Фронт. Behav. Neurosci., 23 March 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 Масрор Р. Шариф1, Арнаулд Белмер1, Мэтью Дж. Фогарти2, Erica WH Mu2,
Масрор Р. Шариф1, Арнаулд Белмер1, Мэтью Дж. Фогарти2, Erica WH Mu2, 

- 1Институт трансляционных исследований и Институт здоровья и биомедицинских инноваций, Квинслендский технологический университет, Брисбен, QLD, Австралия
- 2Школа биомедицинских наук, Университет Квинсленда, Брисбен, QLD, Австралия
Современная диета стала очень подслащенной, что привело к беспрецедентному уровню потребления сахара, особенно среди подростков. Известно, что хроническое долгосрочное потребление сахара способствует развитию метаболических расстройств, включая ожирение и диабет типа II, мало известно о прямых последствиях длительного употребления алкоголя в мозгу в мозге. Вчто сахар может вызвать высвобождение допамина в ядре accumbens (NAc) аналогично наркотикам, мы исследовали изменения в морфологии нейронов в этой области мозга после коротких (4 недель) и длительных (12 недель) выпивок - как потребление сахарозы, используя прерывистую парадигму выбора с двумя бутылками. Мы использовали окрашивание Golgi-Cox для пропитки средних колючих нейронов (MSN) из ядра NAc и оболочки коротко- и долгосрочных сахарозы, потребляющих крысы, и сравнивали их с контрольными по возрасту контрольными веществами. Мы показываем, что длительное потребление по-закусной сахарозы значительно уменьшало общую длину дендритных оболочек ДНК NAc по сравнению с контрольными крысами с возрастом. Мы также обнаружили, что реструктуризация этих нейронов в основном обусловлена уменьшением дистальной дендритной сложности. Напротив, мы наблюдали увеличение плотности позвоночника на дистальных порядках ветвления ДНК-оболочки NAc у долгосрочных потребляющих сахарозу крыс. В сочетании эти результаты подчеркивают эффекты нейронов длительного заушного приема сахарозы на морфологию оболочки NAc shell.
Введение
За последние 40 лет был зафиксирован рост потребления сахаристых напитков и продуктов, содержащих добавленные сахара (Nielsen и др., 2002; Попкин, 2010; Ng и др., 2012), с отчетами, оценивающими, что до 75% всех продуктов питания и напитков содержат большое количество добавленных сахаров (Ford и Dietz, 2013; Брей и Попкин, 2014). В течение этого периода также наблюдалось одновременное увеличение распространенности ожирения и диабета типа II, особенно среди подростков (Арсланян, 2002; Reinehr, 2013; Dabelea и др., 2014; Fryar и др., 2014). Недавние исследования показали, что дети с избыточным весом и ожирением часто потребляют большое количество добавленного сахара, однако вклад диет с высоким содержанием сахара в увеличение заболеваемости избыточным весом и ожирением детей остается спорным (Ху, 2013; Брей и Попкин, 2014; Bucher Della Torre и др., 2015).
В то время как растущее количество доказательств свидетельствует о том, что потребление диеты с высоким содержанием сахара может частично способствовать увеличению веса среди детей и подростков (Malik et al., 2010; Te Morenga и др., 2013; Брей и Попкин, 2014), меньше внимания уделяется неблагоприятным не-метаболическим последствиям, возникающим из-за чрезмерного потребления сахара. Интересно, что некоторые распространенные поведенческие и психологические закономерности часто возникают среди подгруппы тех, кто переваривает и поддерживает диеты с высоким содержанием сахара. Наиболее заметными являются развитие расстройств пищевого поведения, включая выпивку, в сочетании с одновременным началом психологических симптомов, включая отсутствие мотивации и депрессии (см. Шихан и Герман, 2015). Кроме того, поскольку люди, употребляющие алкоголь, часто демонстрируют потерю контроля и неспособность самостоятельно ограничить потребление сахара, вполне вероятно, что это поведение возникает в результате неврологических адаптаций в областях мозга, которые оценивают гедоническую ценность очень вкусной пищи (Saper et al., 2002; Люттер и Нестлер, 2009; Kenny, 2011). Это обоснование также подтверждается доказательствами на людях, демонстрирующими, что сахар и сладость могут вызывать тягу, которая похожа на тягу, вызванную привыканием к наркотикам, таким как алкоголь и никотин (Volkow и др., 2012).
Хотя аддиктивные свойства сахара все еще являются спекулятивными, эти наблюдения сочетаются с исследованиями, демонстрирующими вклад чрезмерного потребления сахара в изменения схемы вознаграждения и развития привыкания к привыканию и эмоциональных состояний на животных моделях (Avena и др., 2008; Бентон, 2010; Ventura et al., 2014), требует дальнейшего изучения. Предыдущие исследования грызунов показали, что прерывистый доступ к сахарозе изменяет активность нескольких нейротрансмиттеров внутри мезолимбической системы, включая допамин, опиоиды и ацетилхолин (см. Avena и др., 2008). Было показано, что потребление пигментной сахарозы облегчает высвобождение допамина в ядре accumbens (NAc), подобно лекарственным средствам злоупотребления (Avena и др., 2008). Кроме того, мы показали, что долгосрочное потребление сахарозы с использованием 24 h прерывистого доступа с двумя бутылками парадигмы выбора (Simms и др., 2008) модулирует экспрессию никотинового рецептора ацетилхолина (nAChR) в NAc (Шариф и др., В прессе). Интересно отметить, что мы также наблюдали, что соединения nAChR, известные модуляции активности допамина и ацетилхолина в NAc, оказывают различное влияние на потребление сахарозы после кратковременного и длительного приема (Шариф и др., В прессе).
Хотя эти исследования продемонстрировали сходство в поведенческих и нейрохимических изменениях, вызванных прерывистым доступом к сахару и наркотикам, не известно, облегчают ли эти эффекты изменения морфологии нейронов в NAc. Tего отличие от веществ злоупотребления, включая кокаин, амфетамин и никотин, которые производят хорошо охарактеризованные изменения в морфологии средних колючих нейронов (MSN) в NAc, включая повышенную плотность позвоночника и измененную дендритную сложность (Робинсон и Колб, 1999, 2004; Li et al., 2003; Crombag et al., 2005). Поскольку мы ранее показали, что длительное воздействие (неделя 12) на спирт и сахарозу с использованием прерывистой парадигмы выбора с двумя бутылками дает дифференциальный ответ на фармакотерапевтические вмешательства по сравнению с краткосрочным потреблением (4 недель; Steensland et al., 2007; Шариф и др., В прессе), мы оценили влияние краткосрочного и долгосрочного потребления сахарозы на морфологию MSN в NAc. Мы разрешали крысам-подросткам потреблять сахарозу по-разному для 4 (краткосрочных) или 12 (долгосрочных) недель, а затем анализировали морфологию NAc MSN от кратковременных и долгосрочных сахарозы, потребляющих крысы, и сравнивали это с контролируемые возрастом, которым был предоставлен доступ только к воде. Наши результаты показывают, что MSN из оболочки NAc изменяются после длительного, но не кратковременного потребления сахарозы, уменьшая длину дендритов, но увеличивая плотность дистального дендритного позвоночника. Кроме того, мы обнаружили, что морфология MSN с ядра NAc оставалась относительно неповрежденной после краткосрочного и долгосрочного потребления сахарозы. Эти результаты подчеркивают прямое неврологическое последствие длительного потребления сахарозы в виде выпивки. Кроме того, эти данные демонстрируют необходимость дальнейших исследований, направленных на выяснение молекулярных и нейрохимических изменений, которые сопровождают морфологическую реструктуризацию НС оболочек щитовидной железы, вызванных пролонгированным, выпитообразным поглощением сахарозы.
Материалы и методы
Заявление о этике
Все экспериментальные процедуры проводились в соответствии с Австралийским кодексом по уходу и использованию животных для научных целей, 8th Edition (Национальный совет по здравоохранению и медицинским исследованиям, 2013). Протоколы были одобрены Комитетом по этике животных в Квинсленде и Комитетом по этике животных в Квинсленде.
Животные и жильё
Пятинедельные (подростковые) самцы крыс Wistar (контроль: 176.4 ± 4.8 g; Sucrose: 178.3 ± 5.0 g) (ARC, WA, Australia) были индивидуально размещены в вентилируемом двухслойном плексигласе® садки. Крысы были акклиматизированы для индивидуальных условий проживания, обработки и обратного света за 5 за несколько дней до начала экспериментов. Все крысы были размещены в контролируемом климатом 12-hr обратном свете / темном цикле (выключается в 9 am) комнате со стандартной крысиной чау и доступной водой вволю.
Парадигма питьевого доступа с двумя бутылками
Временная доступность 5% сахарозы с двумя бутылочными вариантами питьевой парадигмы (Simms и др., 2008) была адаптирована из Мудрый (1973), Все жидкости были представлены в пластмассовых бутылках 300 мл с напитками из нержавеющей стали, вставленными через две прокладки в передней части клетки после начала цикла темного света. Вес каждой бутылки регистрировали до презентации бутылки. Две бутылки были представлены одновременно: одна бутылка, содержащая воду; второй флакон, содержащий 5% (мас. / об.) сахарозы. Размещение бутылки сахарозы 5% (мас. / Об.) Чередуется с каждым воздействием контроля на боковые предпочтения. Бутылки взвешивали 24 h после подачи жидкостей и проводили измерения до ближайшего 0.1 g. Вес каждой крысы также измеряли для расчета граммов потребления сахарозы на килограмм массы тела. В день 1 периода выпивки крысы (n = 6-9) получили доступ к одной бутылке сахарозы 5% (мас. / Об.) И одной бутылке воды. После 24 h бутылку сахарозы заменяли второй бутылкой с водой, которая была доступна для следующего 24 h. Эта картина повторялась по средам и пятницам. Крысы имели неограниченный доступ к воде в другие дни. Потребление сахарозы, подобное употреблению, привело к эскалации общего потребления сахарозы (мл) с течением времени (дополнительная диаграмма 1) и сопровождался стабильными исходными уровнями питья в расчете на массу тела [20 ± 5 г / кг 5% (мас. / об.)] в течение коротких [~ 4 недель (сеансы употребления 13)] и долгосрочные [ ~ 12 недель (сеансы питья 37)]. Отдельная группа контрольных крыс (n = 6-9) получили доступ к воде в обеих бутылках (то есть без сахарозы) в тех же условиях, которые описаны выше. Средняя масса тела контрольных и потребляющих сахарозу крыс в конце краткосрочного воздействия составляла 405.7 ± 40.8 g и 426.4 ± 31.2 g соответственно. В конце длительного воздействия средний вес тела для групп контроля и сахарозы составлял 578.8 ± 53.4 g и 600.2 ± 45.2 g.
Окрашивание Гольджи-Кокса
После последнего сеанса питья крысы были перенесены с животного объекта, чтобы обеспечить обработку образцов головного мозга на объекте гистологии в Школе биомедицинских наук Университета Квинсленда (Сент-Люсия, Австралия). Все одобренные меры были приняты для снижения стресса во время транспортировки, после чего крысам разрешалось восстанавливаться в течение ночи. На следующий день крыс умерщвляли передозировкой пентобарбиталом натрия (60-80 мг / кг, ip Vetcare, Брисбен, Австралия) и внутрисердечно перфузировали искусственной спинномозговой жидкостью 300 мл, которая содержала (в мМ): 130 NaCl, 3 KCl, 26 NaHCO3, 1.25 NaH2PO4, 5 MgCl2, 1 CaCl2, и D-глюкозы 10. Затем каждое животное декапитировали и мозг удаляли и инкубировали в темноте в растворе Гольджи-Кокса, который содержал 5% дихромата калия, 5% хромата калия и 5% хлорида ртути (все химические вещества от Sigma-Aldrich), которые были сделаны свежими 3-днями до жертвоприношения, как описано ранее (Rutledge и др., 1969). Методы инкубации пятен и постобработки Golgi-Cox были изменены из Ranjan and Mallick (2010), Мозги от кратковременных потребляющих сахарозу животных инкубировали в течение 6-дней при 37 ° C, в то время как мозг от долгосрочных потребляющих сахарозу животных инкубировали в течение дней 10 с одним изменением на свежий раствор Гольджи-Кокса после 4 дней инкубации.
После инкубации корональные срезы 300 мкм разрезали с использованием вибрирующего микротома Zeiss Hyrax V50 (Carl Zeiss, Германия). Затем срезы помещали последовательно в планшеты с 24-лунками, заполненные 30% (мас. / Об.) Сахарозой в забуференном фосфатом солевом растворе 0.1 M и обрабатывали, как описано в (Ranjan and Mallick, 2010). Вкратце разделы дегидратировали в 50% этаноле для 5 мин, затем помещали в 0.1 M NH4OH для 30 мин, дважды промывали дистиллированной водой в течение 5 мин и помещали в фиксатор пленки Fujihunt (Fujifilm, Singapore) для 30 мин в темноте. Затем срезы дважды промывали в дистиллированной воде для каждого 2 мин и обезвоживали в 70, 90, 95 и 100% этаноле дважды по 5 мин каждый. Затем секции были очищены в растворе CXA (1: 1: 1 хлороформ: ксилол: спирт) для 10 мин и смонтированы в DPX (Sigma-Aldrich) на слайдах Superfrost Plus (Menzel-Glaser, Lomb Scientific, Австралия) и покрыты покровным (Мензель-Глейзер, Германия). Слайды оставляли в темноте для сушки при комнатной температуре в течение ночи.
Выбор и отслеживание нейронов в пределах Nucleus Accumbens
Корональные срезы между bregma + 2.8 и + 1.7 были обследованы для MSNs внутри ядра и оболочки NAc, используя боковой желудочек и переднюю комиссуру в качестве ориентиров с помощью атласа мозга крысы (Паксинос и Ватсон, 2007) (Рисунок 1). Контурная функция в Neurolucida 7 (MBF Bioscience, VT, USA) использовалась для демаркации ядра NAc и оболочки NAc на каждом фрагменте (рис. 2). Между нейронами 2 и 9 на одну область на одно животное были прослежены параметры дендритной длины с использованием объектива 63x или плотности позвоночника (сообщено как шипы на 100 мкм) с использованием цели 100x на Zeiss Axioskop II (Carl Zeiss, Германия) с использованием автоматизированного хуг этап под управлением Neurolucida® 7 (MBF Biosciences, VT, США). Все следы проводились вслепую по отношению к лечению. Морфологические параметры импрегнированных нейронов Гольджи-Кокса анализировали аналогично предыдущим сообщениям (Klenowski et al., 2015).
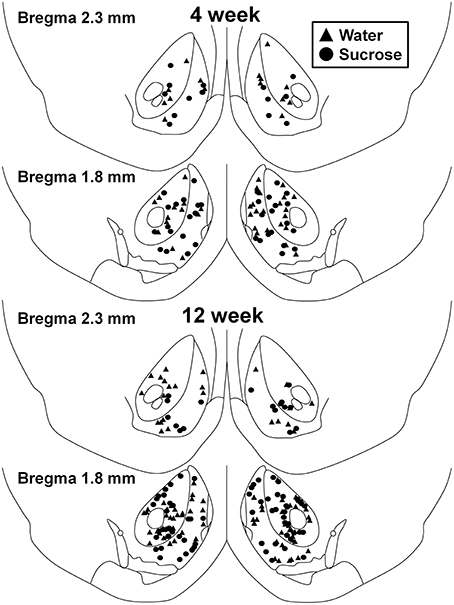
Рисунок 1. Карта, показывающая местоположения средних колючих нейронов, отобранных из ядра ядра и оболочки 4 и 12, потребляющих сахарозу, и контролируемых возрастом, На двух верхних панелях показаны местоположения нейронов, отобранных из ядра ядра и оболочки 4 (треугольники) и сахарозы (круги) животных. В нижних двух панелях показаны положения нейронов, отобранных из контрольной недели 12 (треугольники) и сахарозы (круги) животных.
Статистический анализ
Среднюю и стандартную ошибку среднего (SEM) рассчитывали для каждого набора данных с животным как n, используя средние данные морфометрии от всех ядер или оболочек NAc MSNs (n = 7 для оболочки NAc и n = 6 для ядра NAc 4-неделя, n = 9 для 12-недельных групп). Где указано, непарные двуххвостые Студенческие t- тесты или двухсторонние ANOVA с пост-тестами Bonferroni были проведены для всех анализов, связанных с сравнением групповых средств, с использованием GraphPad Prism версии 6.02 (GraphPad Software, Сан-Диего, Калифорния). Статистическая значимость была принята на P <0.05. Все данные в разделе результатов представлены как средние значения ± SEM. Процентные изменения рассчитываются относительно контрольного значения.
Итоги
Средние колючие нейроны из оболочки Nucleus Accumbens уменьшают длину дендритов, уменьшают дендритную сложность, но увеличивают среднюю плотность позвоночника при дистальных отростчатых порядках после длительного, но не краткосрочного потребления сахарозы
После кратковременных (4 недель) потребления сахарозы существенных различий в морфометрических параметрах оболочки NAc оболочки (Таблица 1). Также не было существенных различий между кратковременным потреблением сахарозы и контролем воды NAc с оболочкой NAc в анализах, связанных с порядком центробежной ветви. А именно, дендритные сегменты на порядок ветвлений (P = 0.4111), средняя дендритная длина на порядок ветвлений (P = 0.5581) и средней плотности позвоночника на порядок ветвления (P = 0.2977, двухсторонние ANOVA) не были существенно различны между группами. Карта местоположения, показывающая приблизительные положения отобранных нейронов, показана на рисунке 1.
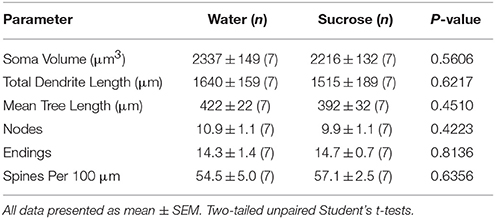
Таблица 1. Общие морфологические параметры средних колючих нейронов из раковины accumbens оболочки кратковременных сахарозы, потребляющих крысы, и контролируемых по возрасту контролей воды.
После длительных (12 недель) потребления сахарозы общая длина древовидной длины нитей оболочки NAc уменьшилась на 21% по сравнению с контролем потребления воды (вода: 1827 ± 148 мкм, n = 9; Сахароза 1449 ± 78 мкм, n = 9, *P = 0.0384, двусторонняя непарная Студенческая t-test, Figure 2, Таблица 2). Сравнение среднего числа дендритных бифуркаций (узлов) и дендритных окончаний между группами воды и сахарозы показало снижение (хотя и незначительного) уровня дендритной сложности в оболочках NAc shell (узлы: вода 12.9 ± 1.4 n = 9, Sucrose 10.1 ± 0.8 n = 9, P = 0.0879; окончания: Вода 17.9 ± 1.4 n = 9, Sucrose 14.8 ± 0.7 n = 9, P = 0.0657, двусторонняя непарная Студенческая t-test, Table 2). Не было изменений в объеме сомы (P = 0.9400), средняя длина дендритного дерева (P = 0.1646) или общей плотности позвоночника (P = 0.3662) в оболочках NAc от долгосрочных сахарозы, потребляющих крысы по сравнению с контролем воды. Эти морфометрические параметры подробно описаны в таблице 2.
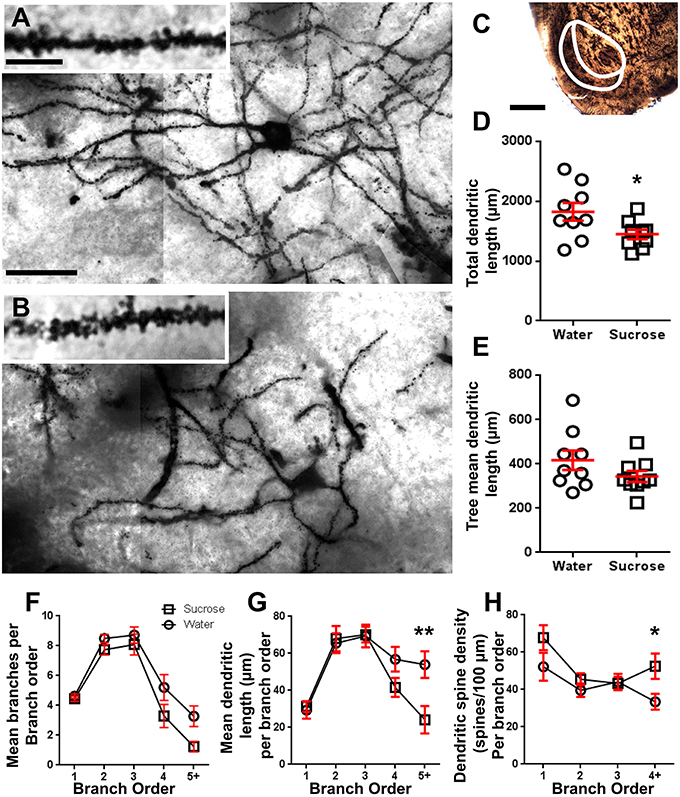 Рисунок 2. Снижение длины дендритной беседки и увеличение дистальной плотности дендритного позвоночника средних колючих нейронов (MSN) из оболочки ядра (NAc) у крыс с длительной обработкой сахарозой по сравнению с контрольными крысами. (А, В) показывают представления контрольной (верхней) и долгосрочной (12 недели) сахарозы (снизу) обработанных ярких полей z-статическая мозаика пропитанных Гольджи MSN из оболочки NAc (увеличение 63x). Вставка (А, В) показывает контроль и долгосрочные обработанные сахарозой яркие изображения пропитанных Гольджи дендритов MSN и дендритных шипов из оболочки NAc (увеличение 100x). (С) показывает анатомические области, в которых были отобраны MSN в этом исследовании. (D) показывает рассеянную диаграмму уменьшенной общей дендритной беседки MSN (среднее ± SEM) из оболочки NAc у долгосрочных животных сахарозы (квадраты) по сравнению с контрольными (кругами), непарными учениками t-тестовое задание, *P <0.05, n = 9; контроля и n = 9; 12 неделя сахароза. (Е) показывает разброс неизмененной средней длины дендритного дендрита (средняя ± SEM) у ДНК NAc у долгосрочных сахарозных животных (квадраты) по сравнению с контрольными (кругами), непарными учениками t-тестовое задание, P > 0.05, n = 9; контроля и n = 9; 12 неделя сахароза. Анализ порядка филиала (среднее ± SEM) номера дендритного сегмента на порядок ветвлений (F), средняя дендритная длина на порядок ветвей (G) и плотность дендритного позвоночника на порядок ветвей (H), Длительное потребление сахарозы уменьшало дендритную длину при дистальных отрядах (5 +) и увеличивала плотность дендритного позвоночника при дистальных отрядах (4 +) по сравнению с контрольными (G, H), двухсторонние ANOVA с пост-тестами Bonferroni, *P <0.05 **P <0.01, n = 9; контроля и n = 9; долгосрочная сахароза. Масштабные бары: (A, B) = 20 мкм; вставка (A, B) = 10 мкм; (С) = 1 мм.
Рисунок 2. Снижение длины дендритной беседки и увеличение дистальной плотности дендритного позвоночника средних колючих нейронов (MSN) из оболочки ядра (NAc) у крыс с длительной обработкой сахарозой по сравнению с контрольными крысами. (А, В) показывают представления контрольной (верхней) и долгосрочной (12 недели) сахарозы (снизу) обработанных ярких полей z-статическая мозаика пропитанных Гольджи MSN из оболочки NAc (увеличение 63x). Вставка (А, В) показывает контроль и долгосрочные обработанные сахарозой яркие изображения пропитанных Гольджи дендритов MSN и дендритных шипов из оболочки NAc (увеличение 100x). (С) показывает анатомические области, в которых были отобраны MSN в этом исследовании. (D) показывает рассеянную диаграмму уменьшенной общей дендритной беседки MSN (среднее ± SEM) из оболочки NAc у долгосрочных животных сахарозы (квадраты) по сравнению с контрольными (кругами), непарными учениками t-тестовое задание, *P <0.05, n = 9; контроля и n = 9; 12 неделя сахароза. (Е) показывает разброс неизмененной средней длины дендритного дендрита (средняя ± SEM) у ДНК NAc у долгосрочных сахарозных животных (квадраты) по сравнению с контрольными (кругами), непарными учениками t-тестовое задание, P > 0.05, n = 9; контроля и n = 9; 12 неделя сахароза. Анализ порядка филиала (среднее ± SEM) номера дендритного сегмента на порядок ветвлений (F), средняя дендритная длина на порядок ветвей (G) и плотность дендритного позвоночника на порядок ветвей (H), Длительное потребление сахарозы уменьшало дендритную длину при дистальных отрядах (5 +) и увеличивала плотность дендритного позвоночника при дистальных отрядах (4 +) по сравнению с контрольными (G, H), двухсторонние ANOVA с пост-тестами Bonferroni, *P <0.05 **P <0.01, n = 9; контроля и n = 9; долгосрочная сахароза. Масштабные бары: (A, B) = 20 мкм; вставка (A, B) = 10 мкм; (С) = 1 мм.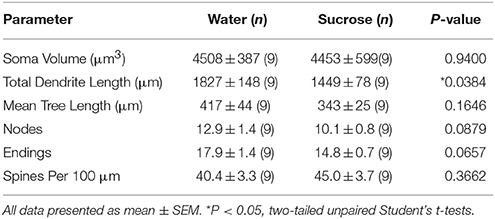
Таблица 2. Общие морфологические параметры средних колючих нейронов из раковины accumbens у многолетних сахарозы, потребляющих крысы, и контролируемых по возрасту контролей воды.
Следуя характеристике общей дендритной морфологии долговременной сахарозы, потребляющей NAc shell MSNs, мы проанализировали дендритные арборизации и плотности позвоночника в отношении их характеристик порядка ветвления. Наша всесторонняя оценка дендритных деревьев определяла количественно количество дендритных сегментов на порядок ветвей, среднюю длину дендритных сегментов по порядку ветвления и среднюю плотность позвоночника на порядок ветвления оболочки NAc для контроля воды и долгосрочных потребляющих сахарозу крыс. Краткое описание данных и анализа заказа филиала представлено в таблице 3.
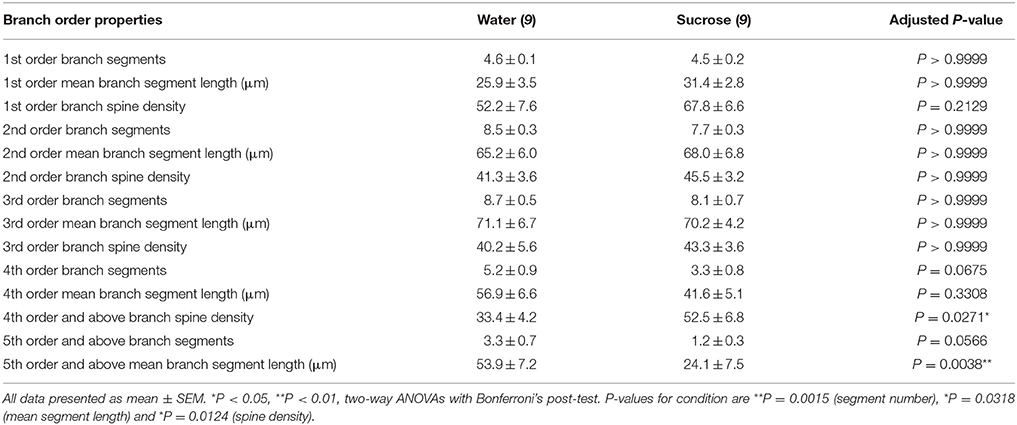 Таблица 3. Специфические характеристики отделения средних колючих нейронов от многолетних сахарозы и водяных крыс.
Таблица 3. Специфические характеристики отделения средних колючих нейронов от многолетних сахарозы и водяных крыс.Среднее число сегментов дендритной ветви на порядок ветвей ДНК-оболочки NAc значительно уменьшалось у длительных крыс, потребляющих сахарозу, по сравнению с контролем воды (**P = 0.0015, двухсторонняя ANOVA). Пост-тесты Бонферрони показали тенденцию к уменьшенному числу сегментов ветви на 4th (Вода: 5.2 ± 0.9, n = 9; Сахароза 3.3 ± 0.8, n = 9, P = 0.0675, Рисунок 2F, Таблица 3), и 5-й порядок и выше порядки ветвления (Вода: 3.3 ± 0.7, n = 9; Сахароза 1.2 ± 0.3, n = 9, P = 0.0566, Рисунок 2F, Таблица 3). Средняя длина дендритного сегмента по одному порядку размножения ДНК NAc оболочки также была значительно снижена у длительных крыс, потребляющих сахарозу, по сравнению с контролем воды (*P = 0.0444, двухсторонняя ANOVA). Пост-тесты Bonferroni показали уменьшение 55% в филиалах 5-го порядка и далее (Вода: 53.9 ± 7.2 мкм, n = 9; Сахароза 24.1 ± 7.5 мкм, n = 9, **P = 0.0038, Рисунок 2G, Таблица 3).
Анализ распределения отделений показал значительное увеличение плотности дендритного позвоночника в раковинах NAc у пациентов с длительной потребляющей сахарозой крыс по сравнению с контрольными (*P = 0.0124, двухсторонняя ANOVA). Пост-тесты Bonferroni показали увеличение плотности позвоночника 57% в дистальных ветвях 4-го порядка и далее (Вода: 33.4 ± 4.2, n = 9; Сахароза 52.5 ± 6.8, n = 9, P = 0.0271 *, вставка цифр 2A, В, Н, Таблица 3). Репрезентативные изображения общей архитектуры MSN и плотности дистального позвоночника (вставка) изображены на рисунках 2A, В.
В совокупности эти результаты показывают, что кратковременное потребление сахарозы мало влияет на морфологические параметры MSN в оболочке NAc. Однако после длительного потребления наблюдается значительное уменьшение длины и сложности нейронной беседки, особенно в дистальных дендритных ветвях. Сопутствующая дистальная плотность позвоночника увеличивается также проявляется в оболочках NAc оболочки длительных сахарозы, потребляющих крысы.
Средние колючие нейроны ядра ядра Nucleus уменьшают сложность ветвления после длительного, но не краткосрочного потребления сахарозы
После кратковременного потребления сахарозы существенных различий в морфометрических показателях сердечной недостаточности NAc (таблица 4). Также не было выявлено существенных различий между потреблением сахарозы 4 и контрольным ядром МСВ в анализах, относящихся к порядку центробежной ветви. А именно, дендритные сегменты на порядок ветвлений (P = 0.7717), средняя дендритная длина на порядок ветвлений (P = 0.2096) и средняя плотность позвоночника на порядок ветвлений (P = 0.3521, двухсторонние ANOVA) не различались между группами.
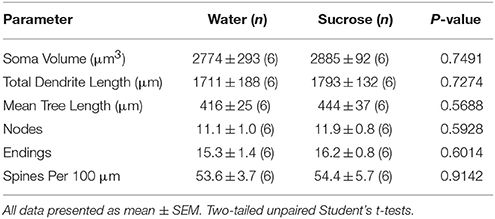 Таблица 4. Общие морфологические параметры средних колючих нейронов ядра ядра, поглощающего кратковременные сахарозы, потребляющие крысы и контролируемые возрастной контроль воды.
Таблица 4. Общие морфологические параметры средних колючих нейронов ядра ядра, поглощающего кратковременные сахарозы, потребляющие крысы и контролируемые возрастной контроль воды.Длительное потребление сахарозы также не имело существенного влияния на морфометрические параметры сердечника MSN NAc (Таблица 5). Среднее число сегментов дендритной ветви на порядок ветвей NAc основных MSNs было значительно снижено у длительных крыс, потребляющих сахарозу, по сравнению с контролем воды (*P = 0.0416, двухсторонний ANOVA), однако не было существенных различий в средней дендритной длине на порядок ветвлений (P = 0.0995) и средней плотности позвоночника на порядок ветвления (P = 0.4888, двухсторонние ANOVAs) между MSNs в ядре NAc у многолетних крыс, потребляющих сахарозу, по сравнению с контролем воды. В совокупности наши данные показывают, что ядро NAc не так реагирует на долгосрочное потребление сахарозы по сравнению с MSN из области оболочки NAc.
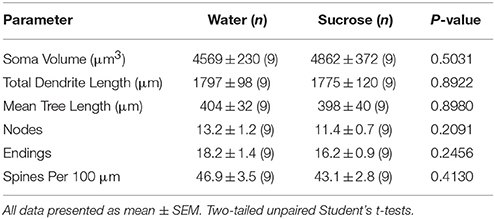 Таблица 5. Общие морфологические параметры средних колючих нейронов из ядра ядра ядро долгоживущих сахарозы, потребляющих крысы, и контролируемых возрастом воды.
Таблица 5. Общие морфологические параметры средних колючих нейронов из ядра ядра ядро долгоживущих сахарозы, потребляющих крысы, и контролируемых возрастом воды.Обсуждение
Увеличение доступности сильно подслащенных продуктов питания в западной диете не только способствовало увеличению распространенности и экономического бремени ожирения и диабета типа II, но и привело к возникновению расстройств пищевого поведения, таких как выпивка (Swanson и др., 2011; Kessler et al., 2013; Дэвис, 2015). Хотя аддиктивные свойства сахаров, включая фруктозу и сахарозу, остаются спекулятивными, наблюдается поразительное сходство в поведенческих и нервных коррелятах, которые проявляются в результате чрезмерного употребления в пищу и длительного употребления наркотиков (Avena и др., 2008, 2011). Кроме того, сахар активирует схему вознаграждения мозга аналогично наркотикам, вызывающим злоупотребление (Volkow и др., 2012), а результаты исследований на людях свидетельствуют о том, что сахар и сладость могут вызывать тягу, сравнимую по величине с теми, которые вызываются аддиктивными лекарственными средствами, такими как алкоголь и никотин (Volkow и др., 2012). Поэтому мы использовали модель потребления бинго-сахарозы у крыс для определения эффектов коротких (4 недель) и долгосрочного (12 недель) потребления сахарозы на морфологию нейронов MSN в NAc, ключевом компоненте схемы перекрывающихся вознаграждений который модулируется сахаром и наркотиками. Мы показываем, что у MSN из оболочки NAc у хронических многолетних сахарозы, потребляющих крысы, значительно снижается дендритная длина и сложность, но увеличивается дистальная плотность дендритного позвоночника. Долгосрочное потребление сахарозы не влияло на морфологию MSN из ядра NAc, в то время как кратковременное потребление сахарозы также не оказывало существенного влияния на морфологию MSN из ядра NAc или оболочки. Эти результаты не только демонстрируют прямой эффект длительного заедания, как прием сахарозы на морфологию нейронов оболочек NAc shell, но также указывают на потенциально вредные последствия длительного потребления диетических продуктов с высоким содержанием сахара.
NAc, который является частью брюшного полосатого тела, состоит в основном из MSNs, которые морфологически характеризуются как нейроны среднего размера с обширными дендритными арборизациями и высокой плотностью позвоночника (Кемп и Пауэлл, 1971; Graveland и DiFiglia, 1985; Rafols и др., 1989; Kawaguchi et al., 1990). Glutamatergic и допаминергические нейроны являются двумя основными афферентными входами в NAc, в первую очередь с контактом с дендритными валами и шипами MSN (Groves, 1980; Кайя и Намба, 1981; Groves и др., 1994). В частности, оболочка и сердечник NAc получают глутаматергический вход от функционально отличных областей коры (Brog et al., 1993). Оболочка NAc также иннервируется возбуждающими афферентами из подкорковых областей, таких как гиппокамп, таламус и базалатеральная амигдала (Brog et al., 1993; Райт и Гроненеген, 1995). Предыдущие исследования продемонстрировали, что эти глютаматергические факторы играют ключевую роль в мотивации и целенаправленном поведении, таких как поиск продуктов питания и вознаграждений (Maldonado-Irizarry и др., 1995; Kelley и Swanson, 1997; Рейнольдс и Берридж, 2003; Ричард и Берридж, 2011). Другим преобладающим вкладом в NAc MSN является от дофаминергических афферентов, которые выступают из брюшной тегментальной области (Lindvall и Björklund, 1978; Veening и др., 1980; Каливас и Миллер, 1984). Интересно отметить, что в предыдущих исследованиях, в которых использовались сходные модели прерывистого доступа к сахару, было показано, что полученное в результате подобное выпивке потребление приводит к увеличению внеклеточного допамина в NAc аналогично (хотя и в меньшей степени) к наркотикам (Rada и др., 2005; Avena и др., 2006) и может модулировать экспрессию рецептора допамина (Colantuoni и др., 2001, 2002) в ядре NAC и оболочке. Интересно, что подобное употреблению сахарозы потребление, вызванное выпивкой, вызывает эскалацию потребления во времени аналогично самоуправлению наркотических средств, таких как кокаин и героиn (Ахмед и Кооб, 1998; Ahmed и др., 2000, 2003), что связано с развитием «зависимого типа».
Наш анализ морфометрии ветвящегося порядка показывает, что общее снижение дендритной длины ДНК-оболочки NAc, вызванное долгосрочным потреблением сахарозы, в основном связано с уменьшением сложности дистальных порядков ветвления. Мы наблюдали уменьшение дистального ветвления (порядки 4th и 5th порядка и выше) и значительно уменьшенную среднюю длину в 5-м порядке и выше дендритов в сочетании с увеличенной плотностью позвоночника при этих порядках ветвления. Общим фактором, который может повлиять на этот тип дендритной реструктуризации, является изменение синаптической связи и / или функции (Russo и др., 2010). Предыдущие исследования показали, что глутаматергические синапсы у MSN формируются, главным образом, на шипах, особенно на дистальных дендритах (Groenewegen et al., 1999). Кроме того, совместная локализация дофаминовых и глутаматергических входов из префронтальной коры (Sesack и Pickel, 1992), гиппокамп (Тотерделл и Смит, 1989; Sesack и Pickel, 1990), и миндалины (Johnson et al., 1994) наблюдались на дендритных шипах MSN. Эти наблюдения в сочетании с увеличением плотности позвоночника после длительного потребления сахарозы, наблюдаемого в нашем исследовании, способствуют формированию увеличенных возбуждающих входов. Поэтому возникает вероятность того, что стойкие эффекты, вызванные затяжным поглощением сахарозы, могут способствовать усилению возбуждающей синаптической активности у дистальных дендритов MSN в оболочке NAc. Следовательно, редукция и / или ретракция дистальных дендритов может быть результатом синаптического гомеостатического механизма (Рейсснер и Каливас, 2010), однако это еще предстоит определить.
Интересно отметить, что Crombag и его коллеги показали, что не было увеличения плотности позвоночника в оболочке NAc после потребления сахарозы в течение 4 недель через парадигму самообслуживания носа, несмотря на более надежное получение и более высокий коэффициент ответа на сахарозу при сравнении с амфетамином (Crombag et al., 2005). Их наблюдение за отсутствием изменения плотности позвоночника в 4-недели отражает наши результаты. Напротив, однако, наше исследование показывает, что после длительного (12 week) воздействия хронического потребления сахарозы наблюдается значительное увеличение плотности дистального отдела позвоночника у MSNs у сахароз-опытных крыс. Кроме того, наша лаборатория ранее показала, что долгосрочное потребление сахарозы в течение недели (12) облегчает дифференциальный фармакологический ответ на фармакотерапевтические средства, которые, как было показано, модулируют ответы допамина и ацетилхолина на уровне NAc (Шариф и др., В прессе). В совокупности это говорит о том, что долгосрочное (12 недельное и последующее) воздействие сахарозы, которое более точно отражает реальные сценарии, приводит к адаптации морфологии на уровне NAc.
С точки зрения злоупотребления наркотиками повторное воздействие различных препаратов вызывает длительные изменения структуры дендритов и дендритных шипов. Например, амфетамины и кокаин увеличивают плотность позвоночника в NAc как в оболочке, так и в сердечнике (Робинсон и Колб, 2004). Показано, что воздействие никотина также увеличивает плотность позвоночника в оболочке NAc. И наоборот, воздействие морфина приводит к уменьшению плотности позвоночника и сложности дендритной отрасли (Робинсон и Колб, 2004). Что касается долгосрочного потребления сахарозы, мы наблюдали увеличение плотности позвоночника, аналогичное амфетамину, кокаину и никотину и противоположным эффекту морфина. Однако, в отличие от амфетамина и кокаина, но похожего на никотин, увеличение плотности позвоночника при длительном воздействии сахарозы ограничивается оболочкой NAc. Интересно также, что изменения как дендритного ветвления (Робинсон и Колб, 1999) и плотности позвоночника (Li et al., 2003), произведенные амфетамином или кокаином, ограничены дистальными дендритами MSN в NAc, что отражает результаты нашего исследования, Кроме того, и подтверждающие изменения, описанные выше, потребление сахарозы также ранее было показано, чтобы усилить синергетическую силу возбуждения на аккремальные дофаминовые нейроны (Stuber et al., 2008b), а также другие компоненты пути мезолимбической награды (Stuber et al., 2008a; Chen et al., 2010). В совокупности это означает сахарозу в качестве мощного модулятора нейроновой морфологии после длительного интенсивного использования, что сродни эффектам, наблюдаемым от наркотических средств.
Хотя для выявления клеточных и синаптических механизмов, способствующих морфологическим изменениям, наблюдаемым в этом исследовании, необходимы дальнейшие исследования, наши результаты демонстрируют значительные эффекты нейронов, вызванные долгосрочным потреблением сахарозы. В частности, рассмотрение, не рассмотренное в нашем исследовании, заключается в том, могут ли наблюдаемые морфологические эффекты сахарозы быть вызваны некалорийными подсластителями, такими как сахарин. В этой связи важно отметить, что Ленуар и его коллеги показали, что интенсивная сладость превосходит награду кокаина, будь то сахарин или сахароза (Lenoir и др., 2007). Кроме того, недавнее исследование, опубликованное нашей лабораторией (Шариф и др., В прессе) демонстрирует, что варениклин, неполный агонист рецептора никотинового ацетилхолина, уменьшает потребление сахарозы и сахарина у грызунов в соответствии с тем же режимом длительного прерывистого доступа, который использовался в настоящем исследовании. Интересно, что в предыдущих исследованиях было показано сходство в острых эффектах некалорийных подсластителей, таких как сахарин и сахароза, на уровне NAc (Scheggi et al., 2013; Tukey и др., 2013; Карелли и Запад, 2014). Тем не менее, дальнейшим исследованиям необходимо определить, могут ли некалорийные подсластители вызывать долгосрочные эффекты, подобные изменениям морфологии НС оболочки оболочки, вызванному долгосрочным потреблением сахарозы, описанным здесь.
Отсутствие влияния на морфологию NAc MSN после кратковременного потребления сахарозы подчеркивает важность проведения долгосрочных исследований для оценки воздействия длительного злоупотребления наркотиками или природных наград, таких как сахароза. В терминах зависимости не только повторяющиеся циклы употребления алкоголя и абстиненции ключевых компонентов цикла зависимости, все больше доказательств показывает, что переход к зависимости является прогрессивным процессом, который часто происходит в течение длительного периода времени. Хотя аддиктивные свойства сахара остаются неопределенными, все больше расследуется правдоподобие зависимости от других нелекарственных вознаграждений, таких как секс, азартные игры и еда. Результаты этого исследования добавляют достоинства к гипотезе о том, что сахара, такие как сахароза, потенциально обладают аддиктивными свойствами после длительного потребления, напоминающего выпивку. Наши результаты также имеют последствия для растущего числа детей и подростков, которые поддерживают нездоровые привычки в еде (высокий уровень потребления сахара и выпивка) во взрослую жизнь. В соответствии с повышенным риском развития метаболических эффектов также возможно, что неврологические и психиатрические последствия, влияющие на настроение и мотивацию, также могут быть результатом этого поведения.
Авторские вклады
Участвовал в исследовании: PK, SB. Проведенные эксперименты: PK, MS, AB, MF, EM. Анализ данных: ПК, МФ, МС. Интерпретировал данные и способствовал написанию рукописи: ПК, MS, MF, EM, MB, SB. Все авторы прочитали и утвердили окончательную рукопись для подачи.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Рецензенты SC, SA и редактор обработки заявили о своей совместной принадлежности, а редактор обработки заявляет, что этот процесс, тем не менее, соответствовал стандартам справедливого и объективного обзора.
Благодарности
Эта работа была поддержана финансированием из грантов Австралийского исследовательского совета (FT1110884) для SB и Национального совета по здравоохранению и медицинским исследованиям (1061979) для SB и MB.
Дополнительный материал
Дополнительный материал для этой статьи можно найти в Интернете по адресу: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
Дополнительный рисунок 1. Потребление сахарозы и предпочтение от 4 и 12 недельных сахарозы, потребляющих крыс. (А, В) показывают эскалацию общего потребления сахарозы (мл) в течение 4 и 12 недель воздействия. (CD) показывают высокое предпочтение сахарозы над водой в периоды представления сахарозы.
Рекомендации
Ahmed, SH и Koob, GF (1998). Переход от умеренного к чрезмерному употреблению наркотиков: изменение гедонистической уставки. Наука 282, 298-300. doi: 10.1126 / science.282.5387.298
Ahmed, SH, Lin, D., Koob, GF и Parsons, LH (2003). Эскалация самообеспечения кокаина не зависит от изменения уровней допамина, вызванного кокаином. J. Neurochem. 86, 102-113. doi: 10.1046 / j.1471-4159.2003.01833.x
Ahmed, SH, Walker, JR и Koob, GF (2000). Постоянное увеличение мотивации принимать героин у крыс с историей эскалации наркотиков. Нейропсихофармакологии 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
Арсланян С. (2002). Тип диабета 2 у детей: клинические аспекты и факторы риска. Horm Res 57 (комплект 1), 19-28. doi: 10.1159 / 000053308
Avena, NM, Bocarsly, ME, Hoebel, BG и Gold, MS (2011). Перекрытия в нозологии злоупотребления психоактивными веществами и переедания: трансляционные последствия «пищевой зависимости». Тек. Преступность по наркотикам 4, 133-139. doi: 10.2174 / 1874473711104030133
Avena, NM, Rada, P. и Hoebel, BG (2008). Доказательства сахарной зависимости: поведенческие и нейрохимические эффекты прерывистого, чрезмерного потребления сахара. Neurosci. Biobehav. Rev. 32, 20-39. doi: 10.1016 / j.neubiorev.2007.04.019
Avena, NM, Rada, P., Moise, N. и Hoebel, BG (2006). Подгузник сахарозы, питающийся расписанием выпивки, регулярно высвобождает дофамин и устраняет реакцию насыщения ацетилхолина. неврология 139, 813-820. doi: 10.1016 / j.neuroscience.2005.12.037
Бентон, Д. (2010). Правдоподобие сахарной зависимости и ее роль в ожирении и нарушениях питания. Clin. Nutr. 29, 288-303. doi: 10.1016 / j.clnu.2009.12.001
Bray, GA и Popkin, BM (2014). Диетический сахар и масса тела: мы достигли кризиса в эпидемии ожирения и диабета ?: здоровье быть проклятым! Налейте сахар. Лечение диабета 37, 950-956. doi: 10.2337 / dc13-2085
Brog, JS, Salyapongse, A., Deutch, AY и Zahm, DS (1993). Образцы афферентной иннервации ядра и оболочки в «прилежащей» части вентральной полосатой крысы: иммуногистохимическое обнаружение ретроградно-переносимого фторо-золота. J. Comp. Neurol. 338, 255-278. doi: 10.1002 / cne.903380209
Bucher Della Torre, S., Keller, A., Laure Depeyre, J. и Kruseman, M. (2015). Подслащенные сахаром напитки и риск ожирения у детей и подростков: систематический анализ того, как методологическое качество может влиять на выводы. J. Acad. Nutr. Рацион питания. [Epub перед печатью]. doi: 10.1016 / j.jand.2015.05.020
Carelli, RM и West, EA (2014). Когда хороший вкус становится плохим: нервные механизмы, лежащие в основе возникновения негативного аффекта и связанной с ним естественной девальвации вознаграждения кокаином. Нейрофармакология 76 (Pt B), 360-369. doi: 10.1016 / j.neuropharm.2013.04.025
Chen, BT, Hopf, FW и Bonci, A. (2010). Синаптическая пластичность в мезолимбической системе: терапевтические последствия для злоупотребления психоактивными веществами. Энн. NY Acad. Sci. 1187, 129-139. doi: 10.1111 / j.1749-6632.2009.05154.x
Colantuoni, C., Rada, P., McCarthy, J., Patten, C., Avena, NM, Chadeayne, A., et al. (2002). Доказательства того, что прерывистое, чрезмерное потребление сахара вызывает эндогенную опиоидную зависимость. Obes. Местожительство 10, 478-488. doi: 10.1038 / oby.2002.66
Colantuoni, C., Schwenker, J., McCarthy, J., Rada, P., Ladenheim, B., Cadet, JL, et al. (2001). Чрезмерное потребление сахара меняет связывание с дофамином и му-опиоидными рецепторами в головном мозге. Neuroreport 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
Crombag, HS, Gorny, G., Li, Y., Kolb, B. и Robinson, TE (2005). Противоположные эффекты опыта самоконтроля амфетамина на дендритных шипах в медиальной и орбитальной префронтальной коре. Cereb. кора головного мозга 15, 341-348. doi: 10.1093 / cercor / bhh136
Dabelea, D., Mayer-Davis, EJ, Saydah, S., Imperatore, G., Linder, B., Divers, J., et al. (2014). Распространенность типа 1 и диабет типа 2 среди детей и подростков с 2001 до 2009. JAMA 311, 1778-1786. doi: 10.1001 / jama.2014.3201
Davis, C. (2015). Эпидемиология и генетика расстройства пищевого поведения (BED). CNS Spectr. 20, 522-529. doi: 10.1017 / s1092852915000462
Ford, ES и Dietz, WH (2013). Тенденции потребления энергии среди взрослых в Соединенных Штатах: результаты NHANES. Am. J. Clin. Nutr. 97, 848-853. doi: 10.3945 / ajcn.112.052662
Фрайар, CD, Кэрролл, MD и Огден, CL (2014). Распространенность избыточного веса, ожирения и экстремального ожирения среди взрослых: США, 1960-1962 через 2011-2012, Атланта, Джорджия: CDC.
Graveland, GA и DiFiglia, M. (1985). Частота и распределение нейронов среднего размера с зазубренными ядрами приматов и грызунов neostriatum. Brain Res. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
Groenewegen, HJ, Wright, CI, Beijer, AV и Voorn, P. (1999). Конвергенция и сегрегация вентральных стригальных входов и выходов. Энн. NY Acad. Sci. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
Groves, PM (1980). Синаптические окончания и их постсинаптические мишени в неостриатуме: синаптические специализации, выявленные при анализе серийных разделов. Труды. Natl. Изд-во АН. США 77, 6926-6929. doi: 10.1073 / pnas.77.11.6926
Groves, PM, Linder, JC и Young, SJ (1994). 5-гидроксидопамин-меченые допаминергические аксоны: трехмерные реконструкции аксонов, синапсов и постсинаптических мишеней у крыс neostriatum. неврология 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
Hu, FB (2013). Решено: имеется достаточное количество научных доказательств того, что снижение потребления сахара, обогащенного сахаром, уменьшит распространенность ожирения и связанных с ожирением заболеваний. Obes. Rev. 14, 606-619. doi: 10.1111 / obr.12040
Johnson, LR, Aylward, RL, Hussain, Z. и Totterdell, S. (1994). Вход от миндалины к ядру крысы accumbens: ее связь с тирозин гидроксилазой иммунореактивность и идентифицированные нейроны. неврология 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
Кайя, Х. и Намба, М. (1981). Два типа дофаминергических нервных терминалов в крысах neostriatum. Ультраструктурное исследование. Neurosci. Lett. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
Kalivas, PW и Miller, JS (1984). Нейротензивные нейроны в брюшной тегментальной области проектируются к медиальному ядру accumbens. Brain Res. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
Kawaguchi, Y., Wilson, CJ и Emson, PC (1990). Подтипы проецирования клеток неоспиралиновой клетки крысы выявлены путем внутриклеточной инъекции биоцитина. J. Neurosci. 10, 3421-3438.
Kelley, AE и Swanson, CJ (1997). Кормление, вызванное блокадой AMPA и каинатных рецепторов в брюшном полосатом теле: исследование микроинфузионного картирования. Behav. Brain Res. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
Kemp, JM и Powell, TP (1971). Синаптическая организация хвостатодного ядра. Philos. Сделка R. Soc. Лонд. B Biol. Sci. 262, 403-412. doi: 10.1098 / rstb.1971.0103
Кенни, PJ (2011). Механизмы вознаграждения при ожирении: новые идеи и будущие направления. Нейрон 69, 664-679. doi: 10.1016 / j.neuron.2011.02.016
Kessler, RC, Berglund, PA, Chiu, WT, Deitz, AC, Hudson, JI, Shahly, V., et al. (2013). Распространенность и корреляция расстройства пищевого поведения во Всемирных исследованиях психического здоровья Всемирной организации здравоохранения. Biol. психиатрия 73, 904-914. doi: 10.1016 / j.biopsych.2012.11.020
Klenowski, PM, Fogarty, MJ, Belmer, A., Noakes, PG, Bellingham, MC и Bartlett, SE (2015). Структурная и функциональная характеристика дендритных беседок и ГАМКергических синаптических входов на интернейронах и основных клетках в базалатеральной миндалине крысы. J. Neurophysiol. 114, 942-957. doi: 10.1152 / jn.00824.2014
Lenoir, M., Serre, F., Cantin, L. и Ahmed, SH (2007). Интенсивная сладость превосходит награду кокаина. PLoS ONE 2: e698. doi: 10.1371 / journal.pone.0000698
Li, Y., Kolb, B. и Robinson, TE (2003). Расположение постоянных изменений, вызванных амфетамином, в плотности дендритных шипов на средних колючих нейронах в ядре accumbens и caudate-putamen. Нейропсихофармакологии 28, 1082-1085. doi: 10.1038 / sj.npp.1300115
Lindvall, O. и Björklund, A. (1978). Анатомия дофаминергических нейронных систем в мозге крысы. Adv. Biochem. Psychopharmacol. 19, 1-23.
Люттер, М. и Нестлер, EJ (2009). Гомеостатические и гедонические сигналы взаимодействуют в регуляции приема пищи. J. Nutr. 139, 629-632. doi: 10.3945 / jn.108.097618
Maldonado-Irizarry, CS, Swanson, CJ и Kelley, AE (1995). Глютаматные рецепторы в ядре accumbens оболочки контролируют поведение кормления через латеральный гипоталамус. J. Neurosci. 15, 6779-6788.
Malik, VS, Popkin, BM, Bray, GA, Després, JP и Hu, FB (2010). Подслащенные сахаром напитки, ожирение, сахарный диабет типа 2 и риск сердечно-сосудистых заболеваний. Обращение 121, 1356-1364. doi: 10.1161 / CIRCULATIONAHA.109.876185
Ng, SW, Slining, MM и Popkin, BM (2012). Использование калорийных и некалорических подсластителей в пищевых продуктах, упакованных в США, 2005-2009. J. Acad. Nutr. Рацион питания. 112, 1828-1834 e1821-e1826. doi: 10.1016 / j.jand.2012.07.009
Nielsen, SJ, Siega-Riz, AM и Popkin, BM (2002). Тенденции потребления энергии в США между 1977 и 1996: аналогичные сдвиги наблюдаются по возрастным группам. Obes. Местожительство 10, 370-378. doi: 10.1038 / oby.2002.51
Paxinos, G., Watson, C. (2007). Мозг крысы в стереотаксических координатах, Амстердам; Boston, MA: Academic Press / Elsevier.
Попкин, Б.М. (2010). Что не так с американским подходом к ожирению? Виртуальный наставник 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
Рада, П., Авена, Н. М. и Хебель, Б. Г. (2005). Ежедневный bingeing по сахару неоднократно высвобождает допамин в оболочке accumbens. неврология 134, 737-744. doi: 10.1016 / j.neuroscience.2005.04.043
Rafols, JA, Cheng, HW и McNeill, TH (1989). Исследование Гольджи полосатого тела: возрастные дендритные изменения в разных популяциях нейронов. J. Comp. Neurol. 279, 212-227. doi: 10.1002 / cne.902790205
Ranjan, A. и Mallick, BN (2010). Модифицированный метод для последовательного и надежного окрашивания Golgi-cox значительно сокращает время. Фронт. Neurol. 1: 157. doi: 10.3389 / fneur.2010.00157
Рейнехр, Т. (2013). Тип 2 сахарный диабет у детей и подростков. Мир Дж. Диабет 4, 270-281. doi: 10.4239 / wjd.v4.i6.270
Reissner, KJ и Kalivas, PW (2010). Использование глютамата гомеостаза как мишень для лечения зависимых расстройств. Behav. Pharmacol. 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
Reynolds, SM и Berridge, KC (2003). Мотивационные ансамбли глутамата в прилежащих ядрах: рострокаудальные градиенты оболочки страха и питания. Евро. J. Neurosci. 17, 2187-2200. doi: 10.1046 / j.1460-9568.2003.02642.x
Richard, JM и Berridge, KC (2011). Nucleus accumbens взаимодействует с дофамином / глутаматом, чтобы генерировать желание против страха: только D (1) для аппетитного питания, но D (1) и D (2) вместе для страха. J. Neurosci. 31, 12866-12879. doi: 10.1523 / JNEUROSCI.1339-11.2011
Robinson, TE и Kolb, B. (1999). Изменения в морфологии дендритов и дендритных шипов в прилежащем ядре и префронтальной коре после повторного лечения амфетамином или кокаином. Евро. J. Neurosci. 11, 1598-1604. doi: 10.1046 / j.1460-9568.1999.00576.x
Robinson, TE и Kolb, B. (2004). Структурная пластичность, связанная с воздействием наркотиков. Нейрофармакология 47 (комплект 1), 33-46. doi: 10.1016 / j.neuropharm.2004.06.025
Russo, SJ, Dietz, DM, Dumitriu, D., Morrison, JH, Malenka, RC и Nestler, EJ (2010). Зависимый синапс: механизмы синаптической и структурной пластичности в прилежащих ядрах. Тенденции Neurosci. 33, 267-276. doi: 10.1016 / j.tins.2010.02.002
Rutledge, LT, Duncan, J. и Beatty, N. (1969). Изучение коллатералей аксонов пирамидальных клеток в интактной и частично изолированной коре головного мозга. Brain Res. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper, CB, Chou, TC и Elmquist, JK (2002). Потребность в корме: гомеостатический и гедонический контроль за едой. Нейрон 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
Scheggi, S., Secci, ME, Marchese, G., De Montis, MG и Gambarana, C. (2013). Влияние вкусовых качеств на стимуляцию для работы на калорийную и некалорийную пищу у непитанных и без пищи лишенных крыс. неврология 236, 320-331. doi: 10.1016 / j.neuroscience.2013.01.027
Sesack, SR и Pickel, VM (1990). В крысах медиального ядра accumbens, гиппокампальные и катехоламинергические терминалы сходятся на колючих нейронах и находятся в приложении друг к другу. Brain Res. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
Sesack, SR и Pickel, VM (1992). Префронтальные кортикальные эфференты в синапсе крыс на немеченых нейронных мишенях терминалов катехоламинов в сепсисах ядра и на дофаминовых нейронах в брюшной тегментальной области. J. Comp. Neurol. 320, 145-160. doi: 10.1002 / cne.903200202
Shariff, M., Quik, M., Holgate, JY, Morgan, M., Patkar, OL, Tam, V., et al. (в прессе). Нейронные никотиновые ацетилхолиновые рецепторные модуляторы снижают потребление сахара. PLoS ONE.
Шихан, Д.В. и Герман, Б.К. (2015). Психологические и медицинские факторы, связанные с необработанным расстройством пищевого поведения. Прим. Care Companion CNS Disord. 17. doi: 10.4088 / PCC.14r01732
Simms, JA, Steensland, P., Medina, B., Abernathy, KE, Chandler, LJ, Wise, R., et al. (2008). Периодический доступ к этанолу 20% вызывает высокое потребление этанола у крыс Long-Evans и Wistar. Алкоголь. Clin. Exp. Местожительство 32, 1816-1823. doi: 10.1111 / j.1530-0277.2008.00753.x
Steensland, P., Simms, JA, Holgate, J., Richards, JK и Bartlett, SE (2007). Varenicline, α4beta2 никотиновый ацетилхолиновый рецептор агониста, выборочно уменьшает потребление этанола и поиск. Труды. Natl. Изд-во АН. США 104, 12518-12523. doi: 10.1073 / pnas.0705368104
Stuber, GD, Hopf, FW, Hahn, J., Cho, SL, Guillory, A. и Bonci, A. (2008a). Добровольное потребление этанола усиливает синтаптическую силу возбуждения в брюшной тегментальной области. Алкоголь. Clin. Exp. Местожительство 32, 1714-1720. doi: 10.1111 / j.1530-0277.2008.00749.x
Stuber, GD, Klanker, M., de Ridder, B., Bowers, MS, Joosten, RN, Feenstra, MG, et al. (2008b). Вознаграждение - прогностические сигналы усиливают синтаптическую силу возбуждения на нейронах дофамина среднего мозга. Наука 321, 1690-1692. doi: 10.1126 / science.1160873
Swanson, SA, Crow, SJ, Le Grange, D., Swendsen, J. и Merikangas, KR (2011). Распространенность и корреляция нарушений питания у подростков. Результаты опроса, посвященного репродукции подростковой популяции. Архипелаг Ген. Психиатрия 68, 714-723. doi: 10.1001 / archgenpsychiatry.2011.22
Te Morenga, L., Mallard, S. и Mann, J. (2013). Диетические сахара и масса тела: систематический обзор и метаанализ рандомизированных контролируемых исследований и когортных исследований. BMJ 346: e7492. doi: 10.1136 / bmj.e7492
Totterdell, S. и Smith, AD (1989). Сходимость гиппокампа и дофаминергического входа на идентифицированные нейроны в ядре прикрытия крысы. J. Chem. Neuroanat. 2, 285-298.
Тьюки, Д.С., Феррейра, Дж. М., Антуан, С. О., Д'Амур, Дж. А., Нинан, И., Кабеса де Вака, С. и др. (2013). Прием сахарозы вызывает быстрое перемещение рецепторов AMPA. J. Neurosci. 33, 6123-6132. doi: 10.1523 / JNEUROSCI.4806-12.2013
Veening, JG, Cornelissen, FM и Lieven, PA (1980). Актуальная организация афферентов к caudatoputamen крысы. Исследование пероксидазы хрена. неврология 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
Вентура, Т., Сантандер, Дж., Торрес, Р. и Контрерас, А. М. (2014). Нейробиологическая основа тяги к углеводам. Питание 30, 252-256. doi: 10.1016 / j.nut.2013.06.010
Volkow, ND, Wang, GJ, Fowler, JS, Tomasi, D. и Baler, R. (2012). Продовольственная и лекарственная награда: перекрывающиеся цепи в человеческом ожирении и наркомании. Тек. Верхний. Behav. Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
Мудрый, РА (1973). Добровольное потребление этанола у крыс после контакта с этанолом в разных графиках. Psychopharmacologia 29, 203-210. doi: 10.1007 / BF00414034
Райт, CI и Groenewegen, HJ (1995). Образцы сходимости и сегрегации в срединном ядре прикрытия крысы: отношения префронтальной коры головного мозга, средней линии талама и базальные амигдалоидные афференты. J. Comp. Neurol. 361, 383-403. doi: 10.1002 / cne.903610304
Ключевые слова: потребление, напоминающее выпивку, долгосрочный, средний колючий нейрон, прилежащие ядра, сахароза
Образец цитирования: Klenowski PM, Shariff MR, Belmer A, Fogarty MJ, Mu EWH, Bellingham MC и Bartlett SE (2016) Длительное потребление сахарозы в способе, напоминающем выпивку, изменяет морфологию средних колючих нейронов в оболочке Nucleus Accumbens. Фронт. Behav. Neurosci, 10: 54. doi: 10.3389 / fnbeh.2016.00054
Поступило в редакцию: 03 December 2015; Принято: 07 March 2016;
Опубликовано: 23 March 2016.
Под редакцией:
Djoher Nora Abrous, Институт неврологии де Бордо, Франция
Рассмотрено:
Серж Х. Ахмед, Центр национального развития ЮНЕСКО, Франция
Стефани Кайль, Центр национального развития ЮНЕСКО, Франция
Copyright © 2016 Кленовски, Шариф, Белмер, Фогарти, Му, Беллингем и Бартлетт. Это статья с открытым доступом, распространяемая в соответствии с условиями Лицензия Creative Commons Attribution (CC BY), Использование, распространение или воспроизведение на других форумах разрешено при условии, что оригинальный автор (ы) или лицензиар зачисляются и что оригинальная публикация в этом журнале цитируется в соответствии с принятой академической практикой. Запрещается использование, распространение или воспроизведение, которое не соответствует этим условиям.
* Переписка: Селена Е. Бартлетт, [электронная почта защищена]
