Dev Rev. 2008; 28(1): 62-77. doi: 10.1016 / j.dr.2007.08.003
Abstrakcja
Okres dojrzewania to okres rozwojowy charakteryzujący się suboptymalnymi decyzjami i działaniami, które powodują zwiększoną częstość niezamierzonych urazów i przemocy, nadużywania alkoholu i narkotyków, niezamierzonej ciąży i chorób przenoszonych drogą płciową. Tradycyjne neurobiologiczne i kognitywne wyjaśnienia zachowania nastolatków nie uwzględniają nieliniowych zmian w zachowaniu obserwowanych w okresie dojrzewania, w stosunku do dzieciństwa i dorosłości. Przegląd ten zapewnia biologicznie wiarygodną konceptualizację mechanizmów neuronalnych leżących u podstaw tych nieliniowych zmian w zachowaniu, jako podwyższoną reakcję na bodźce, podczas gdy kontrola impulsów jest nadal stosunkowo niedojrzała w tym okresie. Najnowsze badania obrazowe na ludziach i badania na zwierzętach stanowią biologiczną podstawę tego poglądu, co sugeruje zróżnicowany rozwój limbicznych systemów nagrody w stosunku do odgórnych systemów kontroli w okresie dojrzewania w stosunku do dzieciństwa i dorosłości. Ten wzorzec rozwojowy może się nasilić u młodzieży z predyspozycjami do podejmowania ryzyka, zwiększając ryzyko złych wyników.
Według National Center for Health Statistics każdego roku w Stanach Zjednoczonych odnotowuje się ponad 13,000 nastolatków. Około 70% tych zgonów wynika z wypadków samochodowych, niezamierzonych obrażeń, zabójstw i samobójstw (Eaton i in., 2006). Wyniki Krajowego Badania Zachowań Młodzieży 2005 (YRBS) pokazują, że młodzież angażuje się w zachowania, które zwiększają prawdopodobieństwo śmierci lub choroby poprzez prowadzenie pojazdu po piciu lub bez pasa bezpieczeństwa, noszenie broni, używanie nielegalnych substancji i uprawianie seksu bez zabezpieczenia skutkujące niezamierzonymi ciążami i chorobami przenoszonymi drogą płciową, w tym zakażeniem HIV (Eaton i in., 2006). Statystyki te podkreślają znaczenie zrozumienia ryzykownych wyborów i działań u młodzieży.
Postulowano wiele hipotez poznawczych i neurobiologicznych, dlaczego nastolatki angażują się w suboptymalne zachowania związane z wyborem. W niedawnym przeglądzie literatury dotyczącej rozwoju ludzkiego mózgu u młodzieży, Yurgelun-Todd (2007) sugeruje, że rozwój poznawczy w latach młodzieńczych wiąże się z coraz większą skutecznością zdolności kontroli poznawczej. Wydajność ta jest opisana jako zależna od dojrzewania kory przedczołowej, o czym świadczy zwiększona aktywność w ogniskowych obszarach przedczołowych (Rubia i in., 2000; Tamm, Menon i Reiss, 2002) i zmniejszona aktywność w nieistotnych regionach mózgu (Brown i wsp., 2005; Durston i in., 2006).
Ten ogólny wzorzec poprawy kontroli poznawczej z dojrzewaniem kory przedczołowej sugeruje liniowy wzrost rozwoju od dzieciństwa do dorosłości. Jednak suboptymalne wybory i działania obserwowane w okresie dojrzewania stanowią nieliniową zmianę w zachowaniu, którą można odróżnić od dzieciństwa i dorosłości, o czym świadczy Narodowe Centrum Statystyki Zdrowia dotyczące zachowań i śmiertelności wśród młodzieży. Jeśli kontrola poznawcza i niedojrzała kora przedczołowa były podstawą suboptymalnego zachowania, dzieci powinny wyglądać niezwykle podobnie lub nawet gorzej niż młodzież, biorąc pod uwagę ich mniej rozwiniętą korę przedczołową i zdolności poznawcze. Tak więc sama niedojrzała funkcja przedczołowa nie może wyjaśnić zachowania nastolatków.
Dokładna konceptualizacja zmian poznawczych i neurobiologicznych w okresie dojrzewania musi traktować dorastanie jako przejściowy okres rozwojowy (Włócznia, 2000), zamiast pojedynczej migawki w czasie (Casey, Tottenham, Liston i Durston, 2005). Innymi słowy, aby zrozumieć ten okres rozwoju, przejścia do i poza okresem dojrzewania są konieczne, aby odróżnić odrębne atrybuty tego etapu rozwoju. Ustalenie trajektorii rozwojowych dla procesów poznawczych i neuronowych jest niezbędne do scharakteryzowania tych przejść i ograniczenia interpretacji dotyczących zmian w zachowaniu w tym okresie. Na poziomie poznawczym lub behawioralnym nastolatki charakteryzują się impulsywnością (tj. Brakiem kontroli poznawczej) i podejmowaniem ryzyka przy użyciu tych konstruktów używanych synonimicznie i bez uznania dla różnych trajektorii rozwoju każdego z nich. Na poziomie neurobiologicznym obrazowanie na ludziach i badania na zwierzętach sugerują odmienne podstawy neurobiologiczne i trajektorie rozwojowe dla systemów neuronowych, które leżą u podstaw tych oddzielnych konstrukcji kontroli impulsów i ryzykownych decyzji.
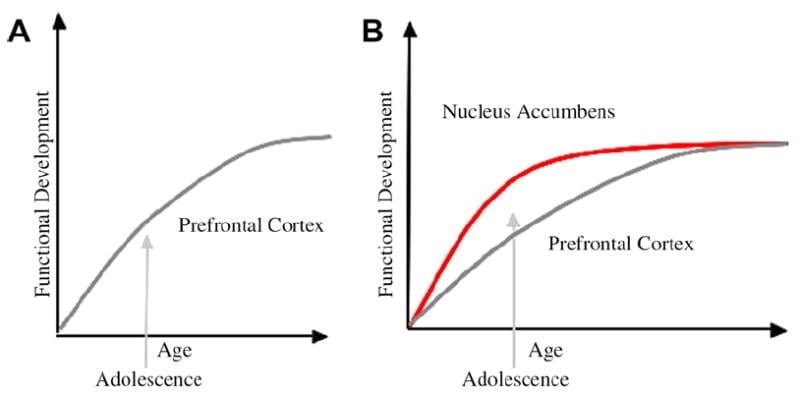
Opracowaliśmy neurobiologiczny model rozwoju młodzieży w ramach tego modelu, który opiera się na modelach gryzoni (Laviola, Adriani, Terranova i Gerra, 1999; Włócznia, 2000) i ostatnie badania obrazowe okresu dojrzewania (Ernst i in., 2005; Galvan, Hare, Voss, Glover i Casey, 2007; Galvan i in., 2006). Rys. 1 poniżej przedstawia ten model. Po lewej stronie znajduje się tradycyjna charakterystyka okresu dojrzewania jako związana prawie wyłącznie z niedojrzałością kory przedczołowej. Po prawej stronie znajduje się nasz proponowany model neurobiologiczny, który ilustruje, w jaki sposób należy rozpatrywać limbiczne podkorowe i przedczołowe odgórne obszary kontrolne. Rysunek ilustruje różne trajektorie rozwojowe dla tych systemów, z układami limbicznymi rozwijającymi się wcześniej niż przedczołowe obszary kontrolne. Zgodnie z tym modelem osobnik jest bardziej obciążony przez dojrzałe funkcjonalnie regiony limbiczne w okresie dojrzewania (tj. Brak równowagi limbicznej w stosunku do kontroli przedczołowej), w porównaniu z dziećmi, dla których te układy (tj. Limbiczne i przedczołowe) nadal się rozwijają; iw porównaniu do dorosłych, dla których te systemy są w pełni dojrzałe. Ta perspektywa stanowi podstawę dla nieliniowych przesunięć w zachowaniu w trakcie rozwoju, z powodu wcześniejszego dojrzewania tego limbicznego w stosunku do mniej dojrzałego odgórnego obszaru przedczołowego. Dzięki rozwojowi i doświadczeniu funkcjonalna łączność między tymi regionami zapewnia mechanizm odgórnej kontroli tych regionów (Hare, Voss, Glover i Casey, 2007a). Ponadto model pogodzi sprzeczność statystyk zdrowotnych o ryzykownych zachowaniach w okresie dojrzewania, z wnikliwą obserwacją przez Reyna i Farley (2006) że młodzież jest w stanie rozumieć i rozumieć ryzyko zachowań, w które się angażuje. Zgodnie z naszym modelem, w sytuacjach emocjonalnie istotnych układ limbiczny wygrywa z systemami kontroli, biorąc pod uwagę jego dojrzałość w stosunku do przedczołowego systemu kontroli. Dowody z badań behawioralnych i badań obrazowych u ludzi w celu wsparcia tego modelu są dostarczane w kontekście działań w kontekstach nagradzających i emocjonalnych (Galvan i in., 2006, 2007; Hare, Voss, Glover i Casey, 2007b; Hare i in., 2007a). Ponadto spekulujemy, dlaczego mózg może się rozwijać w ten sposób i dlaczego niektóre nastolatki mogą być bardziej narażone na podejmowanie nieoptymalnych decyzji prowadzących do gorszych wyników długoterminowych (Galvan i in., 2007; Hare i in., 2007b).
Sugeruje się, że tradycyjne wyjaśnienie zachowania młodzieży jest spowodowane przedłużającym się rozwojem kory przedczołowej (A). Nasz model bierze pod uwagę rozwój kory przedczołowej wraz z podkorowymi regionami limbicznymi (np. Jądro półleżące), które biorą udział w ryzykownych wyborach i działaniach (B).
Rozwój zachowań ukierunkowanych na cel
Podstawą rozwoju poznawczego jest zdolność do tłumienia nieodpowiednich myśli i działań na rzecz tych, które są ukierunkowane na cel, zwłaszcza w obecności atrakcyjnych zachęt (Casey, Galvan i Hare, 2005; Casey i in., 2000b; Casey, Thomas, David-son, Kunz i Franzen, 2002a; Casey, Tottenham i Fossella, 2002b). Szereg klasycznych badań rozwojowych wykazało, że zdolność ta rozwija się w dzieciństwie i okresie dojrzewania (Etui, 1972; Flavell, Feach i Chinsky, 1966; Keating & Bobbitt, 1978; Pascual-Leone, 1970). Kilku teoretyków twierdziło, że rozwój poznawczy jest spowodowany wzrostem szybkości i wydajności przetwarzania, a nie wzrostem zdolności umysłowych (np. Bjorkland, 1985; Bjorkland, 1987; Etui, 1985). Inni teoretycy uwzględnili konstrukcję procesów „hamujących” w swoim opisie rozwoju poznawczego (Harnishfeger & Bjorkland, 1993). Zgodnie z tym stwierdzeniem niedojrzałe poznanie charakteryzuje się podatnością na zakłócenia ze strony konkurencyjnych źródeł, które muszą być tłumione (np. Brainerd & Reyna, 1993; Casey, Thomas, Davidson, Kunz i Franzen, 2002a; Dempster, 1993; Diamond, 1985; Munakata i Yerys, 2001). Tak więc zachowanie ukierunkowane na cel wymaga kontroli impulsów lub opóźnienia gratyfikacji w celu optymalizacji wyników, a zdolność ta wydaje się dojrzewać w dzieciństwie i okresie dojrzewania.
Zachowanie nastolatków zostało opisane jako impulsywne i ryzykowne, niemal synonimicznie, jednak konstrukty te opierają się na różnych procesach poznawczych i neuronalnych, które sugerują odmienne konstrukcje o różnych trajektoriach rozwojowych. W szczególności przegląd literatury sugeruje, że impulsywność zmniejsza się wraz z wiekiem w dzieciństwie i okresie dojrzewania (Casey i in., 2002a; Casey, Galvan i in., 2005; Galvan i in., 2007) i wiąże się z przedłużającym się rozwojem kory przedczołowej (Casey, Galvan i in., 2005), chociaż istnieją różnice w stopniu, w jakim dana osoba jest impulsywna lub nie, niezależnie od wieku.
W przeciwieństwie do kontroli impulsowej / poznawczej, podejmowanie ryzyka wydaje się zwiększać w okresie dorastania w stosunku do dzieciństwa i dorosłości i jest związane z systemami podkorowymi, o których wiadomo, że biorą udział w ocenie nagród. Badania obrazowe u ludzi, które zostaną poddane przeglądowi, sugerują wzrost aktywacji podkorowej (np. Półleżącej) podczas podejmowania ryzykownych wyborów (Kuhnen i Knutson, 2005; Matthews i in., 2004; Montague i Berns, 2002), który jest przesadzony u nastolatków, w stosunku do dzieci i dorosłych (Ernst i in., 2005; Galvan i in., 2006). Odkrycia te sugerują różne trajektorie zachowania opartego na nagradzaniu lub zachętach, z wcześniejszym rozwojem tych systemów w stosunku do systemów kontroli, które wykazują przedłużony i liniowy przebieg rozwoju, w kategoriach nadpisywania niewłaściwych wyborów i działań na rzecz tych ukierunkowanych na cel.
Dowody z badań neuroobrazowania rozwoju mózgu człowieka
Niedawne badania nad rozwojem mózgu nastolatków opierają się na postępach w metodach neuroobrazowania, które można łatwo stosować w rozwoju populacji ludzkich. Metody te opierają się na metodach obrazowania metodą rezonansu magnetycznego (MRI) (patrz Rys. 2) i obejmują: strukturalny MRI, który jest używany do pomiaru wielkości i kształtu struktur; funkcjonalny MRI, który jest używany do pomiaru wzorców aktywności mózgu; i obrazowanie dyfuzyjnego tensora (DTI), które jest stosowane do indeksowania połączeń włókien włóknistych. Dowody na nasz rozwojowy model współzawodnictwa między regionami korowymi i podkorowymi są wspierane przez niedojrzałą strukturalną i funkcjonalną łączność mierzoną odpowiednio przez DTI i fMRI.

Badania MRI nad rozwojem ludzkiego mózgu
W kilku badaniach wykorzystano strukturalny MRI do odwzorowania anatomicznego przebiegu normalnego rozwoju mózgu (patrz przegląd przez Durston i in., 2001). Chociaż całkowity rozmiar mózgu jest w przybliżeniu 90% jego dorosłego rozmiaru do szóstego roku życia, szare i podskładniki istoty białej mózgu nadal podlegają dynamicznym zmianom w okresie dojrzewania. Dane z ostatnich badań podłużnego MRI wskazują, że objętość istoty szarej ma odwrócony kształt litery U, z większą zmiennością regionalną niż istota biała (Giedd, 2004; Gogtay i in., 2004; Sowell i in., 2003; Sowell, Thompson i Toga, 2004). Ogólnie rzecz biorąc, regiony zachowujące podstawowe funkcje, takie jak układy motoryczne i czuciowe, dojrzewają najwcześniej; obszary asocjacji wyższego rzędu, które integrują te podstawowe funkcje, dojrzewają później (Gogtay i in., 2004; Sowell, Thompson i Toga, 2004). Na przykład badania z wykorzystaniem pomiarów MRI pokazują, że utrata istoty szarej korowej występuje najwcześniej w pierwotnych obszarach czuciowo-ruchowych, a najpóźniej w grzbietowo-bocznych przedczołowych i bocznych kory skroniowej (Gogtay i in., 2004). Ten wzorzec jest zgodny z nieludzkimi badaniami naczelnych i pośmiertnych ludzi, które pokazują, że kora przedczołowa jest jednym z ostatnich regionów mózgu dojrzałych (Bourgeois, Goldman-Rakic i Rakic, 1994; Huttenlocher, 1979). W przeciwieństwie do istoty szarej, objętość istoty białej wzrasta w przybliżeniu liniowo, wzrastając przez cały okres rozwoju aż do dorosłości (Gogtay i in., 2004). Zmiany te prawdopodobnie odzwierciedlają trwającą mielinizację aksonów przez oligodendrocyty wzmacniające przewodnictwo neuronalne i komunikację.
Chociaż mniej uwagi poświęcono regionom podkorowym podczas badania zmian strukturalnych, niektóre z największych zmian w mózgu w trakcie rozwoju są widoczne w tych regionach, szczególnie w zwojach podstawy mózgu (Sowell i in., 1999, Patrz Rys. 3), a zwłaszcza u mężczyzn (Giedd i in., 1996). Zmiany rozwojowe w objętości strukturalnej w zwojach podstawy i obszarach przedczołowych są interesujące w świetle znanych procesów rozwojowych (np. Arborizacja dendrytyczna, śmierć komórek, przycinanie synaptyczne, mielinizacja), które występują w dzieciństwie i okresie dojrzewania. Procesy te pozwalają na precyzyjne dostrojenie i wzmocnienie połączeń między regionami przedczołowymi i podkorowymi z rozwojem i uczeniem się, które mogą pokrywać się z większą kontrolą poznawczą. Jak te zmiany strukturalne odnoszą się do zmian poznawczych? Szereg badań dotyczyło dojrzewania strukturalnego płata czołowego i funkcji poznawczych z wykorzystaniem środków neuropsychologicznych i poznawczych (np. Sowell i in., 2003). W szczególności odnotowano powiązania między objętością regionów przedczołowych na poziomie przedczołowym i zwojami podstawnymi MRI a pomiarami kontroli poznawczej (tj. Zdolność do obejścia niewłaściwej odpowiedzi na korzyść innej lub do tłumienia uwagi w kierunku nieistotnego atrybutu bodźca na korzyść odpowiedniego atrybutu bodźca (Casey, Trainor i in., 1997). Odkrycia te sugerują, że zmiany poznawcze znajdują odzwierciedlenie w strukturalnych zmianach w mózgu i podkreślają znaczenie rozwoju podkorowego (jądra podstawnego), jak również korowego (np. Kory przedczołowej).

Badania DTI nad rozwojem ludzkiego mózgu
Analizy morfometrii oparte na MRI sugerują, że połączenia korowe są dopracowywane poprzez eliminację nadmiaru synaps i wzmocnienie odpowiednich powiązań z rozwojem i doświadczeniem. Ostatnie postępy w technologii rezonansu magnetycznego, takie jak DTI, stanowią potencjalne narzędzie do badania roli określonych układów istoty białej w rozwoju mózgu i zachowaniu z większą szczegółowością. Istotne dla tego artykułu są badania neuroobrazowe, które powiązały rozwój dróg włóknistych z poprawą zdolności poznawczych. W szczególności wykazano związki między pomiarami istoty białej przedczołowej opartymi na DTI a kontrolą poznawczą u dzieci. W jednym z badań rozwój tej zdolności był dodatnio skorelowany z przewodami włókien przedczołowo-ciemieniowych (Nagy, Westerberg i Klingberg, 2004) zgodne z funkcjonalnymi badaniami neuroobrazowymi wykazującymi zróżnicowaną rekrutację tych regionów u dzieci w stosunku do dorosłych.
Stosując podobne podejście, Liston i in. (2005) wykazali, że układy istoty białej między zwojami przedczołowo-podstawnymi a przewodami włóknistymi -posterior nadal rozwijają się w dzieciństwie do wieku dorosłego, ale tylko te odcinki między korą przedczołową i zwojami podstawnymi są skorelowane z kontrolą impulsów, jak zmierzono na podstawie wyników na go / nogo zadanie. Przedczołowe odcinki włókien zostały określone przez regiony zainteresowania zidentyfikowane w badaniu fMRI przy użyciu tego samego zadania. W obu badaniach rozwojowych DTI, pomiary przewodu pokarmowego były skorelowane z rozwojem, ale specyficzność poszczególnych dróg włóknistych z wydajnością poznawczą wykazano poprzez oddzielenie danego przewodu (Liston i in., 2005) lub zdolności poznawcze (Nagy i in., 2004). Odkrycia te podkreślają znaczenie badania nie tylko zmian regionalnych, ale i związanych z obwodami, gdy twierdzimy o zależnych od wieku zmianach w neuronalnych substratach rozwoju poznawczego.
Funkcjonalne badania MRI behawioralnego i rozwoju mózgu
Chociaż zmiany strukturalne mierzone za pomocą MRI i DTI są związane ze zmianami behawioralnymi podczas rozwoju, bardziej bezpośrednim podejściem do badania związku struktura-funkcja jest zmierzenie zmian w mózgu i zachowanie jednocześnie, jak w przypadku fMRI. Zdolność do pomiaru zmian funkcjonalnych w rozwijającym się mózgu za pomocą rezonansu magnetycznego ma znaczący potencjał w dziedzinie nauki o rozwoju. W kontekście obecnego artykułu fMRI zapewnia środki do ograniczania interpretacji zachowań młodzieży. Jak stwierdzono wcześniej, uważa się, że rozwój kory przedczołowej odgrywa ważną rolę w dojrzewaniu wyższych zdolności poznawczych, takich jak podejmowanie decyzji i kontrola poznawcza (Casey, Tottenham i Fossella 2002b; Casey, Trainor i in., 1997). Wykorzystano wiele paradygmatów wraz z fMRI do oceny neurobiologicznych podstaw tych umiejętności, w tym zadań flankera, Stroopa i go / nogo (Casey, Castellanos i in., 1997; Casey, Giedd i Thomas, 2000a; Durston i in., 2003). Podsumowując, badania te pokazują, że dzieci podczas rekrutacji wykonują te różne, ale często większe, bardziej rozproszone regiony przedczołowe niż dorośli. Wzorzec aktywności w regionach mózgu kluczowych dla wykonywania zadań (tj. Korelujących z wydajnością poznawczą) staje się bardziej ogniskowy lub dopracowany wraz z wiekiem, podczas gdy regiony nieskorelowane z wykonywaniem zadań zmniejszają się wraz z wiekiem. Ten wzór zaobserwowano w obu przekrojach (Brown i wsp., 2005) i badania podłużne (Durston i in., 2006) i przez różne paradygmaty. Chociaż badania neuroobrazowe nie mogą definitywnie scharakteryzować mechanizmu takich zmian rozwojowych (np. Arborizacja dendrytyczna, przycinanie synaptyczne), wyniki odzwierciedlają rozwój wewnątrz i udoskonalenie projekcji do i z, aktywowanych regionów mózgu z dojrzewaniem i sugerują, że zmiany te zachodzą przez dłuższy czas okres czasu (Brown i wsp., 2005; Bunge, Dudukovic, Thomason, Vaidya i Gabrieli, 2002; Casey, Trainor i in., 1997; Casey i in., 2002a; Crone, Donohue, Honomichl, Wendelken i Bunge, 2006; Luna i in., 2001; Moses i in., 2002; Schlaggar i in., 2002; Tamm i in., 2002; Thomas i wsp., 2004; Turkeltaub, Gareau, Flowers, Zeffiro i Eden, 2003).
W jaki sposób ta metodologia może nas poinformować, czy nastolatkom rzeczywiście brakuje wystarczającej kontroli poznawczej (impulsywnej) lub czy są ryzykowni w swoich wyborach i działaniach? Kontrola impulsów mierzona za pomocą zadań kontroli poznawczej, takich jak zadanie go / nogo, pokazuje liniowy wzór rozwoju w okresie dzieciństwa i dorastania, jak opisano powyżej. Jednak ostatnie badania neuroobrazowe rozpoczęły badanie przetwarzania związanego z nagrodami, specyficznego dla podejmowania ryzyka u młodzieży (Bjork i in., 2004; Ernst i in., 2005; May i in., 2004). Badania te skupiały się przede wszystkim na regionie półleżących, części jąder podstawnych zaangażowanych w przewidywanie nagrody, a nie na charakterystykę rozwoju tego regionu w połączeniu z odgórnymi regionami kontrolnymi (kora przedczołowa). Pomimo, że ostatnie doniesienie o mniejszej brzusznej aktywności przedczołowej u młodzieży w porównaniu z dorosłymi podczas pieniężnego podejmowania decyzji dotyczących zachowań ryzykownych, zostało wykazane (Eshel, Nelson, Blair, Pine i Ernst, 2007).
Ogólnie rzecz biorąc, w niewielu badaniach zbadano, w jaki sposób rozwój obwodów nagrody w regionach podkorowych (np. Półleżących) zmienia się wraz z rozwojem korowych obszarów przedczołowych. Co więcej, w jaki sposób te zmiany neuronalne pokrywają się z poszukiwaniem nagrody, impulsywnością i zachowaniami związanymi z podejmowaniem ryzyka, pozostaje stosunkowo nieznane. Nasz model neurobiologiczny sugeruje, że połączenie wzmożonej reakcji na nagrody i niedojrzałości w obszarach kontroli behawioralnej może zniechęcić młodzież do poszukiwania natychmiastowych, a nie długoterminowych zysków, być może wyjaśniając ich wzrost ryzykownych decyzji i zachowań impulsywnych. Śledzenie rozwoju podkorowego (np. Półleżącego) i korowego (np. Przedczołowego) procesu podejmowania decyzji w okresie dzieciństwa do dorosłości, zapewnia dodatkowe ograniczenia dotyczące tego, czy zmiany zgłaszane w okresie dojrzewania są specyficzne dla tego okresu rozwoju, czy odzwierciedlają dojrzewanie, które stale występuje w nieco liniowy wzór od dzieciństwa do dorosłości.
Dowody empiryczne z niedawnego badania fMRI pomagają wspierać nasz model neurobiologiczny i przyjmują przejściowe podejście do zrozumienia okresu dojrzewania poprzez badanie zmian przed i po okresie dojrzewania. W tym badaniu (Galvan i in., 2006), zbadaliśmy reakcje behawioralne i neuronalne w celu manipulacji nagrodami w rozwoju, koncentrując się na regionach mózgu zaangażowanych w uczenie się związane z nagrodami i zachowanie u zwierząt (Hikosaka i Watanabe, 2000; Pecina, Cagniard, Berridge, Aldridge i Zhuang, 2003; Schultz, 2006) i badania obrazowania dorosłych (np. Knutson, Adams, Fong i Hommer, 2001; O, Doherty, Kringelbach, Rolls, Hornak, Andrews, 2001; Zald i in., 2004) oraz w badaniach nad uzależnieniem (Hyman i Malenka, 2001; Volkow i Li, 2004). Na podstawie modeli gryzoni (Laviola i wsp., 1999; Włócznia, 2000) i poprzednie prace obrazowe (Ernst i in., 2005), postawiliśmy hipotezę, że w stosunku do dzieci i dorosłych młodzież wykazywałaby przesadną aktywację półleżących, w połączeniu z mniej dojrzałą rekrutacją odgórnych obszarów kontroli przedczołowej. Ostatnie prace pokazujące opóźnioną łączność funkcjonalną między tymi przedczołowymi i limbicznymi podkorowymi regionami w okresie dojrzewania w stosunku do dorosłych, zapewniają mechanizm braku odgórnej kontroli tych regionów (Hare i in., 2007a).
Nasze wyniki były zgodne z modelami gryzoni (Laviola, Macri, Morley-Fletcher i Adriani, 2003) i wcześniejsze badania obrazowe (Ernst i in., 2005) sugerując zwiększoną aktywność półleżących nagród w okresie dojrzewania. W rzeczywistości, w stosunku do dzieci i dorosłych, młodzież wykazywała przesadną odpowiedź półleżącą w oczekiwaniu na nagrodę. Jednak zarówno dzieci, jak i młodzież wykazywały mniej dojrzałą odpowiedź w obszarach kontroli przedczołowej niż dorośli. Odkrycia te sugerują, że różne trajektorie rozwojowe dla tych regionów mogą leżeć u podstaw wzmocnienia aktywności półleżącej w stosunku do dzieci lub dorosłych, co z kolei może wiązać się ze zwiększonymi impulsywnymi i ryzykownymi zachowaniami obserwowanymi w tym okresie rozwoju (patrz Rys. 4).

Różnicowa rekrutacja regionów przedczołowych i podkorowych została opisana w wielu badaniach rozwojowych fMRI (Casey i in., 2002b; Monk i in., 2003; Thomas i wsp., 2004). Zazwyczaj wyniki te interpretowano w kategoriach niedojrzałych obszarów przedczołowych, a nie nierównowagi między rozwojem regionalnym przedczołowym i podkorowym. Biorąc pod uwagę dowody dotyczące obszarów przedczołowych, kierując odpowiednimi działaniami w różnych kontekstach (Miller i Cohen, 2001) niedojrzała aktywność przedczołowa może utrudniać odpowiednie oszacowanie przyszłych wyników i ocenę ryzykownych wyborów, a zatem może mieć mniejszy wpływ na wycenę nagród niż wyniki półleżące. Ten wzorzec jest zgodny z wcześniejszymi badaniami wykazującymi podwyższone podkorowe, w stosunku do aktywności korowej, gdy decyzje są zniekształcone przez natychmiastowe długoterminowe zyski (McClure, Laibson, Loewenstein i Cohen, 2004). Ponadto wykazano aktywność półleżącą w przypadku fMRI, która pozytywnie koreluje z późniejszymi zachowaniami podejmującymi ryzyko (Kuhnen i Knutson, 2005). W okresie dojrzewania, w stosunku do dzieciństwa lub dorosłości, niedojrzała brzuszna kora przedczołowa może nie zapewniać wystarczającej kontroli odgórnej silnie aktywowanych regionów przetwarzania nagrody (np. Półleżących), powodując mniejszy wpływ systemów przedczołowych (kora oczodołowo-czołowa) w stosunku do półleżących w nagrodzie wycena.
Dlaczego mózg miałby być zaprogramowany do rozwoju w ten sposób?
Okres dojrzewania to okres przejściowy między dzieciństwem a dorosłością, często współwystępujący z okresem dojrzewania. Dojrzewanie oznacza początek dojrzewania płciowego (Graber i Brooks-Gunn, 1998) i mogą być zdefiniowane przez markery biologiczne. Dojrzewanie można opisać jako stopniowe przejście w dorosłość z mglistym kursem czasu ontogenetycznego (Włócznia, 2000). Ewolucyjnie mówiąc, okres dojrzewania to okres, w którym nabywane są umiejętności niezależności w celu zwiększenia sukcesu po oddzieleniu od ochrony rodziny, choć zwiększają szanse na szkodliwe okoliczności (np. Uraz, depresja, lęk, używanie narkotyków i uzależnienie (Kelley, Schochet i Landry, 2004). Zachowania poszukujące niezależności są powszechne wśród gatunków, takie jak wzrost interakcji społecznych kierowanych przez rówieśników i intensyfikacja poszukiwań nowości i zachowań ryzykownych. Czynniki psychospołeczne wpływają na skłonność młodzieży do ryzykownych zachowań. Jednak ryzykowne zachowania są wynikiem biologicznie nierównowagi między zwiększoną nowością a poszukiwaniem wrażeń w połączeniu z niedojrzałymi „kompetencjami samoregulacji” (Steinberg, 2004). Nasze dane neurobiologiczne sugerują, że dzieje się to poprzez zróżnicowany rozwój tych dwóch systemów (limbiczny i kontrolny).
Spekulacje sugerują, że ten wzorzec rozwojowy jest cechą ewolucyjną. Musisz angażować się w zachowania wysokiego ryzyka, aby opuścić rodzinę i wioskę, aby znaleźć partnera i podjąć ryzyko w tym samym czasie, w którym hormony prowadzą młodzież do poszukiwania partnerów seksualnych. W dzisiejszym społeczeństwie, gdy okres dojrzewania może trwać w nieskończoność, gdy dzieci mieszkają z rodzicami i mają zależność finansową oraz wybierają partnerów w późniejszym życiu, ewolucja ta może zostać uznana za niewłaściwą.
Istnieją dowody na istnienie wśród gatunków zwiększonego poszukiwania nowości i podejmowania ryzyka w okresie młodzieńczym. Szukając rówieśników i walcząc z rodzicami, które pomagają młodym ludziom oddalić się od rodzinnego terytorium w celu krycia, można zaobserwować u innych gatunków, w tym u gryzoni, naczelnych innych niż ludzie i niektórych ptaków (Włócznia, 2000). W porównaniu z dorosłymi szczurami o dojrzałym wieku wykazują zwiększone zachowania związane z poszukiwaniem nowości w paradygmacie nowatorskiego wyboru (Laviola i wsp., 1999). Dowody neurochemiczne wskazują, że równowaga w młodzieńczym mózgu między korowymi i podkorowymi układami dopaminy zaczyna się przesuwać w kierunku wyższych poziomów dopaminy w korze mózgowej w okresie dojrzewania (Włócznia, 2000). Podobny przedłużający się wzrost dopaminergiczny w okresie dojrzewania do dorosłości wykazano również w korze przedczołowej naczelnych innych niż ludzie (Rosenberg & Lewis, 1995). Tak więc to podwyższone pozorne podejmowanie ryzyka wydaje się występować wśród gatunków i ma ważne cele adaptacyjne.
Biologiczne predyspozycje, rozwój i ryzyko
Indywidualne różnice w kontroli impulsów i podejmowaniu ryzyka są od pewnego czasu rozpoznawane w psychologii (Benthin, Slovic i Severson, 1993). Być może jednym z klasycznych przykładów różnic indywidualnych zgłaszanych w tych zdolnościach w literaturze psychologii społecznej, poznawczej i rozwojowej jest opóźnienie gratyfikacji (Mischel, Shoda i Rodriguez, 1989). Opóźnienie gratyfikacji jest zazwyczaj oceniane u małych dzieci w wieku 3-do 4. Maluch jest pytany, czy wolałby małą nagrodę (jedno ciasteczko) czy dużą nagrodę (dwa pliki cookie). Następnie mówi się dziecku, że eksperymentator opuści pokój, aby przygotować się do nadchodzących zajęć i wyjaśnia dziecku, że jeśli pozostanie na swoim miejscu i nie zje ciasteczka, otrzyma dużą nagrodę. Jeśli dziecko nie może lub nie może czekać, powinna zadzwonić, aby wezwać eksperymentatora, a tym samym otrzymać mniejszą nagrodę. Kiedy stanie się jasne, że dziecko rozumie zadanie, siedzi przy stole z dwoma nagrodami i dzwonkiem. Rozproszenia w pomieszczeniu są zminimalizowane, bez zabawek, książek i zdjęć. Eksperymentator wraca po min. 15 lub po tym, jak dziecko zadzwoni do dzwonka, zje nagrodę lub przejawi jakiekolwiek oznaki cierpienia. Mischel pokazał, że dzieci zazwyczaj zachowują się na jeden z dwóch sposobów: (1) albo dzwonią prawie natychmiast, aby mieć ciasteczko, co oznacza, że otrzymują tylko jeden; lub (2) czekają i optymalizują swoje zyski i otrzymują oba pliki cookie. Ta obserwacja sugeruje, że niektórzy ludzie są lepsi od innych pod względem zdolności do kontrolowania impulsów w obliczu bardzo istotnych zachęt, a to uprzedzenie można wykryć we wczesnym dzieciństwie (Mischel i in., 1989) i wydają się pozostawać przez cały okres dojrzewania i młodości (Eigsti i wsp., 2006).
Co może wyjaśniać indywidualne różnice w optymalnym podejmowaniu decyzji i zachowaniu? Niektórzy teoretycy twierdzą, że dopaminergiczne obwody mezolimbiczne, związane z przetwarzaniem nagrody, stanowią podstawę ryzykownych zachowań. Indywidualne różnice w tym obwodzie, takie jak warianty alleliczne w genach związanych z dopaminą, powodujące zbyt małą lub zbyt dużą ilość dopaminy w regionach podkorowych, mogą mieć związek z skłonnością do podejmowania ryzykownych zachowań (O'Doherty, 2004). Wykazano, że jądro półleżące zwiększa aktywność bezpośrednio przed dokonaniem ryzykownych wyborów paradygmatów ryzyka monetarnego (Kuhnen i Knutson, 2005; Matthews i in., 2004; Montague i Berns, 2002) i jak opisano wcześniej, młodzież wykazuje przesadną aktywność półleżącą w stosunku do satysfakcjonujących wyników w stosunku do dzieci lub dorosłych (Ernst i in., 2005; Galvan i in., 2006). Łącznie dane te sugerują, że młodzież może być bardziej narażona na ryzykowne wybory jako grupa (Ogrodnik i Steinberg, 2005), ale niektóre nastolatki będą bardziej skłonne do angażowania się w ryzykowne zachowania, co potencjalnie zwiększa ryzyko negatywnych wyników. Dlatego ważne jest rozważenie indywidualnej zmienności podczas badania złożonych relacji mózg-zachowanie związanych z podejmowaniem ryzyka i przetwarzaniem nagrody w populacjach rozwojowych.
Aby zbadać indywidualne różnice w zachowaniach ryzykownych, Galvan i in. (2007) niedawno zbadał związek między aktywnością w obwodach nerwowych związanych z nagrodami w oczekiwaniu na dużą nagrodę pieniężną z cechami osobowości podejmującymi ryzyko i impulsywnością w okresie dojrzewania. Funkcjonalne obrazowanie metodą rezonansu magnetycznego i anonimowe skale oceny samooceny ryzykownych zachowań, percepcji ryzyka i impulsywności uzyskano u osób w wieku 7 i 29. Pomiędzy aktywnością półleżącą a prawdopodobieństwem angażowania się w ryzykowne zachowania w ramach rozwoju wystąpił pozytywny związek. Aktywność ta różniła się w zależności od ocen indywidualnych przewidywanych pozytywnych lub negatywnych konsekwencji takiego zachowania. Osoby, które postrzegały ryzykowne zachowania jako prowadzące do tragicznych konsekwencji, aktywowały mniejszą siłę nagradzania. Stowarzyszenie to było napędzane głównie przez dzieci, a dorośli oceniali konsekwencje takiego zachowania, jak to możliwe. Oceny impulsywności nie były związane z aktywnością półleżącą, ale raczej z wiekiem. Odkrycia te sugerują, że w okresie dojrzewania niektóre osoby mogą być bardziej skłonne do podejmowania ryzykownych zachowań z powodu zmian rozwojowych w połączeniu ze zmiennością predyspozycji danej osoby do angażowania się w ryzykowne zachowania, niż do prostych zmian w impulsywności (patrz Rys. 5).

Zachowanie młodzieży często było określane jako impulsywne i ryzykowne (Steinberg, 2004, 2007), jednak ten przegląd literatury obrazowej sugeruje różne substraty neurobiologiczne i różne trajektorie rozwojowe dla tych zachowań. W szczególności impulsywność jest związana z niedojrzałym brzusznym rozwojem przedczołowym i stopniowo zmniejsza się od dzieciństwa do dorosłości (Casey, Galvan i in., 2005). Negatywna korelacja między ocenami impulsywności a wiekiem w badaniu wg Galvan i in. (2007) dalej popiera to pojęcie. Przeciwnie, podejmowanie ryzyka wiąże się ze wzrostem aktywności półleżącej (Kuhnen i Knutson, 2005; Matthews i in., 2004; Montague i Berns, 2002), to jest przesadzone u nastolatków, w stosunku do dzieci i dorosłych (Ernst i in., 2005; Galvan i in., 2006). Tak więc wyborów i zachowań nastolatków nie można wytłumaczyć impulsywnością ani przedłużającym się rozwojem samej kory przedczołowej, ponieważ dzieci byłyby wtedy przewidywane jako osoby podejmujące większe ryzyko. Odkrycia dostarczają neuronalnych podstaw, dlaczego niektóre nastolatki są bardziej narażone na ryzyko niż inne, ale dodatkowo stanowią podstawę dla tego, jak zachowania nastolatków różnią się od dzieci i dorosłych podejmujących ryzyko.
Łącznie dane te sugerują, że chociaż młodzież jako grupa jest uważana za osoby podejmujące ryzyko (Ogrodnik i Steinberg, 2005), niektóre nastolatki będą bardziej skłonne do angażowania się w ryzykowne zachowania, narażając je na potencjalnie większe ryzyko negatywnych wyników. Odkrycia te podkreślają znaczenie uwzględnienia zmienności indywidualnej podczas badania złożonych relacji mózg-zachowanie związanych z podejmowaniem ryzyka i przetwarzaniem nagrody w populacjach rozwojowych. Co więcej, te różnice indywidualne i rozwojowe mogą pomóc wyjaśnić podatność niektórych osób na podejmowanie ryzyka związanego z używaniem substancji, a ostatecznie uzależnienie.
wnioski
Badania obrazowe u ludzi wykazują zmiany strukturalne i funkcjonalne w rejonach frontostriatalnych (Giedd i in., 1996, 1999; Jernigan i in., 1991; Sowell i in., 1999; do wglądu, Casey, Galvan i in., 2005), które wydają się równolegle zwiększać kontrolę poznawczą i samoregulację (Casey, Trainor i in., 1997; Luna & Sweeney, 2004; Luna i in., 2001; Rubia i in., 2000; Steinberg, 2004; Zobacz też Steinberg, 2008, ten przypadek). Zmiany te wydają się wykazywać przesunięcie w aktywacji regionów przedczołowych z rozproszonej do bardziej ogniskowej rekrutacji w czasie (Brown i wsp., 2005; Bunge i in., 2002; Casey, Trainor i in., 1997; Durston i in., 2006; Moses i in., 2002) i zwiększona rekrutacja regionów podkorowych w okresie dojrzewania (Casey i in., 2002a; Durston i in., 2006; Luna i in., 2001). Chociaż badania neuroobrazowe nie mogą definitywnie scharakteryzować mechanizmu takich zmian rozwojowych, te zmiany objętości i struktury mogą odzwierciedlać rozwój wewnątrz i udoskonalanie projekcji do iz tych obszarów mózgu podczas dojrzewania, sugerując dostrojenie systemu wraz z rozwojem.
Podsumowując, wyniki syntezy tutaj wskazują, że zwiększone ryzyko podejmowania zachowań w okresie dojrzewania jest związane z różnymi trajektoriami rozwojowymi podkorowych przyjemności i regionów kontroli korowej. Te zmiany rozwojowe mogą być pogłębione przez indywidualne różnice w aktywności systemów nagród. Chociaż okres dojrzewania został wyróżniony jako okres charakteryzujący się poszukiwaniem nagród i zachowaniem ryzykownym (Ogrodnik i Steinberg, 2005; Włócznia, 2000) indywidualne różnice w odpowiedziach neuronalnych na nagrodę predysponują niektóre nastolatki do podejmowania większego ryzyka niż inne, narażając je na większe ryzyko negatywnych wyników. Odkrycia te stanowią kluczowy grunt, syntetyzując różne odkrycia związane z podejmowaniem ryzykownych zachowań w okresie dojrzewania oraz rozumiejąc indywidualne różnice i markery rozwojowe, aby skłonność do angażowania się w negatywne zachowania.
Podziękowanie
Prace te zostały częściowo dofinansowane przez Narodowy Instytut Narkomanii R01 DA18879 i Narodowy Instytut Zdrowia Psychicznego 1P50 MH62196.
Referencje
- Benthin A, Slovic P, Severson H. Psychometryczne badanie percepcji ryzyka młodzieży. Journal of Adolescence. 1993;16: 153-168. [PubMed]
- Bjork JM, Knutson B, Fong GW, Caggiano DM, Bennett SM, Hommer DW. Aktywacja mózgu wywołana zachętą u młodzieży: podobieństwa i różnice między młodymi dorosłymi. Journal of Neuroscience. 2004;24: 1793-1802. [PubMed]
- Bjorkland DF. Rola wiedzy pojęciowej w rozwoju organizacji w pamięci dzieci. W: Brainerd CJ, Pressley M, redaktorzy. Podstawowe procesy w rozwoju pamięci: postęp w badaniach rozwoju poznawczego. Nowy Jork: Springer-Verlag; 1985. str. 103 – 142.
- Bjorkland DF. Jak zmiany wieku w bazie wiedzy przyczyniają się do rozwoju pamięci dzieci: przegląd interpretacyjny. Przegląd rozwojowy. 1987;7: 93-130.
- Bourgeois JP, Goldman-Rakic PS, Rakic P. Synaptogeneza w korze przedczołowej małp rezus. Kora mózgowa. 1994;4: 78-96. [PubMed]
- Brainerd CJ, Reyna VF. Niezależność pamięci i ingerencja pamięci w rozwój poznawczy. Przegląd psychologiczny. 1993;100: 42-67. [PubMed]
- Brown TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. Zmiany rozwojowe w ludzkiej funkcjonalnej organizacji mózgowej dla generowania słów. Kora mózgowa. 2005;15: 275-290. [PubMed]
- Bunge SA, Dudukovic NM, Thomason ME, Vaidya CJ, Gabrieli JD. Niedojrzały wpływ płata czołowego na kontrolę poznawczą u dzieci: dowody z fMRI. Neuron. 2002;33: 301-311. [PubMed]
- Przypadek R. Walidacja neo-Piagetowskiej konstrukcji pojemności. Journal of Experimental Child Psychology. 1972;14: 287-302.
- Sprawa R. Rozwój intelektualny: Narodziny do dorosłości. New York: Academic Press; 1985.
- Casey BJ, Castellanos FX, Giedd JN, Marsh WL, Hamburger SD, Schubert AB. et al. Implikacja prawego przedniego układu obwodowego w hamowaniu odpowiedzi i zaburzeniach uwagi / nadpobudliwości. Journal of American Academy of Child and Adolescent Psychiatry. 1997;36: 374-383. [PubMed]
- Casey BJ, Galvan A, Hare TA. Zmiany w funkcjonalnej organizacji mózgu podczas rozwoju poznawczego. Aktualna opinia w neurobiologii. 2005;15: 239-244. [PubMed]
- Casey BJ, Giedd JN, Thomas KM. Strukturalny i funkcjonalny rozwój mózgu i jego związek z rozwojem poznawczym. Psychologia biologiczna. 2000a;54: 241-257. [PubMed]
- Casey BJ, Thomas KM, Davidson MC, Kunz K, Franzen PL. Rozdzielanie funkcji prążkowia i hipokampa rozwojowo za pomocą zadania kompatybilności z bodźcem. Journal of Neuroscience. 2002a;22: 8647-8652. [PubMed]
- Casey BJ, Thomas KM, Welsh TF, Badgaiyan RD, Eccard CH, Jennings JR, et al. Dysocjacja konfliktu odpowiedzi, dobór uwagi i oczekiwanie z funkcjonalnym rezonansem magnetycznym. Materiały z Narodowej Akademii Nauk. 2000b;97: 8728-8733.
- Casey BJ, Tottenham N, Fossella J. Kliniczne, obrazowe, zmiany i genetyczne podejścia do modelu kontroli poznawczej. Psychobiologia rozwojowa. 2002b;40: 237-254. [PubMed]
- Casey BJ, Tottenham N, Liston C, Durston S. Obrazowanie rozwijającego się mózgu: Czego dowiedzieliśmy się o rozwoju poznawczym? Trendy w kognitywistyce. 2005;9: 104-110.
- Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN, et al. Badanie funkcjonalnego rezonansu magnetycznego MRI dotyczące przedczołowej aktywacji podczas wykonywania zadania go-no-go. Journal of Cognitive Neuroscience. 1997;9: 835-847.
- Crone E, Donohue S, Honomichl R, Wendelken C, Bunge S. Regiony mózgu pośredniczące w elastycznym stosowaniu reguł podczas rozwoju. Journal of Neuroscience. 2006;26: 11239-11247. [PubMed]
- Dempster FN. Odporność na zakłócenia: zmiany rozwojowe w podstawowym mechanizmie przetwarzania. W: Howe ML, Pasnak R, redaktorzy. Nowe tematy w rozwoju poznawczym Tom 1: Podstawy. Nowy Jork: Springer; 1993. str. 3 – 27.
- Diament A. Opracowanie możliwości użycia przypomnienia do kierowania działaniem, na co wskazuje wydajność niemowlęcia na AB. Rozwój dziecka. 1985;56: 868-883. [PubMed]
- Durston S, Davidson MC, Thomas KM, Worden MS, Tottenham N, Martinez A, et al. Parametryczna manipulacja konkurencją konfliktu i odpowiedzi za pomocą szybkiego fMRI związanego z badaniami mieszanymi. Neuroimage. 2003;20: 2135-2141. [PubMed]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella J, i in. Przejście od rozproszonej do ogniskowej aktywności korowej z rozwojem. Nauka o rozwoju. 2006;1: 18-20. [PubMed]
- Durston S, Hulshoff Pol HE, Casey BJ, Giedd JN, Buitelaar JK, van Engeland H. Anatomiczny MRI rozwijającego się mózgu człowieka: Czego się nauczyliśmy? Journal of American Academy of Child Adolescent Psychiatry. 2001;40: 1012-1020.
- Eaton LK, Kinchen S, Ross J, Hawkins J, Harris WA, Lowry R, i in. Nadzór nad zachowaniem ryzyka młodzieży - Stany Zjednoczone, 2005, podsumowania nadzoru. Raport tygodniowy "Śmiertelność i śmiertelność". 2006;55: 1-108. [PubMed]
- Eigsti IM, Zayas V, Mischel W, Shoda Y, Ayduk O, Dadlani MB, et al. Przewidywanie kontroli poznawczej od przedszkola do późnej młodości i młodości dorosłych. Nauka psychologiczna. 2006;17: 478-484. [PubMed]
- Ernst M, Nelson EE, Jazbec S, McClure EB, Monk CS, Leibenluft E, et al. Amygdala i jądro półleżące w odpowiedziach na otrzymanie i pominięcie przyrostów u dorosłych i młodzieży. Neuroimage. 2005;25: 1279-1291. [PubMed]
- Eshel N, Nelson EE, Blair RJ, Pine DS, Ernst M. Neuralne substraty z wyboru do wyboru u dorosłych i młodzieży: Rozwój kory przedczołowej przedniej i przedniej obręczy. Neuropsychologia. 2007;45: 1270-1279. [Artykuł bezpłatny PMC] [PubMed]
- Flavell JH, Feach DR, Chinsky JM. Spontaniczna próba słowna w zadaniu pamięci jako funkcja wieku. Rozwój dziecka. 1966;37: 283-299. [PubMed]
- Galvan A, Hare TA, Parra CE, Penn J, Voss H, Glover G, et al. Wcześniejszy rozwój półleżących w stosunku do kory oczodołowo-czołowej może być podstawą ryzykownych zachowań u młodzieży. Journal of Neuroscience. 2006;26: 6885-6892. [PubMed]
- Galvan A, Hare T, Voss H, Glover G, Casey BJ. Podejmowanie ryzyka a mózg młodzieży: Kto jest zagrożony? Nauka o rozwoju. 2007;10: F8-F14. [PubMed]
- Ogrodnik M, Steinberg L. Wpływ rówieśników na podejmowanie ryzyka, preferencje ryzyka i ryzykowne podejmowanie decyzji w okresie dojrzewania i dorosłości: badanie eksperymentalne. Psychologia rozwojowa. 2005;41: 625-635. [PubMed]
- Giedd JN. Obrazowanie strukturalnego rezonansu magnetycznego mózgu nastolatka. Roczniki z nowojorskiej Akademii Nauk. 2004;1021: 77-85. [PubMed]
- Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A, et al. Rozwój mózgu w dzieciństwie i okresie dojrzewania: podłużne badanie MRI. Natura Neuroscience. 1999;2: 861-863.
- Giedd JN, Snell JW, Lange N, Rajapakse JC, Casey BJ, Kozuch PL, et al. Ilościowe obrazowanie rezonansu magnetycznego w rozwoju ludzkiego mózgu: wiek 4 – 18. Kora mózgowa. 1996;6: 551-560. [PubMed]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, et al. Dynamiczne mapowanie rozwoju kory ludzkiej w dzieciństwie do wczesnej dorosłości. Postępowanie Narodowej Akademii Nauk Stanów Zjednoczonych Ameryki. 2004;101: 8174-8179. [Artykuł bezpłatny PMC] [PubMed]
- Graber JA, Brooks-Gunn J. Dojrzewanie. W: Blechman EA, Brownell KD, redaktorzy. Medycyna behawioralna a kobiety obszerny podręcznik. New York, NY: Guilford Press; 1998. str. 51 – 58.
- Hare TA, Voss HU, Glover GH, Casey BJ. Mózg młodzieńczy i potencjalne ryzyko lęku i depresji. 2007 Przesłany do publikacji.
- Hare TA, Voss HU, Glover GH, Casey BJ. Konkurencja między układami limbicznymi przedczołowymi i podkorowymi leży u podstaw reaktywności emocjonalnej w okresie dojrzewania. 2007b Przesłany do publikacji.
- Harnishfeger KK, Bjorkland F. Ontogeneza mechanizmów hamowania: odnowione podejście do rozwoju poznawczego. W: Howe ML, Pasnek R, redaktorzy. Nowe tematy w rozwoju poznawczym. Vol. 1. Nowy Jork: Springer-Verlag; 1993. str. 28 – 49.
- Hikosaka K, Watanabe M. Opóźniona aktywność orbitalnych i bocznych neuronów przedczołowych małpy różniących się różnymi nagrodami. Kora mózgowa. 2000;10: 263-271. [PubMed]
- Huttenlocher PR. Gęstość synaptyczna w korze czołowej człowieka - zmiany rozwojowe i skutki starzenia się. Brain Research. 1979;163: 195-205. [PubMed]
- Hyman SE, Malenka RC. Uzależnienie i mózg: neurobiologia przymusu i jego wytrwałość. Nature Reviews Neuroscience. 2001;2: 695-703.
- Jernigan TL, Zisook S, Heaton RK, Moranville JT, Hesselink JR, Braff DL. Nieprawidłowości obrazowania rezonansu magnetycznego w jądrach soczewkowatych i korze mózgowej w schizofrenii. Archiwa Psychiatrii Ogólnej. 1991;48: 811-823.
- Keating DP, Bobbitt BL. Indywidualne i rozwojowe różnice w komponentach przetwarzania poznawczego zdolności umysłowych. Rozwój dziecka. 1978;49: 155-167.
- Kelley AE, Schochet T, Landry C. Roczniki z nowojorskiej Akademii Nauk. 2004;1021: 27-32. [PubMed]
- Knutson B, Adams CM, Fong GW, Hommer D. Przewidywanie rosnącej nagrody pieniężnej selektywnie rekrutuje jądro półleżące. Journal of Neuroscience. 2001;21: RC159. [PubMed]
- Kuhnen CM, Knutson B. Neuralna podstawa podejmowania ryzyka finansowego. Neuron. 2005;47: 763-770. [PubMed]
- Laviola G, Adriani W, Terranova ML, Gerra G. Psychobiologiczne czynniki ryzyka podatności na psychostymulanty u młodzieży i modeli zwierzęcych. Neuroscience and Biobehavioral Reviews. 1999;23: 993-1010. [PubMed]
- Laviola G, Macri S, Morley-Fletcher S, Adriani W. Abstrakcyjne zachowania ryzykowne u młodzieńczych myszy: determinanty psychobiologiczne i wczesny wpływ epigenetyczny. Neuroscience and Biobehavioral Reviews. 2003;27: 19-31. [PubMed]
- Liston C, Watts R, Tottenham N, Davidson MC, Niogi S, Ulug AM, et al. Mikrostruktura frontostriatalna moduluje efektywną rekrutację kontroli poznawczej. Kora mózgowa. 2005;16: 553-560. [PubMed]
- Luna B, Sweeney JA. Pojawienie się wspólnych funkcji mózgu: badania FMRI nad rozwojem hamowania odpowiedzi. Roczniki z nowojorskiej Akademii Nauk. 2004;1021: 296-309. [PubMed]
- Luna B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ i in. Dojrzewanie szeroko rozproszonej funkcji mózgu podporządkowuje rozwój poznawczy. Neuroimage. 2001;13: 786-793. [PubMed]
- Matthews SC, et al. Selektywna aktywacja jądra półleżącego podczas podejmowania decyzji podejmujących ryzyko. Neuroreport. 2004;15: 2123-2127. [PubMed]
- Maj JC, Delgado MR, Dahl RE, Stenger VA, Ryan ND, Fiez JA, et al. Funkcjonalne obrazowanie rezonansu magnetycznego związanego z wydarzeniami w obwodach mózgu związanych z nagrodami u dzieci i młodzieży. Psychiatria biologiczna. 2004;55: 359-366. [PubMed]
- McClure SM, Laibson DI, Loewenstein G, Cohen JD. Oddzielne systemy neuronowe cenią natychmiastowe nagrody pieniężne. Science. 2004;306: 503-507. [PubMed]
- Miller EK, Cohen JD. Integracyjna teoria funkcji kory przedczołowej. Coroczny przegląd neuronauki. 2001;24: 167-202.
- Mischel W, Shoda Y, Rodriguez MI. Opóźnienie gratyfikacji u dzieci. Science. 1989;244: 933-938. [PubMed]
- Monk CS, McClure EB, Nelson EE, Zarahn E, Bilder RM, Leibenluft E, et al. Niedojrzałość młodzieży w związanym z uwagą zaangażowaniu mózgu w emocjonalną mimikę. Neuroimage. 2003;20: 420-428. [PubMed]
- Montague PR, Berns GS. Ekonomia neuronowa i biologiczne substraty wyceny. Neuron. 2002;36: 265-284. [PubMed]
- Moses P, Roe K, Buxton RB, Wong EC, Frank LR, Stiles J. Funkcjonalny MRI globalnego i lokalnego przetwarzania u dzieci. Neuroimage. 2002;16: 415-424. [PubMed]
- Munakata Y, Yerys BE. Teraz wszyscy razem: kiedy znikają podziały między wiedzą a działaniem. Nauka pscychologiczna. 2001;12: 335-337.
- Nagy Z, Westerberg H, Klingberg T. Dojrzewanie istoty białej wiąże się z rozwojem funkcji poznawczych w dzieciństwie. Journal of Cognitive Neuroscience. 2004;16: 1227-1233. [PubMed]
- O'Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Abstrakcyjne reprezentacje nagród i kar w ludzkiej korze oczodołowo-czołowej. Natura Neurosci. 2001;4: 95-102. [PubMed]
- O'Doherty JP. Reprezentacje nagród i związane z nagrodami uczenie się w ludzkim mózgu: wnioski z neuroobrazowania. Aktualne opinie w neurobiologii. 2004;14: 769-776.
- Pascual-Leone JA. Model matematyczny przejścia w etapach rozwoju Piageta. Acta Psychologica. 1970;32: 301-345.
- Pecina S, Cagniard B, Berridge KC, Aldridge JW, Zhuang X. Myszy z mutacją Hyperdopaminergic mają wyższe „pragnienie”, ale nie „lubienie” słodkich nagród. Journal of Neuroscience. 2003;23: 9395-9402. [PubMed]
- Reyna VF, Farley F. Ryzyko i racjonalność w podejmowaniu decyzji dotyczących młodzieży: konsekwencje dla teorii, praktyki i polityki publicznej. Nauka psychologiczna w interesie publicznym. 2006;7: 1-44.
- Rosenberg DR, Lewis DA. Postnatalne dojrzewanie unerwienia dopaminergicznego kory przedczołowej i motorycznej małpy: analiza immunohistochemiczna hydroksylazy tyrozynowej. The Journal of Comparative Neurology. 1995;358: 383-400. [PubMed]
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A, et al. Frontalizacja funkcjonalna wraz z wiekiem: mapowanie trajektorii neurorozwojowych za pomocą fMRI. Neuroscience and Biobehavioral Reviews. 2000;24: 13-19. [PubMed]
- Schlaggar BL, Brown TT, Lugar HM, Visscher KM, Miezin FM, Petersen SE. Funkcjonalne różnice neuroanatomiczne między dorosłymi a dziećmi w wieku szkolnym w przetwarzaniu pojedynczych słów. Science. 2002;296: 1476-1479. [PubMed]
- Schultz W. Teorie behawioralne i neurofizjologia nagrody. Roczne recenzje psychologii. 2006;57: 87-115.
- Sowell ER, Peterson BS, Thompson PM, Welcome SE, Henkenius AL, Toga AW. Mapowanie zmiany korowej w całym okresie życia człowieka. Natura Neuroscience. 2003;6: 309-315.
- Sowell ER, Thompson PM, Holmes CJ, Jernigan TL, Toga AW. Dowody in vivo na dojrzewanie mózgu u młodzieży w regionach czołowych i prążkowiu. Natura Neuroscience. 1999;2: 859-861.
- Sowell ER, Thompson PM, Toga AW. Mapowanie zmian w ludzkiej korze w ciągu całego życia. Neurobiolog. 2004;10: 372-392. [PubMed]
- Spear LP. Młodzieżowy mózg i związane z wiekiem objawy behawioralne. Neuroscience and Biobehavioral Reviews. 2000;24: 417-463. [PubMed]
- Steinberg L. Podejmowanie ryzyka w okresie dorastania: Jakie zmiany i dlaczego? Roczniki z nowojorskiej Akademii Nauk. 2004;1021: 51-58. [PubMed]
- Steinberg L. Podejmowanie ryzyka w okresie dojrzewania: nowe perspektywy z mózgu i nauk behawioralnych. Aktualne kierunki w naukach psychologicznych. 2007;16: 55-59.
- Steinberg L. Perspektywa neuronauki społecznej na temat podejmowania ryzyka przez młodzież. Przegląd rozwojowy. 2008;28: 78-106. [Artykuł bezpłatny PMC] [PubMed]
- Tamm L, Menon V, Reiss AL. Dojrzewanie funkcji mózgu związane z hamowaniem odpowiedzi. Journal of American Academy of Child and Adolescent Psychiatry. 2002;41: 1231-1238. [PubMed]
- Thomas KM, Hunt RH, Vizueta N, Sommer T, Durston S, Yang Y, et al. Dowody różnic rozwojowych w uczeniu się sekwencji ukrytych: badanie FMRI dzieci i dorosłych. Journal of Cognitive Neuroscience. 2004;16: 1339-1351. [PubMed]
- Turkeltaub PE, Gareau L, Flowers DL, Zeffiro TA, Eden GF. Rozwój mechanizmów neuronowych do czytania. Natura Neuroscience. 2003;6: 767-773.
- Volkow ND, Li TK. Uzależnienie od narkotyków: neurobiologia zachowań nie działa. Nature Reviews Neuroscience. 2004;5: 963-970.
- Yurgelun-Todd D. Zmiany emocjonalne i poznawcze w okresie dojrzewania. Aktualna opinia w neurobiologii. 2007;17: 251-257. [PubMed]
- Zald DH, Boileau I, El-Dearedy W, Gunn R, McGlone F, Dichter GS, et al. Transmisja dopaminy w ludzkim prążkowiu podczas zadań z nagrodami pieniężnymi. Journal of Neuroscience. 2004;24: 4105-4112. [PubMed]