកគេបានទទួល។ ការផ្សាយពាណិជ្ជកម្ម access to food and water, and daily handling by an experimenter for 2 weeks prior to the study. Each group was conditioned under one of four variable reward schedules: 0, 25, 50, or 100%. The 75% group was omitted in this initial study, as Fiorillo et al. (2003) found equivalent post-CS DA release under 25 and 75% reward schedules, such that both conditions led to greater DA release than did the 100% CS-US condition, but less than the 50% condition.
បរិធាន
ការចូលទៅកាន់បទបង្ហាញ sucrose និង CS ត្រូវបានផ្តល់ជូនជាលក្ខណៈបុគ្គលនៅក្នុងប្រអប់ម៉ាស៊ីនប្រតិបត្តិការ (33 × 31 × 29 សង់ទីម៉ែត្រ) ។ ប្រអប់នីមួយៗត្រូវបានបំពាក់ដោយទស្សនាវដ្តីពង្រឹងដែលមានទីតាំងនៅជញ្ជាំងខាងមុខ។ ពន្លឺនៅលើកំពូលនៃទស្សនាវដ្តីបានបម្រើការជាអេសអេស។ ឌុយទ័ររាវដែលគ្រប់គ្រងដោយម៉ូទ័រអាចត្រូវបានដំឡើងនៅជាន់នៃទស្សនាវដ្តី។ ព្រឹត្តិការណ៍ក្នុងប្រអប់ត្រូវបានគ្រប់គ្រងដោយឧបករណ៍និងសូហ្វវែរឌែលេសដោយប្រើកម្មវិធីនៅក្នុងផ្ទះសរសេរនៅក្នុង MED-PC ។ ការធ្វើតេស្តក្បាលម៉ាស៊ីនត្រូវបានធ្វើឡើងជាលក្ខណៈបុគ្គលនៅក្នុងទ្រុង Plexiglas (27 × 48 × 20 សង់ទីម៉ែត្រ) ។ ទ្រុងនីមួយៗត្រូវបានបំពាក់ដោយប្រព័ន្ធត្រួតពិនិត្យដែលមានកោសិកាធ្នឹមរូបថតចំនួន ៦ ដើម្បីចាប់យកចលនាផ្តេក។
និតិវិធី
ការបណ្តុះបណ្តាល។ ការសិក្សានេះត្រូវបានធ្វើឡើងដោយអនុលោមតាមគោលការណ៍សីលធម៌ដែលបានកំណត់ដោយក្រុមប្រឹក្សាកាណាដាស្តីពីការថែទាំសត្វ។ កណ្តុរត្រូវបានដាក់កម្រិតលើចំណីអាហារដល់ 90% នៃទំងន់រាងកាយរបស់ពួកគេសម្រាប់រយៈពេលនៃការសិក្សានិងដាក់ជាលក្ខណៈបុគ្គល។ កណ្តុរនីមួយៗទទួលបានការបណ្តុះបណ្តាលរង្វាន់ XcrX ថ្ងៃចំនួន 15 (ដំណោះស្រាយទឹក 10% នៅ 0.06 មីល្លីលក្នុងមួយរង្វាន់) ៈ 5 ថ្ងៃជាប់ៗគ្នា weeks 3 សប្តាហ៍ដោយមានថ្ងៃចុងសប្តាហ៍។ សត្វត្រូវបានថែរក្សានៅលើជង្រុកស្តង់ដារមុននិងក្រោយវគ្គបណ្តុះបណ្តាល។ ការប៉ះពាល់ sucrose ត្រូវបានកំណត់ត្រឹមវគ្គបណ្តុះបណ្តាលដប់ប្រាំ ~ 40 នាទី។ សម័យប្រជុំនីមួយៗមានបទបង្ហាញរំញោច 15 (ពន្លឺមួយស៊ីអេសអេស) ដែលនីមួយៗត្រូវបានបំបែកដោយចន្លោះពេលនៃការសាកល្បង 120 s ។ ពន្លឺត្រូវបានគេដែលមានទីតាំងស្ថិតនៅបន្ទះខាងលើនៃទស្សនាវដ្តីហើយនៅតែបន្តសម្រាប់ 25 s ជាមួយនឹង sucrose ដែលអាចរកបានក្នុងកំឡុងពេល 5 ចុងក្រោយ។ ក្នុងករណីក្រុម 0 ដាប់ប៊្លុកស៊ុលត្រូវបានលើកឡើងរាល់ដង 140 s (សម្រាប់ 5 s) ប៉ុន្តែពន្លឺរំញោចមិនត្រូវបានបំភ្លឺទេ។ នេះស្មើនឹងចន្លោះពេលរវាងបទបង្ហាញរបស់ឌិបភឺរនៅក្នុងក្រុម 0 និងក្រុមផ្សេងទៀត (120 + 25 s) ។ វគ្គព្យាបាលនីមួយៗមានរយៈពេល ~ 40 នាទី។ ជាមធ្យមក្រុម 25 ទទួលបាន sucrose ម្តងសម្រាប់រាល់បទបង្ហាញរបស់ស៊ីបួន។ ក្រុម 50 ទទួលបាន sucrose ម្តងសម្រាប់រាល់បទបង្ហាញពីររបស់ CS ហើយក្រុម 100 ទទួល sucrose បន្ទាប់ពីរាល់បទបង្ហាញរបស់ CS ។
ការសាកល្បង។ ពីរសប្តាហ៍បន្ទាប់ពីការចូលប្រើ sucrose ចុងក្រោយ (ឬ“ ម៉ាស៊ីនត្រជាក់”) វគ្គឆ្លើយតបនឹងក្បាលរថភ្លើងទៅនឹងអេមហ្វេតាមីន (AMPH; ip) ត្រូវបានគេវាយតម្លៃ។ សត្វកណ្តុរត្រូវបានផ្តល់វគ្គ 2-h ចំនួន ៣ ដងដើម្បីទំលាប់ទៅនឹងប្រអប់ក្បាលរថភ្លើងបន្ទាប់មកមានវគ្គសាកល្បង AMPH ចំនួន ៦ ។ ថ្ងៃសាកល្បង AMPH បានកើតឡើងនៅចន្លោះពេល 1-wk ។ នៅថ្ងៃធ្វើតេស្តសត្វកណ្តុរត្រូវបានគេផ្តល់ឱ្យចំនួន 30 នាទីដើម្បីទំលាប់ទៅនឹងប្រអប់បន្ទាប់មកបានទទួលកំរិតថ្នាំអេសអិលអេចអិម ១០ មីលីក្រាម / គីឡូក្រាមក្នុងមួយថ្ងៃបន្ទាប់មកតាមវគ្គនីមួយៗដាច់ដោយឡែកពីគ្នាដោយចំនួនប្រាំដង 0.5 មីលីក្រាម / គីឡូក្រាម (មួយដូសក្នុងមួយថ្ងៃ) នៅថ្ងៃសាកល្បង 1.0 ដល់ 1 ។ ក្បាលរថយន្តក្រោយ AMPH ត្រូវបានគេវាយតម្លៃសម្រាប់រយៈពេល 5 នាទីលើវគ្គនីមួយៗ។
វិធីសាស្រ្តវិភាគទិន្នន័យ។
ការវិភាគស្ថិតិត្រូវបានធ្វើឡើងជាមួយអេសភីអេសអេស (អេស។ ស៊ី។ អេស។ ស។ ស៊ី។ ធី។ និងស។ ស៊ី។ អេស។ ស៊ី។ អេស។ ស៊ី។ អេស។ ស៊ី។ អាយ។ ស៊ី។ អាយ។ ) ។ ការឆ្លើយតបអាកប្បកិរិយាភ្លាមៗទៅកាន់ស៊ីអេសអេសត្រូវបានគេវាយតម្លៃទាក់ទងនឹងការច្រមុះតាមច្រមុះទៅក្នុងជំរៅដែលកាំរស្មីត្រូវបានគេចែកចាយ។ ចំនួនមធ្យមនៃផេសច្រមុះក្នុងចន្លោះពេលនេះ (16 s ក្នុងការសាកល្បង) បន្ទាប់មកត្រូវបានប្រៀបធៀបទៅនឹងចំនួនមធ្យមនៃកេសច្រមុះសម្រាប់រយៈពេលដូចគ្នា (21 s) ជាមធ្យមក្នុងរយៈពេលដែល CS អវត្តមាន។ ក្រុមអាយអូអូអេសអេសអេសអេសអេសអេសអេសអេសនិងអវត្តមានបានតាមដានការទទួលយកការឆ្លើយតបដែលរើសអើងចំពោះការឆ្លើយតបនិងការញាក់ច្រមុះដោយមិនរើសអើងក្រោមកាលវិភាគផ្សេងៗគ្នាក្នុងវគ្គបណ្តុះបណ្តាលស៊ីអ៊ិចអេសអិលអេចអរ។
ប្រសិទ្ធភាពនៃការព្យាបាលលើការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនត្រូវបានគេវាយតម្លៃជាមួយក្រុម Group អាយអេអូអេអាអេសសម្រាប់ដំណាក់កាលទំលាប់ដោយគ្រឿងញៀន (បីវគ្គ) ការរំញោចមុន 0.5 មីលីក្រាម / គីឡូក្រាមបញ្ហា AMPH (មួយវគ្គ) និងក្នុងអំឡុងពេលប្រាំវគ្គ 1 មីលីក្រាម / គីឡូក្រាម AMPH ។ តំរែតំរង់អារម្មណ៍នៅពេលក្រុមត្រូវបានគេរំពឹងថាខុសគ្នាក្នុងការឆ្លើយតបទៅនឹងកំរិតថ្នាំ AMPH ម្តងហើយម្តងទៀត។ ក្រុមអាយអូវអាយអេសអេសអេសអេសអេសអេសអេសអេសអេសអេសអេសអេសអេសក៏បានវាយតំលៃការឆ្លើយតបរបស់ម៉ូទ័រអាក់ទ័រដោយមិនប្រើថ្នាំក្នុងអំឡុងពេលនៃការចាក់ថ្នាំអេឡិចត្រូនិចដំណាក់កាលទី ១០ ។ ការប្រៀបធៀបដែលបានគ្រោងទុកបានវាយតម្លៃភាពខុសគ្នានៃលទ្ធផលមធ្យមសម្រាប់ក្រុមទី 30 ទល់នឹងក្រុម 50 (គ្មានការគ្រប់គ្រងរយៈពេលរំពឹងទុក) និងក្រុម 0 (គ្មានការគ្រប់គ្រងមិនច្បាស់លាស់) ដោយមធ្យោបាយ tការសាកល្បង (Howell, 1992), using the MS error and df error terms for the relevant effect (i.e., group or group × session interaction) from the ANOVA (Winer, 1971) ។ ការវិភាគនិន្នាការពហុវិជ្ជាបានសាកល្បងទម្រង់នៃការផ្លាស់ប្តូរក្នុងរយៈពេលនៃវគ្គ។
ដើម្បីកំណត់ថាតើវិធីសាស្រ្តឆ្លើយតបនៅក្នុងវត្តមាននិងអវត្តមាននៃស៊ីអេសក្នុងអំឡុងពេលវគ្គបណ្តុះបណ្តាល sucrose 15 បានរួមចំណែកដល់បំរែបំរួលនៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹង AMPH ឬភាពខុសគ្នានៃក្រុមសម្របសម្រួលនៅក្នុងការឆ្លើយតប AMPH ការវិភាគតាមដាននៃ covariance (ANCOVAs) ត្រូវបានអនុវត្តនៅលើ AMPH ។ ទិន្នន័យក្បាលម៉ាស៊ីនរួមទាំងភួយច្រមុះសរុប (ផលបូកសំរាប់វគ្គទី 15) នៅពេល CS អវត្តមានជា covariate ។ ផលប៉ះពាល់សំខាន់នៃ covariate នឹងបង្ហាញថាការឆ្លើយតបដោយមិនប្រើថ្នាំមានឥទ្ធិពល (ឥទ្ធិពលរបស់កម្លាំង) នៃក្រុមឬវគ្គ។ ការថយចុះនូវសារៈសំខាន់នៃផលប៉ះពាល់នៃក្រុមឬវគ្គនៅក្នុងវត្តមាននៃសម្ព័ន្ធភាពដ៏សំខាន់អាចបង្ហាញថាការឆ្លើយតបឆ្លើយតបទៅនឹងផលប៉ះពាល់របស់ក្រុមឬវគ្គ។ ការថយចុះនូវសារៈសំខាន់នៃក្រុមឬផលប៉ះពាល់វេននៅក្នុងអវត្តមាននៃឥទ្ធិពលវណ្ណៈអភិជនគឺគ្រាន់តែឆ្លុះបញ្ចាំងពីការបាត់បង់ថាមពលស្ថិតិដោយសារតែការផ្ទេរឌីអេហ្វអេពីពាក្យកំហុសទៅវណ្ណៈកម្មហើយនឹងមិនមានឥទ្ធិពលលើការបកស្រាយ ផលប៉ះពាល់នៃក្រុមឬវគ្គ។
លទ្ធផល
ច្រមុះហៀរក្នុងកំឡុងពេលវគ្គ sucrose ។
CS មានវត្តមាន។។ រូបភាព 1A បង្ហាញភេជ្ជៈច្រមុះមធ្យមសម្រាប់ក្រុមទី 25, 50 និង 100 ខណៈពេលដែល CS មានវត្តមាននៅលើវគ្គម៉ាស៊ីនត្រជាក់ 15 (ផែច្រមុះមិនត្រូវបានគេដាក់កូដសម្រាប់ក្រុម 0 ដែលមិនបានទទួល CS) ។ សម័យ 3 គ្រុប NUM 15 វគ្គ ANOVA បានផ្តល់នូវផលប៉ះពាល់សំខាន់ៗនៃក្រុម, F(2, 21) = 5.63, p = 0.011, និងសម័យ, F(14, 294) = 14.00, p <០.០០១ រួមជាមួយនឹងអន្តរកម្មជាក្រុម, សម័យ F(28, 294) = 2.93, p <០.០០១ ។ រូបភាព 1A បង្ហាញថាឥទ្ធិពលសំខាន់នៃសម័យបានឆ្លុះបញ្ចាំងពីការកើនឡើងនៃផែ្នកច្រមុះនៅទូទាំងវគ្គទាំងបីក្រុមហើយឥទ្ធិពលសំខាន់នៃក្រុមឆ្លុះបញ្ចាំងជាទូទៅពិន្ទុខ្ពស់ជាងជាទូទៅនៅក្នុងក្រុម 100 ទល់នឹងក្រុម 25 ដែលមានពិន្ទុមធ្យមនៅក្នុងក្រុម 50 ។ អន្តរកម្មជាក្រុម significant រយៈពេលដ៏សំខាន់សម្រាប់និន្នាការគូប។ F(2, 21) = 4.42, p = 0.030, ចង្អុលបង្ហាញពីការកើនឡើងយ៉ាងរហ័សការធ្លាក់ចុះនិងការធ្លាក់ចុះតាមច្រមុះនៅក្នុងវគ្គសិក្សានៅក្នុងក្រុម 100 ដូចជាការកើនឡើងលីនេអ៊ែរលើវគ្គនៅក្នុងក្រុម 50 និងការកើនឡើងលីនេអ៊ែររាក់ក្នុងរយៈពេលវគ្គនៅក្នុងក្រុម 25 ។
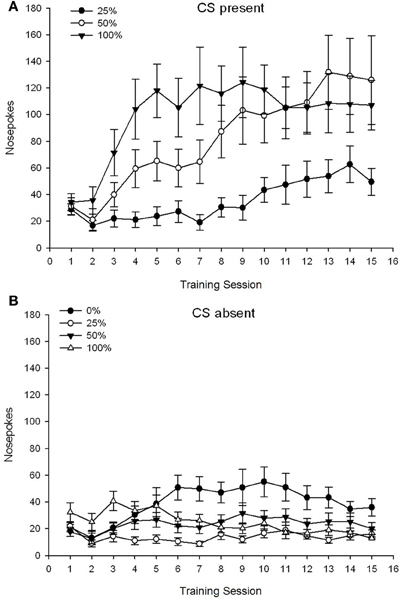
Figure 1. Mean (SE) approach responses (nose pokes) on 15 sucrose training sessions in groups of Sprague Dawley rats (n = 8 / ក្រុម) បានប៉ះពាល់នឹងរង្វាន់ sucrose (ដំណោះស្រាយ 10%) ដែលបានផ្តល់ជូននៅក្រោម 0, 25, 50, ឬកាលវិភាគអថេរ 100%. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. (ក) Scores when CS was present (5 s × 15 trials). (ខ) Scores when CS was absent (average for 5 × 15 s while light was off).
CS អវត្តមាន។។ រូបភាព 1B បង្ហាញមេដាយច្រមុះមធ្យមសម្រាប់ក្រុមទាំងបួនសម្រាប់រយៈពេលស្មើគ្នា (ការសាកល្បង 5 s s 15) ជាមធ្យមក្នុងរយៈពេលដែល CS អវត្តមាន។ សម័យ 4 គ្រុប NUM 15 វគ្គ ANOVA បានផ្តល់នូវផលប៉ះពាល់សំខាន់ៗនៃក្រុម, F(3, 28) = 7.06, p = 0.001, និងសម័យ។ F(14, 392) = 2.84, p <០.០០១ រួមជាមួយនឹងអន្តរកម្មជាក្រុម, សម័យ F(42, 392) = 3.93, p <០.០០១ ។ អន្តរកម្មជាក្រុម significant សម័យដ៏សំខាន់សម្រាប់និន្នាការបួនជ្រុង។ F(3, 28) = 3.91, p = 0.019 រួមជាមួយគ្មានអន្តរកម្មសម្រាប់និន្នាការគូប។ F(3, 28) <០០០០១, p > ០.៤៤ ឆ្លុះបញ្ចាំងពីទម្រង់នៃច្រមុះរបស់ច្រមុះដែលបញ្ចោញលើក្រុមនៅក្នុងក្រុម ០ ក៏ដូចជាប្រឆាំងនឹងទម្រង់ដែលមានស្ថេរភាពជាទូទៅលើវគ្គនៅក្នុងក្រុមផ្សេងទៀត។
ជំរកទៅបន្ទប់ក្បាលម៉ាស៊ីន។
ក្រុម 4 គ្រុប 3 សម័យ ANOVA បានផ្តល់នូវឥទ្ធិពលសំខាន់នៃសម័យប្រជុំ, F(2, 56) = 5.67, p = 0.006, និងគ្មានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត, F(3, 28) <០០០០១, p > ០.២១ ។ ការបំផ្លាញធ្នឹមមធ្យម (SE) ក្នុងមួយម៉ោង ២ នៅក្នុងប្រអប់ក្បាលម៉ាស៊ីនគឺ ១៦៨១១ (១២៣) នៅវគ្គទី ១ ១៥២៥ (១៤០) នៅវគ្គទី ២ និង ១២៦៩ (៩៦) លើវគ្គទី ៣ ។ ការប្រៀបធៀបដែលបានគ្រោងទុកមិនមានភាពខុសគ្នាខ្លាំងរវាងក្រុម ៥០ និងក្រុមឡើយ។ ០ ឬក្រុម ១០០ នៅវគ្គជំរកដំបូងឬចុងក្រោយ, t(84) <០០០០១, p > ០.០៥ ។ ដូច្នេះក្នុងករណីអវត្តមានរបស់អេមភីអេការប៉ះពាល់ម្តងហើយម្តងទៀតទៅនឹងប្រអប់តេស្តត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការថយចុះថេរនៃសកម្មភាពក្បាលរថភ្លើងដោយឯកឯងនៅក្នុងក្រុមទាំងបួន (ពោលគឺបែបផែនសម័យ) និងគ្មានការឆ្លើយតបឌីផេរ៉ង់ស្យែលដែលជាមុខងារនៃកាលវិភាគបណ្តុះបណ្តាល sucrose (គ្មានអន្តរកម្ម) ។
វគ្គសាកល្បង។
ប្រសិទ្ធភាពនៃការគិតមុន 0.5 មីលីក្រាម / គីឡូក្រាមបញ្ហា AMPH ។.
ក្បាលម៉ាស៊ីនចាក់មុន។។ អ័ក្សអ័រអេសអេសអេសស៊ីអេសអេសស៊ីអេសអេសនៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនក្នុងកំឡុងពេលនៃការចាក់ថ្នាំ 4 នាទីមិនមានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ទេ។ F(3, 28) <០០០០១, p > ០.៣៨ ។ ការប្រៀបធៀបដែលបានគ្រោងទុកមិនមានភាពខុសគ្នាខ្លាំងរវាងក្រុម ៥០ និងក្រុម ០ ឬក្រុម ១០០, t(32) <០០០០១, p > ០,៤០ ។ ដូច្នេះភាពខុសគ្នានៃមូលដ្ឋានគ្រឹះក្នុងក្បាលរថភ្លើងចាក់បញ្ចូលមុនមិនគិតពីភាពខុសគ្នានៃក្រុមនៅក្នុងការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹងអេមភីអេ។ ការបំបែកធ្នឹមមធ្យម (SE) សម្រាប់គំរូគឺ ៥៥៩ (៧៧) ។
ក្បាលម៉ាស៊ីនក្រោយចាក់ថ្នាំទល់នឹងវគ្គជម្រុះគ្រឿងញៀនចុងក្រោយ។។ ៤ ក្រុម× ២ វេន ANOVA បានប្រៀបធៀបការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនរបស់ក្រុមទាំងនោះនៅលើវគ្គជម្រុះចុងក្រោយហើយភ្លាមៗបន្ទាប់ពីការរំញោចមុន ០,៥ មីលីក្រាម / គីឡូក្រាមបញ្ហា AMPH ។ ពិន្ទុសម្រាប់វេននៃទំលាប់ (១២០ នាទី) ត្រូវបានគេធ្វើមាត្រដ្ឋានអោយត្រូវគ្នាទៅនឹងរយៈពេលនៃវគ្គសាកល្បងអេមភីអេស (៩០ នាទី) (ពិន្ទុទំលាប់ទំលាប់× ៩០/១២០) ។ ការវិភាគបានផ្តល់នូវឥទ្ធិពលសំខាន់នៃសម័យប្រជុំ F(1, 28) = 34.16, p <០.០០១ និងមិនមានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត F(3, 28) <០០០០១, p > ០.១០ ។ បែបផែនសម័យបានឆ្លុះបញ្ចាំងពីការកើនឡើងនៃការបំផ្លាញធ្នឹមមធ្យម (SE) ក្នុងការឆ្លើយតបទៅនឹងកំរិតប្រើចាប់ពី ៩៥២ (៧២) ដល់ ១៨៥៩ (១៥១) ។ ការប្រៀបធៀបដែលបានគ្រោងទុកមិនមានភាពខុសគ្នាខ្លាំងរវាងក្រុម ៥០ និងក្រុម ០ ឬក្រុម ១០០ ក្នុងការឆ្លើយតបទៅនឹងកំរិតថ្នាំនោះទេ។ t(56) <០០០០១, p > ០.១០ ។ ទោះយ៉ាងណាក៏ដោយលំដាប់ចំណាត់ថ្នាក់នៃពិន្ទុបំបែកធ្នឹម (M; SE) ស្របទៅនឹងសម្មតិកម្ម: ក្រុម ៥០ (២២០៥; ២៦៤)> ក្រុម ០ (២០២៥; ២០៣)> ក្រុម ១០០ (១៩០៩; ៤០៧)> ក្រុម ២៥ (១២៩៦; ២៩៩) ។
ផលប៉ះពាល់នៃ 1 មីលីក្រាម / គីឡូក្រាម AMPH ។.
ក្បាលម៉ាស៊ីនចាក់មុន។។ វគ្គ 4 គ្រុប× 5 វគ្គ ANOVA នៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនក្នុងកំឡុងពេលនៃការចាក់ថ្នាំស៊ីម៉ងត៍ដំណាក់កាលទី 30 នាទីនៅលើវគ្គសាកល្បង AMPH 1 មីលីក្រាម / គីឡូក្រាមផ្តល់នូវឥទ្ធិពលសំខាន់នៃសម័យ F(4, 112) = 43.64, p <០.០០១ និងមិនមានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត F(3, 28) <០០០០១, p > ០.៤២ ។ ការប្រៀបធៀបដែលបានគ្រោងទុកមិនមានភាពខុសគ្នាខ្លាំងរវាងក្រុម ៥០ និងក្រុម ០ ឬក្រុម ១០០ នៅវគ្គសាកល្បងដំបូងឬចុងក្រោយទេ។ t(140) <០០០០១, p > ០.៣០ ។ ដូច្នេះភាពខុសគ្នានៃមូលដ្ឋាននៅក្នុងក្បាលរថភ្លើងមិនរាប់បញ្ចូលភាពខុសគ្នានៃក្រុមនៅក្នុងការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹងអេមភីអេ។ ពិន្ទុបំបែកធ្នឹមមធ្យម (SE) សម្រាប់ដំណាក់កាលទំលាប់នៃកម្រិតដូសនៅវគ្គទី ១-៥ គឺៈ ៤៥៤ (៣០) ៨០៩ (៥៣) ៧៦០ (៣៦) ៥០៥ (៣៥) ៧៥៦ (៣៩) ។
ក្បាលរថភ្លើងក្រោយពេលចាក់។។ រូបភាព 2 បង្ហាញពីផលប៉ះពាល់នៃការចាក់ចំនួនប្រាំដងនៃ 1 mg / kg AMPH (មួយដងក្នុងមួយសប្តាហ៍) ទៅលើពិន្ទុសកម្មភាពក្បាលម៉ាស៊ីនក្នុងបួនក្រុម។ ក្រុម 4 គ្រុប 5 សម័យ ANOVA បានផ្តល់នូវឥទ្ធិពលសំខាន់នៃសម័យប្រជុំ, F(4, 112) = 8.21, p <០,០០១, ផលប៉ះពាល់សំខាន់នៃក្រុម, F(2, 45) = 3.28, p = 0.085, និងមិនមានអន្តរកម្មគួរឱ្យកត់សម្គាល់, F(12, 122) <០០០០១, p > ០.៦៨ ។
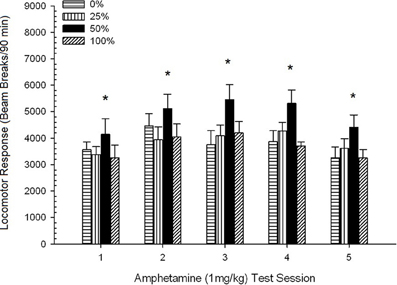
Figure 2. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) to 1 mg/kg d-amphetamine (i.p.) on 5 weekly sessions in groups of Sprague Dawley rats (n = 8/group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. *p < 0.05 for mean difference between group 50 and group 0 as well as group 100, based on planned comparisons.
ការប្រៀបធៀបដែលបានគ្រោងទុកបានបង្ហាញថាពិន្ទុក្រុម 50 ខុសគ្នាយ៉ាងខ្លាំងពីក្រុម 0, t(14) = 2.19, p = 0.037, និងក្រុម 100, t(14) = 2.36, p = 0.025 [និងខុសគ្នាបន្តិចបន្តួចពីក្រុម 25, t(14) = 2.03, p = 0.051] ។ ដូច្នេះនៅក្នុងក្រុម 50 ការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹង 1 មីលីក្រាម / គីឡូក្រាម AMPH អាចទុកចិត្តបានលើសពីក្រុមបីផ្សេងទៀតនៅទូទាំងវគ្គសាកល្បងទាំងប្រាំ។ ការវិភាគនិន្នាការពហុធាបានរកឃើញនិន្នាការជ្រុងបួនជ្រុងសំខាន់ៗនៅទូទាំងវគ្គនីមួយៗ F(1, 28) = 32.47, p <០,០០០១ និងគ្មាននិន្នាការសំខាន់ផ្សេងទៀត F(1, 28) <០០០០១, p > ០.១៩ ។ រូបភាព 2 បង្ហាញថាលទ្ធផលនេះបានឆ្លុះបញ្ចាំងពីលំនាំ“ បញ្ច្រាស់យូ” ឆ្លងកាត់វគ្គ។
គ្រប់គ្រងការផ្លាស់ប្តូរនៃការញ័រច្រមុះឆ្លើយតបក្នុងកំឡុងពេលហ្វឹកហាត់ sucrose ។
ការតាមដាន ANCOVA នៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹង 1 មីលីក្រាម / គីឡូក្រាម AMPH ដោយមានប្រហោងច្រមុះ (ស៊ីអេសអេស) ជា covariate ដែលស្ថិតក្នុងក្រុមទាំងបីដែលទទួលបានអេសអេសផ្តល់លទ្ធផលសំខាន់បន្ទាប់បន្សំនៃក្រុម។ F(2, 20) = 3.07, p = 0.069, និងមិនមានឥទ្ធិពលទាក់ទងនឹង covariate, F(4, 80) <០០០០១, p > ០.៨៥ ។ ដូច្នេះវិធីសាស្រ្តដែលត្រូវបានឆ្លើយតបក្នុងកំឡុងពេលហ្វឹកហាត់មិនបានពន្យល់ពីការប្រែប្រួលគួរឱ្យកត់សម្គាល់នៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹង AMPH 0.85 មីលីក្រាម / គីឡូក្រាមក្នុងក្រុម 1, 25, ឬ 50 ទេ។
ការតាមដាន ANCOVA នៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹង 1 មីលីក្រាម / គីឡូក្រាម AMPH ដោយមានប្រហោងច្រមុះ (CS អវត្តមាន) ជា covariate បានផ្តល់នូវផលប៉ះពាល់សំខាន់នៃ covariate, F(1, 27) = 6.17, p = 0.020, ផលប៉ះពាល់សំខាន់នៃក្រុម, F(3, 27) = 4.13, p = 0.016 វគ្គតូច interaction អន្តរកម្ម Covariate, p = 0.080, និងគ្មានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត, F(4, 108) <០០០០១, p > ០.២១ ។ ដូច្នេះវិធីសាស្រ្តដែលមិនមានការបដិសេធ (មិនរើសអើង) ក្នុងការឆ្លើយតបក្នុងកំឡុងពេលបណ្តុះបណ្តាលបានពន្យល់ពីការប្រែប្រួលគួរឱ្យកត់សម្គាល់នៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹងអេមភីអេស ១ មីលីក្រាម / គីឡូក្រាម។ ទោះជាយ៉ាងណាក៏ដោយបំរែបំរួលនេះមិនត្រូវបានត្រួតលើគ្នាជាមួយវ៉ារ្យង់ដែលទាក់ទងនឹងក្រុមនោះទេព្រោះការដាក់បញ្ចូលរបស់វ៉ាក់សាំងក្នុងការវិភាគបានកើនឡើងជាជាងការថយចុះសារៈសំខាន់នៃឥទ្ធិពលរបស់ក្រុម។
ការពិភាក្សា
ទិន្នន័យស្តីពីការញាក់ច្រមុះខណៈពេលដែល CS មានវត្តមានបង្ហាញថាក្រុមដែលទទួលបានការផ្សារភ្ជាប់គ្នារវាងស៊ីអេសនិងការចែកជូនតាមការឆ្លុះបញ្ចាំងពីការកើនឡើងនៃការឆ្លើយតបដែលត្រូវបានគេយកចិត្តទុកដាក់ជាងវគ្គបណ្តុះបណ្តាល។ ទម្រង់នៃការឆ្លើយតបលើវគ្គសិក្សានានាខណៈពេលដែលបច្ចុប្បន្ន CS បានបង្ហាញថាកាលវិភាគរបស់ 100 និង 50% ស៊ីអេសអេសមានប្រសិទ្ធភាពដូចគ្នាក្នុងវិធីដោះស្រាយរីឯកាលវិភាគ 25% បង្កើតបានជាការកើនឡើងតិចតួចនៃវិធីសាស្រ្តដែលបានបង្កើត។ ទិន្នន័យកណ្តាស់ច្រមុះខណៈពេលដែល CS អវត្តមានបានបង្ហាញថាក្រុមដែលបានទទួលកាលវិភាគបណ្តុះបណ្តាលស៊ីអេសអេសបី (ក្រុមទី 25, 50, 100) បានរៀនយ៉ាងឆាប់រហ័សដើម្បីកាត់បន្ថយកោរសក់ច្រមុះរបស់ពួកគេនៅពេលអវត្ដមាននៃ CS ចំណែកសត្វនៅក្នុងក្រុម 0 ។ ដែលមិនបានទទួលស៊ីអេសគ្រាន់តែរៀនកាត់បន្ថយអាកប្បកិរិយារបស់ពួកគេឱ្យនៅកំរិតមួយបន្ទាប់ពីការបណ្តុះបណ្តាលយ៉ាងទូលំទូលាយ។
ទិន្នន័យនៃទំលាប់បង្ហាញថាក្រុមទាំងនោះមិនមានភាពខុសប្លែកគ្នាមុនពេល AMPH និងការប៉ះពាល់ម្តងហើយម្តងទៀតទៅនឹងប្រអប់តេស្តត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការថយចុះនៃការឆ្លើយតបរបស់ឧបករណ៍បន្ថយក្បាលម៉ាស៊ីន។ ដូច្នេះភាពខុសគ្នារវាងក្រុមនិងការឆ្លើយតបកើនឡើងចំពោះការប្រើម្តងហើយម្តងទៀតនៃអេអឹមអេចមិនអាចត្រូវបានគេសន្មតថាមានភាពខុសគ្នាពីមុននៃឥរិយាបថក្បាលម៉ាស៊ីន។
លទ្ធផលនៃការប្រកួតប្រជែងមុនពេលដឹងច្បាស់ជាមួយ 0.5 មីលីក្រាម / គីឡូក្រាម AMPH បានបញ្ជាក់ថាថ្នាំនេះបង្កើនសកម្មភាពក្បាលរថភ្លើងទាក់ទងនឹងថ្ងៃឈប់ជក់គ្រឿងញៀនចុងក្រោយ។ ស្របតាមសម្មតិកម្មក្រុម 50 ចំណាត់ថ្នាក់ខ្ពស់ជាងក្រុម 0 ឬ 100 (ក៏ដូចជាក្រុម 25) ទាក់ទងនឹងការឆ្លើយតបមធ្យមចំពោះកំរិតទោះបីជាភាពខុសគ្នារវាងក្រុមមិនសំខាន់ក៏ដោយ។
សម្រាប់វគ្គប្រែប្រួលការប្រៀបធៀបដែលបានគ្រោងទុករវាងក្រុមបានបង្ហាញថាការប៉ះពាល់មុននៃរង្វាន់រង្វាន់ ៥០ ភាគរយនាំឱ្យមានការកើនឡើងយ៉ាងខ្លាំងនៃការឆ្លើយតបនៃក្បាលរថភ្លើងទៅនឹងកម្រិតថ្នាំអាហ្វេតាមីនចំនួន ១,០ មីលីក្រាម / គីឡូក្រាមទាក់ទងនឹងកាលវិភាគចំនួន ៣ ផ្សេងទៀត។ ប្រសិទ្ធិភាពនេះត្រូវបានបង្ហាញឱ្យឃើញពីកំរិតដំបូងហើយមិនបានផ្លាស់ប្តូរការចង់បានលើសពីកំរិតម្តងហើយម្តងទៀត។ ការវិភាគនិន្នាការបានបង្ហាញពីការឆ្លើយតបជាទ្វេភាគី (សម្រាប់គំរូពេញ) ទៅនឹងកំរិតថ្នាំ AMPH ម្តងហើយម្តងទៀតដែលកើនឡើងដល់កំរិតទីបីនិងថយចុះ។ លទ្ធផលនៃការតាមដាន ANCOVA ដោយប្រើច្រមុះច្រមុះ (អវត្តមាន CS) ខណៈដែល covariate បានបញ្ជាក់ថាភាពខុសគ្នានៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនរបស់ក្រុមទាំងបួនទៅ ១ មីលីក្រាម / គីឡូក្រាម AMPH មិនត្រូវបានសំរបសំរួលដោយវិធីសាស្រ្តដែលមិនមានការយកចិត្តទុកដាក់ក្នុងកំឡុងពេលហ្វឹកហាត់បឺត។
ផលប៉ះពាល់ជាក្រុមក្នុងអំឡុងពេលនៃការចាប់អារម្មណ៍គឺស្របនឹងសម្មតិកម្មរបស់យើង។ បែបផែនសម័យឆមាសមិនត្រូវគ្នានឹងការកើនឡើងជាលំដាប់ដែលរំពឹងទុកនៅក្នុងការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនជាមួយនឹងការចាក់ថ្នាំ AMPH ម្តងហើយម្តងទៀត។ នេះអាចទាក់ទងនឹងចន្លោះពេលនៃការប្រើថ្នាំ។ ដើម្បីដោះស្រាយបញ្ហានេះនីតិវិធីមួយ (ដូសប្រចាំថ្ងៃជំនួស) បានបង្ហាញដើម្បីជំរុញឱ្យមានការកើនឡើងជាលំដាប់នៅក្នុងការឆ្លើយតបនៃក្បាលម៉ាស៊ីនទៅនឹង 1.0 មីលីក្រាម / គីឡូក្រាមនៃអេមភីអេស (មានន័យថាការផ្លាស់ប្តូរអាកប្បកិរិយា) គួរតែត្រូវបានប្រើ។ ផលប៉ះពាល់នៃរបបដែលរំញោចនៃអេអឹមអេជទៅលើការឆ្លើយតបជាបន្តបន្ទាប់ចំពោះបញ្ហាប្រឈម 0.5 មីលីក្រាម / គីឡូក្រាមនឹងជួយគាំទ្រដល់ភាពទូទៅនៃឥទ្ធិពលនេះ។ ការដាក់បញ្ចូលបញ្ហាប្រឈមនឹងជាតិប្រៃមុនពេល AMPH នឹងកំណត់តួនាទីនៃផលប៉ះពាល់ការរំពឹងទុកឬការចាក់ (ឧទាហរណ៍ស្ត្រេស) ទៅលើការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅអេអឹមអេជ។ ការដាក់បញ្ចូលក្រុម sucrose ដែលមានលក្ខខណ្ឌចំនួន 75% នឹងជួយបញ្ជាក់ពីតួនាទីនៃភាពមិនប្រាកដប្រជានៃរង្វាន់និងភាពមិនប្រាកដប្រជានៃរង្វាន់ចំពោះគំរូនៃការឆ្លើយតបសម្រាប់ក្រុម 50 និងក្រុម 25 ។ លើសពីនេះទៀតដើម្បីអនុញ្ញាតឱ្យមានការវាយតម្លៃ (ដោយ ANCOVA) នៃការចូលរួមចំណែកនៃវិធីសាស្រ្តដោះស្រាយដោយគ្មានគ្រឿងញៀនចំពោះចលនាក្បាលម៉ាស៊ីនក្រោមអេអឹមអេជ (ប្រើផតខាស់ច្រមុះជាមួយស៊ីអេសអេសជា covariate) ផែ្នកច្រមុះក៏ត្រូវបានគេសរសេរកូដសម្រាប់ក្រុម 0 ក្នុងចន្លោះពេលនៅពេល CS មានវត្តមាននៅក្នុងក្រុមចំនួនបួនផ្សេងទៀត (មានន័យថាច្រមុះហៀរចេញពីក្រុមទាំងប្រាំ - រួមទាំងក្រុម 0 ដែលមិនបានទទួលស៊ីអេសអេស - អាចត្រូវបានរាប់បញ្ចូលក្នុងការវិភាគនៃ covariance ជាមួយស៊ីអេសអេលដែលជាសម្ព័ន្ធមិត្ត) ។ ការកែលម្អទាំងនេះត្រូវបានដាក់បញ្ចូលក្នុងការពិសោធន៍ 2 ។
ពិសោធន៍ 2
សំភារៈនិងវិធីសាស្រ្ត
The methodology of experiment 2 was similar to that of experiment 1 but revised to better approximate a regimen found to reliably induce AMPH sensitization (Fletcher et al ។ , 2005) ។ ការផ្លាស់ប្តូរមានដូចខាងក្រោមៈ (ក) ក្រុម 75% CS-sucrose (n = 8) ត្រូវបានរួមបញ្ចូល; (ខ) ក្នុងអំឡុងពេលហ្វឹកហាត់ sucrose កណ្តុរ (លើកលែងតែក្រុម 0) បានទទួលបទបង្ហាញ 20 CS (ពន្លឺ) (ផ្ទុយពី 15 ក្នុងការពិសោធន៍ 1); (c) បទបង្ហាញ CS ត្រូវបានបំបែកគ្នាដោយចន្លោះពេលនៃការសាកល្បងជាមធ្យមនៃ 90 s; ជួរ: 30 – 180 s (ទល់នឹង 120 s នៅក្នុងការពិសោធន៍ 1) ដែលទូទាត់នឹងការកើនឡើងនៃការសាកល្បងហ្វឹកហាត់ដើម្បីស្មើនឹងរយៈពេលនៃវគ្គបណ្តុះបណ្តាលនីមួយៗទៅនឹងការពិសោធ 1; (ឃ) រយៈពេលនៃវគ្គទំលាប់ទាំងបីត្រូវបានថយចុះពី 120 ដល់ 90 នាទីដើម្បីឆ្លើយតបទៅនឹងរយៈពេលនៃវគ្គសាកល្បង; (ង) បញ្ហាប្រឈមនឹងទឹកអំបិល (ip, 1 មីលីលី / គីឡូក្រាម) (90 នាទី) ត្រូវបានបន្ថែម (ថ្ងៃបណ្តុះបណ្តាលក្រោយ sucrose 8) ដើម្បីវាយតម្លៃផលប៉ះពាល់ក្បាលម៉ាស៊ីននៃការចាក់។ ក្នុងមួយ SE (ឧទាហរណ៍ការរំពឹងទុកភាពតានតឹង); (ច) វគ្គរំញោចអារម្មណ៍ 1 មីលីក្រាម / គីឡូក្រាមត្រូវបានធ្វើឡើងនៅថ្ងៃធ្វើការឆ្លាស់គ្នា (ថ្ងៃហ្វឹកហាត់ក្រោយ 12 – 21) ជាជាងនៅចន្លោះពេលប្រចាំសប្តាហ៍ដូចនៅក្នុងការពិសោធន៍ 1; (ក្រាម) រួមជាមួយនឹងការត្រៀមលក្ខណៈអេមអេស៊ាអិមអេសអេជអេជអេជអេជអេលអេសអេជអេលអេសអេសអេសអេសអេសអេសអេសអេសអេសអេសអេសអេសអេសអេលអេសអេជអេ (អេ។ អេ។ អេ។ អេ។ អេ។ អេ។ អេ។ អេ។ ប្រសិទ្ធិភាពភាពរំជើបរំជួលនៅទូទាំងកិតើ; (h) ផែ្នកច្រមុះខណៈពេលដែល CS បច្ចុប្បន្នត្រូវបានដាក់កូដសម្រាប់ក្រុមទាំងអស់ (រួមទាំងក្រុម 0.5); (ខ្ញុំ) ផែ្នកច្រមុះខណៈពេលដែល CS អវត្តមានត្រូវបានគេកត់ត្រាជាពិសេសពីចន្លោះពេល 9-s ភ្លាមៗមុនពេលចាប់ផ្តើមនៃអេសអេសដើម្បីចង្អុលបង្ហាញវិធីសាស្រ្តឆ្លើយតបមិនគ្រប់ខែ។
លទ្ធផល
ច្រមុះហៀរក្នុងកំឡុងពេលវគ្គ sucrose ។
ដំណាក់កាលទី 5 ក្រុម×វគ្គទី 15 វគ្គ× 2 ដំណាក់កាល (បច្ចុប្បន្ន CS, អវត្តមាន CS) ANOVA នៃច្រមុះច្រមុះផ្តល់នូវផលប៉ះពាល់សំខាន់ៗនៃក្រុម, F(4, 19) = 2.89, p = 0.050, សម័យ។ F(14, 266) = 2.28, p = 0.006, និងតំណាក់កាល, F(1, 19) = 14.72, p = 0.001 ក៏ដូចជាអន្តរកម្មបីផ្លូវយ៉ាងសំខាន់។ F(56, 266) = 1.38, p = 0.050 ។ បន្ទះ។ (A, B) នៃរូប។ 3 រៀបចំផែនការដាក់ពិន្ទុច្រមុះសម្រាប់ច្រមុះសម្រាប់ក្រុមបច្ចុប្បន្ននិង CS អវត្តមានដំណាក់កាលរៀងគ្នា។ ការប្រៀបធៀបនៃបន្ទះទាំងពីរបង្ហាញថាឥទ្ធិពលសំខាន់នៃដំណាក់កាលបានឆ្លុះបញ្ចាំងពីការឆ្លើយតបនៃការស្ទុះច្រមុះជាទូទៅនៅពេលដែល CS មានវត្តមានទល់នឹងអវត្តមាន។ ហេតុដូច្នេះហើយការឆ្លើយតបដោយយកចិត្តទុកដាក់បានកើតឡើងជាញឹកញាប់ជាងការឆ្លើយតបដែលមិនធ្លាប់មានពីមុន។ ផលប៉ះពាល់សំខាន់ៗនៃក្រុមនិងសម័យមិនត្រូវបានបកស្រាយងាយទេដោយសារអន្តរកម្មលំដាប់ខ្ពស់។ លទ្ធផលចុងក្រោយនេះបានឆ្លុះបញ្ចាំងពីចំណុចរួមបញ្ចូលគ្នាសម្រាប់ក្រុមទាំង ៥ នៅកម្រិតទាបមួយដែលមានស្ថេរភាពឆ្លងកាត់វគ្គជាច្រើននៅពេលដែលអ 3B) រួមជាមួយនឹងការបង្វែរពិន្ទុទៅជាខ្ពស់ (ក្រុម 75 ក្រុម 100) កម្រិតមធ្យម (ក្រុម 50) និងកំរិតទាប (ក្រុម 0, ក្រុមទី 25) នៃច្រមុះហៀរសំបោរឆ្លើយតបនៅវគ្គជាច្រើននៅពេលដែល CS មានវត្តមាន (រូបភាពទី 3A) ។ ក្នុងចំណោមនិន្នាការពហុធាដែលមានលំដាប់ទាប (លីនែអ៊ែនធ័របួនជ្រុងគូប) មានតែអន្តរកម្មបីផ្លូវសម្រាប់និន្នាការលីនេអ៊ែរឈានដល់សារៈសំខាន់។ F(4, 19) = 2.32, p = 0.094, ឆ្លុះបញ្ចាំងពីការកើនឡើងជាទូទៅនៃការលេងល្បែងតាមច្រមុះលើវគ្គនៅក្នុងក្រុម 75 និងស្ថេរភាពគួរឱ្យកត់សម្គាល់លឿនជាងមុននៅកម្រិតខ្ពស់, មធ្យមនិងកម្រិតទាបនៃការឆ្លើយតបនៅក្នុងក្រុមដទៃទៀតនៅពេល CS មានវត្តមាន។
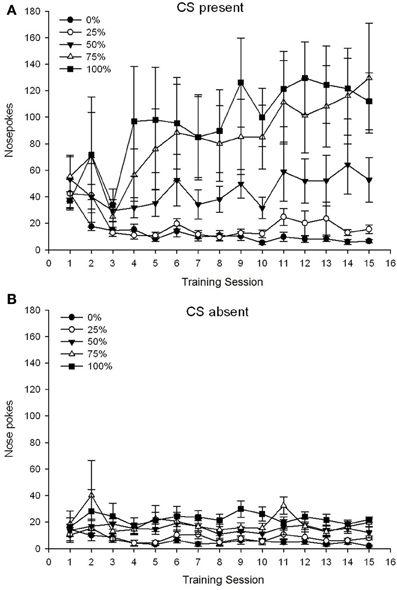
Figure 3. Mean (SE) approach responses (nose pokes) on 15 sucrose training sessions in groups of Sprague Dawley rats (n = 8 / ក្រុម) ប្រឈមនឹងរង្វាន់ sucrose (ដំណោះស្រាយ 10%) ដែលបានផ្តល់ជូននៅក្រោម 0, 25, 50, 75, ឬកាលវិភាគអថេរ 100%. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. (ក) Scores when CS was present (5 s × 20 trials). (ខ) Scores when CS was absent (average for 5 × 20 s while light was off).
ជំរកទៅនឹងប្រអប់ក្បាលម៉ាស៊ីន។
ការឆ្លើយតបនឹងក្បាលរថភ្លើងគ្មានគ្រឿងញៀន 5 Group NUM 3 សម័យ ANOVA ផ្តល់នូវឥទ្ធិពលសំខាន់នៃសម័យប្រជុំ។ F(2, 70) = 60.01, p <០.០០១ និងមិនមានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត F(4, 35) <០០០០១, p > ០.៦០ ។ ការប្រៀបធៀបដែលបានគ្រោងទុកនៃក្រុម ៥០ ជាមួយក្រុម ០ និងជាមួយក្រុម ១០០ នៅវគ្គទីជំរកទីមួយនិងចុងក្រោយមិនមានផលប៉ះពាល់អ្វីគួរឱ្យកត់សម្គាល់ទេ។ t'<០.៨៤, p > ០,៤០ ។ ដូច្នេះមានន័យថាការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនដែលគ្មានគ្រឿងញៀននៅក្នុងក្រុមសំខាន់ៗមិនខុសគ្នាទេមុនពេលធ្វើតេស្ត។ ចំនួននៃការបំបែកធ្នឹមក្នុង ៩០ នាទីគឺ ២១៦២ (១១៨) នៅវគ្គទី ១, ១៤៧០ (១១៦) នៅវគ្គទី ២ និង ១២៥០ (៩៨) នៅវគ្គទី ៣ ។
វគ្គសាកល្បង។
Saline។ 5 Group × 2 Session ANOVA បានប្រៀបធៀបការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីននៅលើវគ្គជម្រុះចុងក្រោយនិងវគ្គប្រឈមនឹងជាតិប្រៃ។ ANOVA ផ្តល់លទ្ធផលសំខាន់នៃសម័យប្រជុំ F(1, 35) = 62.46, p <០.០០១ និងមិនមានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត F(4, 35) <០០០០១, p > ០.១៩ ។ រូបភាព 4 ផែនការក្រុមមានន័យថានិងបង្ហាញថាផលប៉ះពាល់វេនឆ្លុះបញ្ចាំងពីការថយចុះជាទូទៅនៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនពីវេនជក់គ្រឿងញៀនចុងក្រោយដល់វគ្គអំបិលដែលមិនខុសគ្នាតាមក្រុម។ ដូច្នេះការធ្លាក់ចុះនៃការឆ្លើយតបនឹងក្បាលរថភ្លើងដែលត្រូវបានគេមើលឃើញក្នុងរយៈពេលវគ្គជម្រុះចំនួនបីបានបន្តនៅលើការប៉ះពាល់គ្រឿងញៀនទីបួនទៅនឹងប្រអប់សាកល្បង។
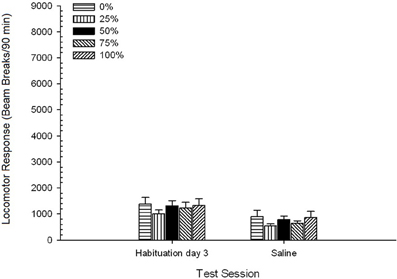
Figure 4. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) on the last of 3 drug-free habituation sessions and on a subsequent session after saline injection (i.p., 1 ml/kg) in groups of Sprague Dawley rats (n = 8/group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, 75, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli.
ផលប៉ះពាល់នៃ 0.5 មីលីក្រាម / គីឡូក្រាម AMPH ។.
ក្បាលម៉ាស៊ីនចាក់មុន។។ 5 Group × 2 Session ANOVA នៃក្បាលរថភ្លើងចាក់មុន (30- នាទី) នៅមុនការធ្វើតេស្ត 0.5 mg / kg ថ្ងៃសាកល្បង AMPH បានផ្តល់នូវឥទ្ធិពលសំខាន់នៃសម័យប្រជុំ, F(1, 35) = 13.39, p = 0.001, និងគ្មានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត, F(4, 35) <០០០០១, p > ០.១៥ ។ ការប្រៀបធៀបដែលបានគ្រោងទុកមិនមានភាពខុសគ្នាខ្លាំងរវាងក្រុម ៥០ និងក្រុម ០ ឬក្រុម ១០០ នៅវគ្គទី ១ ទេ។ t(70) <០០០០១, p > ០.៣០ ។ ទោះយ៉ាងណានៅលើកទី ២ (ក្រោយការចាប់អារម្មណ៍) ក្រុមវេនទី ៥០ (១២០៣; ១២១) បានបង្ហាញពីការបំបែកធ្នឹមចាក់ថ្នាំ (M; SE) ច្រើនជាងក្រុមទី ១០០ (៧៥៦; ១០៣) ។ t(70) = 5.11, p <០.០០១ ប៉ុន្តែមិនខុសគ្នាពីក្រុម ០ (១១២៦; ២១១), t(7) <០០០០១, p > ០,៤០ ។ ដូច្នេះភាពខុសគ្នានៃមូលដ្ឋាននៅក្នុងក្បាលរថភ្លើងមិនរាប់បញ្ចូលភាពខុសគ្នានៃក្រុមក្នុងការឆ្លើយតបនឹងក្បាលរថភ្លើងចំពោះកម្រិតថ្នាំ AMPH ០.៥ មីលីក្រាម / គីឡូក្រាមដំបូងទេប៉ុន្តែអាចរួមចំណែកដល់ភាពខុសគ្នារវាងក្រុម ៥០ និងក្រុម ១០០ ក្នុងការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹងកម្រិតថ្នាំ AMPH ០.៥ មីលីក្រាម / គីឡូក្រាម។ ។ ការបំបែកធ្នឹមមធ្យម (SE) សម្រាប់ដំណាក់កាលចាក់មុននៅវគ្គទី ១ នៃ AMPH ០,៥ មីលីក្រាម / គីឡូក្រាមគឺ ៧៥៧ (៤១) និង ៩៧៤ (៥៩) ។
ក្បាលរថភ្លើងក្រោយពេលចាក់។។ ក្រុម 5 ក្រុម NUM 2 សម័យ ANOVA នៃក្បាលម៉ាស៊ីនក្បាលម៉ាស៊ីនឆ្លើយតបទៅនឹង 0.5 មីលីក្រាម / គីឡូក្រាម AMPH មុននិងក្រោយតំរែតំរង់តំរែតំរង់ 5 បានផ្តល់នូវឥទ្ធិពលសំខាន់នៃសម័យ។ F(1, 35) = 76.05, p <០.០០១ និងមិនមានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត F(4, 35) <០០០០១, p > ០.១៩ ។ រូបភាព 5 បង្ហាញពិន្ទុមធ្យមសម្រាប់ក្រុមនិងវគ្គនីមួយៗ។
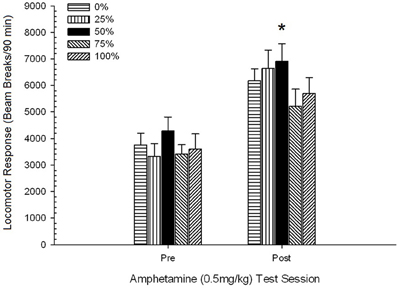
Figure 5. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) to 0.5 mg/kg d-amphetamine on separate sessions before and after a 5-session sensitizing regimen of d-amphetamine (1.0 mg/kg; i.p. per session) in groups of Sprague Dawley rats (n = 8/group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, 75, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. *p < 0.05 for mean difference between group 50 and group 0 as well as group 100, based on planned comparisons.
តួលេខបង្ហាញថាផលប៉ះពាល់ Session ពាក់ព័ន្ធនឹងការកើនឡើងគួរឱ្យកត់សម្គាល់នៃការបំបែកធ្នឹមមធ្យម (SE) ក្នុងមួយ 90 នាទីពី 0.5 មីលីក្រាម / កិត 1, 3674 (216) ដល់ 0.5 មីលីក្រាម / កិតើកិតើ 2, 6123 (275) ។ កង្វះនៃអន្តរកម្មឬក្រុមមានប្រសិទ្ធិភាពបានបង្ហាញថាការប្រែលប្រួល AMPH មិនមានភាពខុសគ្នារវាងក្រុមនីមួយៗទេ។ ទោះបីជាកង្វះនៃផលប៉ះពាល់ទាក់ទងនឹងក្រុមសំខាន់ៗនៅក្នុង ANOVA ក៏ដោយក៏ការត្រួតពិនិត្យតួលេខបង្ហាញថាក្រុម 50 បង្ហាញនូវការឆ្លើយតបដ៏អស្ចារ្យបំផុតចំពោះកំរិតថ្នាំ 0.5 mg / គីឡូក្រាមចំនួន ១ និង ២ ។ ការប្រៀបធៀបដែលបានគ្រោងទុកនៃការឆ្លើយតបទៅនឹងកំរិតថ្នាំ 0.5 មីលីក្រាម / គីឡូក្រាមដំបូងបានបង្ហាញថាមិនមានភាពខុសគ្នាខ្លាំងរវាងក្រុម 50 និងក្រុម 0 ឬក្រុម 100, t's(35) <០០០០១, p > ០.៥០ ។ ទោះយ៉ាងណាក៏ដោយដើម្បីឆ្លើយតបទៅនឹងការចាក់ថ្នាំ ០,៥ មីលីក្រាម / គីឡូក្រាមលើកទី ២ ក្រុមទី ៥០ បង្ហាញក្បាលរថភ្លើងធំជាងក្រុម ០, t(35) = 2.00, p <0.05 ក៏ដូចជាក្រុម ១០០, t(35) = 3.29, p <0.01 ។
ដោយមើលឃើញពីភាពខុសគ្នានៃក្រុមសំខាន់ក្នុងក្បាលរថភ្លើងចាក់នៅលើកទី ២ នៃ 0.5 mg / kg AMPH ដែលបានរាយការណ៍ខាងលើការតាមដាន 5 Group × 2 Session ANCOVA នៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹង 0.5 mg / kg AMPH ត្រូវបានធ្វើឡើងដែលគ្រប់គ្រងមុន ចាក់ក្បាលរថភ្លើងនៅវគ្គទី ២ ។ ការវិភាគនេះបានផ្តល់លទ្ធផលគួរឱ្យកត់សំគាល់នៃ covariate, F(1, 34) = 8.65, p = 0.006, ឥទ្ធិពលសំខាន់នៃសម័យ។ F(1, 34) = 10.83, p = 0.002, និងគ្មានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត, F(4, 34) <០០០០១, p > ០.៥០ ។ សំខាន់ការប្រៀបធៀបដែលបានគ្រោងទុកដោយផ្អែកលើកំហុស MS និង df ពី ANCOVA បានបញ្ជាក់ថាការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹងដូសអេមភីអេ ០,៥ មីលីក្រាម / គីឡូក្រាមលើកទី ២ នៅតែមានចំនួនច្រើនជាងក្រុម ៥០ ជាងក្រុម ១០០ t(34) = 3.09, p <0.01, និងក្រុម 0, t(34) = 1.88, p <០.០៥ (កន្ទុយមួយ) នៅពេលបំរែបំរួលមុនចាក់ពីវគ្គ ២ ត្រូវបានគ្រប់គ្រង។ ដូច្នេះក្រុមទី ៥០ បានបង្ហាញក្បាលម៉ាស៊ីនក្រោយការចាប់អារម្មណ៏កាន់តែខ្លាំងទៅនឹងអេអឹមភី ០.៥ មីលីក្រាម / គីឡូក្រាមច្រើនជាងក្រុម ១០០ ឬក្រុម ០ ហើយភាពខុសគ្នានៃក្រុមទាំងនេះមិនត្រូវបានសម្របសម្រួលដោយក្បាលរថភ្លើងចាក់មុនថ្ងៃសាកល្បងឡើយ។
ផលប៉ះពាល់នៃ 1.0 មីលីក្រាម / គីឡូក្រាម AMPH ។.
ក្បាលម៉ាស៊ីនចាក់មុន។។ វគ្គចាក់បញ្ចូល 5 គ្រុប× 5 វគ្គ ANOVA នៃ 30 នាទីមុនសម្រាប់ការទទួលអារម្មណ៍នៃអរម៉ូនអេសអេជអេជអិមអេជអរអេសអិលអេជអរអេសបានទទួលឥទ្ធិពលសំខាន់នៃសម័យ។ F(4, 140) = 16.70, p <០.០០១ និងមិនមានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត F(4, 35) <០០០០១, p > ០.៤៥ ។ ការប្រៀបធៀបដែលបានគ្រោងទុកមិនមានភាពខុសប្លែកគ្នាគួរឱ្យកត់សម្គាល់នៅក្នុងក្បាលម៉ាស៊ីនចាក់មុនរវាងក្រុម ៥០ និងក្រុម ០ ឬក្រុម ១០០ នៅវគ្គទី ១ ទេ។ t(175) <០០០០១, p > ០.១០ ។ ទោះយ៉ាងណានៅវគ្គចុងក្រោយក្រុម ៥០ (១១៦៧; ១៤០) បង្ហាញការបំបែកធ្នឹម (M; SE) ច្រើនជាងក្រុម ១០០ (១០០០; ៩៩), t(175) = 2.35, p <០.០៥ ប៉ុន្តែមិនខុសគ្នាពីក្រុម ០ (១០៨៥, ១២០), t(175) <០០០០១, p > ០.២០ ។ ដូច្នេះភាពខុសគ្នានៃក្បាលរថភ្លើងចាក់បញ្ចូលគ្នាធ្វើឱ្យមានភាពខុសគ្នារវាងក្រុម 0.20 និង 50 ក្នុងការឆ្លើយតបនឹងក្បាលរថភ្លើងទៅនឹងកម្រិតថ្នាំ AMPH 100 មីលីក្រាម / គីឡូក្រាមចុងក្រោយ។ មធ្យម (SE) ការបំបែកធ្នឹមសរុបសម្រាប់គំរូក្នុងដំណាក់កាលចាក់មុនសម្រាប់វគ្គទី ១ ដល់ ៥ គឺ ៨១០ (៤៦) ៧៨៤ (៥២) ៧៦០ (៥៣) ៧២៦ (៤៦) ១០០៩ (៥១) ។
ក្បាលរថភ្លើងក្រោយពេលចាក់។។ ក្រុម 5 ក្រុម× 5 សម័យ ANOVA នៃការឆ្លើយតបចំពោះ 1 មីលីក្រាម / គីឡូក្រាមអេអឹមអេផ្តល់ផលសំខាន់យ៉ាងសំខាន់នៃសម័យ។ F(4, 140) = 6.72, p <០.០០១, ក្រុមតូច interaction អន្តរកម្មវេន, F(16, 140) = 1.57, p = 0.085, និងគ្មានផលប៉ះពាល់សំខាន់នៃក្រុម, F(4, 35) <០០០០១, p > ០.៧៧ ។ ការវិភាគនិន្នាការពហុវិជ្ជាបានបង្ហាញពីនិន្នាការលីនេអ៊ែរគួរឱ្យកត់សម្គាល់ F(1, 35) = 9.19, p = 0.005 និងនិន្នាការគូប F(1, 35) = 21.63, p <០.០០១, លើវគ្គ ១ ដល់ ៥ 6 បង្ហាញពិន្ទុក្បាលរថភ្លើងមធ្យមសម្រាប់ក្រុមនិងវគ្គនីមួយៗ។
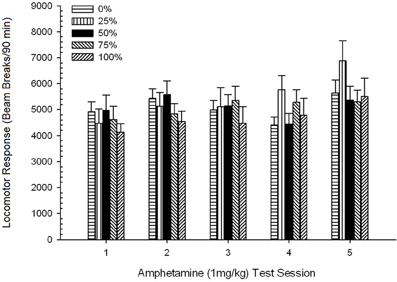
Figure 6. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) to 1 mg/kg d-amphetamine (i.p.) on 5 weekly sessions in groups of Sprague Dawley rats (n = 8/ group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, 75, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli.
តួលេខបង្ហាញថាផលប៉ះពាល់សម័យបានឆ្លុះបញ្ចាំងពីការកើនឡើងគួរឱ្យកត់សម្គាល់នៃការបំបែកធ្នឹមមធ្យម (SE) សម្រាប់គំរូពេញលេញពីវគ្គទី 1, 4624 (213) ដល់វគ្គទី 5, 5736 (272) ដែលបញ្ជាក់ពីការកើតឡើងនៃអារម្មណ៍ទៅ AMPH ។ និន្នាការគូបបង្ហាញពីអតិបរិមាទាក់ទងនឹងវគ្គទី 1, 3, និង 5, ជាមួយនឹងការធ្លាក់ចុះនៅលើវគ្គ 2 និង 4 ជាពិសេសសម្រាប់ក្រុម 0 និង 50 ។ តួលេខនេះក៏បានបង្ហាញផងដែរថាទោះបីជាកង្វះនៃអន្តរកម្មសំខាន់ក្រុម 25 បង្ហាញការឆ្លើយតបក្បាលម៉ាស៊ីនកាន់តែច្រើនឡើង ៗ នៅលើវគ្គហើយខុសគ្នាគួរឱ្យកត់សម្គាល់ពីក្រុមដទៃទៀតនៅលើវគ្គទី 4 និង 5 (9 និង 22% ធំជាងក្រុមខ្ពស់បន្ទាប់) ។ ការប្រៀបធៀបដែលបានគ្រោងទុកបានរកឃើញថាក្រុមទី 50 មិនខុសគ្នាខ្លាំងពីក្រុម 0 ឬ 100, t(175) <០០០០១, p > ០,៤០ លើវគ្គតេស្ត AMPH ១ ឬ ១ មីលីក្រាម / ចុងក្រោយ។
គ្រប់គ្រងការផ្លាស់ប្តូរនៃការញ័រច្រមុះឆ្លើយតបក្នុងកំឡុងពេលហ្វឹកហាត់ sucrose ។
ក្រុម 5 ពីរក្រុម NUM 2 វគ្គ ANCOVA នៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹង 0.5 មីលីក្រាម / គីឡូក្រាម AMPH មុននិងក្រោយតំរែតំរង់អារម្មណ៍រួមទាំងផែ្នកច្រមុះសរុបក្នុងកំឡុងពេលហ្វឹកហាត់ sucrose ជាមួយ CS បច្ចុប្បន្ននិងជាមួយ CS អវត្តមាន covariates ដាច់ដោយឡែករកមិនឃើញផលប៉ះពាល់គួរអោយកត់សំគាល់ណាមួយឡើយ។ F(1, 18) <០០០០១, p > ០.៣១ ។ ដូច្នេះវិធីសាស្រ្តឆ្លើយតបក្នុងកំឡុងពេលហ្វឹកហាត់មិនបានសម្របសម្រួលភាពខុសគ្នានៃក្រុមក្នុងការឆ្លើយតបទៅនឹង AMPH 0.31 មីលីក្រាម / គីឡូក្រាម។
ក្រុម 5 ពីរ× 5 សម័យ ANCOVA នៃក្បាលម៉ាស៊ីនឆ្លើយតបទៅនឹង 1 មីលីក្រាម / គីឡូក្រាមក្នុងអំឡុងពេលវគ្គរំញោចអារម្មណ៍ជាមួយនឹងផតថលសរុប (អេសអេសអេសអេសអេកអេក) ខណៈដែលសារធាតុariatesដាច់ដោយឡែកមិនបានផ្តល់ផលប៉ះពាល់សំខាន់នៃ covariate ខណៈពេលដែល CS មានវត្តមាន។ F(4, 104) <០០០០១, p > ០,៣៨ និងផលប៉ះពាល់សំខាន់នៃ covariate ខណៈ CS អវត្តមាន F(1, 18) = 3.32, p = 0.085 ។
ការពិភាក្សា
លទ្ធផលនៃការសិក្សានេះមិនគាំទ្រដល់សម្មតិកម្មដែលថាក្រុម 50 នឹងបង្ហាញការឆ្លើយតបខ្ពស់ជាងក្បាលម៉ាស៊ីននៅលើវគ្គបើប្រៀបធៀបទៅនឹងក្រុមផ្សេងទៀត។ ទិន្នន័យ AMPH 1 មីលីក្រាម / គីឡូក្រាមបានបញ្ជាក់ពីការកើតឡើងនៃការប្រែលប្រួលជាមួយនឹងរបបប្រើប្រចាំថ្ងៃជំនួស។ លំនាំឆ្លងកាត់ក្រុមបានបង្ហាញពីនិន្នាការសម្រាប់ការប្រែលប្រួលកាន់តែខ្លាំងក្នុងវគ្គចុងក្រោយនៅក្នុងក្រុម 25 ដោយមិនមានភស្តុតាងបែបនេះសម្រាប់ក្រុម 50 ។ ផ្ទុយទៅវិញលទ្ធផលនៃកំរិតដូស 0.5 មីលីក្រាម / គីឡូក្រាមបានបង្ហាញពីនិន្នាការនៃការប្រែលប្រួលខ្លាំងនៅក្នុងក្រុម 50 ខណៈពេលដែលការបញ្ជាក់អំពីការកើនឡើងជាទូទៅនៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីននៅទូទាំងក្រុមទៅក្រុមទី ២ ទល់នឹងកម្រិតថ្នាំ AMPH 0.5 mg / kg ដំបូង។ ផលប៉ះពាល់នៃការចាក់ជាតិប្រៃបានបញ្ជាក់ថាភាពតានតឹងឬភាពតានតឹងដែលទាក់ទងនឹងការចាក់មិនបានរួមចំណែកដល់ផលប៉ះពាល់ AMPH ទេ។
The nose poke data again revealed an overall increase in approach responding over the course of training sessions when the CS was present, with no corresponding increase when the CS was absent. Therefore, the animals appeared to acquire the association between the CS and the prospect of sucrose reward. Group differences in the frequency of nose pokes when the CS was present conformed roughly to the frequency of reward delivery under the respective schedules, with groups 75 and 100 displaying the most nose pokes, group 50 displaying intermediate numbers of nose pokes, and groups 0 and 25 displaying the fewest nose pokes. These results suggest that the CS came to control approach responding in a manner consistent with the overall probability of reward. Although speculative, one possible explanation for the lower nose poke rates with CS present in group 50 in experiment 2 vs. experiment 1 may be the shortening of the inter-trial interval, as longer inter-trial intervals (experiment 1) appear to encourage impulsive tendencies and this is associated with increased turnover of DA in anterior cingulate, prelimbic and infralimbic cortices (Dalley et al ។ , 2002) ។ ដូច្នេះការកាត់បន្ថយ 30% នៃចន្លោះពេលនៃការសាកល្បងក្នុងការពិសោធន៍ 2 (និង 3) អាចមានការផ្លាស់ប្តូរកំរិត Cortical DA និងជំរុញអោយមានការជ្រើសរើសកាន់តែច្រើន (មានន័យថាត្រូវបានណែនាំដោយប្រេកង់នៃរង្វាន់) និងវិធីសាស្រ្តជំរុញ (មិនណែនាំដោយប្រេកង់រង្វាន់) ។ ឆ្លើយតបជាក្រុម 50 ក្នុងកំឡុងពេលហ្វឹកហាត់ក្នុងការពិសោធ 2 ប្រៀបធៀបនឹងការពិសោធ 1 ។
កង្វះនៃផលប៉ះពាល់ដែលទាក់ទងទៅនឹង covariate សម្រាប់ផែ្នកច្រមុះនៅក្នុងស្ថានភាពបច្ចុប្បន្នរបស់ CS នៅក្នុង ANCOVAs បង្ហាញថាវិធីសាស្រ្តឆ្លើយតបក្នុងកំឡុងពេលហ្វឹកហាត់ sucrose មិនបានសំរបសំរួលផលប៉ះពាល់នៃកាលវិភាគ CS-sucrose ផ្សេងគ្នាលើការឆ្លើយតបទៅនឹង AMPH ។ ផលប៉ះពាល់សំខាន់នៃ covariate សម្រាប់ស្ថានភាពអវត្តមាន CS នៅក្នុង ANCOVA នៃការឆ្លើយតបនៃក្បាលម៉ាស៊ីនទៅនឹង 1 មីលីក្រាម / គីឡូក្រាម AMPH បានបង្ហាញថាទំនោរទៅរកការឆ្លើយតបដោយថ្នាំមិនគ្រប់ខែបានពន្យល់ពីភាពប្រែប្រួលខ្លះនៃផលប៉ះពាល់ក្បាលម៉ាស៊ីនរបស់ AMPH ក្នុងកំឡុងពេលចាប់អារម្មណ៍។
ភស្តុតាងរួមបង្ហាញថាផលប៉ះពាល់នៃប្រវត្តិម៉ាស៊ីនត្រជាក់អាចត្រូវបានគេស្គាល់ច្បាស់ជាមួយ 0.5 AMPH ជាង 1 mg / kg AMPH ហើយពិធីសារដែលបង្កើតការប្រែលប្រួលក្នុងករណីដែលមិនមានឧបាយកលផ្សេងទៀតអាចនឹងធ្វើឱ្យមើលមិនឃើញឬបង្ហាញពីផលប៉ះពាល់នៃផលប៉ះពាល់ដែលមិនសមហេតុផល។ ឧបាយកល - លើកកម្ពស់ឧបាយកលអាកប្បកិរិយា (មានន័យថារង្វាន់អថេររ៉ាំរ៉ៃ) ។
Behavioral sensitization to AMPH is a robust effect in the laboratory. However, outside the laboratory, only a minority of individuals who gamble chronically escalate to pathological levels. Although risk for sensitization is related to risk for addiction (or drug seeking), especially for psychostimulants (Vezina, 2004; Flagel et al ។ , 2008), many factors aside from sensitization risk may predispose one to addiction (e.g., Verdejo-Garcia et al ។ , 2008; Conversano et al., 2012; Volkow et al ។ , 2012) ។ ទោះយ៉ាងណាកត្តាចរិតដែលផ្តល់ភាពងាយរងគ្រោះដល់ការប្រែលប្រួលអាចមានអន្តរកម្មជាមួយប្រវត្ដិសាស្ដ្រដើម្បីបញ្ជាក់ពីផលប៉ះពាល់នៃរង្វាន់ដែលមិនអាចទាយទុកជាមុនបាន (ឧទាហរណ៍កាលវិភាគ 50% ស៊ី។ អេស។ អេស។ អេស) លើប្រតិកម្មប្រព័ន្ធឌីអេ។ ដើម្បីស៊ើបអង្កេតពីលទ្ធភាពនេះការពិសោធន៍ 3 បានប្រើនីតិវិធីដូចគ្នានឹងការពិសោធ 2 ដែរប៉ុន្តែបានប្រើសំពាធ Lewis ជំនួសឱ្យ Sprague Dawley ។
Sprague Dawley rats display intermediate levels of DA transporters, with lower levels than Wistar strain rats (Zamudio et al., 2005), but higher levels than Wistar-Kyoto rats (a “depressive”-like strain) in the nucleus accumbens, amygdala, ventral tegmental area and substantia nigra (Jiao et al., 2003). This profile may render Sprague Dawley rats only moderately sensitive to environmental or pharmacological manipulations of DA function. In contrast, Lewis rats exhibit low levels of DA transporters as well as D2 and D3 DA receptors in the nucleus accumbens and dorsal striatum compared to other strains (e.g., F344) (Flores et al ។ , 1998). These morphological differences may contribute to Lewis rats’ differential response to DA manipulations. Lewis rats also exhibit a range of accentuated responses to experimental drug manipulations compared to other strains (e.g., F344). Most importantly, Lewis rats display greater sensitization to methamphetamine, characterized by low response to initial doses but higher response to later doses (Camp et al., 1994). Lewis rats also exhibit greater locomotor sensitization to a range of doses of cocaine (Kosten et al ។ , 1994; Haile et al ។ , 2001) ។ ផ្អែកលើគំរូនៃផលប៉ះពាល់នេះយើងសន្មតថាកណ្តុរឡឺវីសអាចឱ្យយើងធ្វើការស៊ើបអង្កេតថាតើភាពងាយនឹងទទួលបានការចាប់អារម្មណ៍នឹងពង្រីកឥទ្ធិពលនៃកាលវិភាគម៉ាស៊ីនត្រជាក់លើការឆ្លើយតបជាបន្តបន្ទាប់ចំពោះអេអឹមអេជ។
ពិសោធន៍ 3
សំភារៈនិងវិធីសាស្រ្ត
វិធីសាស្រ្តគឺដូចគ្នានឹងការពិសោធ 2 ដែរក្រៅពីការប្រើប្រាស់កណ្តុរឡេវីស (200 – 225 ក្រាមនៅពេលមកដល់ទន្លេឆាលឃិនឃ្វីបប្រទេសកាណាដា) ។
លទ្ធផល
ច្រមុះហៀរក្នុងកំឡុងពេលវគ្គ sucrose ។
ដំណាក់កាលទី 5 ក្រុម×វគ្គទី 15 វគ្គ× 2 ដំណាក់កាល (បច្ចុប្បន្ន CS, អវត្តមាន CS) ANOVA នៃច្រមុះច្រមុះផ្តល់នូវផលប៉ះពាល់សំខាន់ៗនៃក្រុម, F(4, 34) = 6.12, p = 0.001, សម័យ, F(14, 476) = 3.42, p <០.០០១ និងដំណាក់កាល F(1, 34) = 20.83, p <០,០០១ ក៏ដូចជាអន្តរកម្មបីផ្លូវយ៉ាងសំខាន់។ F(56, 476) = 1.56, p = 0.008 ។ បន្ទះ។ (A, B) នៃរូប។ 7 រៀបចំផែនការដាក់ពិន្ទុច្រមុះសម្រាប់ច្រមុះសម្រាប់ក្រុមបច្ចុប្បន្ននិង CS អវត្តមានដំណាក់កាលរៀងគ្នា។ ការប្រៀបធៀបនៃបន្ទះទាំងពីរបង្ហាញថាឥទ្ធិពលសំខាន់នៃដំណាក់កាលបានឆ្លុះបញ្ចាំងពីការឆ្លើយតបនៃការស្ទុះច្រមុះជាទូទៅនៅពេលដែល CS មានវត្តមានទល់នឹងអវត្តមាន។ ហេតុដូច្នេះហើយការឆ្លើយតបដែលបានលើកឡើងបានកើតឡើងជាញឹកញាប់ជាងការឆ្លើយតបមុនពេញវ័យ។ ផលប៉ះពាល់សំខាន់ៗនៃក្រុមនិងសម័យមិនត្រូវបានបកស្រាយងាយទេដោយសារអន្តរកម្មលំដាប់ខ្ពស់។ អន្តរកម្មទាំងបីបានឆ្លុះបញ្ចាំងពីការរួមបញ្ចូលគ្នានៃពិន្ទុសម្រាប់ក្រុមទាំង ៥ ក្នុងកម្រិតទាបមួយដែលមានស្ថេរភាពឆ្លងកាត់វគ្គនានានៅពេលដែលអវត្តមាន [បន្ទះ (ខ)] រួមជាមួយការបង្វែរពិន្ទុនៅពេលដែលអេស។ ស៊ី។ ឌី។ អេសមានវត្តមានក្នុងទម្រង់ខុសគ្នាដាច់ពីគ្នាសម្រាប់ក្រុមនីមួយៗដែលស្របគ្នានឹងលំដាប់នៃចំនួនប្រេកង់រង្វាន់របស់ពួកគេ៖ ពីខ្ពស់បំផុត (ក្រុម 100) ទៅទាបបំផុត (ក្រុម 25) [បន្ទះ (ក)] ។ មានតែនិន្នាការលីនេអ៊ែរសម្រាប់អន្តរកម្មប៉ុណ្ណោះដែលសំខាន់។ F(4, 34) = 4.03, p = 0.009, ឆ្លុះបញ្ចាំងពីការកើនឡើងប្រហាក់ប្រហែលនៃផែ្នកច្រមុះលើវគ្គនៅក្នុងក្រុម 100 នៅពេលដែល CS មានវត្តមានប្រឆាំងនឹងទម្រង់មិនមានការកើនឡើងគួរអោយកត់សំគាល់នៃការកើនឡើងនៃច្រមុះតាមរយៈវគ្គផ្សេងៗនៅក្នុងក្រុមផ្សេងទៀតក្នុងដំណាក់កាលនេះ។
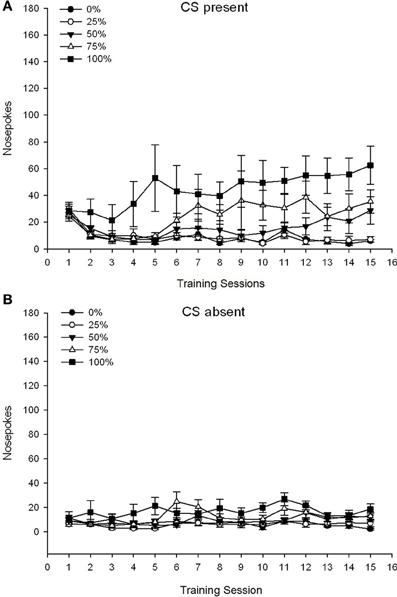
Figure 7. Mean (SE) approach responses (nose pokes) on 15 sucrose training sessions in groups of Lewis rats (n = 8 / ក្រុម) ប្រឈមនឹងរង្វាន់ sucrose (ដំណោះស្រាយ 10%) ដែលបានផ្តល់ជូននៅក្រោម 0, 25, 50, 75, ឬកាលវិភាគអថេរ 100%. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. (ក) Scores when CS was present (5 s × 20 trials). (ខ) Scores when CS was absent (average for 5 × 20 s while light was off).
ជំរកទៅនឹងប្រអប់ក្បាលម៉ាស៊ីន។
ក្រុម 5 គ្រុប 3 សម័យ ANOVA បានផ្តល់នូវឥទ្ធិពលសំខាន់នៃសម័យប្រជុំ, F(2, 70) = 23.07, p <០.០០១ និងមិនមានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត F(8, 70) <០០០០១, p > ០.១៨ ។ លំនាំក្បាលរថភ្លើងកោងនៃមធ្យម (SE) បានផុសចេញពីវគ្គទី ១, ១០៧៦ (៧៤), តាមរយៈវគ្គទី ២, ៦៤៤ (៤៨), ដល់វគ្គទី ៣, ៧៦២ (៥៩) ។ ការប្រៀបធៀបដែលបានគ្រោងទុកនៃក្រុម ៥០ ជាមួយក្រុម ០ និងក្រុម ១០០ នៅវគ្គជម្រុះដំបូងនិងចុងក្រោយបានបង្ហាញថាការបំបែកធ្នឹមមានតិចជាងនៅក្នុងក្រុម ៥០ (M = 911; SE = 109) ទល់នឹងក្រុម 0 (M = 1103; SE = 176) លើវគ្គជំរកទី 1, t(105) = 2.02, p <០.០៥ ប៉ុន្តែមិនខុសគ្នារវាងក្រុម ៥០ និងក្រុម ១០០ (M = 1066; SE = 150), t(105) <០០០០១, p > ០.២០ នៅសម័យនេះ។ ក្រុម ៥០ មិនខុសគ្នាខ្លាំងពីក្រុម ០ ឬក្រុម ១០០ ក្នុងវគ្គជម្រុះចុងក្រោយទេ។ t(105) <០០០០១, p > ០.៣០ ។ ដូច្នេះមានន័យថាការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនដែលគ្មានគ្រឿងញៀននៅក្នុងក្រុមសំខាន់ៗមិនមានភាពខុសគ្នាជាប្រចាំទេមុនពេលធ្វើតេស្ត។
វគ្គសាកល្បង។
Saline។ 5 Group × 2 Session ANOVA នៃក្បាលម៉ាស៊ីនក្បាលម៉ាស៊ីននៅលើវគ្គជំរកចុងក្រោយនិងវគ្គសាកល្បងអំបិលផ្តល់នូវឥទ្ធិពលសំខាន់នៃសម័យ។ F(1, 35) = 50.12, p <០.០០១ និងមិនមានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត F(4, 35) <០០០០១, p > ០.១៩ ។ រូបភាព 8 បង្ហាញក្រុមពិន្ទុមធ្យមសម្រាប់វគ្គទាំងពីរហើយបង្ហាញថាផលប៉ះពាល់វេនឆ្លុះបញ្ចាំងពីការធ្លាក់ចុះគួរឱ្យកត់សម្គាល់ពីទំលាប់ទៅការធ្វើតេស្តជាតិប្រៃ។ ដូច្នេះការទទួលការចាក់។ ក្នុងមួយ SE (ឧទាហរណ៍ភាពតានតឹងភាពតានតឹង) មិនបានបង្កើនការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទេ។
Figure 8. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) on the last of 3 drug-free habituation sessions and on a subsequent session after saline injection (i.p., 1 ml/kg) in groups of Lewis rats (n = 8/group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, 75, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli.
ផលប៉ះពាល់នៃ 0.5 មីលីក្រាម / គីឡូក្រាម AMPH ។.
ក្បាលម៉ាស៊ីនចាក់មុន។។ ក្បាលម៉ាស៊ីន 5 គ្រុប× 2 សម័យ ANOVA នៃក្បាលម៉ាស៊ីនចាក់មុនទទួលបានលទ្ធផលសំខាន់គួរឱ្យកត់សំគាល់នៃសម័យ។ F(1, 35) = 15.04, p <០.០០១ និងមិនមានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត F(4, 35) <០០០០១, p > ០.៣៣ ។ ការប្រៀបធៀបដែលបានគ្រោងទុកមិនមានភាពខុសគ្នាខ្លាំងរវាងក្រុម ៥០ និងក្រុម ០ ឬក្រុម ១០០ ទាំងវគ្គសាកល្បងទេ។ t(70) <០០០០១, p > ០.៣០ ។ ដូច្នេះភាពខុសគ្នានៃមូលដ្ឋាននៅក្នុងក្បាលរថភ្លើងចាក់បញ្ចូលមុនមិនគិតពីភាពខុសគ្នានៃក្រុមនៅក្នុងការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹង 0.30 មីលីក្រាម / គីឡូក្រាម AMPH ទេ។ ការបំបែកធ្នឹមមធ្យម (SE) សម្រាប់ដំណាក់កាលចាក់មុនសម្រាប់វគ្គទី ១ និងទី ២ (ក្រោយពេលចាប់អារម្មណ៍) ០,៥ មីលីក្រាម / គីឡូក្រាមគឺ ៣២៥ (២៥) និង ៤៧៣ (៣៦) ។
ក្បាលរថភ្លើងក្រោយពេលចាក់។។ ក្រុម 5 ក្រុម× 2 សម័យ ANOVA នៃក្បាលម៉ាស៊ីនក្បាលម៉ាស៊ីនឆ្លើយតបនឹង 0.5 មីលីក្រាម / កិតើគីឡូក្រាមដែលបានផ្តល់មុននិងក្រោយការរ៉ាំរ៉ៃ 1 មីលីក្រាម / គីឡូក្រាម AMPH មានឥទ្ធិពលសំខាន់នៃសម័យ។ F(1, 34) = 87.44, p <០.០០១ និងមិនមានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត F(4, 34) <០០០០១, p > ០.១៩ ។ រូបភាព 9 គ្រោងពិន្ទុក្បាលរថភ្លើងមធ្យមសម្រាប់ក្រុមនិងវគ្គនីមួយៗហើយបង្ហាញថាបែបផែនសម័យបានឆ្លុះបញ្ចាំងពីការឆ្លើយតបជាទូទៅកើនឡើងចំពោះកម្រិតថ្នាំ 0.5 មីលីក្រាម / គីឡូក្រាមលើកទីពីរស្របនឹងការគិត។ តួលេខនេះក៏បង្ហាញផងដែរថាក្រុមទាំងនោះបានអនុវត្តស្រដៀងគ្នានៅលើវគ្គទី 1 ប៉ុន្តែក្រុមនោះ 50 បានបង្ហាញសកម្មភាពក្បាលម៉ាស៊ីនច្រើនជាងក្រុមដទៃទៀតនៅលើវគ្គ 2 ។ ការប្រៀបធៀបដែលបានគ្រោងទុកក្នុងការឆ្លើយតបទៅនឹងកម្រិតថ្នាំ 0.5 មីលីក្រាម / គីឡូក្រាមដំបូងបានបង្ហាញថាមិនមានភាពខុសគ្នាខ្លាំងរវាងក្រុម 50 និងក្រុម 0 ឬក្រុម 100, t(35) <០០០០១, p > ០.២០ ។ ទោះយ៉ាងណាក្រុមទី ៥០ បានបង្ហាញនូវការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនកាន់តែធំទៅនឹងកម្រិតថ្នាំ ០.៥ មីលីក្រាម / គីឡូក្រាមលើកទី ២ ជាងក្រុម ០, t(35) = 4.32, p <០.០០១ ឬក្រុម ១០០, t(35) = 2.24, p <0.05 ។
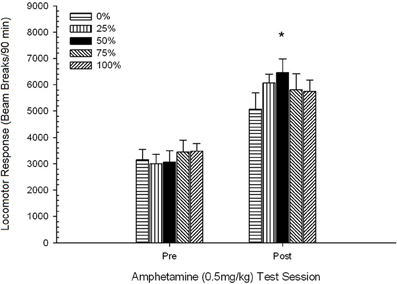
Figure 9. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) to 0.5 mg/kg d-amphetamine on separate sessions before and after a 5-session sensitizing regimen of d-amphetamine (1.0 mg/kg; i.p. per session) in groups of Lewis rats (n = 8/group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, 75, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. *p < 0.05 for mean difference between group 50 and group 0 as well as group 100, based on planned comparisons.
ផលប៉ះពាល់នៃ 1 មីលីក្រាម / គីឡូក្រាម AMPH ។.
ក្បាលម៉ាស៊ីនចាក់មុន។។ 5 Group × 5 Session ANOVA នៃ 30- នាទីមុនការចាក់បញ្ចូលពិន្ទុសម្រាប់វគ្គដែលទទួលបានលទ្ធផលមានឥទ្ធិពលសំខាន់នៃសម័យ។ F(4, 140) = 4.10, p = 0.004, និងគ្មានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត, F(4, 35) = 1.25, p > ០.៣១ ។ ការប្រៀបធៀបដែលបានគ្រោងទុកបានរកឃើញថាការបំបែកធ្នឹមក្នុងដំណាក់កាលមុនចាក់ (M; SE) គឺទាបជាងក្នុងក្រុម ៥០ (៣៩៥; ៦២) ជាងក្រុម ១០០ (៥០៨; ៦២) ។ t(175) = 2.58, p <0.01 ប៉ុន្តែមិនមែនក្រុម ០, t(175) <០០០០១, p > ០.១០ វគ្គ AMPH ១ មីលីក្រាម / គីឡូក្រាម ១. វគ្គចុងក្រោយ AMPH ១ មីលីក្រាម / គីឡូក្រាមការប្រៀបធៀបដែលបានគ្រោងទុកក៏បានរកឃើញថាក្បាលម៉ាស៊ីនចាក់មុនក្នុងក្រុម ៥០ (៣៧៨; ៦០) គឺទាបជាងក្នុងក្រុម ១០០ (៦៥០; ៧៥) ។ ), t(175) = 6.17, p <០.០០១ ប៉ុន្តែមិននៅក្នុងក្រុម ០, t(175) <០០០០១, p > ០.១០ ។ ដោយសារទិសដៅនៃភាពខុសគ្នានៃក្រុមទាំងនេះ (ក្រុមត្រួតពិនិត្យ = ក្រុម ៥០) គឺផ្ទុយពីគំរូដែលបានសម្មតិកម្មភាពខុសគ្នារបស់ក្រុមនៅក្នុងក្បាលរថភ្លើងក្រោយចាក់បញ្ចូលគ្នាដែលសន្មតជាមួយសម្មតិកម្មមិនអាចត្រូវបានសន្មតថាជាភាពខុសគ្នានៃមូលដ្ឋានចាក់មុន។ មធ្យម (SE) ការបំបែកធ្នឹមទាំងមូលក្នុងដំណាក់កាលមុនចាក់សម្រាប់វគ្គ ១ ដល់ ៥ គឺ៖ ៤៤២ (៣៤) ៤៥២ (៣២) ៥៤២ (៤០) ៤១១ (២៦) ៥០៤ (៣៧) ។
ក្បាលរថភ្លើងក្រោយពេលចាក់។។ ក្រុម 5 ក្រុម× 5 វគ្គ ANOVA នៃការឆ្លើយតបចំពោះដូស 1 មីលីក្រាម / គីឡូក្រាមបានផ្តល់នូវឥទ្ធិពលសំខាន់នៃសម័យប្រជុំ, F(4, 140) = 6.15, p <០.០០១ និងមិនមានផលប៉ះពាល់គួរឱ្យកត់សម្គាល់ផ្សេងទៀត F(4, 35) <០០០០១, p > ០.៧៧ ។ ការវិភាគនិន្នាការពហុវិជ្ជាបានបង្ហាញពីនិន្នាការលីនេអ៊ែរគួរឱ្យកត់សម្គាល់ F(1, 35) = 9.34, p = 0.004 និងនិន្នាការគូប F(1, 35) = 5.08, p = 0.031, លទ្ធផលចុងក្រោយបង្ហាញអតិបរិមាទាក់ទងនៅវគ្គទី 3 និង 5 ។ រូបភាព។ 10 គ្រោងពិន្ទុទាំងនេះហើយបង្ហាញថាទោះបីខ្វះអន្តរកម្មសំខាន់នៅក្នុង ANOVA ក៏ដោយក៏ក្រុម 50 បានបង្ហាញក្បាលរថភ្លើងធំជាងក្រុម ៤ ផ្សេងទៀតក្នុងការឆ្លើយតបទៅនឹងកំរិតថ្នាំ 1 mg / kg ចុងក្រោយ។ ដូច្នោះហើយការប្រៀបធៀបដែលបានគ្រោងទុកបានបង្ហាញពិន្ទុមធ្យមគួរឱ្យកត់សម្គាល់លើវគ្គទី 5 ក្នុងក្រុម 50 ជាងក្រុមដទៃទៀតទាំងអស់ t(35) > ១៥, p <0.001 ។
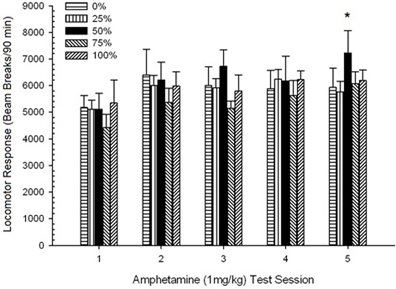
Figure 10. Mean (SE) locomotor response (number of beam breaks in an electronic array per 90 min) to 1 mg/kg d-amphetamine (i.p.) on 5 weekly sessions in groups of Lewis rats (n = 8/group) previously exposed to 15 daily conditioning sessions with sucrose reward (10% solution) delivered under 0, 25, 50, 75, or 100% variable schedules. The conditioned stimulus was a light (120 s). Group 0 received the same number of rewards as group 100 in the absence of conditioned stimuli. *p < 0.05 for mean difference between group 50 and group 0 as well as group 100, based on planned comparisons.
គ្រប់គ្រងការផ្លាស់ប្តូរនៃការញ័រច្រមុះឆ្លើយតបក្នុងកំឡុងពេលហ្វឹកហាត់ sucrose ។
ក្រុម 5 ពីរក្រុម NUM 2 វគ្គ ANCOVA នៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹង 0.5 មីលីក្រាម / គីឡូក្រាម AMPH មុននិងក្រោយតំរែតំរង់អារម្មណ៍រួមទាំងផែ្នកច្រមុះសរុបក្នុងកំឡុងពេលហ្វឹកហាត់ sucrose ជាមួយ CS បច្ចុប្បន្ននិងជាមួយ CS អវត្តមាន covariates ដាច់ដោយឡែករកមិនឃើញផលប៉ះពាល់គួរអោយកត់សំគាល់ណាមួយឡើយ។ F(1, 32) <0.44 p > ០.៥១ ។ ANCOVAs រយៈពេល ៥ ក្រុម× ៥ នៃការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹង AMPH ១ មីលីក្រាម / គីឡូក្រាមក្នុងកំឡុងពេលនៃការចាប់អារម្មណ៍ជាមួយនឹងផែ្នកច្រមុះសរុប (អេកអេសអេសអេសអេកអេក) ខណៈពេលដែលសារធាតុariatesដាច់ដោយឡែកមិនបានផ្តល់ផលប៉ះពាល់សំខាន់នៃ covariate ទេខណៈពេលដែល CS មានវត្តមានឬអវត្តមាន។ F(1, 33) <០០០០១, p > ០.៧១ ។ ដូច្នេះវិធីសាស្រ្តក្នុងការឆ្លើយតបចំពោះគ្រឿងញៀនមិនគិតពីភាពខុសគ្នានៃក្រុមនៅក្នុងការឆ្លើយតបនឹងក្បាលម៉ាស៊ីនចំពោះកម្រិតអេមភីអេ។
ការពិភាក្សា
Sensitization developed to the effects of repeated 1.0 mg/kg amphetamine. The habituation and saline data confirm that this effect was not due to pre-existing differences, expectancy, or stress-related responses to the injection. The ANCOVAs with nose pokes confirm that these effects were not due to drug-free approach behavior. The nose poke data themselves indicated that the groups acquired the association between the CS and prospect of sucrose reward. The groups’ rank level of nose-poke responding at the end of training matched the overall frequency of reward under the different schedules from highest (group 100) to lowest (group 0), as it did in experiment 2. The relatively lower overall mean nose poke levels in this experiment compared to experiments 1 and 2 may reflect more selective approach responding to cues for reward in Lewis rats (Kosten et al ។ , 2007).
ទិន្នន័យកំរិតដូស 0.5 មីលីក្រាម / គីឡូក្រាមបានបង្ហាញថាការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនដំបូងចំពោះអេអឹមភីអេនៅក្នុងកណ្តុរឡេវីស (រូប។ 9) ត្រូវបានបង្ក្រាបខ្លះបើប្រៀបធៀបទៅនឹងកណ្តុរ Sprague Dawley (ពិសោធន៍ 2; រូបភាព។ 5) ប៉ុន្តែការកើនឡើងជាក្រុមក្នុងការឆ្លើយតបទៅនឹងកិតទី ២ នៃកណ្តុរឡឺវីសគឺគួរឱ្យកត់សម្គាល់ (ជិតទ្វេដងនៃការឆ្លើយតបទៅនឹងកំរិតថ្នាំ 0.5 មីលីក្រាម / គីឡូក្រាមដំបូង) បន្ទាប់ពីរបប 5-session AMPH តំរែតំរង់គួរអោយកត់សំគាល់បំផុតក្រុម 50 បង្ហាញការឆ្លើយតបក្បាលម៉ាស៊ីនកាន់តែធំ ច្រើនជាងក្រុមទាំងអស់លើកលែងតែក្រុម 25 ដល់វិនាទី (មានន័យថាកំរិតក្រោយ) កំរិតថ្នាំ 0.5 mg / kg កំរិត AMPH និងការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនធំជាងក្រុមដទៃទៀតរួមទាំងក្រុម 25 ដល់កំរិតថ្នាំ 1 mg / kg កំរិត AMPH ចុងក្រោយ (វគ្គចុងក្រោយ) ។
ការវិភាគសង្ខេបនៃចំណាត់ថ្នាក់ក្រុមឆ្លងកាត់ការសាកល្បង។
To determine the reliability of group differences in sensitization, a non-parametric analysis assessed the contingency between group and rank of mean locomotor response to the second (post-chronic AMPH) 0.5 mg/kg dose and the final 1.0 mg/kg dose of AMPH from the 3 experiments. The analysis yielded a significant effect, φ = 0.986, p = 0.025, ឆ្លុះបញ្ចាំងពីការពិតដែលថាក្រុម 50 ស្ថិតនៅលំដាប់ទី ១ នៅក្នុងទាំងអស់ប៉ុន្តែមានតែការប្រៀបធៀបមួយប៉ុណ្ណោះ។ ចំណាត់ថ្នាក់ខ្ពស់នៃក្រុម 50 បើប្រៀបធៀបទៅនឹងក្រុមដទៃទៀតទាំងអស់ក្នុងការឆ្លើយតបទៅនឹងកំរិតថ្នាំទី ២ (ក្រោយរ៉ាំរ៉ៃ AMPH) 0.5 mg / kg ដូសត្រូវបានបង្ហាញក្នុងរូបភាព។ 5 (ពិសោធន៍ 2) និងរូបភាព។ 9 (ពិសោធន៍ 3) ។ ចំណាត់ថ្នាក់ខ្ពស់នៃក្រុម 50 ទាក់ទងនឹងក្រុមដទៃទៀតក្នុងការឆ្លើយតបទៅនឹងកំរិតថ្នាំ 1.0 មីលីក្រាម / គីឡូក្រាមចុងក្រោយត្រូវបានបង្ហាញនៅក្នុងរូបភាព 2 (ពិសោធន៍ 1) និងរូបភាព។ 10 (ពិសោធន៍ 3) ។ ការលើកលែងតែមួយគត់ចំពោះគំរូនេះគឺការឆ្លើយតបទៅនឹងកំរិតថ្នាំ 1.0 មីលីក្រាម / គីឡូក្រាមចុងក្រោយនៅក្នុងកណ្តុរ Sprague-Dawley ក្នុងការពិសោធន៍ 2 ។
ការពិភាក្សាទូទៅ
The present series of experiments tested the hypothesis that chronic exposure to a gambling-like schedule of reward can sensitize brain DA pathways much like chronic exposure to drugs of abuse. Evidence for such an effect would suggest that neuroplasticity, of the same kind thought to contribute to drug addiction, can be induced by chronic exposure to unpredictable reward schedules. In line with the literature on drug addiction, locomotor response to 0.5 and 1.0 mg/kg doses of AMPH indexed DA system reactivity, with greater locomotion in response to later doses operationally defining sensitization (cf. Robinson និង Berridge, 1993; Pierce និង Kalivas, 1997; Vanderschuren និង Kalivas, 2000).
សរុបមកលទ្ធផលគឺស្របនឹងសម្មតិកម្មរបស់យើង។ ទោះយ៉ាងណាក៏ដោយពួកគេក៏បង្ហាញពីភាពប្រែប្រួលគួរឱ្យកត់សម្គាល់នៅក្នុងផលប៉ះពាល់ពិសោធន៍ដោយសារតែកត្តានីតិវិធី។ ផលប៉ះពាល់នៃកាលវិភាគម៉ាស៊ីនត្រជាក់គឺមានកម្រិតមធ្យមប៉ុន្តែស្របគ្នាជាមួយនឹងក្រុម 50 បង្ហាញនូវការឆ្លើយតបច្រើនជាងក្រុម ៤ ផ្សេងទៀតចំពោះថ្នាំទាំងពីរនេះបន្ទាប់ពីលេបថ្នាំ ៥ ដូស។ បើទោះបីជារួម។ Fភាពខុសគ្នានៃផលប៉ះពាល់ទាក់ទងនឹងក្រុមនៅក្នុងការវិភាគបំរែបំរួលជាញឹកញាប់មិនសំខាន់ភាពខុសគ្នានៃក្រុមសំខាន់ៗត្រូវបានបញ្ជាក់ជាមួយនឹងការប្រៀបធៀបដែលបានគ្រោងទុកជាគូ។ ក្នុងន័យនេះវាគួរតែត្រូវបានកត់សម្គាល់ថា“ ការគិតបច្ចុប្បន្នទោះយ៉ាងណាគឺមានសារៈសំខាន់ជារួម [សម្រាប់។ F នៅក្នុង ANOVA] គឺមិនចាំបាច់ទេ។ ជាបឋមសម្មតិកម្មដែលត្រូវបានសាកល្បងដោយការធ្វើតេស្តទាំងមូលនិងការធ្វើតេស្តប្រៀបធៀបច្រើនគឺខុសគ្នាជាមួយនឹងកម្រិតថាមពលខុសគ្នា។ ឧទាហរណ៍ជារួម។ F ពិតជាចែកចាយភាពខុសគ្នាក្នុងចំណោមក្រុមនៅទូទាំងចំនួនដឺក្រេនៃសេរីភាពសម្រាប់ក្រុម។ នេះមានប្រសិទ្ធិភាពនៃការរលាយជាទូទៅ។ F in the situation where several group means are equal to each other but different from some other mean” (Howell, 1992, ទំ។ 338) ។ នេះគឺជាស្ថានភាពជាក់ស្តែងដែលបានអនុវត្តនៅក្នុងការពិសោធន៍បច្ចុប្បន្នដែលក្រុម 50 ត្រូវបានគេរំពឹងថាខុសគ្នាពីក្រុម 0 និងក្រុម 100 គ្រប់គ្រងប៉ុន្តែគ្មានភាពខុសគ្នារវាងក្រុមត្រួតពិនិត្យទាំងនេះត្រូវបានគេព្យាករណ៍សម្រាប់ក្រុម 25 ឬក្រុម 75 ។
ទិន្នន័យនៃការញ័រច្រមុះបានបញ្ជាក់ថានៅក្នុងរាល់ការពិសោធន៍សត្វបានទទួលការផ្សារភ្ជាប់គ្នារវាងស៊ីអេសនិងការរំពឹងទុកនៃរង្វាន់លើកទឹកចិត្ត។ ការឆ្លើយឆ្លងគ្នារវាងភាពញឹកញាប់នៃការញាក់ច្រមុះសម្រាប់ក្រុមផ្សេងៗគ្នានិងរង្វាន់ទូទៅនៅក្រោមកាលវិភាគបណ្តុះបណ្តាលរៀងៗខ្លួនបង្ហាញថាអត្រាមធ្យមនៃវិធីសាស្រ្តប្រើថ្នាំមិនផ្តល់រង្វាន់ឆ្លើយតប។ ទោះយ៉ាងណាក៏ដោយកង្វះប្រសិទ្ធភាពនៃការសំរបសំរួលនៃច្រមុះហៀរនៅលើការឆ្លើយតបនៃក្បាលម៉ាស៊ីនទាក់ទងទៅនឹងក្រុមទៅអេអឹមអេអេអេអេអេអេអេអេអូអេវ៉ាអេសបានបង្ហាញថាដំណើរការដាច់ដោយឡែកបង្ហាញពីឥរិយាបថទាំងពីរ។
ក្នុងករណីខ្លះឥទ្ធិពលនៃកាលវិភាគម៉ាស៊ីនត្រជាក់ត្រូវបានបង្ហាញយ៉ាងច្បាស់ក្នុងការឆ្លើយតបទៅនឹងកំរិត AMPH ដំបូង។ ក្នុងករណីផ្សេងទៀតវាកើតឡើងតែបន្ទាប់ពីប្រើម្តងហើយម្តងទៀត។ ភាពខុសគ្នានៃក្រុមនៅក្នុងក្បាលម៉ាស៊ីនក្បាលម៉ាស៊ីនឆ្លើយតបនឹងកំរិត AMPH ដំបូងបានបង្ហាញថាការប៉ះពាល់ទៅនឹងកាលវិភាគរង្វាន់ដូចល្បែងគឺគ្រប់គ្រាន់ដោយខ្លួនវាផ្ទាល់ដើម្បីជំរុញឱ្យមានការរំញោចអារម្មណ៍។ ភាពខុសគ្នានៃក្រុមនៅក្នុងក្បាលរថភ្លើងបន្ទាប់ពីការប្រើថ្នាំ AMPH ច្រើនបង្ហាញពីឥទ្ធិពលដែលអាចត្រូវបានកំណត់ថាជាភាពងាយទទួលរងគ្រោះដែលមានតែការបង្ហាញនៅពេលបញ្ចូលជាមួយការប៉ះពាល់ជាបន្តទៅនឹងភ្នាក់ងាររំញោចបឋម (ឧទាហរណ៍ថ្នាំអំហ្វេតាមីន) ។
Differences in the pattern of response across experiments suggest that a longer interval between training and initial AMPH challenge may maximize the opportunity to detect the inherent sensitizing effect of the conditioning treatment. This in turn suggests that effects of conditioned reward exposure may incubate over time, a phenomenon also seen with stimulant sensitization (Grimm et al ។ , 2006) ។ លំនាំនៃការឆ្លើយតបទៅនឹងថ្នាំអាហ្វេតាមីនពីរដូសបង្ហាញថាកំរិតដូស 0.5 មីលីក្រាម / គីឡូក្រាមអាចមានប្រសិទ្ធិភាពជាងមុនក្នុងការបង្ហាញពីផលប៉ះពាល់នៃប្រវត្តិម៉ាស៊ីនត្រជាក់។ នេះបង្ហាញថាផលប៉ះពាល់ម៉ាស៊ីនត្រជាក់ក្រោមពិធីសារនៃការបណ្តុះបណ្តាលនាពេលបច្ចុប្បន្ននេះគឺមិនច្បាស់ទេហើយអាចត្រូវបានក្លែងបន្លំដោយផលប៉ះពាល់នៃពិដានក្រោមកំរិតនៃ AMPH និងលក្ខខណ្ឌដែលបង្កើត de novo ការលើកទឹកចិត្ត។
In experiment 3, the biphasic pattern of response to the 0.5 mg/kg doses and progressive emergence of superiority in group 50 is consistent with the expected profile for Lewis rats in response to methamphetamine (Camp et al., 1994) ។ នេះផ្តល់ការគាំទ្រដល់សុពលភាពនៃលទ្ធផលដែលបានរកឃើញនាពេលបច្ចុប្បន្ននេះនិងបានបង្ហាញពីការត្រួតស៊ីគ្នារវាងកត្តាដែលងាយរងផលប៉ះពាល់កម្រិតមធ្យមទៅនឹងភាពរំជើបរំជួលចិត្តសាស្ត្រនិងតារាងពេលវេលានៃរង្វាន់។
Across experiments, the post-sensitization locomotor response of group 50 generally exceeded that of the other groups under different doses of amphetamine and in different strains of animals. However, the high within-group variability and modest between-group effect sizes indicate a role for other factors in DA system reactivity to amphetamine following exposure to varying schedules of conditioned sucrose reward. Although responses of DA neurons to reward signals may provide a coarse model of gambling (Fiorillo et al ។ , 2003) ដូចម៉ូដែលទាំងអស់ដែរដែលមានការបាត់បង់ព័ត៌មានសម្រាប់ជាប្រយោជន៍នៃការឃ្លាំមើលពោលគឺដើម្បីបង្ហាញដំណើរការសំខាន់មួយ។ ជាលទ្ធផលលំនាំនៃផលប៉ះពាល់នៅទូទាំងលក្ខខណ្ឌស៊ីអេសអេសនៅក្នុងហ្វ័រឌូលី et al ។ ការសិក្សាមិនបានធ្វើឱ្យទូទៅទៅនឹងការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹងថ្នាំអំហ្វេតាមីនទេ។ ការកែលម្អបន្ថែមទៀតនៃគំរូត្រូវបានអំពាវនាវឱ្យចាប់យកទិដ្ឋភាពនៃការលេងល្បែងដែលមានឥទ្ធិពលលើមុខងាររបស់ប្រព័ន្ធឌីអេ។
Taken together, the results of this series of experiments provide provisional support for the hypothesis that chronic exposure to gambling-like schedules of reward enhances the reactivity of the brain DA system to psychostimulant challenge. As such, they extend the findings of Singer et al. (2012) who demonstrated that, relative to a fixed schedule, prior exposure to a variable reinforcement schedule in an operant paradigm enhances subsequent locomotor response to amphetamine. More specifically, the present findings point to uncertainty of reward delivery as the critical factor underlying the effects of variable reward. The magnitude of effects in the operant paradigm was substantially greater than the effects found in the present experiments. This may reflect greater chronic exposure to the gambling-like activity (55 vs. 15 days); it may reflect the effects of requiring an operant response to elicit the reward (i.e., a role for agency) rather than passive exposure, as in the present study. Increasing the duration of training in the present paradigm would help to resolve these questions.
The validity of variable reward and reinforcement schedules as models of gambling cannot be gleaned from these experiments. Future research that examines the impact of conditioning history on risk-taking behavior in rodent gambling tasks could address this issue. Similarly, the correspondence between the behavioral sensitization found here and the elevated striatal DA response to amphetamine recently found in pathological gamblers must await further investigation (Boileau et al ។ , 2013) ។ ការធ្វើកោសល្យវិច័យខ្នាតតូចអាចឆ្លើយសំនួរនេះហើយការព្យាករណ៍ផ្អែកលើទិន្នន័យរបស់មនុស្សអាចថាការចេញផ្សាយឌីអេដូកាន់តែច្រើននៅក្នុងក្រុម 50“ ល្បែងលេងល្បែង” នឹងត្រូវបានគេសង្កេតឃើញយ៉ាងច្បាស់នៅក្នុងឌីអ័រឌីស៊ែរឌីត្រុសជាជាងឌីស្កូបឺរី។ ។ សុពលភាពនៃការបង្ហាញរង្វាន់អថេរ X +XX%% ស៊ីឌីក្នុងគំរូផ្សេងទៀតទាំងនេះនឹងគាំទ្រដល់ឧបករណ៍ប្រើប្រាស់របស់វាដែលជាគំរូនៃការសាកល្បងផូអេសអេស។
Whereas some forms of gambling clearly entail an instrumental response (e.g., slot machines), in other forms of gambling (e.g., lottery) the link between the action (purchasing the ticket, i.e., placing the bet), the cues for reward (i.e., lottery numbers) and the reward itself (the winning number and monetary payoff) is much more diffuse. Nevertheless, activation of DA during the CS-US interval may well occur. This may explain why, when the “winning number” is announced, attention is riveted as each individual lottery ball drops in succession to compose the specific sequence of digits in the winning number. Although the probability of a specific digit occurring is mathematically defined, the outcome for each individual lottery ball is binary—hit (matches the player’s number) or miss (does not match the player’s number)—and the outcome on any given trial is unknown. Such a scenario may better characterize the experience of group 50 in the present experiments, where reward was provided non-contingently but also unpredictably and the CS merely indicated the potential for reward without revealing whether it would occur on a given trial. Slot machines are more strongly linked with PG than are lottery tickets (Cox et al ។ , 2000; Bakken et al ។ , 2009), indicating an important role for instrumental factors (and immediacy) in the rewarding aspects of gambling for this population (Loba et al., 2001) ។ ទោះយ៉ាងណាដំណើរការផូវូវៀនបានយកគំរូតាមការពិសោធន៍បច្ចុប្បន្ន (រង្វាន់រង្វាន់មិនប្រាកដប្រជា +) ហាក់ដូចជាចាំបាច់ប្រសិនបើមិនមានធាតុផ្សំគ្រប់គ្រាន់នៃបទពិសោធន៍នៃល្បែង។
ទន្ទឹមនឹងកង្វះខាតតំរូវការឧបករណ៍ច្បាស់លាស់លក្ខណៈពិសេសនៃការរចនាមួយចំនួនទៀតអាចរួមចំណែកដល់គំរូនៃផលប៉ះពាល់ពិសោធន៍និងតិចតួច។ ក្រុមទាំងនោះមានភាពខុសគ្នានៅក្នុងការប៉ះពាល់ជារួមនិងភាពជាប់គ្នារវាង CS និងរង្វាន់ sucrose ។ ទោះបីជាកត្តានេះអាចរួមចំណែកដល់ការផ្លាស់ប្តូរអន្តរក្រុមក៏ដោយក៏វាមិនអាចពន្យល់បានភ្លាមៗថាហេតុអ្វីបានជាសត្វដែលមានបញ្ហា sucrose ខ្ពស់បំផុត (ក្រុម 100) បង្ហាញការប្រែលប្រួលតិចជាងក្រុម 50 ។ លើសពីនេះទៀតក្រុម 0 មិនបានទទួលការរំញោចមុនពេលប៉ះពាល់ដល់ sucrose នៅរាល់ការសាកល្បង។ ទោះបីជាវាបានរារាំងការរំពឹងទុកនៃលទ្ធផលដែលមិនសមហេតុផលក៏ដោយវាមិនបានគ្រប់គ្រងសម្រាប់វត្តមាននៃការជម្រុញមុនពេលការផ្តល់រង្វាន់ដែលមាននៅក្នុងក្រុមដទៃទៀតទាំងអស់។ ដើម្បីដោះស្រាយបញ្ហានេះការស្រាវជ្រាវនាពេលអនាគតគួរតែរួមបញ្ចូលលក្ខខណ្ឌមួយដែលសត្វទទួលបានរង្វាន់នៅរាល់ការសាកល្បងបន្ទាប់ពីការប៉ះពាល់ដោយចៃដន្យទៅនឹងការជម្រុញអព្យាក្រឹត (មានន័យថាវត្តមានរបស់វាមិនបង្ហាញពីសក្តានុពលនៃរង្វាន់) ។
Another design limitation is the potential emergence of adjunctive behavior that could influence the effects of training schedule. In the face of uncertainty, animals may develop superstitious behaviors designed to enhance perceived control and reduce uncertainty-induced DA activation (cf. Harris et al ។ , 2013) ។ ដូច្នេះវាអាចទៅរួចដែលថាទិដ្ឋភាពដែលមិនមានការត្រួតពិនិត្យនៃការរចនាពិសោធន៍អាចឱ្យសត្វអាចប៉ះប៉ូវផលប៉ះពាល់នៃកាលវិភាគម៉ាស៊ីនត្រជាក់។ ប្រសិទ្ធិភាពបែបនេះអាចចូលរួមចំណែកក្នុងការឆ្លើយតបតិចតួចនិងអថេរទៅនឹងថ្នាំអំហ្វេតាមីនក្នុងក្រុមទី 50 បន្ទាប់ពីការបណ្តុះបណ្តាលស៊ីអេស + ។ ការស្រាវជ្រាវនាពេលអនាគតគួរតែកត់ត្រាអាកប្បកិរិយាដោយឯកឯងក្រៅពីផែសច្រមុះក្នុងកំឡុងពេលហ្វឹកហាត់ដើម្បីសាកល្បងលទ្ធភាពនេះនិងគ្រប់គ្រងវាតាមស្ថិតិប្រសិនបើវាលេចចេញមក។ ដោយសារតែអាកប្បកិរិយាបែបនេះត្រូវបានគេរំពឹងថានឹងប្រឆាំងឬធ្វើឱ្យថយចុះផលប៉ះពាល់នៃភាពមិនច្បាស់លាស់នៃកាលវិភាគការឆ្លើយតបរបស់ក្បាលម៉ាស៊ីនទៅនឹងថ្នាំអំហ្វេតាមីនក្នុងក្រុម 50 គួរតែត្រូវបានពង្រឹងនៅពេលដែលវាត្រូវបានគ្រប់គ្រង (តាមនីតិវិធីឬស្ថិតិ) ។ ដូច្នេះការរចនាបច្ចុប្បន្ន (មិនមានការត្រួតពិនិត្យ) ផ្តល់នូវការសាកល្បងអភិរក្សនៃផលប៉ះពាល់នៃរង្វាន់ 50% CS + លើការចាប់យកថ្នាំអាហ្វេតាមីន។
In terms of external validity, the use of male rats also limits the generalizability of the results. The lack of a clear “punishment” condition also differs from gambling, where large monetary losses are common and exert important motivational effects (Nieuwenhuis et al ។ , 2005; Singh and Khan, 2012). The ability to accumulate reward is also absent from the present paradigm and cumulative winnings in a slot machine game have been found to interact with DA manipulations in humans (Tremblay et al ។ , 2011 ។; Smart et al ។ , 2013) ។ ស្រដៀងគ្នានេះដែរឱកាសសម្រាប់ជេកផតគឺជាភាពខុសគ្នាដ៏សំខាន់រវាងគំរូបច្ចុប្បន្ននិងល្បែងជាក់ស្តែង។
Despite these limitations, the present results suggest that 50% variable CS + reward can engage DA pathways implicated in the reinforcing effects of gambling (Fiorillo et al ។ , 2003; អេហ្វអេសអេមអេសអេសស៊ីធី XX ។). Cross-sensitization of response to AMPH following this gambling-like schedule is consistent with a pivotal role for DA in gambling and psychostimulant drug effects (Zack និង Poulos, 2009), and extends earlier studies on cross-priming of motivation to gamble by AMPH in pathological gamblers (Zack និង Poulos, 2004). The present results also indirectly suggest that modest doses of AMPH, which do not cause supra-physiological DA release, may better model brain activity in response to intermittent reward signals (i.e., during gambling) than exposure to high (i.e., binge-like) doses of stimulant drugs (cf. Vanderschuren and Pierce, 2010) ។ ការគាំទ្រដោយផ្ទាល់សម្រាប់ការឆ្លើយឆ្លងនេះអាចត្រូវបានទទួលដោយវាយតម្លៃការចេញផ្សាយឌីអេសអេដើម្បីឆ្លើយតបទៅនឹងកាលវិភាគអថេរស៊ីអេសអេសអេសអេសអេសអេសអេសអេសអេសនិងដូសអេអេអេអេអេអេអេអេអេដោយប្រើអតិសុខុមប្រាណ។
តាមទស្សនៈពិសោធន៍គំរូប៉ូវ៉ូវៀនាពេលបច្ចុប្បន្ននិងគំរូប្រតិបត្ដិការមុននៃការពង្រឹងអថេរទាំងពីរបង្ហាញឱ្យឃើញនូវផេណូទីណូដែលស្រដៀងនឹងអ្នកលេងល្បែងដែលមានរោគសាស្ត្ររបស់មនុស្ស។ ដូចនេះពួកគេបានផ្តល់នូវការបំពេញបន្ថែមដ៏មានតំលៃមួយដល់ភារកិច្ចលេងល្បែងដែលជាគំរូនៃអាកប្បកិរិយាលេងល្បែង (ជាវិធានការណ៍ពឹងផ្អែក) ប៉ុន្តែរហូតមកដល់បច្ចុប្បន្ននេះប្រើតែសត្វមានសុខភាពល្អស្មើនឹងអ្នកលេងល្បែងសង្គមរបស់មនុស្ស។ ផ្អែកលើអក្សរសិល្ប៍សត្វដែលត្រូវបានប៉ះពាល់រ៉ាំរ៉ៃចំពោះរង្វាន់អថេរអាចមានភាពខុសប្លែកគ្នានៅក្នុងភារកិច្ចទាំងនេះជាពិសេសក្នុងការឆ្លើយតបទៅនឹងថ្នាំ DA-ergic ។ ការរួមបញ្ចូលគ្នារវាងពពួកផេនឃ្វីនលេងល្បែងស៊ីសងជាមួយនឹងភារកិច្ចលេងល្បែងអាចអនុញ្ញាតឱ្យមានការអភិវឌ្ឍជាប្រព័ន្ធនៃថ្នាំសម្រាប់ការព្យាបាលភីអេសជីដែលប្រហែលជាមិនត្រូវបានបំពេញទាំងស្រុងជាមួយសត្វដែលមានសុខភាពល្អនោះទេ។ ការកែលម្អបន្ថែមទៀតនៅក្នុងរចនាបទពិសោធនិងរបបបណ្តុះបណ្តាលដូចដែលបានរៀបរាប់ខាងលើគួរតែកែលម្អការឆ្លើយឆ្លងរវាងសត្វដែលទទួលការបណ្តុះបណ្តាលនៅក្នុងគំរូនេះនិងអ្នកលេងល្បែងដែលមានរោគវិទ្យាពិតប្រាកដ។
From the clinical-sociological standpoint, the finding that exposure to 50% variable CS + reward, which closely matches the reward schedule on a commercial slot machine (Tremblay et al ។ , 2011 ។) ការផ្លាស់ប្តូរប្រព័ន្ធឌីអេសអិននៅក្នុងវិធីដែលអាចទុកចិត្តបាននិងស្ថិតស្ថេរបង្ហាញថាក្នុងករណីខ្លះសកម្មភាពលេងល្បែងដូចជាគ្រឿងញៀននៃការរំលោភបំពានអាចជា“ ភ្នាក់ងារបង្កជំងឺ” ដែលអាចបង្កឱ្យមានការញៀន។ ទោះជាយ៉ាងណាក៏ដោយទំហំផលប៉ះពាល់តិចតួចនិងភាពប្រែប្រួលខ្ពស់ក្នុងការឆ្លើយតបនឹងរង្វាន់ 50% CS + បង្ហាញថាដូចជាគ្រឿងញៀននៃការរំលោភបំពានទំនោរសម្រាប់កាលវិភាគរង្វាន់ដូចល្បែងដើម្បីជំរុញការញៀននឹងពឹងផ្អែកយ៉ាងខ្លាំងទៅលើទម្រង់ហានិភ័យមុនរបស់អ្នកលេងល្បែង។ ទោះយ៉ាងណាដើម្បីទុកឱ្យបុគ្គលដែលមានហានិភ័យខ្ពស់ប្រឈមនឹងផលប៉ះពាល់អវិជ្ជមាននៃការលេងល្បែងវាហាក់ដូចជាសមហេតុផលដែលគោលនយោបាយត្រូវបានអនុវត្តដើម្បីរារាំងការប្រើប្រាស់និងកាត់បន្ថយគ្រោះថ្នាក់ពីគ្រឿងញៀននៃការរំលោភបំពានអាចត្រូវបានពង្រីកដល់ល្បែងផងដែរ។
ទំនាស់ផលប្រយោជន៍
អ្នកនិពន្ធបានប្រកាសថាការស្រាវជ្រាវត្រូវបានធ្វើឡើងដោយគ្មានទំនាក់ទំនងពាណិជ្ជកម្មឬហិរញ្ញវត្ថុណាមួយដែលអាចត្រូវបានបកស្រាយថាជាទំនាស់ផលប្រយោជន៍ដែលអាចមាន។
ការទទួលស្គាល់
ការស្រាវជ្រាវនេះត្រូវបានផ្តល់មូលនិធិដោយជំនួយពីក្រុមប្រឹក្សាស្រាវជ្រាវវិទ្យាសាស្ត្រធម្មជាតិនិងវិស្វកម្មកាណាដាដល់លោក Paul J. Fletcher ។ យើងសូមថ្លែងអំណរគុណយ៉ាងជ្រាលជ្រៅដល់លោកស្រី Djurdja Djordjevic ដែលបានរៀបចំតួលេខ។
ឯកសារយោង
អេហ្វអេសអេម, ភី (2013) ។ ដូប៉ាមីនការលើកទឹកចិត្តនិងសារៈសំខាន់នៃការវិវត្តនៃឥរិយាបទដូចល្បែង។ Behav ។ ខួរក្បាល Res។ 256, 1-4 ។ doi: 10.1016 / j.bbr.2013.07.039
A.P.A. (2013). សៀវភៅរោគវិនិច្ឆ័យនិងស្ថិតិនៃជំងឺផ្លូវចិត្ត, អេសអិល 5។ Arlington, VA: ការបោះពុម្ពផ្សាយផ្នែកចិត្តសាស្រ្តអាមេរិច។
Bakken, I. J., Gotestam, K. G., Grawe, R. W., Wenzel, H. G., and Oren, A. (2009). Gambling behavior and gambling problems in Norway 2007. Scand. J. Psychol។ 50, 333-339 ។ doi: 10.1111 / j.1467-9450.2009.00713.x
Boileau, I., Payer, D., Chugani, B., Lobo, D. S., Houle, S., Wilson, A. A., et al. (2013). In vivo evidence for greater amphetamine-induced dopamine release in pathological gambling: a positron emission tomography study with [C]-(+)-PHNO. Mol ។ ចិត្តសាស្ត្រ doi: 10.1038/mp.2013.163. [Epub ahead of print].
Bolles, R. C. (1972). Reinforcement, expectancy, and learning. Psychol ។ Rev. ៥៥, ៦៧១–៦៧៨។ doi: 79/h394
Camp, D. M., Browman, K. E., and Robinson, T. E. (1994). The effects of methamphetamine and cocaine on motor behavior and extracellular dopamine in the ventral striatum of Lewis versus Fischer 344 rats. ខួរក្បាល Res. 668, 180–193. doi: 10.1016/0006-8993(94)90523-1
Conversano, C., Marazziti, D., Carmassi, C., Baldini, S., Barnabei, G., and Dell’Osso, L. (2012). Pathological gambling: a systematic review of biochemical, neuroimaging, and neuropsychological findings. Harv. Rev. Psychiatry 20, 130-148 ។ doi: 10.3109 / 10673229.2012.694318
Cox, B. J., Kwong, J., Michaud, V., and Enns, M. W. (2000). Problem and probable pathological gambling: considerations from a community survey. អាច។ J. ចិត្តសាស្ត្រ។ 45, 548-553 ។
Dalley, J. W., Theobald, D. E., Eagle, D. M., Passetti, F., and Robbins, T. W. (2002). Deficits in impulse control associated with tonically-elevated serotonergic function in rat prefrontal cortex. Neuropsychopharmacology 26, 716–728. doi: 10.1016/S0893-133X(01)00412-2
Everitt, BJ និង Robbins, TW (2005) ។ ប្រព័ន្ធសរសៃប្រសាទនៃការពង្រឹងសម្រាប់ការញៀនគ្រឿងញៀន: ពីសកម្មភាពទៅទម្លាប់ដើម្បីបង្ខិតបង្ខំ។ ណត។ Neurosci។ 8, 1481-1489 ។ doi: 10.1038 / nn1579
Fiorillo, ស៊ីឌី, Tobler, PN និង Schultz, W. (2003) ។ ការបម្លែងកូដប្រូតូកូលនៃប្រូតេអ៊ីនដូប៉ាមីន។ វិទ្យាសាស្រ្ត 299, 1898-1902 ។ doi: 10.1126 / science.1077349
Flagel, S. B., Watson, S. J., Akil, H., and Robinson, T. E. (2008). Individual differences in the attribution of incentive salience to a reward-related cue: influence on cocaine sensitization. Behav ។ ខួរក្បាល Res។ 186, 48-56 ។ doi: 10.1016 / j.bbr.2007.07.022
Fletcher, P. J., Tenn, C. C., Rizos, Z., Lovic, V., and Kapur, S. (2005). Sensitization to amphetamine, but not PCP, impairs attentional set shifting: reversal by a D1 receptor agonist injected into the medial prefrontal cortex. ពេទ្យចិត្តសាស្ត្រ (Berl ។ ) 183, 190–200. doi: 10.1007/s00213-005-0157-6
Flores, G., Wood, G. K., Barbeau, D., Quirion, R., and Srivastava, L. K. (1998). Lewis and Fischer rats: a comparison of dopamine transporter and receptors levels. ខួរក្បាល Res. 814, 34–40. doi: 10.1016/S0006-8993(98)01011-7
Frascella, J., Potenza, M. N., Brown, L. L., and Childress, A. R. (2010). Shared brain vulnerabilities open the way for nonsubstance addictions: carving addiction at a new joint? Ann ។ ញូអេដអាដ។ Sci។ 1187, 294-315 ។ doi: 10.1111 / j.1749-6632.2009.05420.x
Grimm, J. W., Buse, C., Manaois, M., Osincup, D., Fyall, A., and Wells, B. (2006). Time-dependent dissociation of cocaine dose-response effects on sucrose craving and locomotion. Behav ។ Pharmacol. 17, 143–149. doi: 10.1097/01.fbp.0000190686.23103.f8
Haile, C. N., Hiroi, N., Nestler, E. J., and Kosten, T. A. (2001). Differential behavioral responses to cocaine are associated with dynamics of mesolimbic dopamine proteins in Lewis and Fischer 344 rats. តំណភ្ជាប់កោសិកាសរសៃប្រសាទ 41, 179-190 ។ doi: 10.1002 / syn.1073
Harris, J. A., Andrew, B. J., and Kwok, D. W. (2013). Magazine approach during a signal for food depends on Pavlovian, not instrumental, conditioning. J. Exp ។ ចិត្ត។ ជីវចល។ ប៊ីវ។ ដំណើរការ។។ 39, 107-116 ។ doi: 10.1037 / a0031315
Howell, D. C. (1992). Statistical Methods for Psychology. Boston, MA: Duxbury.
Ito, R., Dalley, J. W., Robbins, T. W., and Everitt, B. J. (2002). Dopamine release in the dorsal striatum during cocaine-seeking behavior under the control of a drug-associated cue. J. Neurosci។ 22, 6247-6253 ។
Jiao, X., Pare, W. P., and Tejani-Butt, S. (2003). Strain differences in the distribution of dopamine transporter sites in rat brain. Prog ។ Neuropsychopharmacol ។ Biol ។ ចិត្តសាស្រ្ត 27, 913–919. doi: 10.1016/S0278-5846(03)00150-7
Koob, G. F., and Le Moal, M. (2008). Review. Neurobiological mechanisms for opponent motivational processes in addiction. ទីក្រុង Philos ។ Trans ។ R. សុ។ លីន។ ខប៊ីល។ ឌី. 363, 3113–3123. doi: 10.1098/rstb.2008.0094
Kosten, T. A., Miserendino, M. J., Chi, S., and Nestler, E. J. (1994). Fischer and Lewis rat strains show differential cocaine effects in conditioned place preference and behavioral sensitization but not in locomotor activity or conditioned taste aversion. J. Pharmacol ។ Exp ។ ពួកវា។ 269, 137-144 ។
Kosten, T. A., Zhang, X. Y., and Haile, C. N. (2007). Strain differences in maintenance of cocaine self-administration and their relationship to novelty activity responses. Behav ។ Neurosci។ 121, 380-388 ។ doi: 10.1037 / 0735-7044.121.2.380
Leeman, R. F., and Potenza, M. N. (2012). Similarities and differences between pathological gambling and substance use disorders: a focus on impulsivity and compulsivity. ពេទ្យចិត្តសាស្ត្រ (Berl ។ ) 219, 469–490. doi: 10.1007/s00213-011-2550-7
Leshner, A. I. (1997). Addiction is a brain disease, and it matters. វិទ្យាសាស្រ្ត 278, 45-47 ។ doi: 10.1126 / science.278.5335.45
Loba, P., Stewart, S. H., Klein, R. M., and Blackburn, J. R. (2001). Manipulations of the features of standard video lottery terminal (VLT) games: effects in pathological and non-pathological gamblers. J. Gambl ។ សិក្សា។. 17, 297–320. doi: 10.1023/A:1013639729908
Martinez, D., Narendran, R., Foltin, R. W., Slifstein, M., Hwang, D. R., Broft, A., et al. (2007). Amphetamine-induced dopamine release: markedly blunted in cocaine dependence and predictive of the choice to self-administer cocaine. Am ។ J. ចិត្តសាស្រ្ត 164, 622-629 ។ doi: 10.1176 / appi.ajp.164.4.622
Mateo, Y., Lack, C. M., Morgan, D., Roberts, D. C., and Jones, S. R. (2005). Reduced dopamine terminal function and insensitivity to cocaine following cocaine binge self-administration and deprivation. Neuropsychopharmacology 30, 1455-1463 ។ doi: 10.1038 / sj.npp.1300687
Nestler, EJ (2001) ។ មូលដ្ឋានម៉ូលេគុលនៃប្លាស្ទិកដែលមានមូលដ្ឋានលើការញៀនយូរ។ ណត។ Rev. Neurosci។ 2, 119-128 ។ doi: 10.1038 / 35053570
Nieuwenhuis, S., Heslenfeld, D. J., von Geusau, N. J., Mars, R. B., Holroyd, C. B., and Yeung, N. (2005). Activity in human reward-sensitive brain areas is strongly context dependent. Neuroimage 25, 1302-1309 ។ doi: 10.1016 / j.neuroimage.2004.12.043
Pierce, R. C., and Kalivas, P. W. (1997). A circuitry model of the expression of behavioral sensitization to amphetamine-like psychostimulants. ខួរក្បាល Res ។ ខួរក្បាល Res ។ Rev. 25, 192. doi: 10.1016/S0165-0173(97)00021-0
Robbins, T. W., and Everitt, B. J. (1999). Drug addiction: bad habits add up. ធម្មជាតិ 398, 567-570 ។ doi: 10.1038 / 19208
Robinson, T. E., Becker, J. B., and Presty, S. K. (1982). Long-term facilitation of amphetamine-induced rotational behavior and striatal dopamine release produced by a single exposure to amphetamine: sex differences. ខួរក្បាល Res. 253, 231–241. doi: 10.1016/0006-8993(82)90690-4
Robinson, TE, និង Berridge, KC (1993) ។ មូលដ្ឋានសរសៃប្រសាទនៃការលោភលន់គ្រឿងញៀន: ទ្រឹស្តីលើកទឹកចិត្ត - រំញោចអារម្មណ៍នៃការញៀន។ ខួរក្បាល Res ។ ខួរក្បាល Res ។ Rev. 18, 247–291. doi: 10.1016/0165-0173(93)90013-P
Robinson, TE, និង Berridge, KC (2001) ។ ការលើកទឹកចិត្ត - ការគិតនិងការញៀន។ ញៀន 96, 103-114 ។ doi: 10.1046 / j.1360-0443.2001.9611038.x
Singer, B. F., Scott-Railton, J., and Vezina, P. (2012). Unpredictable saccharin reinforcement enhances locomotor responding to amphetamine. Behav ។ ខួរក្បាល Res។ 226, 340-344 ។ doi: 10.1016 / j.bbr.2011.09.003
Singh, V., and Khan, A. (2012). Decision making in the reward and punishment variants of the iowa gambling task: evidence of “foresight” or “framing”? មុខ។ Neurosci។ 6: 107 ។ doi: 10.3389 / fnins.2012.00107 ។
Skinner, B. F. (1953). វិទ្យាសាស្រ្តនិងឥរិយាបថមនុស្ស។. New York, NY: Free Press.
Smart, K., Desmond, R. C., Poulos, C. X., and Zack, M. (2013). Modafinil increases reward salience in a slot machine game in low and high impulsivity pathological gamblers. Neuropharmacology 73, 66-74 ។ doi: 10.1016 / j.neuropharm.2013.05.015
Tremblay, A. M., Desmond, R. C., Poulos, C. X., and Zack, M. (2011). Haloperidol modifies instrumental aspects of slot machine gambling in pathological gamblers and healthy controls. អ្នកញៀន។ Biol។ 16, 467-484 ។ doi: 10.1111 / j.1369-1600.2010.00208.x
Vanderschuren, L. J., and Kalivas, P. W. (2000). Alterations in dopaminergic and glutamatergic transmission in the induction and expression of behavioral sensitization: a critical review of preclinical studies. ពេទ្យចិត្តសាស្ត្រ (Berl ។ ) 151, 99-120 ។ doi: 10.1007 / s002130000493
Vanderschuren, L. J., and Pierce, R. C. (2010). Sensitization processes in drug addiction. Curr. Top. Behav. Neurosci. 3, 179–195. doi: 10.1007/7854_2009_21
Vanderschuren, L. J., Schoffelmeer, A. N., Mulder, A. H., and De Vries, T. J. (1999). Dopaminergic mechanisms mediating the long-term expression of locomotor sensitization following pre-exposure to morphine or amphetamine. ពេទ្យចិត្តសាស្ត្រ (Berl ។ ) 143, 244-253 ។ doi: 10.1007 / s002130050943
Verdejo-Garcia, A., Lawrence, A. J., and Clark, L. (2008). Impulsivity as a vulnerability marker for substance-use disorders: review of findings from high-risk research, problem gamblers and genetic association studies. Neurosci ។ Biobehav ។ Rev។ 32, 777-810 ។ doi: 10.1016 / j.neubiorev.2007.11.003
Vezina, P. (2004). Sensitization of midbrain dopamine neuron reactivity and the self-administration of psychomotor stimulant drugs. Neurosci ។ Biobehav ។ Rev។ 27, 827-839 ។ doi: 10.1016 / j.neubiorev.2003.11.001
Volkow, N. D., Wang, G. J., Fowler, J. S., Logan, J., Gatley, S. J., Hitzemann, R., et al. (1997). Decreased striatal dopaminergic responsiveness in detoxified cocaine-dependent subjects. ធម្មជាតិ 386, 830-833 ។ doi: 10.1038 / 386830a0
Volkow, ND, Wang, GJ, Fowler, JS, and Tomasi, D. (2012) ។ សៀគ្វីញៀននៅក្នុងខួរក្បាលមនុស្ស។ Annu. Rev. Pharmacol. Toxicol. 52, 321–336. doi: 10.1146/annurev-pharmtox-010611-134625
Winer, B. (ed.). (1971). Statistical Principles in Experimental Design. New York, NY: McGraw-Hill.
Zack, M., and Poulos, C. X. (2004). Amphetamine primes motivation to gamble and gambling-related semantic networks in problem gamblers. Neuropsychopharmacology 29, 195-207 ។ doi: 10.1038 / sj.npp.1300333
Zack, M. , និង Poulos, CX (2009) ។ តួនាទីប៉ារ៉ាឡែលសម្រាប់ដូប៉ាមីនក្នុងការលេងល្បែងស៊ីសងរោគសាស្ត្រនិងការញៀនផ្លូវចិត្ត។ Curr ។ ការប្រើប្រាស់គ្រឿងញៀន Rev.។ 2, 11-25 ។ doi: 10.2174 / 1874473710902010011
Zamudio, S., Fregoso, T., Miranda, A., De La Cruz, F., and Flores, G. (2005). Strain differences of dopamine receptor levels and dopamine related behaviors in rats. ខួរក្បាល Res ។ Bull. 65, 339–347. doi: 10.1016/j.brainresbull.2005.01.009
Keywords: pathological gambling, sensitization, amphetamine, dopamine, uncertainty
Citation: Zack M, Featherstone RE, Mathewson S and Fletcher PJ (2014) Chronic exposure to a gambling-like schedule of reward predictive stimuli can promote sensitization to amphetamine in rats. មុខ។ Behav ។ Neurosci. 8៖ 36 ។ doi: 10.3389 / fnbeh.2014.00036 ។
បានទទួល: 01 November 2013; បានទទួល: 23 ខែមករា 2014;
Published online: 11 February 2014.
កែសម្រួលដោយ:
Bryan F. តារាចម្រៀង។, សាកលវិទ្យាល័យមីឈីហ្គែនសហរដ្ឋអាមេរិក។
ពិនិត្យឡើងវិញដោយ:
Louk Vanderschuren, University of Utrecht, Netherlands
Ruud Van Den Bos, Radboud University Nijmegen, Netherlands
លោក Patrick Anselme ។, សាកលវិទ្យាល័យ Li ofge, បែលហ្ស៊ិក។
Copyright © 2014 Zack, Featherstone, Mathewson and Fletcher. This is an open-access article distributed under the terms of the អាជ្ញាប័ណ្ណគុណលក្ខណៈ Creative Commons Attribution (CC BY)។ ការប្រើការចែកចាយឬការបន្តពូជនៅក្នុងវេទិកាផ្សេងទៀតត្រូវបានអនុញ្ញាតដែលបានផ្ដល់ឱ្យអ្នកនិពន្ធដើមឬអ្នកផ្តល់អាជ្ញាប័ណ្ណត្រូវបានបញ្ចូលហើយថាការបោះពុម្ពផ្សាយដើមនៅក្នុងទិនានុប្បវត្តិនេះត្រូវបានលើកឡើងស្របតាមការអនុវត្តការសិក្សាដែលទទួលយក។ គ្មានការអនុញ្ញាតការចែកចាយឬការផលិតត្រូវបានអនុញ្ញាតិដែលមិនអនុលោមតាមលក្ខខណ្ឌទាំងនេះ។
*Correspondence: Martin Zack, Cognitive Psychopharmacology Laboratory, Neuroscience Department, Centre for Addiction and Mental Health, 33 Russell Street, Toronto, ON M5S 2S1, Canada e-mail: [អ៊ីមែលការពារ]