Фронт Behav Neurosci. 2015 Май 19; 9: 128. doi: 10.3389 / fnbeh.2015.00128. eCollection 2015.
Andreatta M1, Паули П1.
Абстрактные
В классическом кондиционировании изначально нейтральный стимул (условный стимул, CS) становится связанным с биологически значимым событием (безусловный стимул, US), которым может быть боль (аверсивное кондиционирование) или еда (аппетитное кондиционирование). После нескольких ассоциаций CS способен инициировать либо защитные, либо завершающие реакции соответственно. В отличие от аверсивной обусловленности, аппетитная обусловленность редко исследуется на людях, хотя ее важность для нормального и патологического поведения (например, ожирения, зависимости) неоспорима. Настоящее исследование направлено на то, чтобы передать результаты исследований на животных о формировании аппетита людям, использующим пищу в качестве США. Тридцать три участника были обследованы между 8 и 10 часами утра без завтрака, чтобы убедиться, что они голодны. Во время двух этапов сбора данных одна геометрическая форма (avCS +) предсказывала аверсивный УЗИ (болезненный электрический шок), другая форма (appCS +) предсказывала аппетитное УЗИ (шоколадный или соленый крендель в соответствии с предпочтениями участников), а третья форма (CS– ) не предсказал ни США. В фазе исчезновения эти три формы плюс новая форма (НОВАЯ) были снова представлены без доставки из США. Рейтинги валентности и возбуждения, а также реакции испуга и проводимости кожи (SCR) были собраны в качестве показателей обучения. Мы обнаружили успешную аверсивную и аппетитную обусловленность. С одной стороны, avCS + был оценен как более отрицательный и более возбуждающий, чем CS–, и вызывал потенцирование испуга и усиление SCR. С другой стороны, appCS + был оценен более положительно, чем CS–, и вызвал ослабление испуга и больший SCR. Таким образом, мы успешно подтвердили находки животных у (голодных) людей, продемонстрировав аппетитное обучение и нормальное аверсивное обучение.
Введение
Предсказание угрозы и пищи имеет решающее значение для выживания любого организма. В классическом кондиционировании (Павлов, 1927), несколько раз предшествует аверсивное событие, такое как мягкий болезненный удар электрическим током (отвратительный безусловный стимул, США) или какое-то аппетитное событие, такое как пищевой таблет (аппетитный США). Впоследствии этот единственный стимул (теперь обозначенный CS, CS +) способен вызывать соответственно защитные или завершающие ответы. Первый вид ассоциативного обучения называется аверсивным обучением, в то время как последнее называется аппетитным обучением.
Хотя предсказание аппетитных событий столь же важно для выживания, как и предсказание аверсивных событий, аппетитная обусловленность значительно менее изучена у животных (Бутон и Пек, 1989; Koch et al., 1996; McDannald и др., 2011, для обзора см. Martin-Soelch и др., 2007), а также у людей (Klucken et al., 2009, 2013; Остин и Дука, 2010; Delgado et al., 2011; Леви и Глимчер, 2011). Возможно, это отсутствие исследований может быть связано со сложностью аппетитной парадигмы по сравнению с аверсивной. Например, пища в качестве основного усилителя должна быть доставлена, когда организм голоден, чтобы быть полезным (для недавнего обзора см. Дикинсон и Баллейн, 1994; Clark et al., 2012). В исследовании человека эта трудность была преодолена за счет использования денег (Остин и Дука, 2010; Delgado et al., 2011; Леви и Глимчер, 2011) или эротические фотографии (Klucken et al., 2009, 2013). Тем не менее, в исследованиях нейроотображений указывалось, что первичные (например, закуски или напитки) и вторичные (т. Е. Деньги) усилители активируют некоторые распространенные области мозга (например, стриатум), но также влекут за собой различные формы активации (Delgado et al., 2011; Леви и Глимчер, 2011).
Насколько нам известно, только несколько исследований по кондиционированию человека изучали влияние первичных аппетитных усилителей, таких как запах (Готфрид и др., 2002), воды (Kumar et al., 2008), или пища (Prévost и др., 2012). В первом исследовании, Gottfried et al. (2002) (CS) с неприятным, приятным или нейтральным запахом. Интересно, что они обнаружили большую активацию в орбитофронтальной коре (ОФК) и в брюшном полосатом теле в ответ на аппетитный CS + против аверсивного CS +. Они пришли к выводу, что OFC обрабатывает значение запаха и участвует в передаче аффективной ценности от обонятельной (США) к визуальной (CS) системе. Ветряная активация (вентро) была интерпретирована как отражающая аппетитный CR, вызванный аппетитной CS +. Во втором исследовании, Kumar et al. (2008) приглашали своих участников (здоровый контроль и пациентов с большой депрессией) в лабораторию рано утром и попросили их воздержаться от питья в течение ночи, чтобы убедиться, что они испытывали жажду до сканирования. Фрактальными снимками были CS и 0.1 мл воды в США. Интересно отметить, что среди других активизаций здоровые участники (но не пациенты с депрессией) показали более активную активацию в брюшном полосатом теле с аппетитным CS +, предполагая, что этот стимул был обработан как полезный. В третьем исследовании, Prévost et al. (2012) также представили фрактальные картинки как CS и сладкие или соленые закуски как US в зависимости от предпочтений участников. КС предъявляли в течение 6 с и в течение последней секунды дополнительно представляли картину питания. Каждый раз, когда появлялась картинка с едой, экспериментатор передавал кусочек еды участникам, которым разрешалось сразу же есть закуску. К сожалению, авторы не сообщили об активации мозга аппетитным CS + во время классической фазы кондиционирования; однако они наблюдали меньшее замедление сердечного ритма при вознаграждении CS + по сравнению с CS– без вознаграждения, что указывает на дифференциальные эффекты кондиционирования.
Отсутствие исследований, использующих ответ испуга в качестве показателя аппетитной кондиционирования, вызывает удивление, особенно учитывая его широкое использование в аверсивном обучении. Исходный ответ - это предковый и автоматический защитный ответ на внезапные, неожиданные и сильные отвратительные события (Koch, 1999). Этот защитный ответ опосредуется относительно простым нейронным путем, включающим в себя кохлеарные корневые нейроны, каудальное понтийское ядро ретикулярной формации (PnC) и спинальные мотонейроны (Fendt и Fanselow, 1999; Koch, 1999). Исследования на животных показали, что потенцирование поражений зависит от прогнозов от миндалины до PnC (Fendt и Fanselow, 1999; Koch, 1999), тогда как затухание затмения зависит от интактного ядра accumbens (NAcc, Koch, 1999). Такая модуляция ответного удара является полезной неявной мерой для валентности представленных передних стимулов. Таким образом, потенцирование указывает на отрицательную валентность, в то время как ослабление указывает на положительную валентность, и без значительного влияния на когнитивные процессы (Hamm и Weike, 2005; Andreatta и др., 2010). Насколько нам известно, только одно исследование на животных изучало аппетитную обусловленность, измеряя реакции испуга как зависимую меру. Интересно, что исследованные крысы проявляли затухание пурпа в CS + (то есть свет), связанное с доставкой раствора сахарозы. Примечательно, что такое ослабление ухудшалось у этих животных с NAcc-повреждениями, но не у пациентов с амигдальскими поражениями, что указывает на то, что NAcc играет определенную роль в выявлении аппетитных CR и ослаблении ответов на поражение (Koch et al., 1996).
Нынешнее исследование, насколько нам известно, первым перевело эту парадоксальную парадигму кондиционирования для людей, используя первичные усилители как США, то есть сладкие (шоколадные Smarties®) или соленый (мелкий соленый крендель), а также модуляция пуска в качестве меры CR. Мы ожидали, что appCS + вызовет сильные аппетитные CR, что отразится на ослаблении пуска, увеличении SCR и положительном рейтинге валентности по сравнению с другими стимулами, то есть с avCS + и CS-.
Материалы и методы
Участниками
Сорок два волонтера приняли участие в исследовании и получили зачетные единицы. Девять участников были исключены из анализа, двое из-за технических проблем, трое из-за того, что они были закодированы как не отвечающие (средняя амплитуда испуга <5 мкВ), и четверо из-за того, что они не демонстрировали достаточного количества испуганных ответов на каждое условие (минимум = 2; подробности см. в разделе «Материалы и методы»). В итоге в анализе было рассмотрено 33 участника (16 мужчин; средний возраст: 22.09 года, SD: 2.84; диапазон: 18-29 лет). Четверо участников не были уроженцами немцев, а шесть были левшами. Один участник не знал о ассоциациях CS-US на протяжении всего эксперимента (см. Процедуру), но мы решили не исключать этого участника, потому что его ответы были нормальными и не влияли на результаты.
Материалы
Безусловные стимулы (США)
Использовались два вида США. В качестве отвратительного США мы применили мягко-мучительный электрический удар по недоминируемому предплечью участников. Электрический шок был доставлен с помощью двух электродов с диаметром 9 мм и расстоянием 30 см. Электрический стимул состоял из импульсного стимула с частотой 50 Гц и продолжительностью 200 мс, генерируемого ток-стимулятором (Digitimer DS7A, Digitimer Ltd, Welwyn Garden City, UK, 400 V, максимум 9.99 мА). Интенсивность электрического удара определялась индивидуально с помощью пороговой процедуры, описанной ранее (Andreatta и др., 2010). Вкратце, участники прошли две серии восходящих и нисходящих интенсивностей в интервалах 0.5 мА. Они должны были оценивать каждый стимул в визуальном масштабе от 0 (вообще ничего не чувствуя) до 10 (действительно сильная боль) с 4 как якорь для порога (просто заметная боль). Средняя интенсивность электрического стимула составляла 2.12 мА (SD = 0.56), и он был оценен как болезненный (M = 6.45, SD = 1.73). Прикладные США состояли либо из шоколада (Smarties®) или небольшой соленый крендель. Выбор аппетитного УЗИ зависел от индивидуальных предпочтений участника, о которых сообщалось во время предварительного интервью. А именно, участники должны были сообщить, едят ли они обычно соленую или сладкую пищу во время завтрака. Они также могли свободно выбирать, предпочитают ли они шоколад или соленый крендель во время самого эксперимента. В итоге 22 участника выбрали шоколад, а 11 - небольшой соленый крендель.
Условные стимулы (CS)
Геометрические фигуры (синий квадрат, желтый круг, зеленый треугольник, красный шестиугольник) с диагональю 8 см представлены как CS. Формы были представлены в середине черного экрана компьютера для 8. Одна форма (avCS +) всегда ассоциировалась с аверсивным УЗИ (болезненный удар электрическим током), одна форма (appCS +) всегда ассоциировалась с аппетитным US (шоколадом или соленым кренделем), одна форма (CS-) никогда не ассоциировалась ни с аверсивным или аппетитные USs, а четвертая форма (NEW) была представлена на фазе вымирания, но не во время фазы приобретения, чтобы обеспечить ее нейтральность.
Пусковой зонд
Белый шум 103 дБ длительностью 50 мс использовался в качестве пробника взрыва. Акустические стимулы подавались бинаурально через наушники и возникали случайным образом через 4–6 с после появления формы.
Анкетирование
До и после эксперимента участники должны были заполнить немецкие версии инвентаря тревог состояния (STAI, Laux et al., 1981) и положительного отрицательного графика влияния (PANAS, Krohne et al., 1996). STAI - это инвентарь для оценки черт и / или состояния тревожности участников, который состоит из 20 пунктов для версии черты и состояния. Уровень тревожности участников до (M = 37.06, SD = 7.80) и после (M = 39.33, SD = 9.16), эксперимент существенно не изменился [t(32) = 1.61, p = 0.117]. Оценки тревожных признаков в текущем образце варьировались между 20 и 58 (M = 36.6, SD = 8.98), что сопоставимо с опубликованным нормальным диапазоном взрослых (Laux et al., 1981). PANAS (Krohne et al., 1996) - показатель позитивного и негативного настроения. Люди с высокими баллами по шкале положительных аффектов (PAS) склонны к таким эмоциям, как энтузиазм, в то время как люди с высокими баллами по шкале отрицательных аффектов (NAS) предрасположены к таким эмоциям, как дистресс. Каждый пункт состоит из прилагательного, и участники указывают по шкале от 1 (очень незначительно) до 5 (чрезвычайно), в какой степени прилагательное отражает их чувства в данный конкретный момент. В результате эксперимента не было обнаружено значительных различий в негативном влиянии участников [начало: M = 11.67, SD = 2.29; конец: M = 12.88, SD = 4.69; t(32) = 1.55, p = 0.130]. Так или иначе, участники с самого начала значительно изменили свое позитивное настроение (M = 26.72, SD = 4.70) до конца (M = 23.66, SD = 7.02) эксперимента [t(31) = 3.11, p = 0.004]. Это снижение позитивного настроения участников могло быть связано с неприятной парадигмой (были представлены болезненные удары током, а также отталкивающий белый шум).
Процедура
По прибытии в лабораторию участники прочитали и подписали форму информированного согласия, одобренную этическим комитетом факультета психологии Университета Вюрцбурга. Они не были проинформированы о непредвиденных обстоятельствах между CS и US. После заполнения вопросников электроды были прикреплены и процедура порога болевого синдрома была выполнена, как описано выше.
Во время этап привыкания, четыре геометрические фигуры были представлены дважды с интервалом между интервалами (ITI), изменяющимся между 18 и 25 s (в среднем: 21.5 s). На этом этапе не было поставлено ни одного пробника США или пуска.
До фазы сбора семь шумов белого шума были доставлены каждые 7-15 s, чтобы уменьшить начальную реактивность запуска.
Следующие два фаз сбора были идентичны (рис. 1). Каждая фаза приобретения состояла из испытаний 24: 8 CS-испытаний, испытаний 8 avCS + и испытаний 8 appCS +. Последовательность CS была псевдослучайной, единственным ограничением было то, что один и тот же стимул не мог быть представлен более чем в два раза подряд. Примечательно, что avCS + был представлен вместе с молнией в качестве символа поражения электрическим током, а болезненные США были поставлены на зачет. AppCS + был представлен в составе с изображением Smarties или соленым кренделем, и участник мог выбрать Smarties или крендель из банки. CS - был представлен вместе с символом запрета, и США не были доставлены. Во время трех презентаций 8 CS каждого типа с момента начала стимула между 4 и 6 s был поставлен датчик испуга. В ходе ITI были представлены три дополнительных пробных зонда, чтобы обеспечить их непредсказуемость и уменьшить привычку к запугиванию. ITI, состоящий из черного экрана, варьировался между 18 и 25 s со средним значением 21.5.
Рисунок 1
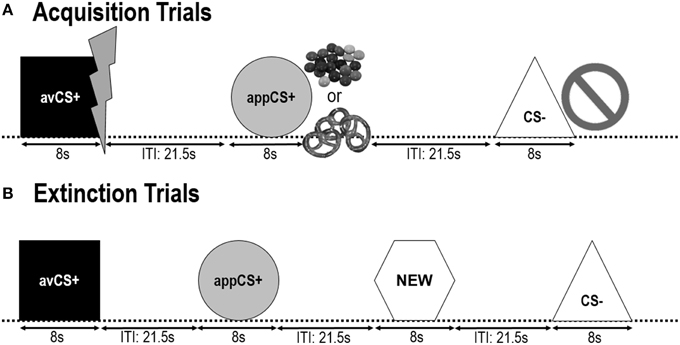
Рисунок 1. Испытания в течение двух фаз сбора (A) и фазы исчезновения (B), Участники узнали, что одна форма (avCS +) предсказала мягкий болезненный электрический шок, одна форма (appCS +) предсказывала либо кусок шоколада, либо небольшой соленый крендель (по их предпочтениям), а третья форма (CS-) не предсказывала никаких биологически значимое событие. Каждая форма была представлена в сочетании с рисунком, изображающим электрический шок, умение / соленый крендель или ничто в зависимости от ассоциации с США. Во время фазы исчезновения три геометрические фигуры были представлены еще раз, но США не были доставлены. Кроме того, в качестве нейтрального контроля была представлена четвертая геометрическая форма (NEW).
Во время фаза исчезновения, участники увидели три геометрические фигуры (например, avCS +, appCS + и CS-), а также новую нейтральную форму (NEW). США не были доставлены, и фигуры не были представлены в сочетании с фотографиями вспышек, шоколадными или солеными кренделями или запретами. Каждый стимул был представлен восемь раз в псевдослучайном порядке (т. Е. Один и тот же стимул не был представлен более двух раз подряд), создавая испытания 32. Начальные стимулы зонда были представлены во время 4 из презентаций стимулов 8 каждого типа CS. Как и в процессе приобретения и фазы привыкания, ITI варьировались между 18 и 25 s, а дополнительные датчики запуска 4 поставлялись непредсказуемо во время ITI.
После каждой фазы участники оценивали валентность (приятность) и возбуждение (возбуждение) CS с визуальными аналоговыми шкалами (VAS) от 1 до 9. Шкала валентности варьировалась от «1», что указывало «очень неприятно» на «9», что указывало «очень приятно», масштаб пробуждения варьировался от «1», обозначающий «спокойствие» до «9», что указывало на «возбуждение». Кроме того, рейтинги непредвиденных обстоятельств были оценивается после двух фаз сбора и фазы экстинкции. Участники увидели геометрическую форму для 1 s, а затем они должны были указать, была ли эта форма связана с поражением электрическим током, с шоколадным / соленым кренделем, ни с чем, или же они не смогли установить какую-либо связь. Примечательно, что все участники (кроме одного) знали о непредвиденных обстоятельствах после приобретения 2.
Сжатие данных
Физиологические ответы были записаны с помощью усилителя V-Amp 16 и программного обеспечения Vision Recorder V-Amp Edition (версия 1.03.0004, BrainProducts Inc., Мюнхен, Германия). Применялась частота дискретизации 1000 Гц и фильтр с надрезом 50 Гц. Анализы в автономном режиме проводились с помощью анализатора мозга Brain Vision (версия 2.0; BrainProducts Inc., Мюнхен, Германия).
Исходный ответ
Начальный ответ измеряли с помощью электромиографии (ЭМГ) слева Круговая мышца глаза мышцы с двумя электродами Ag / AgCl 5 мм. В соответствии с руководящими принципами (Blumenthal и др., 2005), один электрод располагался под зрачком, а второй - 1 см в поперечном направлении. Наземные и эталонные электроды были помещены соответственно на правый и левый мастоиды. Перед установкой электродов кожа была слегка абразирована и очищена спиртом, чтобы поддерживать импеданс ниже 10 kΩ. Электромиографический сигнал был отключен фильтром с фильтром низкого отсечки 28 Гц и фильтром отсечки высокого разрешения 500 Гц. Затем сигнал EMG был исправлен и применено скользящее среднее 50 ms. Мы использовали 50 ms перед запуском зонда в качестве базовой линии (Grillon и др., 2006). Ответы на пусковые зонды оценивались вручную, и испытания с чрезмерными сдвигами базовой линии (± 5 мкВ) или артефактами движения были исключены из дальнейшего анализа. Начальные ответы ниже 5 μV были закодированы как ноль и рассмотрены для расчета величины пуска (Blumenthal и др., 2005). В целом, 10.4% испытаний были отклонены, и минимум 2 из 3 пустых ответов на фазах сбора и 4 из 8 начальных ответов на фазе экстинкции для каждого условия были необходимы, чтобы сохранить участника в пуле анализа. По этой причине четыре участника были исключены. Пиковая амплитуда определялась как максимальный пик относительно базовой линии во время временного окна 20-120 ms после начала зонда зонда. Затем исходные данные были нормированы внутри субъектов, используя z-scores, а затем T-баллы, чтобы уменьшить влияние индивидуальной изменчивости и лучше выявить психологические процессы. T-баллы усреднялись для каждого условия (avCS +, appCS +, CS-, NEW и ITI). Чтобы исследовать потенцирование запаха или затухание при запуске, оценки для ответов ITI startle были вычтены из ответов о пуске для каждого условия.
Реакция электропроводности кожи (SCR)
Реакция проводимости кожи (SCR) регистрировалась с использованием двух электродов 5 mm Ag / AgCl, помещенных на ладонь недоминирующей руки. Гальванический отклик был отключен фильтром с фильтром отсечки высокого разрешения 1 Гц. SCR определяли как разность (в μS) между началом ответа (1-3 s после начала стимула) и пиком ответа (Tranel и Damasio, 1994; Delgado et al., 2011). Испытания, содержащие пусковые зонды, не учитывались при анализе SCR. Ответы ниже 0.02 μS были закодированы как ноль. Еще пять участников были исключены из анализа SCR, поскольку они имели среднее значение SCR ниже 0.02 μS. Необработанные данные о проводимости кожи были преобразованы квадратным корнем для нормализации распределения, и оценки были усреднены для каждого условия отдельно для двух фаз сбора (avCS +, appCS +, CS-) и фазы экстинкции (avCS +, appCS +, CS- и NEW ).
Статистический анализ
Все данные были проанализированы с помощью SPSS для Windows (версия 20.0, SPSS Inc.). Для физиологических реакций были рассчитаны раздельные дисперсионные дисперсии (ANOVA) для двух фаз сбора и фазы исчезновения. У ANOVA для фаз сбора были стимулы (avCS +, appCS +, CS-) и фаза (Приобретение 1, Приобретение 2) как факторы внутри предмета. ANOVA для фазы исчезновения имел только стимул (avCS +, appCS +, CS-, NEW) как фактор внутри предмета. Оценки валентности, возбуждения и непредвиденных обстоятельств анализировались с помощью отдельных ANOVA, содержащих стимулы внутри субъектов (avCS +, appCS +, CS- и NEW) и фазы. Этот коэффициент имел четыре уровня для валентных и возбуждающих оценок (T1: после фазы привыкания, T2: после первой фазы сбора данных T3: после второй фазы сбора данных T4: после фазы исчезновения), но три уровня для рейтингов непредвиденных ситуаций (T1: после первая фаза получения, T2: после второй фазы сбора данных T3: после фазы исчезновения).
Уровень альфа (α) был установлен для 0.05 для всех анализов. Размер эффекта сообщается как частичный η2.
Итоги
Оценки валентности и возбуждения для каждой фазы изображены на рисунке 2; ответы о пуске и SCR показаны на рисунке 3.
Рисунок 2
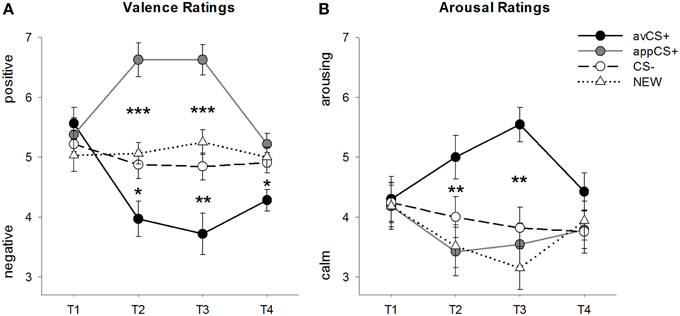
Рисунок 2. Оценки валентности (A) и возбуждения (B), Строки (со стандартными ошибками) отображают рейтинги после фазы привыкания (T1), приобретения 1 (T2), приобретения 2 (T3) и фазы исчезновения (T4). Аверсивная CS + (черная сплошная линия) приобрела отрицательную валентность и высокое возбуждение после двух фаз сбора по сравнению с CS- (черная пунктирная линия) и NEW (черная пунктирная линия). Важно отметить, что аппетитная CS + (серая сплошная линия) приобрела положительную валентность по сравнению с CS- и NEW. *p <0.05 **p > 0.01, ***p <0.001.
Рисунок 3
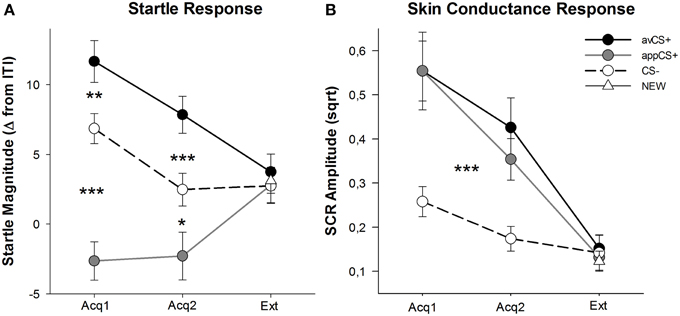
Рисунок 3. Начальные ответы (A) и проводимость кожи (B) (со стандартными ошибками) во время первой фазы сбора данных (Acq1), второй фазы сбора данных (Acq2) и фазы исчезновения (Ext), Исходные ответы были значительно усилены для аверсивной CS + (черная сплошная линия) и значительно ослаблены к аппетитной CS + (серая сплошная линия) по сравнению с CS- (черная пунктирная линия) во время фаз сбора. SCR был значительно больше для avCS + и appCS + по сравнению с CS-. На этапе исчезновения никаких различий не выявлено. *p <0.05 **p > 0.01, ***p <0.001.
Рейтинги
ANOVA для валентные оценки во время приобретения выявили значительные основные эффекты стимула [F(3, 93) = 17.26, GG-ε = 0.801, p <0.001, частичное η2 = 0.358] и фазы [F(3, 93) = 3.30, GG-ε = 0.731, p = 0.039, частичный η2 = 0.096], а также значительное взаимодействие между стимулом и фазой [F(9, 279) = 9.54, GG-ε = 0.463, p <0.001, частичное η2 = 0.235]. Следовать за t- тесты показывают, что валентности четырех геометрических фигур в начале эксперимента были идентичны (ps> 0.19), тогда как после получения 1 и 2 avCS + был оценен как особенно отрицательный, а appCS + - как особенно положительный. В частности, avCS + был оценен как более отрицательно валентный по сравнению с CS– [Acq1: t(31) = 2.34, p = 0.026; Acq2: t(31) = 3.07, p = 0.004], NEW [Acq1: t(31) = 2.70, p = 0.011; Acq2: t(31) = 3.89, p <0.001], а appCS + [Acq1: t(31) = 5.41, p <0.001; Acq2: t(31) = 6.11, p <0.001]. AppCS + был оценен как значительно более положительный, чем CS– [Acq1: t(31) = 4.99, p <0.001; Acq2: t(31) = 5.31, p <0.001] и НОВОЕ [Acq1: t(31) = 4.92, p <0.001; Acq2: t(31) = 4.14, p <0.001]. Различия между CS– и NEW никогда не были значительными (ps> 0.18).
Важно отметить, что никаких существенных различий при сравнении валентных оценок для appCS +, связанных с шоколадом и соленым кренделем [Acq1: t(30) = 0.03, p = 0.477; Acq2: t(30) = 0.29, p = 0.775].
После фазы экстинкции avCS + по-прежнему считался более негативным по сравнению с CS- [t(31) = 2.40, p = 0.023], NEW [t(31) = 3.40, p = 0.002], а appCS + [t(31) = 3.35, p = 0.002]. Напротив, валентность appCS + больше не отличалась от CS- [t(31) = 1.77, p = 0.086] или NEW [t(31) = 1.07, p = 0.293].
ANOVA для рейтинги возбуждения во время приобретения выявлено значительное основное влияние стимула [F(3, 96) = 7.07, GG-ε = 0.737, p = 0.001, частичный η2 = 0.181], но не фазы [F(3, 96) = 1.27, GG-ε = 0.805, p = 0.289, частичный η2 = 0.038], а также значительное взаимодействие между стимулом и фазой [F(9, 288) = 4.53, GG-ε = 0.582, p = 0.001, частичный η2 = 0.124]. Следовать за t- тесты не выявили существенных различий между стимулами относительно их первоначального возбуждения (ps> 0.74). Однако после двух этапов приобретения avCS + был оценен более возбуждающе, чем NEW [Acq1: t(32) = 2.99, p = 0.005; Acq2: t(32) = 5.97, p <0.001] и appCS + [Acq1: t(32) = 2.62, p = 0.013; Acq2: t(32) = 4.42, p <0.001], и немного больше возбуждает, чем CS - после первой фазы сбора данных [t(32) = 1.96, p = 0.058], но значительно больше пробуждения после второй фазы сбора [t(32) = 3.65, p = 0.001]. В отличие от валентных рейтингов возбуждение appCS + не отличалось от возбуждения CS- и NEW (ps > 0.13) после фаз сбора данных.
Точно так же, как и с валентными рейтингами, CS- и NEW не отличались в своем возбуждении (ps> 0.07).
Как и в случае с валентностью, никаких различий не было обнаружено для appCS +, связанных с шоколадом, и appCS +, связанных с соленым кренделем [Acq1: t(31) = 0.26, p = 0.797; Acq2: t(31) = 0.33, p = 0.724].
После фазы исчезновения достоверных различий в рейтингах возбуждения не выявлено (ps> 0.08).
Для дополнительных пост-hoc t- тесты, сравнивающие рейтинги после фазы привыкания, первой и второй фаз сбора и фазы исчезновения, см. в дополнительных материалах.
Исходный ответ
ANOVA для фаз сбора возвращал основной эффект стимула [F(2, 64) = 49.92, GG-ε = 0.964, p <0.001, частичное η2 = 0.609], но не фаза [F(1, 32) = 3.16, p = 0.085, частичный η2 = 0.090] и значительное взаимодействие Стимул × Фаза [F(2, 64) = 3.37, GG-ε = 0.875, p = 0.048, частичный η2 = 0.095]. Следовать за t-tests выявили значительную атакующую атаку на avCS + по сравнению с CS- во время первой [t(32) = 3.27, p = 0.003], а второй [t(32) = 4.00, p <0.001] фазы сбора данных. Реакции испуга на avCS + также были значительно усилены по сравнению с таковыми на appCS +, опять же после того, как Acquisition 1 [t(32) = 8.20, p <0.001] и приобретение 2 [t(32) = 5.74, p <0.001]. Важно отметить, что величина потрясения для appCS + была значительно ослаблена по сравнению с CS– во время получения 1 [t(32) = 6.34, p <0.001] и приобретение 2 [t(32) = 2.91, p = 0.007]. Снова и в соответствии с рейтингами не было обнаружено дифференциальных ответов о путях для appCS +, когда они ассоциировались с шоколадом или соленым кренделем [Acq1: t(31) = 1.04, p = 0.309; Acq2: t(31) = 0.07, p = 0.947]. На этапе исчезновения никакого существенного эффекта не обнаружено [F(3, 96) = 0.26, GG-ε = 0.906, p = 0.833, частичный η2 = 0.008].
Реакция электропроводности кожи (SCR)
Из ANOVA для SCR в течение двух фаз сбора, основной эффект стимула [F(2, 54) = 18.04, GG-ε = 0.908, p <0.001, частичное η2 = 0.401] и фазы [F(1, 27) = 20.91, p <0.001, частичное η2 = 0.436] оказались значимыми, но не их взаимодействие [F(2, 54) = 0.68, GG-ε = 0.637, p = 0.451, частичный η2 = 0.024]. Post-hoc t- тесты показали значительно больший SCR для avCS + [t(27) = 6.46, p <0.001] и в appCS + [t(27) = 4.84, p <0.001] по сравнению с CS–, в то время как участники показали сопоставимый SCR с avCS + и appCS + [t(27) = 0.64, p = 0.527]. Примечательно, что никаких различий в SCR к шоколаду appCS + и к соленой кренделице appCS + не найдено [Acq1: t(26) = 2.55, p = 0.120; Acq2: t(26) = 1.29, p = 0.210]. Как и в случае рейтингов и ответа на поражение, никаких существенных эффектов для фазы исчезновения [F(3, 81) = 0.28, GG-ε = 0.634, p = 0.743, частичный η2 = 0.010].
Обсуждение
Цель этого исследования состояла в том, чтобы перевести результаты животных на людей, используя классическую парадигму условного кондиционирования с первичным подкреплением как безусловный стимул (США). Чтобы сделать это, участники пришли в лабораторию рано утром без завтрака, чтобы гарантировать, что они голодны, и, согласно их предпочтениям, кусочки шоколада или соленых кренделей использовались в качестве аппетитных US. Во время фазы приобретения одна геометрическая форма (avCS +) стала ассоциироваться с умеренно болезненным электрическим током (aversive US), другой формой (appCS +) с аппетитным US и третьей формой (CS-), ни с аппетитными США, ни с аверсивные США. Результаты показывают успешную аверсивную и аппетитную обусловленность на явном вербальном уровне (т. Е. Рейтинги), на неявном поведенческом уровне (т. Е. Напугании) и на физиологическом уровне (т. Е. SCR). В частности, avCS + по сравнению с CS-вызвал более отрицательные оценки валентности, более высокие рейтинги возбуждения, потенцию запуска и больший SCR. Самое главное, что appCS + по сравнению с CS - вызвало более положительные оценки валентности, затухание при запуске и больший SCR. Наши выводы об аутсорсивном обучении соответствовали ожиданиям, поскольку в предыдущих исследованиях было установлено, что угроза прогнозирования (avCS +) оценивается как отвратительная, вызывает больший ответ на страх и увеличивает физиологическое возбуждение (Fendt и Fanselow, 1999; Hamm и Weike, 2005; Andreatta и др., 2010, 2013). Наши результаты по аппетитной кондиции также соответствовали предыдущим исследованиям на людях и животных, показывающим, что вознаграждение, предсказывающее стимул (appCS +), оценивается как положительное, ингибирует ответы от страха и увеличивает физиологическое возбуждение (Koch et al., 1996; Готфрид и др., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Остин и Дука, 2010; Prévost и др., 2012). Насколько нам известно, это первое исследование, демонстрирующее условное ослабление ослабления у людей стимулом, предсказывающим основную награду. Важно отметить, что мы смогли перенести и подтвердить результаты исследования на животных (Koch et al., 1996). Это исследование на животных показало, что поражение при старте у крыс зависит от прогнозов от NAcc (часть вентрального полосатого тела) до PnC. Таким образом, ослабление пучка в нашем исследовании может означать NAcc-активность, которая также соответствует результатам МРТ (Готфрид и др., 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Delgado et al., 2011; Леви и Глимчер, 2011). Следовательно, мы заключаем, что наша аппетитная парадигма кондиционирования была успешной, так как указывались как явные (оценки), так и неявная (начальная затухание) положительная валентность.
В дополнение к этому новому, но предсказуемому выводу заслуживают внимания еще два интересных результата. Во-первых, словесные и физиологические реакции возбуждения на appCS + диссоциированы. Во-вторых, мы обнаружили более быстрое исчезновение аппетитных CR по сравнению с аверсивными CR.
SCR указывает на симпатическую активацию, которая увеличивается как на поощрительный стимул (appCS +), так и на связанный с угрозой стимул (avCS +). Этот результат соответствует предыдущему обучению, в котором эротические картины использовались как аппетитные США (Klucken et al., 2013), а также исследование, показывающее сопоставимый SCR с предсказанием денег и подсказкой, предсказывающей аверсивный шум (Остин и Дука, 2010). Примечательно, что SCR является ориентировочным ответом, связанным с активацией симпатической системы. Предполагается, что этот ответ отражает подготовку поведенческой реакции к мотивационно значимым событиям (Брэдли, 2009). Исходя из этого, мы считаем, что как связанные с угрозой, так и связанные с наградами стимулы вызвали подготовительный ответ на последовательные поведенческие реакции. Другими словами, сигналы питания и боли инициировали подготовку к поведению и предотвращению, соответственно. В отличие от высокого физиологического возбуждения словесные ответы указывали на низкое возбуждение стимула, связанного с наградами. Возможно, что устное возбуждение может быть в большей степени подвержено влиянию природы США, а не физиологической активации сам по себе, На самом деле, мы считаем, что appCS + против avCS + оценивались как сдержанно, так и с высокой степенью возбуждения, потому что они были связаны с низким и высоким спросом на США соответственно. К сожалению, мы не собирали рейтинги возбуждения для США, и поэтому это будет в будущем, чтобы проверить эту гипотезу явно.
На этапе исчезновения США не было доставлено. Возможно, это вызвало новое тормозное обучение, называемое вымиранием (для обзора см. Milad и Quirk, 2012), причем как аверсивные, так и аппетитные CR уменьшаются как следствие. Очевидно, что поведенческий (то есть ответ на пуск) и на физиологические (т. Е. SCR) уровни, никакие дифференциальные реакции на avCS +, appCS + и CS- не были обнаружены больше. Точно так же явное возбуждение условных раздражителей, оцененных после фазы экстинкции, уравнилось на низком уровне, что предполагает успешное обучение исчезновению. Однако avCS + по-прежнему оценивался значительно более негативно, чем CS-, тогда как валентность appCS + больше не отличалась от CS-валентности. Более медленное исчезновение отвратительного явного ответа может быть связано с эволюционным консерватизмом, что означает, что сигналы угроз особенно трудно забыть, потому что отказ от ответа на сигнал угрозы может угрожать жизни.
Остается еще один вопрос: почему тогда окончательный ответ (т. Е. Неявная валентность) полностью погас во время фазы экстинкции? Во-первых, эти ответы были рассчитаны на протяжении всего хода фазы. Поэтому вполне возможно, что дискриминационные CR будут все еще обнаруживаться в ходе первых испытаний фазы исчезновения. Экспериментально мы выполнили эту гипотезу и рассмотрели как исходные ответы, так и SCR на фазе экстинкции (см. Дополнительный материал). Хотя мы не обнаружили существенных различий, мы наблюдали несколько более высокую начальную величину для avCS + по сравнению с CS- и новым контрольным стимулом. Мы также наблюдали немного более сильное ослабление в appCS + по сравнению с CS- и новым контрольным стимулом в самом начале фазы исчезновения, которое, однако, исчезло в течение нескольких испытаний. SCR к appCS + упал уже после испытания 2nd extinction, в то время как SCR для avCS + остался выше для почти всех исчезновений по сравнению с SCR в NEW. Хотя мы должны интерпретировать эти результаты с большой осторожностью, ответы на поражение, казалось, совпадали с валентными рейтингами. Более того, как исходные ответы, так и SCR также поддерживают идею эволюционного консерватизма в отношении стимулов угроз.
Наконец, мы должны признать некоторые ограничения этого исследования. Во-первых, из-за технической проблемы мы не смогли сообщить оценки приятности (и возбуждения) для шоколада и соленого кренделя. Тем не менее, аппетитные CR предлагают предположить, что участники действительно испытали эти две США как аппетитные. Во-вторых, продолжительность отвратительного и аппетитного США сильно отличалась. Таким образом, болезненный удар электрическим током был доставлен на смещение avCS + и продолжался ровно 200 мс, а шоколад и соленый крендель были представлены участникам около 2 s после появления appCS +, и продолжительность была неопределенной, поскольку она зависела от того, насколько быстро человек их съел. Выбор для доставки аппетитных USs таким образом был основан на предыдущем исследовании на людях (Prévost и др., 2012). Тем не менее, было бы методологически более элегантно поставлять аппетитные США, более сопоставимые с аверсивными США, такими как глоток сока или имбирный эль при смещении appCS +. Чтобы компенсировать эту большую разницу в восприятии, мы представили геометрические формы в сочетании с визуальным стимулом, символизирующим США. В-третьих, мы не можем однозначно исключить возможность того, что быстро погашенные физиологические реакции на фазе вымирания обусловлены методологическим аспектом. На самом деле визуальные стимулы в этой фазе не были представлены в сочетании с символом США, как это было в фазах сбора. Возможно, что презентация только CS может повлиять на CR, и по этой причине во время первых испытаний на исчезновение существенных различий не было. Однако наши быстрые потушенные ответы согласуются с потушенными ответами в предыдущем исследовании, в котором CS + (лицо) было представлено в соединении с США (крик) во время приобретения, но не во время вымирания (Lissek et al., 2008).
В заключение мы нашли успешные отвратительные и аппетитные условные реакции на стимул, связанный с угрозой, и стимул, связанный с наградой, соответственно. Интересно, что явный (рейтинги) и неявный (начальный рефлекс) уровень ответов работал синергетическим образом, так как avCS + сообщалось как отрицательное и индуцированное потенцирование поражений, а appCS + сообщалось как положительное и индуцированное затухание. Более того, явные (оценки) и физиологическое (SCR) возбуждение appCS + диссоциируют, отражая два отдельных процесса.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Эта работа была поддержана Центром совместных исследований «Страх, тревога и тревожные расстройства», SFB-TRR 58, проект B1.
Дополнительный материал
Дополнительный материал для этой статьи можно найти в Интернете по адресу: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
Рекомендации
Andreatta, M., Mühlberger, A., Glotzbach-Schoon, E., и Pauli, P. (2013). Предсказуемость боли изменяет валентные оценки рельефно-ассоциированного стимула. Фронт. Сист. Neurosci, 7: 53. doi: 10.3389 / fnsys.2013.00053
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Andreatta, M., Mühlberger, A., Yarali, A., Gerber, B., and Pauli, P. (2010). Разрыв между неявной и явной обусловленной валентностью после изучения боли у людей. Proc. Biol. Sci, 277, 2411-2416. doi: 10.1098 / rspb.2010.0103
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Austin, AJ и Duka, T. (2010). Механизмы внимания к аппетитным и отвратительным результатам в обучении в Павлове. Behav. Brain Res, 213, 19-26. doi: 10.1016 / j.bbr.2010.04.019
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Blumenthal, TD, Cuthbert, BN, Filion, DL, Hackley, S., Lipp, OV и van Boxtel, A. (2005). Доклад Комитета: руководящие принципы для электромиографических исследований с поражением глаз. Психофизиология 42, 1-15. doi: 10.1111 / j.1469-8986.2005.00271.x
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Bouton, ME и Peck, CA (1989). Контекстные эффекты при кондиционировании, вымирании и восстановлении в аппетитной подготовке кондиционирования. Anim. Учить. Behav, 17, 188-198. doi: 10.3758 / BF03207634
Полный текст CrossRef | Google Scholar
Брэдли, М.М. (2009). Естественное избирательное внимание: ориентирование и эмоции. Психофизиология 46, 1-11. doi: 10.1111 / j.1469-8986.2008.00702.x
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Clark, JJ, Hollon, NG и Phillips, PEM (2012). Павловские системы оценки в процессе обучения и принятия решений. Тек. ОПИН. Neurobiol, 22, 1054-1061. doi: 10.1016 / j.conb.2012.06.004
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Delgado, MR, Jou, RL и Phelps, EA (2011). Нейронные системы, лежащие в основе аверсивного кондиционирования у людей с первичным и вторичным артериальными усилителями. Фронт. Neurosci, 5: 71. doi: 10.3389 / fnins.2011.00071
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Dickinson, A., and Balleine, B. (1994). Мотивационный контроль целенаправленного действия. Anim. Учить. Behav, 22, 1-18. doi: 10.3758 / BF03199951
Полный текст CrossRef | Google Scholar
Fendt, M., and Fanselow, MS (1999). Нейроанатомическая и нейрохимическая основа условного страха. Neurosci. Biobehav. оборот, 23, 743-760. doi: 10.1016 / j.ijpsycho.2012.09.006
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Готфрид, Дж. А., О'Догерти, Дж., И Долан, Р. Дж. (2002). Аппетивное и аверсивное обонятельное обучение у людей изучали с помощью функциональной магнитно-резонансной томографии, связанной с событиями. J. Neurosci, 22, 10829-10837. Доступно в Интернете по адресу: http://www.jneurosci.org/content/22/24/10829.full
Grillon, C., Baas, JM, Cornwell, B. and Johnson, L. (2006). Контекстная подготовка и предотвращение поведения в среде виртуальной реальности: эффект предсказуемости. Biol. психиатрия 60, 752-759. doi: 10.1016 / j.biopsych.2006.03.072
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Хамм, А. О. и Уэйк, ИИ (2005). Нейропсихология изучения страха и регулирования страха. Int. J. Psychophysiol, 57, 5-14. doi: 10.1016 / j.ijpsycho.2005.01.006
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Klucken, T., Schweckendiek, J., Merz, CJ, Tabbert, K., Walter, B., Kagerer, S., et al. (2009). Нейронные активации приобретения условного сексуального возбуждения: последствия осознания непредвиденных обстоятельств и пола. J. Sex. Med, 6, 3071-3085. doi: 10.1111 / j.1743-6109.2009.01405.x
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Klucken, T., Wehrum, S., Schweckendiek, J., Merz, CJ, Hennig, J., Vaitl, D., et al. (2013). Полиморфизм 5-HTTLPR связан с измененными гемодинамическими ответами во время аппетитного кондиционирования. Hum. Мозг Мапп, 34, 2549-2560. doi: 10.1002 / hbm.22085
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Кох, М. (1999). Нейробиология испуга. Prog. Neurobiol. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Кох, М., Шмид, А. и Шнитцлер, H.-U. (1996). Удовольствие-затухание испуга нарушается при поражениях ядра. Neuroreport 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Krohne, HW, Egloff, B., Kohmann, C.-W. и Tausch, A. (1996). Untersuchungen mit einer deutschen version der «Положительный и отрицательный график влияния» (PANAS). Диагностика 42, 139-156.
Kumar, P., Waiter, G., Ahearn, T., Milders, M., Reid, I. и Steele, JD (2008). Аномальные временные различия в награждении - обучение сигналов в большой депрессии. Мозг 131, 2084-2093. doi: 10.1093 / brain / awn136
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Laux, L., Glanzmann, P., Schaffner, P. и Spielberger, CD (1981). Достопримечательность Даса Angstinventar, Вайнхайм: тест Белца.
Леви, DJ и Glimcher, PW (2011). Сравнение яблок и апельсинов: использование вознаграждения и общего представления субъективной ценности в мозге. J. Neurosci, 31, 14693-14707. doi: 10.1523 / JNEUROSCI.2218-11.2011
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Lissek, S., Levenson, J., Biggs, AL, Johnson, LL, Ameli, R., Pine, DS, et al. (2008). Повышенный страх, обусловливающий социально значимые безусловные стимулы в социальном тревожном расстройстве. Am. J. Психиатрия 165, 124-132. doi: 10.1176 / appi.ajp.2007.06091513
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Martin-Soelch, C., Linthicum, J. и Ernst, M. (2007). Аппетитное кондиционирование: нейронные основы и последствия для психопатологии. Neurosci. Biobehav. оборот, 31, 426-440. doi: 10.1016 / j.neubiorev.2006.11.002
PubMed Аннотация | Полный текст CrossRef | Google Scholar
McDannald, MA, Lucantonio, F., Burke, KA, Niv, Y. и Schoenbaum, G. (2011). Вентральный стриатум и ортофронтальная кора являются необходимыми для обучения на основе моделей, но не без модели. J. Neurosci, 31, 2700-2705. doi: 10.1523 / JNEUROSCI.5499-10.2011
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Milad, MR и Quirk G. (2012). Исчезновение страха как модель трансляционной нейронауки: десять лет прогресса. Annu. Преподобный Психол, 63, 129-111. doi: 10.1146 / annurev.psych.121208.131631
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Павлов И.П. (1927). Условные рефлексы: исследование физиологической активности коры головного мозга, Лондон: издательство Оксфордского университета.
Превост, К., Лильехольм, М., Тышка, Дж. М., и О'Догерти, Дж. П. (2012). Нейронные корреляты специфической и общей передачи Павлова в инструмент в пределах миндалины человека: исследование фМРТ с высоким разрешением. J. Neurosci, 32, 8383-8390. doi: 10.1523 / jneurosci.6237-11.2012
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Tranel, D., и Damasio, H. (1994). Нейроанатомические корреляты электродермальной реакции проводимости кожи. Психофизиология 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
PubMed Аннотация | Полный текст CrossRef | Google Scholar
Ключевые слова: классическое обусловливание, награда, наказание, рефлекс страха, реакция проводимости кожи
Образец цитирования: Andreatta M and Pauli P (2015) Привлекательный подход к аверсивному обучению у людей. Фронт. Behav. Neurosci. 9: 128. doi: 10.3389 / fnbeh.2015.00128
Поступило в редакцию: 11 February 2015; Принято: 05 May 2015;
Опубликовано: 19 Май 2015.
Под редакцией:
Нильс Бирбаумер, Университет Тюбингена, Германия
Рассмотрено:
Тим Клюкен, Университет Юстуса Либига Гиссен, Германия
Алессандро Ангрилли, Университет Падуи, Италия
Copyright © 2015 Andreatta и Pauli. Это статья с открытым доступом, распространяемая в соответствии с условиями Лицензия Creative Commons Attribution (CC BY), Использование, распространение или воспроизведение на других форумах разрешено при условии, что оригинальный автор (ы) или лицензиар зачисляются и что оригинальная публикация в этом журнале цитируется в соответствии с принятой академической практикой. Запрещается использование, распространение или воспроизведение, которое не соответствует этим условиям.
* Корреспонденция: Марта Андрота, факультет психологии (биологическая психология, клиническая психология и психотерапия), Университет Вюрцбурга, Маркусштрассе 9-11, D-97070 Вюрцбург, Германия, [электронная почта защищена]
