 Мартин Зак
Мартин Зак- 1Лаборатория когнитивной психофармакологии, Отделение нейробиологии, Центр наркомании и психического здоровья, Торонто, О.Н., Канада
- 2Программа трансляционной неврологии, отделение психиатрии, Школа медицины, Пенсильванский университет, Филадельфия, Пенсильвания, США
- 3Секция биопсихологии, отделение нейронауки, Центр наркомании и психического здоровья, Торонто, О.Н., Канада
Зависимость считается заболеванием мозга, вызванным хроническим воздействием наркотиков. Сенсибилизация систем дофамина мозга (DA) частично опосредует этот эффект. Патологическая азартная игра (PG) считается поведенческой зависимостью. Поэтому PG может быть вызвано хроническим воздействием азартных игр. Идентификация асинхронной сенсибилизации систем DA поддержала бы эту возможность. Азартные игры вызывают выпуск DA. Один эпизод игры в игровые автоматы сдвигает ответ DA от доставки вознаграждения до появления реплик (вращающихся барабанов) за вознаграждение в соответствии с принципами обучения во времени. Таким образом, условные раздражители (CS) играют ключевую роль в ответах DA на азартные игры. У приматов ответ DA на CS наиболее силен, когда вероятность вознаграждения равна 50%. В соответствии с этим графиком CS выдает ожидаемое вознаграждение, но не предоставляет никакой информации о том, будет ли оно происходить в ходе данного испытания. Во время азартных игр график 50% должен вызывать максимальный выпуск DA. Это близко соответствует частоте вознаграждения (46%) на коммерческом игровом автомате. DA может способствовать повышению чувствительности, особенно для амфетамина. Хроническое воздействие КС, которое прогнозирует вознаграждение 50% времени, может имитировать этот эффект. Мы протестировали эту гипотезу в трех исследованиях с крысами. Животные получили 15 × 45-min воздействия CS, которые предсказали вознаграждение с вероятностью 0, 25, 50, 75 или 100%. КС был светом; награда была решением 10% сахарозы. После тренировки крысы получали сенсибилизирующий режим пяти отдельных доз (1 мг / кг) d-амфетамина. Наконец, они получили 0.5 или 1 мг / кг амфетамина вызов до 90-мин локомоторный тест активности. Во всех трех исследованиях группа 50% проявляла большую активность, чем другие группы, в ответ на обе дозу дозы. Размеры эффектов были скромными, но непротиворечивыми, что отразилось на значительной группе × ранг-ассоциации (φ = 0.986, p = 0.025). Хроническое воздействие азартных игр графика поощрительных стимулов вознаграждения может способствовать сенсибилизации к амфетамину, как к самому воздействию амфетамина.
Введение
Наркомания была охарактеризована как заболевание мозга, вызванное хроническим воздействием наркотических средств (Leshner, 1997). Предполагается, что нейропластичность опосредует эффекты такого воздействия (Nestler, 2001). Сенсибилизация систем дофамина головного мозга (DA) представляет собой форму нейропластичности, связанной с гиперреактивностью к условным раздражителям (CS) для лекарств и компульсивному поиску наркотиков (Робинсон и Берридж, 2001). Аллергены были оперативно определены увеличением выпуска DA в ответ на КС за вознаграждение и увеличение реакции опорно-двигательного аппарата к фармакологической DA вызову (Робинсон и Берридж, 1993; Пирс и Каливас, 1997; Вандершурен и Каливас, 2000). Хотя сенсибилизация является лишь одним из многих изменений мозга, связанных с наркоманией (ср. Роббинс и Эверитт, 1999; Koob и Le Moal, 2008), были предложены изменения в пресинаптическом высвобождении дофамина, которые представляют собой общие нейроадаптации, связанные с наркоманией, основанной на зависимости (например, рецидив), в тех препаратах, которые индуцируют локомоторную сенсибилизацию к опиатам (например, морфину) или стимулятору (например, амфетамину) , также вызывают восстановление погашенных ответов операндов на самообслуживание героина или кокаина - модель рецидива животных (Vanderschuren et al., 1999). Доказательства того, что стимулирующая сенсибилизация (повышенная ценность лекарственного вознаграждения) наиболее выражена после того, как первоначальное воздействие на наркотики вызывает дополнительные подозрения, предполагает, что сенсибилизация может также участвовать в ранних стадиях зависимости (Вандершурен и Пирс, 2010).
Патологическая азартная игра (PG) была описана как поведенческая зависимость и недавно реклассифицирована в ту же категорию, что и нарушения зависимости от психоактивных веществ в 5-м издании Диагностического и статистического руководства по психическим расстройствам (Frascella et al., 2010; APA, 2013). Это подразумевает, что PG может быть вызвано хроническим воздействием азартной игры, подобно тому, что общие механизмы могут опосредовать последствия азартных игр и воздействия наркотиков (Зак и Пулос, 2009; Лиман и Потенца, 2012); и что сенсибилизация мозговых путей DA может быть одним из важных элементов этого процесса.
Клинические данные косвенно подтверждают эту возможность: используя позитронно-эмиссионную томографию (ПЭТ), Бойлау и его коллеги обнаружили, что у мужских пациентов с ПГ значительно большее сокращение стриала ДА в ответ на амфетамин (0.4 мг / кг), чем у здоровых мужских контролей (Boileau et al., 2013). Общие групповые различия были значительными в ассоциативной и соматосенсорной полосатой. В лимбическом стриатуме, который включает ядро accumbens, группы не различались. Однако у субъектов PG высвобождение DA в лимбической стриатеме напрямую коррелировало с выраженностью симптомов PG. Эти данные согласуются с сенсибилизацией путей DA мозга в PG, но также указывают на некоторые важные различия с людьми, зависящими от человека, и с классической моделью сенсибилизации амфетамина на животных. В отличие от субъектов PG и животных, подвергнутых воздействию низких доз амфетамина (см. Робинсон и др., 1982), люди с зависимостью от вещества последовательно демонстрируют снижение выделения ДА до стимуляции (например,Volkow и др., 1997; Martinez и др., 2007), и данные, полученные от животных, свидетельствуют о том, что это может отражать дефицит функции DA во время начальных стадий абстиненции после укусов моделей злоупотребления психоактивными веществами (Mateo et al., 2005). В исследованиях, где сенсибилизация стимулятора показана у животных, усиленное высвобождение DA обычно наблюдается в лимбическом стриатуме, а не дорсальном (ассоциативном, соматосенсорном) полосатом (Везина, 2004). Однако индуцированный кией (т. Е. Обусловленный) поиск наркотиков у животных, неоднократно подвергавшихся воздействию кокаина, был связан с усиленным высвобождением DA в дорсальном полосатом теле, что, по-видимому, указывало на более привычную форму мотивированного поведения (Ito et al., 2002). Таким образом, общее повышение уровня DA в дорзальных областях у субъектов PG может быть связано с поиском на основе привычки (негибким, рутинным) вознаграждением, включающим «прогрессирование от брюшной до более дорзальных областей полосатого тела» (Эверитт и Роббинс, 2005, п. 1481), тогда как зависимое от тяжести высвобождение DA в лимбической полосатой у этих субъектов может более тесно соответствовать стимулирующей сенсибилизации, как правило, моделируется у животных. Результаты ПЭТ не могут выявить, была ли гиперреактивность DA ранее существовавшей особенностью этих субъектов ПГ, последствием воздействия азартных игр или результатом какого-либо другого процесса полностью. Чтобы решить этот вопрос, необходимо продемонстрировать индукцию сенсибилизации путем хронической азартной игры у субъектов, которые являются нормальными до воздействия. Это вызывает вопросы о том, какие особенности азартных игр, скорее всего, вызовут сенсибилизацию.
Скиннер отметил, что переменный график подкрепления был фундаментальным для привлекательности азартных игр (или, по крайней мере, ее устойчивости) (Скиннер, 1953). Поведение в играх с игровыми автоматами хорошо согласуется с основными принципами инструментального кондиционирования, что отражается на предполагаемой корреляции между денежным выигрышем и размером ставки на последовательные спины (Tremblay и др., 2011). Таким образом, ответчик с переменным отношением, по-видимому, обеспечивает внешнюю действительную модель азартных игр в игровых автоматах.
Недавние исследования с животными обеспечивают сильную первоначальную поддержку причинного эффекта воздействия азартных игр на сенсибилизацию. Певица и его коллеги изучили влияние ежедневных сеансов 55 1-h фиксированного (FR20) или переменного (VR20) сахаринного усиления в экспериментальной парадигме рычага-пресса на последующий локомоторный ответ на амфетамин с низкой дозой (0.5 мг / кг) у здорового мужчины ( Sprague Dawley) крысы (Сингер и др., 2012). Они предположили, что если азартные игры приводят к сенсибилизации, крысы, подвергнутые переменному графику, которые имитируют азартные игры, должны проявлять больший отклик на амфетамин, чем крысы, подвергнутые фиксированному графику. Как и было предсказано, группа VR20 показала 50% больший опорный отклик на амфетамин, чем группа FR20. Напротив, группы проявили эквивалентную локомоцию после инъекции солевого раствора. Эти данные подтверждают, что хроническое воздействие переменного подкрепления является достаточным для того, чтобы вызвать гиперреактивность к DA-инфекции у здоровых животных, рандомизированных к соответствующим графикам.
Из этого результата возникает ряд вопросов: во-первых, в какой степени воспринимаемая непредвиденная ситуация - или ее отсутствие - между ответом операнта и его результатом опосредует эти эффекты? В учебных терминах этот эффект включает в себя «ожидаемое время отклика» или может быть аналогичный эффект наблюдаться при отсутствии ответа операнта, т. Е. «Ожидаемого результата стимула» в павловской парадигме (ср. Bolles, 1972)? Во-вторых, влияет ли степень непредвиденности между предшествующим событием (ответом или стимулом) и его результатом на степень сенсибилизации?
Второй вопрос касается роли неопределенности в сенсибилизации. Например, игры, результат которых действительно случайный, совершенно непредсказуемый, имеют больший потенциал для стимулирования сенсибилизации, чем игры, где шансы на выигрыш четко определены, но не случайны, даже если абсолютная норма вознаграждения низкая? Настоящее исследование рассматривало эти вопросы.
Экспериментальный проект был проинформирован семантическим исследованием о ожидаемой продолжительности вознаграждения и реакции нейронной ДНК у обезьян (Fiorillo и др., 2003). Животные в этом исследовании получили вознаграждение за сок (США) в соответствии с графиками 0, 25, 50, 75 или 100%. Графики были обозначены 1 4 разных CS (значков). График 0% предоставлял вознаграждение так же часто, как график 100%, но опустил CS. Частота стрельбы нейронов DA в интервале между началом CS и доставкой или бездействием в США была ключевой зависимой мерой. Исследование показало, что ответ ДА увеличился в зависимости от неопределенности доставки вознаграждения. Таким образом, в соответствии с графиком 100% CS вызвала небольшую активность в соответствии с графиками 25 и 75%, CS вызвала умеренный и аналогичный уровень активности, а в графике 50% CS вызвала максимальную активность. В каждом случае скорость стрельбы возрастала в течение интервала CS-US, т. Е. По мере приближения ожидаемого результата.
Эти данные показывают, что активность DA зависит не только от того, является ли вознаграждение определенным (фиксированное соотношение), либо неопределенным (переменное отношение), но также изменяется в обратной пропорции к количеству информации о доставке вознаграждения, передаваемой CS. В состоянии 100% CS вызывает ожидаемое вознаграждение, а также прекрасно прогнозирует его доставку. В условиях 25 и 75% CS вызывает ожидание и предсказывает выплату вознаграждения три раза в четыре раза. В состоянии 50% CS вызывает ожидание, но не предоставляет никакой информации о доставке вознаграждения, кроме случая, когда-либо. Основываясь на своих выводах, Fiorillo et al. заключил: «Это увеличение дозы дофамина, вызванное неопределенностью, может способствовать повышению качества азартных игр» (стр. 1901).
Эффект от 50% переменного вознаграждения в одном сеансе не должен меняться в течение нескольких сеансов, потому что вероятность вознаграждения остается совершенно непредсказуемой в каждом испытании. Таким образом, при рассмотрении условий, которые могут максимизировать хроническую активацию нейронов DA в повторяющихся эпизодах азартных игр, 50% -ный график должен вызвать как наиболее стойкий, так и наиболее устойчивый эффект. Это примечательно, учитывая, что долгосрочная ставка вознаграждения (выигрыш> 0), наблюдаемая за тысячи вращений на коммерческом игровом автомате, составила 45.8% (Tremblay и др., 2011). Таким образом, вознаграждение за переменную 50%, как представляется, точно отражает график выплат, управляемый фактическими игровыми устройствами.
В настоящем исследовании использовались те же схемы кондиционирования, что и в Fiorillo et al. при хроническом воздействии - межгрупповой дизайн с крысами. Животные проходили ~ 3 недели ежедневных сеансов кондиционирования, где CS (свет) сочетался с US (небольшое количество сахарозы). После фазы обучения животные отдыхали до оценки сенсибилизации, индексированной по двигательной реакции на амфетамин. На основании литературы было предсказано, что крысы, подвергавшиеся различным схемам поощрения, не будут различаться в их локомоторном поведении без наркотиков, но будут демонстрировать значительно разные уровни локомоции после амфетамина, при этом группа 50% будет демонстрировать больший локомоторный ответ на лекарство по сравнению с другие группы в течение курса доз, образец, который можно было бы ожидать, если бы 50% животных ранее подвергались дополнительным дозам самого амфетамина (т.е. перекрестная сенсибилизация).
Experiment 1
Материалы и методы
Тематика
Четыре группы (n = 8 / группа) взрослых (300-350 g) самцов крыс Sprague-Dawley (Charles River, St. Constant, Quebec, Canada) размещались индивидуально в прозрачных коробках из поликарбоната (20 × 43 × 22 см) под обратным 12: 12 светло-темный цикл. Они получили вволю доступ к еде и воде, а также ежедневную обработку экспериментатором за 2 за несколько недель до начала исследования. Каждая группа была обусловлена одним из четырех графиков вознаграждений с переменными: 0, 25, 50 или 100%. Группа 75% была опущена в этом начальном исследовании, так как Fiorillo et al. (2003) найденный эквивалентный пост-CS DA релиз в соответствии с графиками вознаграждения 25 и 75%, так что оба условия привели к большему выпуску DA, чем условие 100% CS-US, но меньше, чем условие 50%.
Аппараты
Доступ к презентациям сахарозы и к CS был предоставлен индивидуально в коробках для подготовки операндов (33 × 31 × 29 см). Каждая коробка была оборудована журналом для укрепления, расположенным на передней стене. Свет в верхней части журнала служил CS. Моторизованный, управляемый соленоидом жидкий ковш может быть поднят на пол магазина. События в коробке контролировались оборудованием и программным обеспечением Med Associates, используя внутреннюю программу, написанную на MED-PC. Локомоторные испытания проводились индивидуально в клетках из плексигласа (27 × 48 × 20 см). Каждая клетка была оборудована системой мониторинга, состоящей из шести фотобалевых ячеек для обнаружения горизонтального движения.
Процедура
Обучение, Исследование проводилось в соответствии с этическими принципами, установленными Канадским советом по уходу за животными. Крысы были ограничены продуктом 90% их массы тела в течение всего периода исследования и размещались индивидуально. Каждая крыса получила 15 дней тренировки награды сахарозы (10% водного раствора при 0.06 мл за вознаграждение): 5 последовательных дней × 3 недель, с выходными. Животных поддерживали на стандартной чау до и после этапа обучения; воздействие сахарозы было ограничено 15-15-минутными тренировками. Каждый ежедневный сеанс состоял из презентаций стимулов 40 (свет, CS), каждый из которых разделялся промежуточным интервалом 15 s. Свет был расположен на верхней панели магазина и оставался на 120 с сахарозой, доступной в течение последних 25. В случае группы 5 саксоровый ковш был поднят каждый 0 s (для 140 s), но световой стимул не освещался. Это уравнило интервал между представлениями ковша в группе 5 и другими группами (0 + 120 s). Каждый сеанс лечения длился ~ 25 мин. В среднем группа 40 получала сахарозу один раз на каждые четыре презентации CS; группа 25 получала сахарозу один раз для каждых двух презентаций CS, а группа 50 получала сахарозу после каждой презентации CS.
Тестирование, Через две недели после последнего сеанса доступа сахарозы (или «кондиционирования») оценивали локомоторный ответ на d-амфетамин (AMPH; ip). Крысам давали три сеанса 2-h, чтобы приучить себя к локомоторным ящикам, а затем шесть сеансов AMPH. Дни испытаний AMPH произошли с интервалом 1-wk. В тестовые дни крысам давали 30 мин для приучения к ящикам, после чего получали одну дозу AMPH 0.5 мг / кг, а затем на отдельных еженедельных сеансах пятью дозами 1.0 мг / кг (одна доза в день) в испытательные дни 1 через 5 , Локомоция после AMPH была оценена для 90 min на каждой сессии.
Анализ данных
Статистический анализ проводился с SPSS (v. 16 и v. 21; SPSS Inc., Chicago IL). Непосредственный поведенческий ответ на КС оценивали с точки зрения носового окунания в апертуру, где раздавали сахарозу. Среднее количество носовых пиков в течение этого интервала (5 s за пробу) затем сравнивалось со средним числом носовых пиков на ту же самую длительность (5 s), усредненную за время, когда CS отсутствовала. Группа × Сессия ANOVA из носовых пиков с CS присутствовала и отсутствовала, отслеживая получение дискриминационной реакции на кий и неизбирательный ответ на нос, под разными расписаниями в течение учебных занятий сахарозы 15.
Эффекты лечения локомоторных ответов оценивались с помощью групповых ANOVA для группы без рецепта (три сеанса), предварительной сенсибилизации 0.5 мг / кг AMPH-теста (одна сессия) и во время пятисегментного 1 мг / кг AMPH режим сенсибилизации, когда ожидается, что группы будут отличаться в ответ на повторные дозы AMPH. Группа × Сессия ANOVA также оценивали безрецептурные локомоторные ответы во время фазы привыкания к инъекции 30-min из каждой тестовой сессии AMPH. Планируемые сравнения оценивали разницу в средней производительности для группы 50 по сравнению с группой 0 (без контроля ожидаемой продолжительности) и группы 100 (без контроля неопределенности) с помощью t- тесты (Howell, 1992), используя ошибки MS error и df для соответствующего эффекта (т. е. взаимодействие группы или группы × сеанса) с ANOVA (Winer, 1971). Полиномиальные трендовые анализы проверяли профиль изменений в ходе сеансов.
Для того, чтобы определить, является ли подход реакции в присутствии и в отсутствие КС во время подготовки сахарозы сессий 15 способствовали вариация реакции опорно-двигательного аппарата к AMPH или опосредованных различий группы в ответ AMPH, последующий анализ ковариации (ANCOVAs) были выполнены на AMPH локомоторные данные, включая тотальные носовые накопления (сумма для сеансов 15), когда CS отсутствовала в качестве ковариата. Значительный эффект ковариации будет свидетельствовать о том, что реакция на доступ к лекарственным средствам смягчается (влияет на силу) эффектов группы или сеанса. Снижение значимости эффектов группы или сеанса в присутствии значительного ковариата будет указывать на то, что ответы на подход опосредуют (учитывают) эффекты группы или сеанса. Снижение значения групповых или сеансовых эффектов в отсутствие значительного эффекта ковариации просто отразило бы потерю статистической мощности из-за перераспределения df от члена ошибки до ковариата и не повлияло бы на интерпретацию эффекты группы или сеанса.
Итоги
Носовые токи во время сеансов кондиционирования сахарозы
CS присутствует, фигура 1A показывает средние носовые поры для групп 25, 50 и 100, в то время как CS присутствовал на сеансах кондиционирования сахарозы 15 (носовые пики не были закодированы для группы 0, которая не получила CS). Группа 3 × 15 Session ANOVA дала существенные основные эффекты Group, F(2, 21) = 5.63, p = 0.011 и Session, F(14, 294) = 14.00, p <0.001, наряду со значительным взаимодействием Группа × Сеанс, F(28, 294) = 2.93, p <0.001. Рисунок 1A указывает на то, что основной эффект сеанса отражает увеличение количества носа, которое происходит во всех трех группах, и основной эффект группы отражает в целом более высокие общие оценки в группе 100 по сравнению с группой 25 с промежуточными оценками в группе 50. Значительное взаимодействие группы × Session для кубического тренда, F(2, 21) = 4.42, p = 0.030, указывал на быстрый подъем, опускание и выравнивание в носу, запихивает сеансы в группе 100, в отличие от линейного увеличения по сравнению с сеансами в группе 50 и более мелкого линейного увеличения по сравнению с сеансами в группе 25.
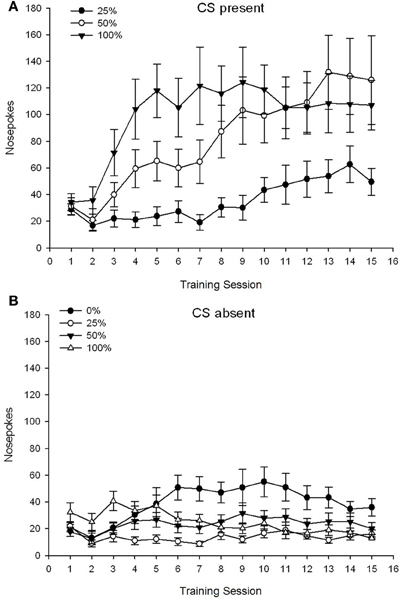
Рисунок 1. Средние (SE) ответы подхода (носовые поры) на 15 сахарозе тренировки в группах крыс Sprague Dawley (n = 8 / группа), подвергнутых вознаграждению за сахарозу (решение 10%), поставляемое в соответствии с графиками 0, 25, 50 или 100%, Условный раздражитель был светом (120 s). Группа 0 получила такое же количество вознаграждений, что и группа 100 в отсутствие условных раздражителей. (A) Оценки при наличии CS (испытания 5 s × 15). (B) Оценки, когда CS отсутствовала (средняя для 5 × 15 s, когда свет был выключен).
CS отсутствует, фигура 1B показывает средние носовые пики для всех четырех групп для эквивалентной продолжительности (5 s × 15), усредненные за время, когда CS отсутствовала. Группа 4 × 15 Session ANOVA дала существенные основные эффекты Group, F(3, 28) = 7.06, p = 0.001 и сеанс F(14, 392) = 2.84, p <0.001, наряду со значительным взаимодействием Группа × Сеанс, F(42, 392) = 3.93, p <0.001. Значимое взаимодействие группы × сеанса для квадратичного тренда, F(3, 28) = 3.91, p = 0.019, наряду с отсутствием взаимодействия для кубического тренда, F(3, 28) <0.93, p > 0.44, отражает «перевернутый-U» профиль сования носа по сеансам в группе 0, в отличие от в целом стабильного профиля по сеансам в других группах.
Пристрастие к локомоторным камерам
Группа 4 × 3 Session ANOVA дала основной эффект сеанса, F(2, 56) = 5.67, p = 0.006, и никаких других значительных эффектов, F(3, 28) <1.60, p > 0.21. Средние (SE) разрывы пучка за 2 часа в локомоторных боксах составили 1681 (123) на сеансе 1, 1525 (140) на сеансе 2 и 1269 (96) на сеансе 3. Планируемые сравнения не обнаружили значительных различий между группой 50 и группой. 0 или группа 100 на первом или последнем сеансе привыкания, t(84) <1.69, p > 0.05. Таким образом, в отсутствие AMPH повторное воздействие тест-боксов было связано с постоянным снижением спонтанной двигательной активности в четырех группах (т. Е. С эффектом сеанса) и отсутствием дифференциального ответа в зависимости от графика тренировок с сахарозой (без взаимодействия). .
Тестирование
Эффекты предварительной сенсибилизации 0.5 мг / кг AMPH.
Предзаводная локомоция, Однофакторный ANOVA группы 4 от локомоторного отклика во время фазы привыкания к инъекции 30-min не давал значительных эффектов, F(3, 28) <1.05, p > 0.38. Запланированные сравнения не обнаружили значительной разницы между группой 50 и группой 0 или группой 100, t(32) <0.87, p > 0.40. Следовательно, исходные различия в локомоции до инъекции не учитывали групповые различия в локомоторном ответе на АМФГ. Среднее значение (SE) разрыва пучка для образца составило 559 (77).
Пост-инъекционная локомоция против заключительной недели без привычек. Сеанс ANOVA из 4 групп × 2 сравнивал локомоторные реакции групп на заключительном сеансе привыкания и сразу после введения предсенсибилизации 0.5 мг / кг AMPH. Баллы для сеанса привыкания (120 минут) были масштабированы, чтобы соответствовать продолжительности тестового сеанса AMPH (90 минут) (исходный показатель привыкания × 90/120). Анализ дал значительный основной эффект сеанса, F(1, 28) = 34.16, p <0.001, и никаких других значимых эффектов, F(3, 28) <2.26, p > 0.10. Эффект сеанса отражал увеличение средних (SE) разрывов пучка в ответ на дозу с 952 (72) до 1859 (151). Запланированные сравнения не обнаружили существенных различий между группой 50 и группой 0 или группой 100 в зависимости от дозы. t(56) <1.72, p > 0.10. Тем не менее, порядок ранжирования баллов разрыва луча (M; SE) соответствует гипотезе: группа 50 (2205; 264)> группа 0 (2025; 203)> группа 100 (1909; 407)> группа 25 (1296; 299). .
Эффекты 1 мг / кг AMPH.
Предзаводная локомоция, Группа 4 × 5-сессия. ANOVA локомоторного ответа во время фазы привыкания 30-min до инъекции на 1 мг / кг сеансов AMPH дал основной эффект сеанса, F(4, 112) = 43.64, p <0.0001, и никаких других значимых эффектов, F(3, 28) <0.97, p > 0.42. Запланированные сравнения не выявили значительных различий между группой 50 и группой 0 или группой 100 на первом или последнем сеансе тестирования. t(140) <0.84, p > 0.30. Следовательно, исходные различия в локомоции не учитывали групповые различия в локомоторном ответе на AMPH. Средние (SE) баллы обрыва луча для фазы привыкания перед приемом на сеансах 1–5 были: 454 (30), 809 (53), 760 (36), 505 (35), 756 (39).
Пост-инъекционная локомоция, фигура 2 показывает влияние пяти инъекций 1 мг / кг AMPH (один раз в неделю) на показатели локомоторной активности в четырех группах. Группа 4 × 5 Session ANOVA дала основной эффект сеанса, F(4, 112) = 8.21, p <0.001, маргинальный основной эффект Группы, F(2, 45) = 3.28, p = 0.085 и никакого значительного взаимодействия, F(12, 122) <0.77, p > 0.68.
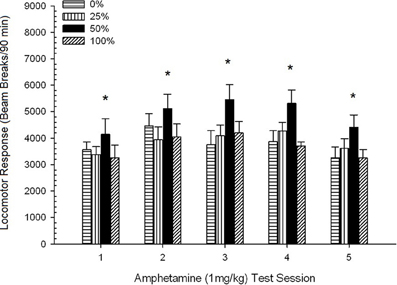
Рисунок 2. Средний (SE) локомоторный отклик (количество разрывов луча в электронной матрице за 90 мин) до 1 мг / кг d-амфетамина (ip) на еженедельных сеансах 5 в группах крыс Sprague Dawley (n = 8 / group), ранее подвергавшихся сеансам суточного кондиционирования 15 с вознаграждением сахарозы (решение 10%), поставляемым в соответствии с графиками 0, 25, 50 или 100%, Условный раздражитель был светом (120 s). Группа 0 получила такое же количество вознаграждений, что и группа 100 в отсутствие условных раздражителей. *p <0.05 для средней разницы между группой 50 и группой 0, а также группой 100 на основе запланированных сравнений.
Запланированные сравнения показали, что оценки группы 50 значительно отличаются от группы 0, t(14) = 2.19, p = 0.037 и группа 100, t(14) = 2.36, p = 0.025 [и незначительно отличается от группы 25, t(14) = 2.03, p = 0.051]. Таким образом, в группе 50 локомоторный отклик на 1 мг / кг AMPH достоверно превышал опорный показатель остальных трех групп во всех пяти сеансах тестирования. Анализ полиномиальных тенденций выявил значительную квадратичную тенденцию в сессиях, F(1, 28) = 32.47, p <0.0001, и никаких других значимых тенденций, F(1, 28) <1.78, p > 0.19. Рисунок 2 показывает, что этот результат отразил шаблон «инвертированный U» в сеансах.
Контроль за вариацией носового мешка в ходе тренировки сахарозы
Последовательный ANCOVA локомоторных ответов на 1 мг / кг AMPH с носовыми парами (CS присутствует) в качестве ковариата в трех группах, получивших CS, дал краевой главный эффект Group, F(2, 20) = 3.07, p = 0.069, и никаких значительных эффектов, связанных с ковариацией, F(4, 80) <0.05, p > 0.85. Таким образом, ответный подход во время тренировки не объясняет значительных изменений локомоторного ответа на 1 мг / кг AMPH в группах 25, 50 или 100.
Последовательный ANCOVA локомоторных ответов на 1 мг / кг AMPH, с носовыми парами (CS отсутствует) в качестве ковариата, дал значительный эффект ковариации, F(1, 27) = 6.17, p = 0.020, значительный основной эффект группы, F(3, 27) = 4.13, p = 0.016, маргинальный сеанс × Ковариальное взаимодействие, p = 0.080, и никаких других значительных эффектов, F(4, 108) <1.48, p > 0.21. Таким образом, неуправляемый (неизбирательный) подход, реагирующий во время тренировки, объясняет значительные различия в локомоторном ответе на 1 мг / кг AMPH. Однако эта вариация не перекрывалась с вариацией, связанной с группой, потому что включение ковариаты в анализ увеличивало, а не уменьшало значимость группового эффекта.
Обсуждение
Данные о носовом суставе, в то время как CS присутствовал, показывают, что группы приобрели связь между CS и доставкой сахарозы, что отражается в увеличении количества ответов на тренинги. Профиль ответа на сеансы в то время как CS присутствовал, предполагал, что расписания 100 и 50% CS-US одинаково эффективны при выработке подхода, тогда как график 25% вызвал более скромное увеличение подхода, вызванного кией. Данные о носовом выступе, в то время как CS отсутствовали, указывают на то, что группы, получившие любой из трех тренировочных графиков CS-сахарозы (группа 25, 50, 100), быстро научились уменьшать свои носовые поры в отсутствие CS, тогда как животные в группе 0 , которые не получили никакой CS, только научились уменьшать свое поведение подхода в ограниченной степени после интенсивного обучения.
Данные по привыканию показывают, что группы не отличались до AMPH, и что повторное воздействие на тестовые ящики было связано с уменьшением реактивного локомоторного ответа без наркотиков. Таким образом, межгрупповые различия и повышенный ответ на повторные дозы AMPH нельзя отнести к ранее существовавшим различиям в локомоторном поведении.
Результаты запроса на предварительную сенсибилизацию с помощью 0.5 мг / кг AMPH подтвердили, что препарат усиливает локомоторную активность по сравнению с окончательным днем без наркотиков. В соответствии с гипотезой группа 50 оценивалась выше, чем группы 0 или 100 (а также группа 25) с точки зрения средней реакции на дозу, хотя средние различия между группами были незначительными.
Для сеансов сенсибилизации запланированные сравнения между группами показали, что предшествующее воздействие 50% условной сахарозы в качестве вознаграждения привело к значительному увеличению локомоторной реакции на дозу амфетамина 1.0 мг / кг по сравнению с тремя другими схемами. Этот эффект был очевиден после первой дозы и существенно не изменился при повторных дозах. Анализ тенденций показал двухфазный ответ (для всего образца) на повторные дозы AMPH, увеличивающийся до третьей дозы и впоследствии снижающийся. Результаты последующего наблюдения ANCOVA с тычками в нос (отсутствие CS) в качестве ковариаты подтвердили, что различия в локомоторных реакциях четырех групп на 1 мг / кг AMPH не были опосредованы подходом без указателей, отвечающим во время тренировок с сахарозой.
Групповой эффект во время сеансов сенсибилизации согласуется с нашей гипотезой. Эффект двухфазной сессии не согласуется с ожидаемой продолжительной эскалацией локомоторных ответов с повторными дозами AMPH. Это может быть связано с интервалом дозирования. Чтобы решить эту проблему, следует использовать процедуру (альтернативные суточные дозы), которая индуцирует последовательную эскалацию локомоторного ответа на дозы 1.0 мг / кг AMPH (т. Е. Поведенческую сенсибилизацию). Воздействие сенсибилизирующего режима AMPH на последующий ответ на вторую проблему 0.5 мг / кг еще больше подтвердит общность этого эффекта. Включение проблемы солевого раствора до AMPH определяло бы роль ожидаемого или связанного с инъекцией (например, стресса) воздействия на локомоторный ответ на AMPH. Включение 75% -устойной сахарозной группы помогло бы выяснить роль неопределенности вознаграждения в сравнении с редкостью вознаграждения по шаблону ответов для групп 50 и группы 25. Кроме того, чтобы дать оценку (по ANCOVA) вклада ответов на подход, вызванный безрецептурным препаратом, в локомоцию под AMPH (с использованием носовых пиков с CS, присутствующими в качестве ковариата), носовые пики были также закодированы для группы 0 в течение интервала, когда CS присутствовал в других четырех группах (т. е. так, чтобы носовые колышки из всех пяти групп, включая группу 0, которая не получала CS, могли быть включены в анализ ковариации с CS, присутствующей в качестве ковариата). Эти уточнения были включены в эксперимент 2.
Experiment 2
Материалы и методы
Методология эксперимента 2 была аналогична методике эксперимента 1, но была пересмотрена, чтобы лучше приблизить схему, которая была достоверно индуцирована сенсибилизацией AMPH (Fletcher и др., 2005). Изменения были следующими: (a) Группа 75% CS-сахарозы (n = 8); (b) Во время обучения сахарозе крысы (кроме группы 0) получали 20 CS (легкие) презентации (в отличие от 15 в эксперименте 1); (c) презентации CS были разделены средним интервалом между судами 90 s; диапазон: 30-180 s (против 120 s в эксперименте 1), что компенсирует увеличение учебных испытаний, чтобы приравнять продолжительность каждой тренировочной сессии к эксперименту 1; (d) продолжительность каждой из трех сеансов привыкания уменьшалась с 120 до 90 min, чтобы соответствовать длительности тестовых сессий; (д) Была добавлена проблема солевого раствора (ip, 1 мл / кг) (90 мин) (день обучения после сахарозы 8), для оценки локомоторных эффектов инъекции сам по себе (например, ожидание, стресс); (f) Сессии сенсибилизации по протоколу 1 мг / кг проводились в альтернативные будни (дни после тренировки 12-21), а не еженедельно, как в эксперименте 1; (g) Наряду с запросом на дозу сенсибилизации 0.5 мг / кг AMPH (после тренировочного дня 9) была добавлена вторая послесенсибилизация 0.5 мг / кг AMPH-пробы (день обучения после сахарозы 28), чтобы проверить общность эффект сенсибилизации через дозы; (h) носовые пики при наличии CS были закодированы для всех групп (включая группу 0); (i) носовые пики, в то время как CS отсутствовали, регистрировались конкретно из интервала 5 непосредственно перед наступлением КС для индексации преждевременного подхода.
Итоги
Носовые токи во время сеансов кондиционирования сахарозы
5 Group × 15 Session × 2 Phase (CS присутствует, CS отсутствует) ANOVA из носовых корешков дало существенные основные эффекты Group, F(4, 19) = 2.89, p = 0.050, сеанс F(14, 266) = 2.28, p = 0.006 и Phase, F(1, 19) = 14.72, p = 0.001, а также значительное трехстороннее взаимодействие, F(56, 266) = 1.38, p = 0.050. Панели (А, В) диаграммы 3 нанесите на график средние баллы групп при наличии CS и отсутствии CS соответственно. Сравнение двух панелей показывает, что основной эффект фазы отражал более общие ответы на тычки носа, когда CS присутствовал, а не отсутствовал. Следовательно, ответные ответы с указанием происходили значительно чаще, чем преждевременные ответы без запроса. Основные эффекты группы и сеанса интерпретировать нелегко из-за взаимодействия более высокого порядка. Этот последний результат отражал конвергенцию оценок для пяти групп на относительно стабильном низком уровне через сеансы, когда CS отсутствовал (рисунок 3B), а также расхождение оценок в высоком (группа 75, группа 100), промежуточное (группа 50) и низкое (группа 0, группа 25) уровни носового мешка, отвечающего на сеансы, когда CS присутствовал (рис. 3A). Из полиномиальных трендов более низкого порядка (линейных, квадратичных, кубических) только трехстороннее взаимодействие для линейного тренда приближалось к значению, F(4, 19) = 2.32, p = 0.094, что отражает общее монотонное нарастание носа в течение сеансов в группе 75 и относительно более быструю стабилизацию при высоких, средних и низких уровнях ответа в других группах, когда CS присутствовал.
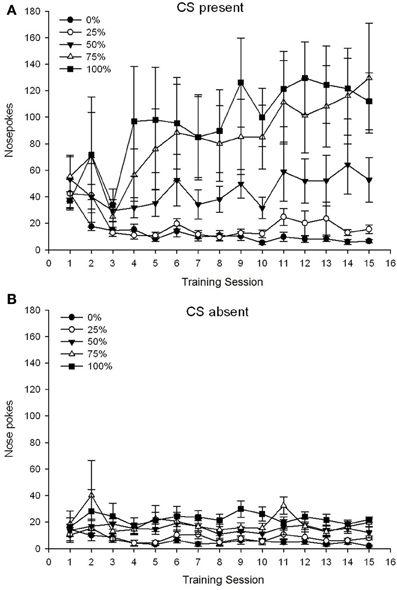
Рисунок 3. Средние (SE) ответы подхода (носовые поры) на 15 сахарозе тренировки в группах крыс Sprague Dawley (n = 8 / группа), получающих вознаграждение за сахарозу (решение 10%), поставляемое в соответствии с графиками 0, 25, 50, 75 или 100%, Условный раздражитель был светом (120 s). Группа 0 получила такое же количество вознаграждений, что и группа 100 в отсутствие условных раздражителей. (A) Оценки при наличии CS (испытания 5 s × 20). (B) Оценки, когда CS отсутствовала (средняя для 5 × 20 s, когда свет был выключен).
Привыкание к локомоторным ящикам
5 Group × 3 Session ANOVA безрецептурных локомоторных ответов дала существенный главный эффект сеанса, F(2, 70) = 60.01, p <0.0001, и никаких других значимых эффектов, F(4, 35) <0.70, p > 0.60. Плановые сравнения группы 50 с группой 0 и с группой 100 на первом и последнем сеансах привыкания не дали значимых эффектов. ts <0.84, p > 0.40. Таким образом, средний локомоторный ответ без лекарств в ключевых группах до тестирования не отличался. Среднее (SE) количество разрывов пучка за 90 минут составило 2162 (118) на сеансе 1, 1470 (116) на сеансе 2 и 1250 (98) на сеансе 3.
Тестирование
Солончаков, Группа 5 × 2 Session ANOVA сравнивает локомоторный отклик на заключительном сеансе привыкания и сеансе сеанса солевого раствора. ANOVA дал основной эффект сеанса, F(1, 35) = 62.46, p <0.0001, и никаких других значимых эффектов, F(4, 35) <0.65, p > 0.64. Рисунок 4 Участки средств группы и показывает, что эффект сеанса отражает общее снижение в ответ опорно-двигательного аппарат от конечной свободных от наркотиков привыкания сессии в солевой сессию, которая не менялась группой. Таким образом, снижение локомоторного ответа, наблюдаемого в течение трех сеансов привыкания, продолжалось в четвертом безрезультатном доступе к тестовым ящикам.
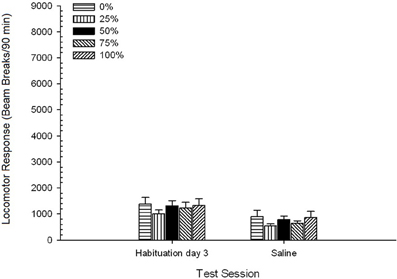
Рисунок 4. Средний (SE) локомоторный отклик (количество разрывов луча в электронной матрице за 90 мин) в последней из сеансов привыкания без рецепта 3 и на последующем сеансе после инъекции солевого раствора (ip, 1 мл / кг) в группах Sprague Dawley крыс (n = 8 / group), ранее подвергавшихся сеансам суточного кондиционирования 15 с вознаграждением сахарозы (решение 10%), поставляемым в соответствии с графиками 0, 25, 50, 75 или 100%, Условный раздражитель был светом (120 s). Группа 0 получила такое же количество вознаграждений, что и группа 100 в отсутствие условных раздражителей.
Эффекты 0.5 мг / кг AMPH.
Предзаводная локомоция, Группа 5 × 2-сессия. ANOVA предторгового локомотива (30-min) на тестовых днях до и после сенсибилизации 0.5 мг / кг AMPH дала значительный основной эффект сеанса, F(1, 35) = 13.39, p = 0.001, и никаких других значительных эффектов, F(4, 35) <1.79, p > 0.15. Запланированные сравнения не обнаружили значительных различий между группой 50 и группой 0 или группой 100 на первом сеансе. t(70) <1.00, p > 0.30. Однако на втором сеансе (после сенсибилизации) группа 50 (1203; 121) показала значительно больше разрывов пучка перед инъекцией (M; SE), чем группа 100 (756; 103), t(70) = 5.11, p <0.001, но не отличался от группы 0 (1126; 211), t(7) <0.88, p > 0.40. Следовательно, базовые различия в локомоции не учитывали групповые различия в локомоторном ответе на первую дозу АМФН 0.5 мг / кг, но могли способствовать различиям между группой 50 и группой 100 в локомоторном ответе на вторую дозу АМФН 0.5 мг / кг. . Средние (SE) значения разрыва пучка для фазы перед инъекцией в первой и второй тестовых сессиях 0.5 мг / кг AMPH составили 757 (41) и 974 (59).
Пост-инъекционная локомоция, Группа 5 × 2 сессия ANOVA от локомоторного ответа до 0.5 мг / кг AMPH до и после сенсибилизирующей схемы дозы 5 дала основной эффект сеанса, F(1, 35) = 76.05, p <0.0001, и никаких других значимых эффектов, F(4, 35) <1.10, p > 0.37. Рисунок 5 показывает средние баллы для каждой группы и сеанса.
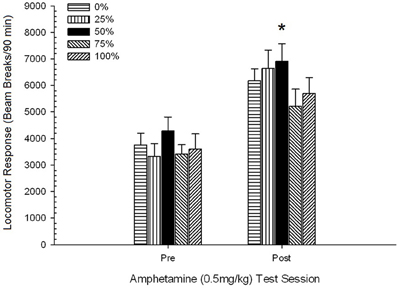
Рисунок 5. Средний (SE) локомоторный отклик (количество разрывов луча в электронной матрице за 90 мин) до 0.5 мг / кг d-амфетамина на отдельных сеансах до и после сенсибилизирующего режима 5-сессии d-амфетамина (1.0 мг / кг; ip за сеанс) в группах крыс Sprague Dawley (n = 8 / group), ранее подвергавшихся сеансам суточного кондиционирования 15 с вознаграждением сахарозы (решение 10%), поставляемым в соответствии с графиками 0, 25, 50, 75 или 100%, Условный раздражитель был светом (120 s). Группа 0 получила такое же количество вознаграждений, что и группа 100 в отсутствие условных раздражителей. *p <0.05 для средней разницы между группой 50 и группой 0, а также группой 100 на основе запланированных сравнений.
На рисунке показано, что эффект сеанса включал значительное увеличение общих разрывов луча (SE) в течение 90 мин от 0.5 мг / кг доза 1, 3674 (216) до 0.5 мг / кг доза 2, 6123 (275). Отсутствие взаимодействия или группового эффекта показало, что сенсибилизация AMPH не зависит от группы. Несмотря на отсутствие значительных эффектов, связанных с группой в ANOVA, проверка этого показателя показывает, что группа 50 показала наибольший ответ как на первую, так и на вторую дозы 0.5 мг / кг. Запланированные сравнения ответа на первую дозу 0.5 мг / кг не выявили существенной разницы между группой 50 и группой 0 или группой 100, t«s(35) <0.48, p > 0.50. Однако в ответ на вторую дозу 0.5 мг / кг (постсенсибилизация) группа 50 продемонстрировала значительно большую двигательную активность, чем группа 0, t(35) = 2.00, p <0.05, а также группа 100, t(35) = 3.29, p <0.01.
В свете существенной разницы между группами в предвзятой локомоции на второй сессии AMNH мг / кг AMNH, описанной выше, была проведена последующая 0.5 Group × 5 сессия ANCOVA с локомоторным ответом на 2 мг / кг AMPH, инъекции на второй сеанс. Этот анализ дал значительный эффект от ковариата, F(1, 34) = 8.65, p = 0.006, основной эффект сеанса F(1, 34) = 10.83, p = 0.002, и никаких других значительных эффектов, F(4, 34) <0.85, p > 0.50. Важно отметить, что запланированные сравнения на основе ошибки MS и ошибки df из ANCOVA подтвердили, что средний локомоторный ответ на вторую дозу AMPH 0.5 мг / кг оставался значительно выше в группе 50, чем в группе 100, t(34) = 3.09, p <0.01, и группа 0, t(34) = 1.88, p <0.05 (односторонний), когда контролировалось изменение перед инъекцией от сеанса 2. Таким образом, группа 50 показала значительно больший локомоторный ответ после сенсибилизации на 0.5 мг / кг AMPH, чем группа 100 или группа 0, и эти групповые различия не были опосредованы передвижением перед инъекцией в дни испытаний.
Эффекты 1.0 мг / кг AMPH.
Предзаводная локомоция, Сеанс 5 × 5-сессия. ANOVA из оценок предварительной инъекции 30-min для сеансов сенсибилизации AMPH 1 мг / кг дало основной эффект сеанса, F(4, 140) = 16.70, p <0.0001, и никаких других значимых эффектов, F(4, 35) <0.94, p > 0.45. Запланированные сравнения не выявили значительной разницы в передвижении перед инъекцией между группой 50 и группой 0 или группой 100 на первом сеансе. t(175) <1.66, p > 0.10. Однако на заключительном сеансе группа 50 (1167; 140) показала значительно больше разрывов пучка (M; SE), чем группа 100 (1000; 99), t(175) = 2.35, p <0.05, но не отличался от группы 0 (1085, 120), t(175) <1.16, p > 0.20. Следовательно, различия в передвижении перед инъекцией вносили вклад в различия между группами 50 и 100 в локомоторном ответе на конечную дозу AMPH 1 мг / кг. Средние (SE) общие разрывы пучка для образца во время фазы предварительной инжекции для сеансов с 1 по 5 составляли: 810 (46), 784 (52), 760 (53), 726 (46), 1009 (51).
Пост-инъекционная локомоция, Группа 5 × 5 Session ANOVA ответов на 1 мг / кг AMPH дала существенный главный эффект сеанса, F(4, 140) = 6.72, p <0.001, маргинальное взаимодействие Группа × Сеанс, F(16, 140) = 1.57, p = 0.085, и никакой основной эффект группы, F(4, 35) <0.44, p > 0.77. Анализ полиномиального тренда выявил значительный линейный тренд, F(1, 35) = 9.19, p = 0.005 и кубический тренд, F(1, 35) = 21.63, p <0.001, за сеансы с 1 по 5. Рисунок 6 показывает средние локомоторные баллы для каждой группы и сеанса.
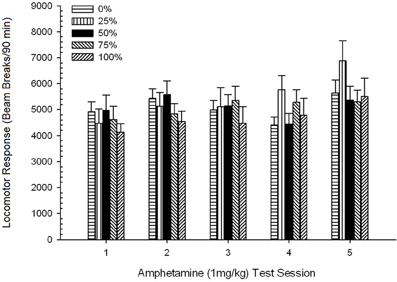
Рисунок 6. Средний (SE) локомоторный отклик (количество разрывов луча в электронной матрице за 90 мин) до 1 мг / кг d-амфетамина (ip) на еженедельных сеансах 5 в группах крыс Sprague Dawley (n = 8 / group), ранее подвергавшихся сеансам суточного кондиционирования 15 с вознаграждением сахарозы (решение 10%), поставляемым в соответствии с графиками 0, 25, 50, 75 или 100%, Условный раздражитель был светом (120 s). Группа 0 получила такое же количество вознаграждений, что и группа 100 в отсутствие условных раздражителей.
На рисунке показано, что эффект сеанса отразил значительное увеличение общих разрывов в среднем (SE) луча для полной выборки из сеанса 1, 4624 (213) на сеанс 5, 5736 (272), что подтверждает появление сенсибилизации к AMPH. Кубический тренд обозначал относительные максимумы на сеансах 1, 3 и 5, с провалами на сессиях 2 и 4, особенно для групп 0 и 50. На рисунке также показано, что, несмотря на отсутствие значительного взаимодействия, группа 25 отображала прогрессивно больший локомоторный ответ по сеансам и значительно отличалась от других групп на сеансах 4 и 5 (9 и 22% соответственно соответственно, чем следующая самая высокая группа). Запланированные сравнения показали, что группа 50 существенно не отличается от групп 0 или 100, t(175) <0.89, p > 0.40 на первом или последнем сеансе теста 1 мг / кг AMPH.
Контроль за вариацией носового мешка в ходе тренировки сахарозы
Две сессии 5 × 2 ANCOVAs от локомоторного ответа до 0.5 мг / кг AMPH до и после режима сенсибилизации, включая тотальные носовые пики при тренировке сахарозы с присутствием CS и отсутствием CS в виде отдельных ковариатов, не обнаружили значимых эффектов ни для ковариата, F(1, 18) <1.03, p > 0.31. Следовательно, подход, отвечающий во время тренировки, не опосредовал групповые различия в ответе на 0.5 мг / кг AMPH.
Две сессии 5 × 5. ANCOVAs локомоторного ответа на 1 мг / кг во время сеансов сенсибилизации с суммарными носовыми парами (CS присутствует, CS отсутствует) в виде отдельных ковариатов не приводили к значительным эффектам ковариации, в то время как CS присутствовал, F(4, 104) <1.04, p > 0.38, и маргинальный главный эффект ковариаты при отсутствии CS, F(1, 18) = 3.32, p = 0.085.
Обсуждение
Результаты этого исследования не всегда подтверждают гипотезу о том, что группа 50 продемонстрирует более высокий локомоторный ответ по сравнению с другими группами. Данные AMPH, содержащие мг / кг 1, подтвердили появление сенсибилизации с использованием режима дозирования на альтернативный день. Структура по группам указывала тенденцию к большей чувствительности во время последних сеансов в группе 25, без таких доказательств для группы 50. Напротив, результаты дозы 0.5 мг / кг показали тенденцию к большей сенсибилизации в группе 50, в то же время подтверждая значительное общее увеличение локомоторного ответа между группами во втором и первой дозе AMNH мг / кг. Нулевой эффект введения физиологического раствора подтвердил, что ожидаемое или связанное с инъекцией стресс не влияет на эффекты AMPH.
Данные о носовом суставе снова показали общее увеличение подхода, отвечающего в ходе учебных занятий, когда CS присутствовал, без соответствующего увеличения, когда CS отсутствовал. Поэтому животные, похоже, приобрели связь между КС и перспективой получения сахарозы. Групповые различия в частоте носовых пиков при присутствии CS соответствовали примерно частоте доставки вознаграждения в соответствии с соответствующими графиками, причем группы 75 и 100 отображали большинство носовых пиков, группа 50 отображала промежуточные числа носовых пиков и группы 0 и 25 показывает наименьшее количество носовых носов. Эти результаты свидетельствуют о том, что КС подошла к тому, чтобы контролировать подход, отвечающий в соответствии с общей вероятностью вознаграждения. Хотя спекулятивное, одно из возможных объяснений низких ставок носового нока с CS, присутствующим в группе 50 в эксперименте 2 против эксперимента 1, может быть сокращением интервала между интервалами, поскольку более длительные интервалы между экспериментами (эксперимент 1), по-видимому, стимулируют импульсивное тенденций, и это связано с увеличением оборота ОД в передних поясницах, предлимбической и инфралимбической коре (Dalley и др., 2002). Таким образом, уменьшение 30% интервала между экспериментами 2 (и 3) может привести к изменению уровней коры головного мозга и способствует более избирательному (т. Е. Руководствуясь относительной частотой вознаграждения) по сравнению с импульсивным (не ориентированным на частоту вознаграждения) отвечая в группе 50 во время учебных испытаний в эксперименте 2 по сравнению с экспериментом 1.
Отсутствие значительных эффектов, связанных с ковариатом для носовых копков в CS-состоянии в ANCOVAs, указывает на то, что подход, отвечающий при тренировке сахарозы, не опосредовал эффекты различных графиков CS-сахарозы при ответах на AMPH. Предельно значимый эффект ковариации для отсутствующего CS-фактора в ANCOVA локомоторных ответов на 1 мг / кг AMPH свидетельствует о том, что тенденция к преждевременному реагированию на наркотики объясняет некоторую изменчивость локомоторных эффектов AMPH во время сеансов сенсибилизации.
В совокупности данные свидетельствуют о том, что эффекты кондиционирования истории могут быть более заметными с 0.5 AMPH, чем с 1 мг / кг AMPH, и что протокол, который генерирует сенсибилизацию в отсутствие каких-либо других манипуляций, может скрыть или сделать излишним эффекты предполагаемого сенсибилизация, способствующая поведенческим манипуляциям (т. е. хроническая переменная награда).
Поведенческая сенсибилизация к AMPH является надежным эффектом в лаборатории. Тем не менее, за пределами лаборатории, только незначительное количество людей, которые играют в азартные игры, хронически перерастают в патологические уровни. Хотя риск сенсибилизации связан с риском для зависимости (или поиска наркотиков), особенно для психостимуляторов (Везина, 2004; Flagel и др., 2008), многие факторы, помимо риска сенсибилизации, могут предрасполагать к зависимости (например, Verdejo-Garcia и др., 2008; Conversano и др., 2012; Volkow и др., 2012). Тем не менее, факторы качества, которые придают уязвимость к сенсибилизации, могут взаимодействовать с историей кондиционирования, чтобы акцентировать влияние непредсказуемой награды (например, 50% CS-US) на реактивность системы DA. Чтобы исследовать эту возможность, эксперимент 3 применял ту же процедуру, что и эксперимент 2, но использовал штамм Льюиса вместо штаммов Sprague Dawley.
Крысы Sprague Dawley демонстрируют промежуточные уровни DA-транспортеров с более низким уровнем, чем крысы штамма Wistar (Zamudio и др., 2005), но более высокие уровни, чем крысы Вистар-Киото («депрессивный» -подобный штамм) в ядре accumbens, миндалине, вентральной тегментальной области и субстанции nigra (Jiao и др., 2003). Этот профиль может приводить к тому, что крысы Sprague Dawley умеренно чувствительны к экологическим или фармакологическим манипуляциям с функцией DA. Напротив, крысы Льюиса демонстрируют низкие уровни DA-переносчиков, а также D2 и D3 DA-рецепторы в ядре accumbens и дорсальной полосатой по сравнению с другими штаммами (например, F344) (Flores и др., 1998). Эти морфологические различия могут вносить вклад в дифференциальную реакцию крыс Льюиса на манипуляции с DA. Крысы Lewis также демонстрируют ряд усиленных реакций на экспериментальные манипуляции с лекарствами по сравнению с другими линиями (например, F344). Что наиболее важно, крысы Lewis демонстрируют большую сенсибилизацию к метамфетамину, характеризующуюся низкой реакцией на начальные дозы, но более высокой реакцией на более поздние дозы (Camp et al., 1994). Крысы Льюиса также проявляют большую локомоторную сенсибилизацию к диапазону доз кокаина (Костен и др., 1994; Haile et al., 2001). Основываясь на этой модели эффектов, мы предположили, что крысы Льюиса позволили нам исследовать, усиливает ли восприимчивость к сенсибилизации эффекты графика подготовки при последующем ответе на AMPH.
Experiment 3
Материалы и методы
Методология была такой же, как в эксперименте 2, кроме использования крыс Льюиса (200-225 g по прибытии, Чарльз-Ривер, Квебек, Канада).
Итоги
Носовые токи во время сеансов кондиционирования сахарозы
5 Group × 15 Session × 2 Phase (CS присутствует, CS отсутствует) ANOVA из носовых корешков дало существенные основные эффекты Group, F(4, 34) = 6.12, p = 0.001, сеанс, F(14, 476) = 3.42, p <0.001, а фаза, F(1, 34) = 20.83, p <0.001, а также значимое трехстороннее взаимодействие, F(56, 476) = 1.56, p = 0.008. Панели (А, В) диаграммы 7 нанесите на график средние баллы групп для фаз присутствия CS и отсутствия CS соответственно. Сравнение двух панелей показывает, что основной эффект фазы отражал более общие ответы на тычки носа, когда CS присутствовал, а не отсутствовал. Следовательно, ответные ответы возникали значительно чаще, чем недозревшие ответы. Основные эффекты группы и сеанса интерпретировать нелегко из-за взаимодействия более высокого порядка. Трехстороннее взаимодействие отражало конвергенцию оценок для пяти групп на относительно стабильном низком уровне через сеансы, когда CS отсутствовал [Панель (B)], а также расхождение баллов, когда CS присутствовала в относительно дискретных профилях для каждой группы, которые совпадали с их рангом порядка частоты вознаграждения: от наивысшего (группа 100) до самого низкого (группа 25) [панель (A)]. Существенным был только линейный тренд взаимодействия, F(4, 34) = 4.03, p = 0.009, что отражает общее постоянное увеличение носа в течение сеансов в группе 100, когда CS присутствовала в сравнении с относительно несогласованным профилем увеличения носа, который заходит на сеансы в других группах на этом этапе.
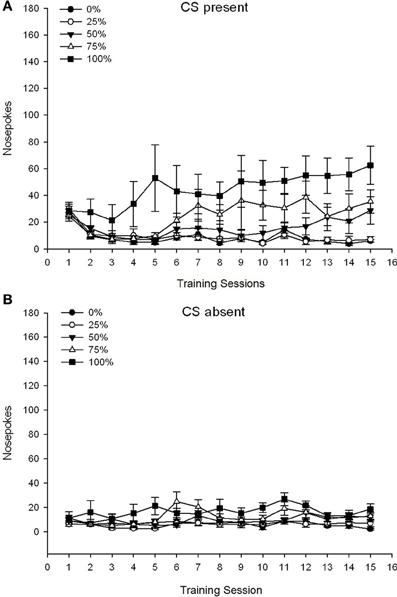
Рисунок 7. Средние (SE) ответы подхода (носовые поры) на 15 сахарозе тренировки в группах крыс Льюиса (n = 8 / группа), получающих вознаграждение за сахарозу (решение 10%), поставляемое в соответствии с графиками 0, 25, 50, 75 или 100%, Условный раздражитель был светом (120 s). Группа 0 получила такое же количество вознаграждений, что и группа 100 в отсутствие условных раздражителей. (A) Оценки при наличии CS (испытания 5 s × 20). (B) Оценки, когда CS отсутствовала (средняя для 5 × 20 s, когда свет был выключен).
Привыкание к локомоторным ящикам
Группа 5 × 3 Session ANOVA дала основной эффект сеанса, F(2, 70) = 23.07, p <0.0001, и никаких других значимых эффектов, F(8, 70) <1.47, p > 0.18. Криволинейный паттерн средних (SE) локомоторных баллов возник с сеанса 1, 1076 (74), через сеанс 2, 644 (48), до сеанса 3, 762 (59). Плановые сравнения группы 50 с группой 0 и с группой 100 на первом и последнем сеансах привыкания выявили значительно меньшее количество перерывов в пучке в группе 50 (M = 911; SE = 109) против группы 0 (M = 1103; SE = 176) на сеансе привыкания 1, t(105) = 2.02, p <0.05, но нет разницы между группой 50 и группой 100 (M = 1066; SE = 150), t(105) <1.20, p > 0.20 на этой сессии. Группа 50 существенно не отличалась ни от группы 0, ни от группы 100 на последнем сеансе привыкания. t(105) <0.93, p > 0.30. Таким образом, средняя двигательная реакция без лекарств в ключевых группах не различалась до тестирования.
Тестирование
Солончаков, Группа 5 × 2 сессия. ANOVA от локомоторных ответов на заключительном сеансе привыкания и сеанса солевого теста дала существенный главный эффект сеанса, F(1, 35) = 50.12, p <0.0001, и никаких других значимых эффектов, F(4, 35) <0.57, p > 0.68. Рисунок 8 показывает средние баллы группы для двух сеансов и указывает, что эффект сеанса отражает значительное снижение от привыкания к физиологическому тесту. Таким образом, получение инъекции сам по себе (например, ожидание, стресс) не улучшали реакцию локомотора.
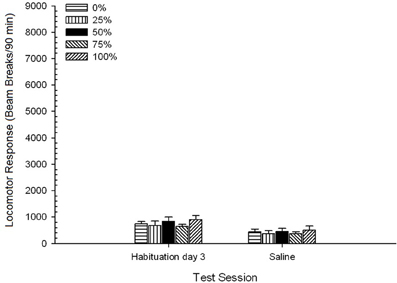
Рисунок 8. Средний (SE) локомоторный отклик (количество разрывов луча в электронной матрице за 90 мин) на последней из сеансов привыкания без рецепта 3 и на последующем сеансе после инъекции солевого раствора (ip, 1 мл / кг) в группах крыс Льюиса (n = 8 / group), ранее подвергавшихся сеансам суточного кондиционирования 15 с вознаграждением сахарозы (решение 10%), поставляемым в соответствии с графиками 0, 25, 50, 75 или 100%, Условный раздражитель был светом (120 s). Группа 0 получила такое же количество вознаграждений, что и группа 100 в отсутствие условных раздражителей.
Эффекты 0.5 мг / кг AMPH.
Предзаводная локомоция, Группа 5 × 2-сессия. ANOVA предвоенной локомоции дала существенный главный эффект сеанса, F(1, 35) = 15.04, p <0.001, и никаких других значимых эффектов, F(4, 35) <1.19, p > 0.33. Запланированные сравнения не выявили значительных различий между группой 50 и группой 0 или группой 100 ни в одной из тестовых сессий, t(70) <0.99, p > 0.30. Таким образом, базовые различия в локомоции до инъекции не учитывали групповые различия в локомоторном ответе на 0.5 мг / кг AMPH. Средние (SE) интервалы пучка для фазы перед инъекцией для первого и второго (после сенсибилизации) сеансов 0.5 мг / кг составили 325 (25) и 473 (36).
Пост-инъекционная локомоция, Группа 5 × 2 сессия ANOVA от локомоторного ответа на дозы 0.5 мг / кг, до и после хронического 1 мг / кг AMPH, дала основной эффект сеанса, F(1, 34) = 87.44, p <0.0001, и никаких других значимых эффектов, F(4, 34) <0.94, p > 0.45. Рисунок 9 подсчитывает средние локомоторные баллы для каждой группы и сеанса и показывает, что эффект сеанса отражает увеличение общего ответа на вторую дозу 0.5 мг / кг в соответствии с сенсибилизацией. На рисунке также показано, что группы выполнялись очень точно в сеансе 1, но эта группа 50 отображала большую локомоторную активность, чем другие группы в сеансе 2. Запланированные сравнения в ответ на первую дозу 0.5 мг / кг не выявили существенных различий между группой 50 и группой 0 или группой 100, t(35) <1.28, p > 0.20. Однако группа 50 продемонстрировала значительно больший локомоторный ответ на вторую дозу 0.5 мг / кг, чем группа 0, t(35) = 4.32, p <0.001, или группа 100, t(35) = 2.24, p <0.05.
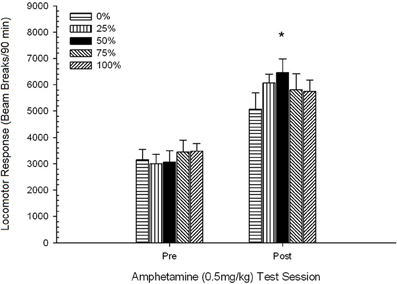
Рисунок 9. Средний (SE) локомоторный отклик (количество разрывов луча в электронной матрице за 90 мин) до 0.5 мг / кг d-амфетамина на отдельных сеансах до и после сенсибилизирующего режима 5-сессии d-амфетамина (1.0 мг / кг; ip за сеанс) в группах крыс Льюиса (n = 8 / group), ранее подвергавшихся сеансам суточного кондиционирования 15 с вознаграждением сахарозы (решение 10%), поставляемым в соответствии с графиками 0, 25, 50, 75 или 100%, Условный раздражитель был светом (120 s). Группа 0 получила такое же количество вознаграждений, что и группа 100 в отсутствие условных раздражителей. *p <0.05 для средней разницы между группой 50 и группой 0, а также группой 100 на основе запланированных сравнений.
Эффекты 1 мг / кг AMPH.
Предзаводная локомоция, Сеанс 5 × 5-сессия. ANOVA из оценок предварительной инъекции 30-min для сеансов сенсибилизации дало основной эффект сеанса, F(4, 140) = 4.10, p = 0.004, и никаких других значительных эффектов, F(4, 35) = 1.25, p > 0.31. Запланированные сравнения показали, что разрывы пучка во время фазы перед инъекцией (M; SE) были значительно ниже в группе 50 (395; 62), чем в группе 100 (508; 62), t(175) = 2.58, p <0.01, но не группа 0, t(175) <1.83, p > 0.10, на сеансе 1 АМФГ 1 мг / кг. На последнем сеансе АМФГ 1 мг / кг запланированные сравнения также показали, что локомоция перед инъекцией в группе 50 (378; 60) была значительно ниже, чем в группе 100 (650; 75). ), t(175) = 6.17, p <0.001, но не в группе 0, t(175) <1.84, p > 0.10. Поскольку направление этих групповых различий (контрольная группа = группа 50) было противоположным гипотетическому паттерну, групповые различия в локомоции после инъекции, которые совпадают с гипотезой, не могут быть отнесены к различиям исходного уровня до инъекции. Средние (SE) общие разрывы пучка во время фазы предварительной инъекции для сеансов с 1 по 5 составляли: 442 (34), 452 (32), 542 (40), 411 (26), 504 (37).
Пост-инъекционная локомоция, Группа 5 × 5 Сеансы ANOVA ответов на дозы 1 мг / кг дали существенный главный эффект сеанса, F(4, 140) = 6.15, p <0.001, и никаких других значимых эффектов, F(4, 35) <0.57, p > 0.68. Анализ полиномиального тренда выявил значительный линейный тренд, F(1, 35) = 9.34, p = 0.004 и кубический тренд, F(1, 35) = 5.08, p = 0.031, последний результат обозначает относительные максимумы на сеансах 3 и 5. фигура 10 подсчитывает эти оценки и показывает, что, несмотря на отсутствие значительного взаимодействия в ANOVA, группа 50 демонстрировала значительно большую локомоцию, чем остальные четыре группы в ответ на конечную дозу 1 мг / кг. Соответственно, запланированные сравнения показали значительно более высокие средние баллы в сеансе 5 в группе 50, чем во всех других группах, t(35) > 3.68, p <0.001.
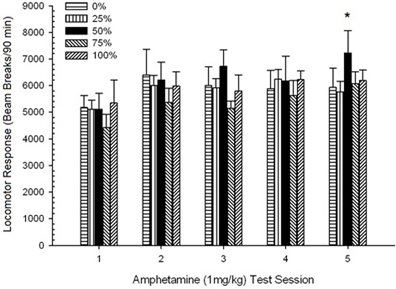
Рисунок 10. Средний (SE) локомоторный отклик (количество разрывов луча в электронной матрице за 90 мин) до 1 мг / кг d-амфетамина (ip) на еженедельных сеансах 5 в группах крыс Льюиса (n = 8 / group), ранее подвергавшихся сеансам суточного кондиционирования 15 с вознаграждением сахарозы (решение 10%), поставляемым в соответствии с графиками 0, 25, 50, 75 или 100%, Условный раздражитель был светом (120 s). Группа 0 получила такое же количество вознаграждений, что и группа 100 в отсутствие условных раздражителей. *p <0.05 для средней разницы между группой 50 и группой 0, а также группой 100 на основе запланированных сравнений.
Контроль за вариацией носового мешка в ходе тренировки сахарозы
Две сессии 5 × 2 ANCOVAs от локомоторного ответа до 0.5 мг / кг AMPH до и после режима сенсибилизации, включая тотальные носовые пики при тренировке сахарозы с присутствием CS и отсутствием CS в виде отдельных ковариатов, не обнаружили значимых эффектов ни для ковариата, F(1, 32) <0.44 p > 0.51. Два 5 групповых × 5 сеансов ANCOVA локомоторного ответа на 1 мг / кг AMPH во время сеансов сенсибилизации с полным тычком в нос (CS присутствует, CS отсутствует) в качестве отдельных ковариат не дали значительных эффектов ковариаты, пока CS присутствовал или отсутствовал. F(1, 33) <0.14, p > 0.71. Таким образом, ответ без лекарств не учитывал групповые различия в локомоторных ответах на любую дозу AMPH.
Обсуждение
Сенсибилизация развивалась к действию повторного приема амфетамина 1.0 мг / кг. Данные о привыкании и физиологическом растворе подтверждают, что этот эффект не был вызван ранее существовавшими различиями, ожиданием или стрессовой реакцией на инъекцию. Результаты анализа ANCOVA с тычками в нос подтверждают, что эти эффекты не были следствием подхода без наркотиков. Сами данные по «тыкаванию носа» показали, что группы приобрели связь между CS и перспективой вознаграждения сахарозой. Ранговый уровень реакции групп на тыканье носа в конце тренировки соответствовал общей частоте вознаграждения по разным графикам от самого высокого (группа 100) до самого низкого (группа 0), как это было в эксперименте 2. Относительно более низкое общее среднее значение Уровни тыкающих носом в этом эксперименте по сравнению с экспериментами 1 и 2 могут отражать более избирательный подход, реагирующий на сигналы для вознаграждения у крыс Льюиса (Костен и др., 2007).
Данные дозы 0.5 мг / кг показали, что исходный локомоторный ответ AMPH у крыс Льюиса (рис. 9) был несколько подавлен по сравнению с крысами Sprague Dawley (эксперимент 2; рисунок 5), но внутригрупповое увеличение реакции на вторую дозу у крыс Льюиса было значительным (почти в два раза больше ответа на первую дозу 0.5 мг / кг) после режима AMPH-сеанса 5. В частности, группа 50 проявляла больший отклик локомотора чем все группы, кроме группы 25, во вторую (то есть постсенсибилизацию) дозу 0.5 мг / кг AMPH и больший локомоторный ответ, чем все другие группы, включая группу 25, до конечной дозы AMPH мг / кг 1 (заключительная сессия сенсибилизации) ,
Сводный анализ группового рейтинга в экспериментах
Для определения достоверности групповых различий в сенсибилизации непараметрический анализ оценивал случайность между группой и рангом среднего локомоторного ответа на вторую (пост-хроническую AMPH) дозу 0.5 мг / кг и конечную дозу AMPH 1.0 мг / кг из экспериментов 3. Анализ дал значительный эффект: φ = 0.986, p = 0.025, что отражает тот факт, что группа 50 занимает первое место во всех, кроме одного из сравнений. Высший ранг группы 50 по сравнению со всеми другими группами в ответ на вторую (пост-хроническую AMPH) дозу 0.5 мг / кг изображен на рисунке 5 (эксперимент 2) и рисунок 9 (эксперимент 3). Высший ранг группы 50 относительно других групп в ответ на конечную дозу 1.0 мг / кг изображен на рисунке 2 (эксперимент 1) и рисунок 10 (эксперимент 3). Единственным исключением из этого шаблона был ответ на конечную дозу 1.0 мг / кг у крыс Sprague-Dawley в эксперименте 2.
Обсуждение
Настоящая серия экспериментов проверила гипотезу о том, что хроническое воздействие азартных игр в виде графика вознаграждения может сенсибилизировать мозговые пути DA, подобно хроническому воздействию наркотиков. Доказательства такого эффекта предполагают, что нейропластичность того же типа, которая, как считается, способствует наркомании, может быть вызвана хроническим воздействием непредсказуемых графиков вознаграждения. В соответствии с литературой по наркотической зависимости, локомоторный ответ на дозы 0.5 и 1.0 мг / кг АМФН с индексированной реакцией DA-системы с большей локомоцией в ответ на более поздние дозы, оперативно определяющие сенсибилизацию (см. Робинсон и Берридж, 1993; Пирс и Каливас, 1997; Вандершурен и Каливас, 2000).
В целом, результаты соответствуют нашей гипотезе. Однако они также указывают на значительную изменчивость экспериментальных эффектов из-за процедурных факторов. Эффект графика кондиционирования был скромным, но последовательным, а группа 50 демонстрировала больший ответ, чем остальные четыре группы, в обе дозы после пяти режимов приема. Хотя в целом F-значения для связанных с группой эффектов в анализе дисперсии часто были незначительными, различия в ключевых группах были подтверждены с помощью парных запланированных сравнений. В этой связи следует отметить, что «настоящее мышление, однако, заключается в том, что общее значение [для F в ANOVA] не требуется. Прежде всего, гипотезы, проверенные общим тестом и тестом с множественным сравнением, совершенно разные, с совершенно разными уровнями мощности. Например, общий F фактически распределяет различия между группами по количеству степеней свободы для групп. Это приводит к размыванию общей F в ситуации, когда несколько групповых средств равны друг другу, но отличаются от некоторого другого среднего »(Howell, 1992, п. 338). Это именно та ситуация, которая применялась в настоящих экспериментах, где ожидается, что группа 50 будет отличаться от элементов управления группы 0 и группы 100, но никакой разницы между этими контрольными группами не было предсказано для группы 25 или группы 75.
Данные носового сустава подтвердили, что в каждом эксперименте животные приобрели связь между КС и перспективой получения сахарозы. Соответствие между частотой носового носа для разных групп и общей частотой вознаграждения в соответствии с их соответствующими графиками обучения свидетельствует о том, что средняя скорость приема сахарозы под руководством безрецептурного подхода отвечает. Тем не менее, отсутствие опосредованного эффекта носовых пиков на групповых локомоторных ответах на AMPH в ANCOVAs указывает на то, что отдельные процессы лежат в основе двух видов поведения.
В некоторых случаях эффект графика подготовки был очевидным в ответ на первую дозу AMPH; в других случаях он появился только после повторных доз. Групповые различия в локомоторном ответе на первую дозу AMPH свидетельствуют о том, что воздействие графиков вознаграждений, связанных с азартными играми, является само по себе достаточным для стимулирования сенсибилизации. Групповые различия в локомоции после нескольких доз AMPH указывают на более тонкий эффект, который можно охарактеризовать как «восприимчивость», которая проявляется только в сочетании с постоянным воздействием первичного сенсибилизирующего агента (то есть амфетамином).
Различия в структуре ответа в экспериментах свидетельствуют о том, что более длительный интервал между тренировкой и первоначальной задачей AMPH может максимизировать возможность обнаружения неотъемлемого сенсибилизирующего эффекта кондиционирующего лечения. Это, в свою очередь, предполагает, что эффекты условного воздействия вознаграждения могут инкубироваться с течением времени, явление, также наблюдаемое при стимулирующей сенсибилизации (Grimm и др., 2006). Модель ответа на две дозы амфетамина предполагает, что доза 0.5 мг / кг может быть более эффективной в выявлении эффектов кондиционирования истории. Это, в свою очередь, предполагает, что эффекты кондиционирования в соответствии с текущим протоколом обучения несколько тонкие и могут быть замаскированы эффектами потолка в дозах AMPH и условиями, которые генерируют De Novo сенсибилизация.
В эксперименте 3 двухфазная картина ответа на дозы 0.5 мг / кг и прогрессирующее появление превосходства в группе 50 согласуется с ожидаемым профилем для крыс Льюиса в ответ на метамфетамин (Camp et al., 1994). Это подкрепляет обоснованность настоящих выводов и предполагает совпадение факторов, которые снижают уязвимость к сенсибилизации психостимулятора и к графикам вознаграждения, подобным азартным играм.
Во всех экспериментах локомоторный отклик после сенсибилизации группы 50 обычно превышал по сравнению с другими группами в разных дозах амфетамина и разных штаммов животных. Тем не менее, высокая внутригрупповая изменчивость и скромные размеры межгрупповых эффектов указывают на роль других факторов в реакционной способности ДС к амфетамину после воздействия различных графиков условной компенсации сахарозы. Хотя ответы нейронов DA на сигналы вознаграждения могут служить грубой моделью азартных игр (Fiorillo и др., 2003), как и все модели, есть потеря информации ради выражения - то есть, чтобы продемонстрировать ключевой процесс. В результате картина эффектов в условиях CS-US в оригинальной Fiorillo et al. исследование не полностью обобщает на локомоторный ответ на амфетамин. Дальнейшие уточнения модели призваны полностью охватить аспекты азартных игр, которые влияют на функцию системы DA.
В совокупности результаты этой серии экспериментов обеспечивают предварительную поддержку гипотезы о том, что хроническое воздействие азартных игр в виде графиков вознаграждения повышает реактивность системы DA мозга на вызов психостимулятора. Таким образом, они расширяют результаты Singer et al. (2012) который продемонстрировал, что по сравнению с фиксированным графиком предварительное воздействие переменного графика подкрепления в оперантной парадигме усиливает последующий локомоторный ответ на амфетамин. Более конкретно, настоящие данные указывают на неопределенность доставки вознаграждения в качестве важного фактора, лежащего в основе влияния переменной вознаграждения. Величина эффектов в оперантной парадигме была значительно больше, чем эффекты, обнаруженные в настоящих экспериментах. Это может отражать большее хроническое воздействие азартной игры (55 против 15 дней); он может отражать эффекты, требующие ответа операндов, чтобы получить вознаграждение (то есть роль для агентства), а не пассивное воздействие, как в настоящем исследовании. Увеличение продолжительности обучения в нынешней парадигме поможет решить эти вопросы.
Из этих экспериментов нельзя почерпнуть правильность графиков вознаграждений за вознаграждение и подкрепления как моделей азартных игр. Будущие исследования, которые анализируют влияние истории кондиционирования на поведение, связанное с риском в играх с грызунами, могут решить эту проблему. Точно так же соответствие между поведенческой сенсибилизацией, найденной здесь, и повышенным стригальным ответом ДА на амфетамин, недавно обнаруженный у патологических игроков, должно ждать дальнейшего исследования (Boileau et al., 2013). Микродиализ мог бы решить этот вопрос, и предсказание, основанное на данных о человеке, будет заключаться в том, что более высокий выброс DA в группе 50 «фенотип азартных игр» будет наиболее четко наблюдаться на дорсальной (сенсомоторной) полосатой, а не вентральной (лимбической) полосатой , Валидация воздействия 50% переменной CS + вознаграждения в этих других парадигмах будет поддерживать его полезность в качестве добросовестной экспериментальной модели PG.
В то время как некоторые формы азартных игр явно влекут за собой инструментальный ответ (например, игровые автоматы), в других формах азартных игр (например, лотерея) связь между действием (покупкой билета, т. Е. Размещением ставки), сигналами для получения вознаграждения (т. Е. , номера лотереи), а сама награда (выигрышный номер и денежная выплата) гораздо более расплывчата. Тем не менее, активация DA во время интервала CS-US вполне может произойти. Это может объяснить, почему, когда объявляется «выигрышный номер», внимание приковывается к тому, что каждый отдельный шар для лотереи последовательно падает, чтобы составить определенную последовательность цифр в выигрышном номере. Хотя вероятность появления конкретной цифры определяется математически, результат для каждого отдельного шара лотереи является двоичным - попадание (совпадает с номером игрока) или промах (не совпадает с номером игрока) - и результат любого заданного испытания неизвестен. Такой сценарий может лучше охарактеризовать опыт группы 50 в настоящих экспериментах, где вознаграждение предоставлялось не случайно, но также и непредсказуемо, и CS просто указывал потенциал вознаграждения, не раскрывая, произойдет ли оно в данном испытании. Игровые автоматы более тесно связаны с PG, чем лотерейные билеты (Cox et al., 2000; Bakken et al., 2009), что указывает на важную роль инструментальных факторов (и непосредственности) в полезных аспектах азартных игр для этого населения (Loba и др., 2001). Тем не менее, павловский процесс, смоделированный в настоящих экспериментах (CS + неопределенное вознаграждение), представляется необходимым, если не достаточным элементом опыта азартных игр.
Наряду с отсутствием четкого инструментального требования, ряд других конструктивных особенностей, возможно, способствовали относительно скромной и переменной структуре экспериментальных эффектов. Группы отличались общим воздействием сахарозы, а также непредвиденным случаем между CS и сахарозой. Хотя это, возможно, способствовало изменчивости между группами, он не может легко объяснить, почему животные с наибольшим воздействием сахарозы (группа 100) проявляют меньшую чувствительность, чем группа 50. Кроме того, группа 0 не получала стимула до воздействия сахарозы на каждом испытании. Хотя это исключало ожидаемое вознаграждение, вызванное кий, оно не контролировало наличие стимула перед доставкой вознаграждения, которая существовала во всех других группах. Чтобы решить эту проблему, будущие исследования должны включать условие, при котором животные получают вознаграждение за каждое испытание после случайного воздействия на нейтральный стимул (т. Е. Присутствие которого не сигнализирует о возможности вознаграждения).
Еще одним ограничением дизайна является потенциальное появление дополнительного поведения, которое может повлиять на последствия графика обучения. Перед лицом неопределенности животные могут развивать суеверное поведение, предназначенное для усиления воспринимаемого контроля и уменьшения вызванной неопределенностью активации DA (см. Harris et al., 2013). Поэтому возможно, что неконтролируемые аспекты экспериментальной конструкции позволили животным компенсировать эффекты графика подготовки. Такой эффект может способствовать относительно скромному и переменному ответу на амфетамин в группе 50 после тренировки CS + сахарозы. В будущих исследованиях следует записывать спонтанное поведение, помимо носовых мышей, во время тренировок, чтобы проверить эту возможность, и контролировать ее статистически, если она возникнет. Поскольку такое поведение, как ожидается, будет противодействовать или ослабить последствия неопределенности, обусловленной графиком, локомоторный ответ на амфетамин в группе 50 должен быть усилен, когда он контролируется (процедурно или статистически). Поэтому настоящий (неконтролируемый) дизайн обеспечивает консервативную проверку эффектов вознаграждения 50% CS + на сенсибилизацию амфетамина.
Что касается внешней действительности, использование самцов крыс также ограничивает обобщаемость результатов. Отсутствие четкого условия «наказания» также отличается от азартных игр, где большие денежные потери являются общими и оказывают важное мотивационное воздействие (Nieuwenhuis et al., 2005; Сингх и Хан, 2012). Было обнаружено, что способность накапливать вознаграждение отсутствует в настоящей парадигме, и накопленные выигрыши в игре с игровым автоматом взаимодействуют с манипуляциями с DA в организме человека (Tremblay и др., 2011; Smart и др., 2013). Аналогичным образом, возможность для джекпота является важным различием между настоящей моделью и фактической азартной игрой.
Несмотря на эти ограничения, настоящие результаты свидетельствуют о том, что вознаграждение CSN за 50% может зависеть от путей DA, связанных с усилением эффектов азартных игр (Fiorillo и др., 2003; Ансельме, 2013). Кросс-сенсибилизация ответа на AMPH после этого графика, подобного азартным играм, согласуется с ключевой ролью DA в азартных играх и эффектах психостимулятора (Зак и Пулос, 2009), и распространяет более ранние исследования по перекрестной загрузке мотивации к азартной игре AMPH у патологических игроков (Зак и Пулос, 2004). Настоящие результаты также косвенно свидетельствуют о том, что умеренные дозы AMPH, которые не вызывают супра-физиологического высвобождения DA, могут лучше моделировать активность мозга в ответ на прерывистые сигналы вознаграждения (т. Е. Во время азартных игр), чем воздействие высокого (то есть, похожего на выпивку) дозы стимулирующих препаратов (см. Вандершурен и Пирс, 2010). Прямая поддержка этого соответствия может быть получена путем оценки выпуска DA в ответ на 50% -ную переменную CS-US и различные дозы AMPH с использованием микродиализа.
С экспериментальной точки зрения, настоящая модель Павлова и предыдущая операционная модель переменного армирования, как представляется, порождают фенотип, похожий на человека, патологический игрок. Таким образом, они обеспечивают ценное дополнение к задачам игры в грызуны, которые моделируют поведение в азартных играх (как зависимую меру), но до сих пор используют только здоровых животных, что эквивалентно социальным игрокам в мире. Основываясь на литературе, животные, хронически подверженные переменному вознаграждению, могут сильно различаться в этих задачах, особенно в ответ на ДА-эргические препараты. Сочетание фенотипа азартных игр с азартными играми с азартными играми может позволить систематическое развитие лекарств для лечения ПГ, что может быть не полностью достигнуто только здоровыми животными. Дальнейшие уточнения в экспериментальной схеме проектирования и обучения, как описано выше, должны улучшить соответствие между животными, обученными этой парадигме и фактическим патологическим игрокам.
С клинико-социологической точки зрения обнаружение того, что подверженность 50% переменной CS + вознаграждение, которое близко соответствует графику вознаграждения на коммерческом игровом автомате (Tremblay и др., 2011), изменяет систему DA мозга надежными и прочными способами, позволяет предположить, что в некоторых случаях активность азартных игр, например наркотики, может быть «патогеном», способным вызвать зависимость. Тем не менее, скромный размер эффекта и высокая изменчивость в ответ на вознаграждение 50% CS + свидетельствуют о том, что, как и наркотики злоупотребления, тенденция к графикам вознаграждений, связанных с азартными играми, для стимулирования наркомании будет в значительной степени зависеть от ранее существовавшего профиля риска игрока. Тем не менее, для того, чтобы не подвергать людей с высоким уровнем риска потенциальным неблагоприятным эффектам, связанным с азартными играми, представляется разумным, что политика, применяемая для предотвращения использования и сведения к минимуму вреда от наркотиков, злоупотребляющих наркотиками, может быть распространена и на азартные игры.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Это исследование было профинансировано грантами Совета по естественным наукам и инженерным исследованиям Канады Павлу Флетчеру. Мы искренне благодарим г-жу Джурджу Джорджевич за подготовку цифр.
Рекомендации
Ансельме, П. (2013). Допамин, мотивация и эволюционное значение азартных игр. Behav. Brain Res, 256, 1-4. doi: 10.1016 / j.bbr.2013.07.039
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
APA (2013). Диагностическое и статистическое руководство по психическим расстройствам, 5th Edn, Арлингтон, VA: Американская психиатрическая публикация.
Bakken, IJ, Gotestam, KG, Grawe, RW, Wenzel, HG и Oren, A. (2009). Азартные игры и проблемы с азартными играми в Норвегии 2007. Сканд. J. Psychol, 50, 333-339. doi: 10.1111 / j.1467-9450.2009.00713.x
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Boileau, I., Payer, D., Chugani, B., Lobo, DS, Houle, S., Wilson, AA, et al. (2013). в естественных условиях доказательство более значительного высвобождения дофамина, вызванного амфетамином, в патологической азартной игре: исследование позитронной эмиссионной томографии с [C] - (+) - PHNO. Mol. психиатрия doi: 10.1038 / mp.2013.163. [Epub перед печатью].
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Bolles, RC (1972). Усиление, ожидание и обучение. Psychol. оборот, 79, 394-409. doi: 10.1037 / h0033120
Camp, DM, Browman, KE и Robinson, TE (1994). Влияние метамфетамина и кокаина на моторное поведение и внеклеточный допамин в брюшном половом члене крыс Льюиса против Fischer 344. Brain Res. 668, 180–193. doi: 10.1016/0006-8993(94)90523-1
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Конверсано, К., Мараццити, Д., Кармасси, К., Балдини, С., Барнабей, Г., и Делль'Оссо, Л. (2012). Патологическая азартная игра: систематический обзор биохимических, нейровизуализационных и нейропсихологических данных. Харв. Преподобная психиатрия 20, 130-148. doi: 10.3109 / 10673229.2012.694318
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Cox, BJ, Kwong, J., Michaud, V. и Enns, MW (2000). Проблема и вероятная патологическая азартная игра: соображения из опроса сообщества. Можно. J. Психиатрия 45, 548-553.
Dalley, JW, Theobald, DE, Eagle, DM, Passetti, F. и Robbins, TW (2002). Дефицит в импульсном контроле, связанный с тонически повышенной серотонинергической функцией в передней префронтальной коре крыс. Нейропсихофармакологии 26, 716–728. doi: 10.1016/S0893-133X(01)00412-2
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Everitt, BJ и Robbins, TW (2005). Нейронные системы арматуры для наркомании: от действий к привычкам к принуждению. Туземный Neurosci, 8, 1481-1489. doi: 10.1038 / nn1579
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Fiorillo, CD, Tobler, PN и Schultz, W. (2003). Дискретное кодирование вероятности вознаграждения и неопределенности с помощью дофаминовых нейронов. Наука 299, 1898-1902. doi: 10.1126 / science.1077349
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Flagel, SB, Watson, SJ, Akil, H. и Robinson, TE (2008). Индивидуальные отличия в атрибуции стимула значимости к рекомендации, связанной с вознаграждением: влияние на сенсибилизацию кокаина. Behav. Brain Res, 186, 48-56. doi: 10.1016 / j.bbr.2007.07.022
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Fletcher, PJ, Tenn, CC, Rizos, Z., Lovic, V. и Kapur, S. (2005). Сенсибилизация к амфетамину, но не PCP, снижает смещение набора внимания: разворачивание агонистом рецептора D1, вводимым в медиальную префронтальную кору. Психофармакология (Berl.) 183, 190–200. doi: 10.1007/s00213-005-0157-6
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Flores, G., Wood, GK, Barbeau, D., Quirion, R. и Srivastava, LK (1998). Льюис и Фишер: сравнение уровней переносчиков допамина и рецепторов. Brain Res. 814, 34–40. doi: 10.1016/S0006-8993(98)01011-7
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Frascella, J., Potenza, MN, Brown, LL и Childress, AR (2010). Общедоступные мозговые уязвимости открывают путь для невнимательных зависимостей: вырезание наркомании в новом суставе? Энн. NY Acad. Sci, 1187, 294-315. doi: 10.1111 / j.1749-6632.2009.05420.x
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Grimm, JW, Buse, C., Manaois, M., Osincup, D., Fyall, A. и Wells, B. (2006). Временно-зависимая диссоциация эффектов доза-реакция кокаина на тягу к сахарозе и локомоцию. Behav. Pharmacol, 17, 143-149. doi: 10.1097 / 01.fbp.0000190686.23103.f8
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Хайле, CN, Hiroi, N., Nestler, EJ и Kosten, TA (2001). Дифференциальные поведенческие реакции на кокаин связаны с динамикой мезолимбических дофаминовых белков у крыс Льюиса и Фишера 344. Synapse 41, 179-190. doi: 10.1002 / syn.1073
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Харрис, JA, Andrew, BJ и Kwok, DW (2013). Подход к журналу при подаче пищи зависит от павловского, а не инструментального, кондиционирования. J. Exp. Psychol. Anim. Behav. Процесс, 39, 107-116. doi: 10.1037 / a0031315
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Howell, DC (1992). Статистические методы психологии, Бостон, Массачусетс: Даксбери.
Ito, R., Dalley, JW, Robbins, TW и Everitt, BJ (2002). Допамин высвобождается в дорсальном полосатом теле во время кокаиноподобного поведения под контролем связанного с наркотиками кия. J. Neurosci, 22, 6247-6253.
Jiao, X., Pare, WP и Tejani-Butt, S. (2003). Различия в растяжении в распределении участков транспортера допамина в мозге крысы. Prog. Neuropsychopharmacol. Biol. Психиатрия 27, 913–919. doi: 10.1016/S0278-5846(03)00150-7
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Koob, GF и Le Moal, M. (2008). Обзор. Нейробиологические механизмы для мотивационных процессов противника в зависимости. Philos. Сделка R. Soc. Лонд. B Biol. Sci, 363, 3113-3123. doi: 10.1098 / rstb.2008.0094
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Kosten, TA, Miserendino, MJ, Chi, S. и Nestler, EJ (1994). Фишерские и Льюисские штаммы крыс демонстрируют дифференциальные эффекты кокаина в обусловленном предпочтении места и поведенческой сенсибилизации, но не в двигательной активности или условном неприятии вкуса. J. Pharmacol. Exp. Ther, 269, 137-144.
Костен, Т.А., Чжан, XY и Хайле, CN (2007). Разделительные различия в поддержании самообслуживания кокаина и их взаимосвязи с реакциями активности новизны. Behav. Neurosci, 121, 380-388. doi: 10.1037 / 0735-7044.121.2.380
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Leeman, RF и Potenza, MN (2012). Сходства и различия между патологическими азартными играми и расстройствами употребления психоактивных веществ: фокус на импульсивность и компульсивность. Психофармакология (Berl.) 219, 469–490. doi: 10.1007/s00213-011-2550-7
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Leshner, AI (1997). Наркомания - это заболевание мозга, и это важно. Наука 278, 45-47. doi: 10.1126 / science.278.5335.45
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Loba, P., Stewart, SH, Klein, RM и Blackburn, JR (2001). Манипуляции с особенностями игр стандартного видео лотерейного терминала (VLT): эффекты у патологических и непатологических игроков. J. Gambl. шпилька, 17, 297-320. doi: 10.1023 / A: 1013639729908
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Martinez, D., Narendran, R., Foltin, RW, Slifstein, M., Hwang, DR, Broft, A., et al. (2007). Освобожденное амфетамином дофаминовое выделение: заметно затуплено в зависимости от кокаина и прогнозирует выбор для самостоятельного введения кокаина. Am. J. Психиатрия 164, 622-629. doi: 10.1176 / appi.ajp.164.4.622
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Mateo, Y., Lack, CM, Morgan, D., Roberts, DC и Jones, SR (2005). Уменьшенная терминальная функция допамина и нечувствительность к кокаину после самоубийства и депривации кокаина. Нейропсихофармакологии 30, 1455-1463. doi: 10.1038 / sj.npp.1300687
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Nestler, EJ (2001). Молекулярная основа долговременной пластичности, лежащая в основе зависимости. Туземный Rev. Neurosci, 2, 119-128. doi: 10.1038 / 35053570
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Nieuwenhuis, S., Heslenfeld, DJ, von Geusau, NJ, Mars, RB, Holroyd, CB и Yeung, N. (2005). Активность в чувствительных к человеку областях мозга сильно зависит от контекста. Neuroimage 25, 1302-1309. doi: 10.1016 / j.neuroimage.2004.12.043
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Pierce, RC и Kalivas, PW (1997). Схематическая модель выражения поведенческой сенсибилизации к амфетаминоподобным психостимуляторам. Brain Res. Brain Res. оборот, 25, 192. doi: 10.1016 / S0165-0173 (97) 00021-0
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Robbins, TW и Everitt, BJ (1999). Наркомания: плохие привычки складываются. природа 398, 567-570. doi: 10.1038 / 19208
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Робинсон, TE, Беккер, JB и Presty, SK (1982). Долгосрочное облегчение индуцированного амфетамином ротационного поведения и освобождения полового члена допамина, вызванного однократным воздействием амфетамина: половые различия. Brain Res. 253, 231–241. doi: 10.1016/0006-8993(82)90690-4
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Robinson, TE и Berridge, KC (1993). Нейронная основа тяги к наркотикам: теория склонности к сенсибилизации. Brain Res. Brain Res. оборот, 18, 247-291. doi: 10.1016 / 0165-0173 (93) 90013-P
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Robinson, TE и Berridge, KC (2001). Инсентив-сенсибилизация и зависимость. Наркомания 96, 103-114. doi: 10.1046 / j.1360-0443.2001.9611038.x
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Певец, Б. Ф., Скотт-Рейлтон, Дж. И Везина П. (2012). Непредсказуемое укрепление сахарина усиливает локомотор, реагирующий на амфетамин. Behav. Brain Res, 226, 340-344. doi: 10.1016 / j.bbr.2011.09.003
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Сингх, В. и Хан, А. (2012). Принятие решений в вариантах вознаграждения и наказания задачи азартных игр в Айове: доказательства «предвидения» или «обрамления»? Фронт. Neurosci, 6: 107. doi: 10.3389 / fnins.2012.00107
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Скиннер, BF (1953). Наука и поведение человека, Нью-Йорк, Нью-Йорк: Свободная пресса.
Smart, K., Desmond, RC, Poulos, CX и Zack, M. (2013). Модафинил увеличивает награду в игре игрового автомата с низкой и высокой импульсивностью патологических игроков. Нейрофармакология 73, 66-74. doi: 10.1016 / j.neuropharm.2013.05.015
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Tremblay, AM, Desmond, RC, Poulos, CX и Zack, M. (2011). Haloperidol изменяет инструментальные аспекты азартных игр в игровых автоматах у патологических игроков и здорового контроля. Addict. Biol, 16, 467-484. doi: 10.1111 / j.1369-1600.2010.00208.x
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Vanderschuren, LJ и Kalivas, PW (2000). Изменения в дофаминергической и глутаматергической передаче при индукции и экспрессии поведенческой сенсибилизации: критический обзор доклинических исследований. Психофармакология (Berl.) 151, 99-120. doi: 10.1007 / s002130000493
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Vanderschuren, LJ и Pierce, RC (2010). Сенсибилизация в наркотической зависимости. Тек. Верхний. Behav. Neurosci, 3, 179-195. doi: 10.1007 / 7854_2009_21
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Vanderschuren, LJ, Schoffelmeer, AN, Mulder, AH и De Vries, TJ (1999). Допаминергические механизмы, опосредующие долгосрочную экспрессию локомоторной сенсибилизации после предварительного воздействия морфина или амфетамина. Психофармакология (Berl.) 143, 244-253. doi: 10.1007 / s002130050943
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Verdejo-Garcia, A., Lawrence, AJ и Clark L. (2008). Импульсивность как маркер уязвимости для расстройств, связанных с употреблением психоактивных веществ: обзор результатов исследований высокого риска, проблемных игроков и исследований генетических ассоциаций. Neurosci. Biobehav. оборот, 32, 777-810. doi: 10.1016 / j.neubiorev.2007.11.003
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Везина П. (2004). Сенсибилизация реактивности дофаминовых нейронов среднего мозга и самолечение психомоторных стимуляторов. Neurosci. Biobehav. оборот, 27, 827-839. doi: 10.1016 / j.neubiorev.2003.11.001
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Volkow, ND, Wang, GJ, Fowler, JS, Logan, J., Gatley, SJ, Hitzemann, R., et al. (1997). Снижение половой допаминергической реакции у детоксифицированных кокаинозависимых субъектов. природа 386, 830-833. doi: 10.1038 / 386830a0
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Volkow, ND, Wang, GJ, Fowler, JS и Tomasi, D. (2012). Схема зависимости в мозге человека. Annu. Rev. Pharmacol. Toxicol, 52, 321-336. doi: 10.1146 / annurev-pharmtox-010611-134625
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Винер, Б. (ред.). (1971). Статистические принципы экспериментального проектирования, Нью-Йорк, Нью-Йорк: Макгроу-Хилл.
Zack, M. и Poulos, CX (2004). Мотивация амфетаминовых цепей для азартных игр и связанных с азартными играми семантических сетей в проблемных игроках. Нейропсихофармакологии 29, 195-207. doi: 10.1038 / sj.npp.1300333
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Zack, M. и Poulos, CX (2009). Параллельные роли допамина в патологической азартной игре и психостимуляторной зависимости. Тек. Преступление за злоупотребление наркотиками, 2, 11-25. doi: 10.2174 / 1874473710902010011
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Zamudio, S., Fregoso, T., Miranda, A., De La Cruz, F., and Flores, G. (2005). Различия в деформации уровней рецепторов допамина и поведения, связанного с допамином у крыс. Brain Res. бык, 65, 339-347. doi: 10.1016 / j.brainresbull.2005.01.009
Pubmed Abstract | Полный текст Pubmed | Полный текст CrossRef
Ключевые слова: патологическая азартная игра, сенсибилизация, амфетамин, допамин, неопределенность
Ссылка: Zack M, Featherstone RE, Mathewson S и Fletcher PJ (2014). Хроническое воздействие азартных игр в виде графика поощрительных стимулов вознаграждения может способствовать повышению чувствительности к амфетамину у крыс. Фронт. Behav. Neurosci. 8: 36. doi: 10.3389 / fnbeh.2014.00036
Поступило в редакцию: 01 Ноябрь 2013; Принято: 23 Январь 2014;
Опубликовано онлайн: 11 Февраль 2014.
Под редакцией:
Брайан Ф. Сингер, Мичиганский университет, США
Рассмотрено:
Louk Vanderschuren, Университет Утрехта, Нидерланды
Рууд Ван Ден Бос, Университет Радбуд Неймеген, Нидерланды
Патрик Ансельм, Университет Льежа, Бельгия
Copyright © 2014 Зак, Фезерстоун, Мэтьюсон и Флетчер. Это статья с открытым доступом, распространяемая в соответствии с условиями Лицензия Creative Commons Attribution (CC BY), Использование, распространение или воспроизведение на других форумах разрешено при условии, что оригинальный автор (ы) или лицензиар зачисляются и что оригинальная публикация в этом журнале цитируется в соответствии с принятой академической практикой. Запрещается использование, распространение или воспроизведение, которое не соответствует этим условиям.
* Корреспонденция: Мартин Зак, Лаборатория когнитивной психофармакологии, Нейронаука, Центр наркомании и психического здоровья, 33 Russell Street, Торонто, ON M5S 2S1, Канада e-mail: [электронная почта защищена]
