Přední. Behav. Neurosci., 23 březen 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2,
Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2, 

- 1Translační výzkumný ústav a institut pro zdravotnické a biomedicínské inovace, Queensland University of Technology, Brisbane, QLD, Austrálie
- 2Škola biomedicínských věd, University of Queensland, Brisbane, QLD, Austrálie
Moderní strava se stala velmi slazenou, což vedlo k bezprecedentní míře spotřeby cukru, zejména mezi adolescenty. Zatímco je známo, že chronický dlouhodobý příjem cukru přispívá k rozvoji metabolických poruch, včetně obezity a diabetu typu II, je známo jen málo, co se týká přímých důsledků dlouhodobé konzumace cukru podobného nárazu na mozek. Bprotože cukr může způsobit uvolňování dopaminu v nucleus accumbens (NAc) podobně jako drogy zneužívání, zkoumali jsme změny v morfologii neuronů v této mozkové oblasti po krátkých (4 týdnech) a dlouhodobých (12 týdnech) binge- jako je spotřeba sacharózy pomocí občasného paradigmatu s výběrem dvou lahví. Barvení Golgi-Cox jsme použili k impregnaci středně ostnatých neuronů (MSN) z jádra NAc a slupky potkanů konzumujících krátkodobou a dlouhodobou sacharózu a porovnali jsme je s kontrolami vody odpovídající věku. Ukazujeme, že prodloužená spotřeba sacharózy typu buldozeru významně snížila celkovou dendritickou délku skořápky MSN NAc ve srovnání s kontrolními krysy odpovídajícími věku. Zjistili jsme také, že restrukturalizace těchto neuronů byla způsobena především sníženou distální dendritickou složitostí. Naopak jsme pozorovali zvýšenou hustotu páteře na distálních větvích MSN NAc shell od dlouhodobých krys konzumujících sacharózu. Souhrnně tyto výsledky zdůrazňují neuronální účinky prodlouženého nadpřirozeného příjmu sacharózy na morfologii MSN NAc shell.
Úvod
V posledních letech 40u byl zaznamenán nárůst spotřeby nápojů slazených cukrem a potravin obsahujících přidané cukry (Nielsen a kol., 2002; Popkin, 2010; Ng a kol., 2012), se zprávami odhadujícími, že až 75% všech potravin a nápojů obsahuje vysoké množství přidaného cukru (Ford a Dietz, 2013; Bray a Popkin, 2014). Během tohoto období došlo také ke zvýšení výskytu obezity a diabetu typu II, zejména u adolescentů (Arslanian, 2002; Reinehr, 2013; Dabelea a kol., 2014; Fryar a kol., 2014). Nedávné studie ukázaly, že děti s nadváhou a obézními dětmi často konzumují vysoké množství přidaného cukru, nicméně příspěvek diet s vysokým obsahem cukru ke zvýšenému výskytu obézních a obézních dětí zůstává kontroverzní (Hu, 2013; Bray a Popkin, 2014; Bucher Della Torre a kol., 2015).
Zatímco rostoucí počet důkazů naznačuje, že konzumace diet s vysokým obsahem cukru může částečně přispět k nárůstu tělesné hmotnosti u dětí a dospívajících (Malik a kol., 2010; Te Morenga a kol., 2013; Bray a Popkin, 2014), byla věnována menší pozornost nepříznivým nemetabolickým důsledkům vyplývajícím z nadměrného příjmu cukru. Je zajímavé, že některé běžné behaviorální a psychologické vzorce se často objevují mezi podmnožinou těch, kteří nad jídlem a udržováním diet s vysokým obsahem cukru. Nejpozoruhodnější je vývoj poruch příjmu potravy, včetně nadměrného požití, v kombinaci se současným nástupem psychologických příznaků, včetně nedostatečné motivace a deprese (shrnuto v Sheehan a Herman, 2015). Kromě toho, protože jednotlivci, kteří jedí binge, často vykazují ztrátu kontroly a neschopnost omezit svůj příjem cukru, je pravděpodobné, že tato chování vznikají v důsledku neurologických adaptací v mozkových oblastech, které hodnotí hedonickou hodnotu vysoce chutného jídla (Saper a kol., 2002; Lutter a Nestler, 2009; Kenny, 2011). Toto zdůvodnění je podporováno i důkazy u lidí, které prokazují, že cukr a sladkost mohou způsobit touhu, která je podobná touhám vyvolaným návykovými drogami, jako je alkohol a nikotin (Volkow a kol., 2012).
Ačkoli návykové vlastnosti cukru jsou stále spekulativní, tato pozorování kombinovaná se studiemi prokazujícími vliv nadměrného příjmu cukru na změnách v odměňování obvodů a rozvoji návykových chování a emočních stavů ve zvířecích modelech (Avena a kol., 2008; Benton, 2010; Ventura a kol., 2014), zaručuje potřebu dalšího vyšetřování. Předchozí studie na hlodavcích ukázaly, že přerušovaný přístup k sacharóze mění aktivitu několika neurotransmiterů v mezolimbickém systému, včetně dopaminu, opioidů a acetylcholinu. Avena a kol., 2008). Ukázalo se, že spotřeba sacharózy podobná nárazům usnadňuje uvolňování dopaminu v nucleus accumbens (NAc), podobně jako zneužívání drog (Avena a kol., 2008). Dále jsme ukázali, že dlouhodobá konzumace sacharózy pomocí paradigmatu volby dvou lahví s přerušovaným přístupem 24 h (Simms a kol., 2008) moduluje expresi nikotinového acetylcholinového receptoru (nAChR) v NAc (Shariff a kol., V tisku). Je zajímavé, že jsme také pozorovali, že sloučeniny nAChR, o kterých je známo, že modulují dopaminovou a acetylcholinovou aktivitu v NAc, mají různé účinky na spotřebu sacharózy po krátkodobém a dlouhodobém příjmu (Shariff a kol., V tisku).
I když tyto studie prokázaly podobnosti v behaviorálních a neurochemických změnách způsobených přerušovaným přístupem k cukru a zneužívání drog, není známo, zda tyto účinky usnadňují změny v morfologii neuronů v NAc. Tjeho je na rozdíl od látek zneužívání, včetně kokainu, amfetaminu a nikotinu, které způsobují dobře charakterizované změny v morfologii středně ostnatých neuronů (MSN) v NAc, včetně zvýšené hustoty páteře a změněné dendritické složitosti (Robinson a Kolb, 1999, 2004; Li et al., 2003; Crombag a kol., 2005). Protože jsme již dříve ukázali, že dlouhodobá expozice (12 týden) alkoholu a sacharóze pomocí intermitentního paradigmatu volby dvou lahví vede k rozdílné odpovědi na farmakoterapeutické intervence ve srovnání s krátkodobým příjmem (4 týdny; Steensland a kol., 2007; Shariff a kol., V tisku), hodnotili jsme účinky krátkodobé a dlouhodobé spotřeby sacharózy na morfologii MSN v NAc. Umožnili jsme dospívajícím krysám konzumovat sacharózu podobným způsobem po dobu 4 (krátkodobé) nebo 12 (dlouhodobé) týdny a poté jsme analyzovali morfologii NAc MSN z krátkodobých a dlouhodobých krys konzumujících sacharózu a porovnali jsme ji s věkově přizpůsobené kontroly, kterým byl umožněn přístup pouze k vodě. Naše výsledky ukazují, že MSN ze skořápky NAc se mění po dlouhodobé, ale ne krátkodobé spotřebě sacharózy, mají sníženou dendritickou délku, ale zvýšenou hustotu distální dendritické páteře. Dále jsme zjistili, že morfologie MSN z jádra NAc zůstala po krátkodobé a dlouhodobé spotřebě sacharózy relativně neporušená. Tyto výsledky poukazují na přímý neurologický důsledek dlouhodobé konzumace sacharózy nárazovým způsobem. Tato data dále prokazují potřebu dalších studií zaměřených na objasnění molekulárních a neurochemických změn, které doprovázejí morfologickou restrukturalizaci MSN ulity NAc navozenou prodlouženým přítokem podobné sacharózy.
Materiály a metody
Etické prohlášení
Všechny experimentální postupy byly provedeny v souladu s australským zákoníkem pro péči o zvířata a jejich použití pro vědecké účely, 8th Edition (National Health and Medical Research Council, 2013). Protokoly byly schváleny Výborem pro etiku zvířat v Queenslandské technologické etice a Výborem pro etiku zvířat v Queenslandské univerzitě.
Zvířata a ustájení
Pět týdnů staré (dospívající) krysy kmene Wistar (kontrola: 176.4 ± 4.8 g; sacharóza: 178.3 ± 5.0 g) (ARC, WA, Austrálie), byly jednotlivě umístěny ve větraných dvojúrovňových plexi® klece. Krysy byly aklimatizovány na individuální podmínky ustájení, manipulaci a cyklus reverzního světla 5 dny před zahájením experimentů. Všechny krysy byly chovány v klimaticky kontrolovaném 12-h cyklu s obráceným světlem / tmou (světla vypnuta v 9 am) se standardní krysou krmivem a vodou podle libosti.
Paradigma pitné vody s přerušovaným přístupem na dvě láhve
Intermitentní přístup 5% sacharóza, dvě láhve, pitné paradigma (Simms a kol., 2008) byl upraven od Moudrý (1973). Všechny tekutiny byly prezentovány v plastových lahvích s odměrkou 300 ml s výtoky z nerezové oceli vloženými dvěma průchodkami v přední části klece po zahájení cyklu temného světla. Hmotnosti každé láhve byly zaznamenány před předložením láhve. Současně byly předloženy dvě láhve: jedna láhev obsahující vodu; druhá láhev obsahující 5% (hm./obj.) sacharózu. Umístění lahvičky se sacharózou 5% (hm./obj.) Bylo střídáno s každou expozicí pro kontrolu vedlejších preferencí. Lahve byly zváženy 24 h po předložení tekutin a měření byla provedena s přesností na 0.1 g. Hmotnost každé krysy byla měřena také pro výpočet gramů příjmu sacharózy na kilogram tělesné hmotnosti. V den 1 doby pití, krysy (n = 6 – 9) byl umožněn přístup k jedné láhvi 5% (w / v) sacharózy a jedné láhvi vody. Po 24 h byla sacharosová láhev nahrazena druhou lahví na vodu, která byla k dispozici pro další 24 h. Tento vzor se opakoval ve středu a v pátek. Krysy měly ve všech ostatních dnech neomezený přístup k vodě. Konzumní spotřeba sacharózy měla za následek eskalaci celkového příjmu sacharózy (ml) v čase (doplňkový obrázek) 1) a byl doprovázen stabilními výchozími úrovněmi pití na základě tělesné hmotnosti [20 ± 5 g / kg 5% (w / v)] během krátkodobých [~ 4 týdnů (13 pití))] a dlouhodobých [ ~ 12 týdny (pití 37)] pití. Samostatná skupina kontrolních potkanů (n = 6 – 9) byl umožněn přístup k vodě v obou lahvích (tj. Bez sacharózy) za stejných podmínek popsaných výše. Průměrná tělesná hmotnost potkanů konzumujících sacharózu na konci krátkodobé expozice byla 405.7 ± 40.8 ga 426.4 ± 31.2 g. Na konci dlouhodobé expozice byla průměrná tělesná hmotnost u kontrolních a sacharózových skupin 578.8 ± 53.4 ga 600.2 ± 45.2 g.
Golgi-Coxovo barvení
Po posledním pití se krysy přenesly ze zvířecího zařízení, aby se umožnilo zpracování vzorků mozku v histologickém zařízení na School of Biomedical Sciences, University of Queensland (St Lucia, Austrálie). Všechna schválená opatření byla přijata ke snížení stresu během přepravy, poté byly krysy ponechány zotavit se přes noc. Následující den byly krysy usmrceny předávkováním pentobarbitalem sodným (60 – 80 mg / kg, ip Vetcare, Brisbane, Austrálie) a intrakardiálně perfundovány umělou mozkomíšní tekutinou 300 ml, která obsahovala (v mM): 130 NaCl, 3 KCl, 26 NaHCO3, 1.25 NaH2PO45 MgCl2, 1 CaCl2a 10 D-glukóza. Každé zvíře bylo poté dekapitováno a mozek odstraněn a inkubován ve tmě v roztoku Golgi-Cox, který obsahoval 5% dichroman draselný, 5% chroman draselný a 5% chlorid rtuťnatý (všechny chemikálie od Sigma-Aldrich), který byl vyroben čerstvý 3 dny před obětováním, jak bylo popsáno dříve (Rutledge a kol., 1969). Metody inkubace barviva Golgi-Cox a metody následného zpracování byly modifikovány Ranjan a Mallick (2010). Mozky od zvířat krátkodobě konzumujících sacharózu byly inkubovány po dobu 6 dní při 37 ° C, zatímco mozky od zvířat dlouhodobě konzumujících sacharózu byly inkubovány po dobu 10 dní, s jednou změnou na čerstvý roztok Golgi-Cox po 4 dnech inkubace.
Po inkubaci byly řezy 300 um koronální řezy pomocí vibračního mikrotomu Zeiss Hyrax V50 (Carl Zeiss, Německo). Plátky byly potom umístěny postupně do destiček pro 24-jamky naplněných 30% (hm./obj.) Sacharózou v 0.1 M fosfátem pufrovaném solném roztoku a zpracovány, jak je uvedeno v (Ranjan a Mallick, 2010). Stručně, řezy byly dehydratovány v 50% ethanolu po dobu 5 min, pak umístěny do 0.1 M NH4OH roztok pro 30 min, dvakrát propláchnutý destilovanou vodou po dobu 5 min a umístěn do fixátoru filmu Fujihunt (Fujifilm, Singapur) na 30 min ve tmě. Plátky byly poté dvakrát propláchnuty v destilované vodě po dobu 2 min a dehydratovány v 70, 90, 95 a 100% ethanolu dvakrát po 5 min. Řezy byly poté vyčištěny v roztoku CXA (1: 1: 1 chloroform: xylen: alkohol) po dobu 10 min a namontovány v DPX (Sigma-Aldrich) na sklíčka Superfrost Plus (Menzel-Glaser, Lomb Scientific, Austrálie) a kryty skluzem (Menzel-Glaser, Německo). Sklíčka byla ponechána ve tmě uschnout při pokojové teplotě přes noc.
Neuronální selekce a sledování v Nucleus Accumbens
Koronální řezy mezi bregma + 2.8 a + 1.7 byly zjišťovány na MSN v jádru a skořápce NAc pomocí boční komory a přední komiseře jako orientačních bodů pomocí atlasu mozku krysy (Paxinos a Watson, 2007) (Obr 1). Funkce obrysu v Neurolucida 7 (MBF Bioscience, VT, USA) byla použita k vymezení jádra NAc a skořápky NAc v každém řezu (obrázek) 2). Mezi neurony 2 a 9 na region na zvíře byly sledovány parametry dendritické délky s použitím objektivu 63x nebo hustoty páteře (hlášeny jako páteře na 100 μm) s použitím objektivu 100x na Zeiss Axioskop II (Carl Zeiss, Německo) pomocí automatizovaného xyz fáze řízená Neurolucida® Software 7 (MBF Biosciences, VT, USA). Veškeré sledování bylo provedeno slepě s ohledem na léčbu. Morfologické parametry neuronů impregnovaných Golgi-Coxem byly analyzovány podobným způsobem jako v předchozích zprávách (Klenowski a kol., 2015).
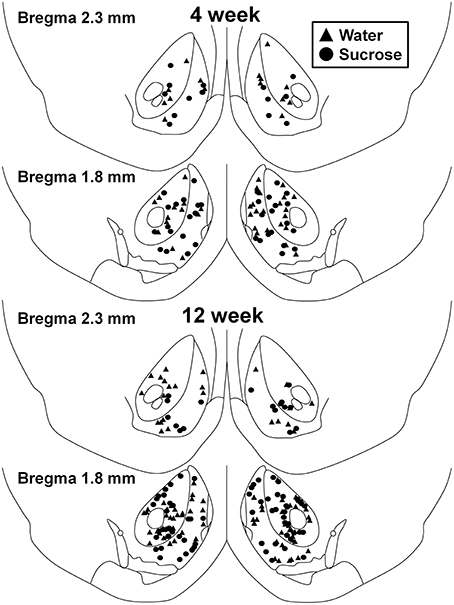
Obrázek 1. Mapa ukazující umístění středních ostnatých neuronů odebraných z jádra accumbens jádra a skořápky 4 a 12 týdně sacharózy konzumující krysy a kontroly podle věku. Horní dva panely ukazují umístění neuronů odebraných z jádra accumbens jádra a skořápky zvířat 4 v týdnu kontroly (trojúhelníky) a sacharózy (kruhy) zvířat. Spodní dva panely ukazují pozice neuronů odebraných z kontrolních zvířat 12 (trojúhelníky) a sacharózy (kruhy).
Statistická analýza
Průměrná a standardní chyba průměru (SEM) byla vypočtena pro každý soubor dat se zvířetem jako n, pomocí středních dat morfometrie ze všech NAN MSN jádra nebo skořepiny (n = 7 pro NAc shell a n = 6 pro jádro NAc 4 za týden, n = 9 pro 12týdenní skupiny). Tam, kde je to uvedeno, nespárovaný Studentův ocas t- byly provedeny testy nebo dvousměrné ANOVA s Bonferroni post-testy pro všechny analýzy zahrnující porovnání skupinových prostředků, s použitím GraphPad Prism verze 6.02 (GraphPad Software, San Diego, CA). Statistická významnost byla přijata na P <0.05. Všechna data v sekci výsledků jsou uvedena jako průměr ± SEM. Procentní změny se počítají jako relativní k kontrolní hodnotě.
výsledky
Středně ostnaté neurony ze skořápky Nucleus Accumbens mají sníženou dendritickou délku, sníženou dendritickou složitost, ale zvýšenou střední hustotu páteře při distálních větvích objednávek po dlouhodobé, ale nikoli krátkodobé spotřebě sacharózy
Po krátkodobé spotřebě sacharózy (4 týdny) nebyly žádné významné rozdíly v morfometrických parametrech MSN NAc shell (tabulka) 1). Rovněž nebyly zjištěny žádné významné rozdíly mezi krátkodobou spotřebou sacharosy a MSNs NAc pro kontrolu vody v analýzách týkajících se pořadí odstředivých větví. Jmenovitě dendritické segmenty na pobočkovou objednávku (P = 0.4111), střední dendritická délka na větev (P = 0.5581) a průměrná hustota páteře na pořadí větví (P = 0.2977, dvousměrné ANOVA) se mezi skupinami významně nelišily. Mapa umístění ukazující přibližné polohy vzorkovaných neuronů je znázorněna na obrázku 1.
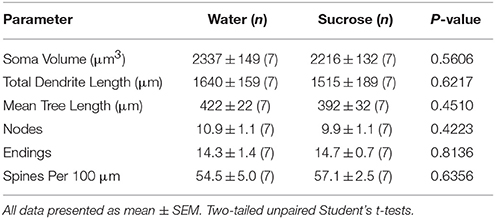
Tabulka 1. Obecné morfologické parametry středně ostnatých neuronů z jádra accumbens slupky potkanů s krátkodobou konzumací sacharózy a kontrolních vod odpovídajících věku.
Po dlouhodobé spotřebě sacharózy (12 týdny) byla celková délka dendritického trnu MSN skořápky NAc snížena o 21% ve srovnání s kontrolami spotřebovávajícími vodu (voda: 1827 ± 148 μm, n = 9; Sacharóza 1449 ± 78 μm, n = 9, *P = 0.0384, dvoustranný nepárový Student t-test, obrázek 2, Stůl 2). Porovnání průměrného počtu dendritických bifurkací (uzlů) a dendritických zakončení mezi skupinami voda a sacharóza odhalilo sníženou (i když nevýznamnou) hladinu dendritické komplexity v MSNs NAc shellu (uzly: Voda 12.9 ± 1.4) n = 9, sacharóza 10.1 ± 0.8 n = 9, P = 0.0879; koncovky: Voda 17.9 ± 1.4 n = 9, sacharóza 14.8 ± 0.7 n = 9, P = 0.0657, dvoustranný nepárový Student t-test, tabulka 2). V soma objemu nedošlo k žádné změně (P = 0.9400), střední délka dendritického stromu (P = 0.1646) nebo celková hustota páteře (P = 0.3662) v MSNs lastury NAc od krys s dlouhodobou konzumací sacharózy ve srovnání s kontrolami s vodou. Tyto morfometrické parametry jsou podrobně uvedeny v tabulce 2.
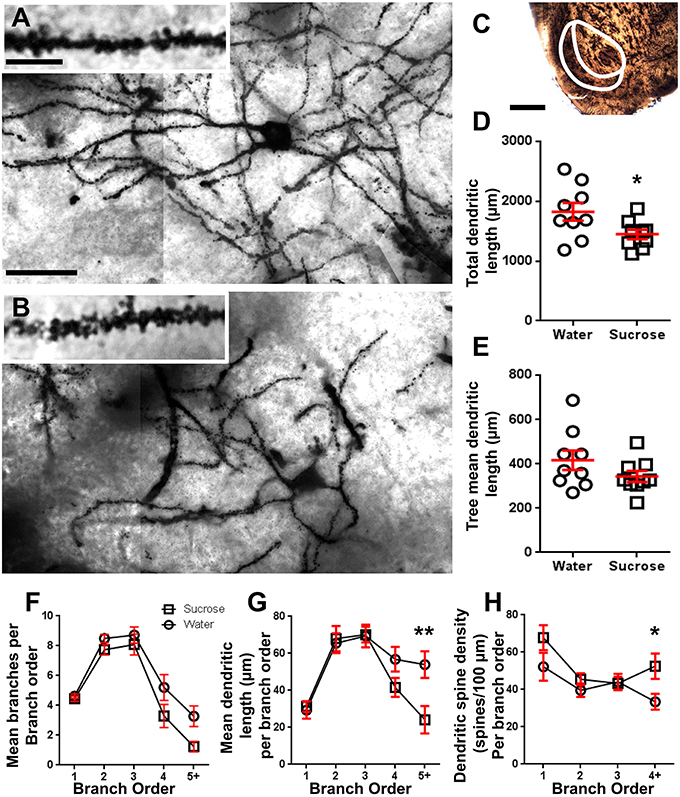 Obrázek 2. Snížená délka dendritického trnu a zvýšená hustota distální dendritické páteře středních ostnatých neuronů (MSNs) ze skořápky nucleus accumbens (NAc) dlouhodobě potkanů ošetřených sacharózou ve srovnání s kontrolními krysy. (A, B) ukazují reprezentace kontrolního (top) a dlouhodobého (12 týdně) sacharosy (dolní) ošetřeného jasného pole z- vyložte mozaiky Golgi-impregnovaných MSN ze skořápky NAc (zvětšení 63x). Začátek (A, B) ukazuje kontrolní a dlouhodobé sacharózou ošetřené světelné polní snímky Golgi-impregnovaných MSN dendritů a dendritických hřbetů z NAc shellu (100x zvětšení). (C) ukazuje anatomické oblasti, ze kterých byly v této studii odebrány vzorky MSN. (D) ukazuje rozptýlený graf sníženého celkového dendritického altánku MSN (průměr ± SEM) ze skořápky NAc u dlouhodobých zvířat se sacharózou (čtverce) ve srovnání s kontrolami (kruhy), nespárovaní studenti t-test, *P <0.05, n = 9; ovládání a n = 9; 12 týdně sacharóza. (E) ukazuje rozptýlený graf nezměněné střední délky dendritického stromu MSN (průměr ± SEM) od skořápky NAc u dlouhodobých zvířat se sacharózou (čtverce) ve srovnání s kontrolami (kruhy), nespárovaní studenti t-test, P > 0.05, n = 9; ovládání a n = 9; 12 týdně sacharóza. Analýza větveného řádu (průměr ± SEM) čísla dendritického segmentu na pořadí větví (F), střední dendritická délka na větev (G) a dendritická hustota páteře na pořadí větví (H). Dlouhodobá spotřeba sacharózy snížila dendritickou délku při distálních větvích (5 +) a zvýšila hustotu dendritické páteře při distálních větvích (4 +) ve srovnání s kontrolami (G, H), dvousměrné ANOVA s Bonferroni po testech, *P <0.05, **P <0.01, n = 9; ovládání a n = 9; dlouhodobá sacharóza. Měřítko: (A, B) = 20 μm; vložený (A, B) = 10 μm; (C) = 1 mm.
Obrázek 2. Snížená délka dendritického trnu a zvýšená hustota distální dendritické páteře středních ostnatých neuronů (MSNs) ze skořápky nucleus accumbens (NAc) dlouhodobě potkanů ošetřených sacharózou ve srovnání s kontrolními krysy. (A, B) ukazují reprezentace kontrolního (top) a dlouhodobého (12 týdně) sacharosy (dolní) ošetřeného jasného pole z- vyložte mozaiky Golgi-impregnovaných MSN ze skořápky NAc (zvětšení 63x). Začátek (A, B) ukazuje kontrolní a dlouhodobé sacharózou ošetřené světelné polní snímky Golgi-impregnovaných MSN dendritů a dendritických hřbetů z NAc shellu (100x zvětšení). (C) ukazuje anatomické oblasti, ze kterých byly v této studii odebrány vzorky MSN. (D) ukazuje rozptýlený graf sníženého celkového dendritického altánku MSN (průměr ± SEM) ze skořápky NAc u dlouhodobých zvířat se sacharózou (čtverce) ve srovnání s kontrolami (kruhy), nespárovaní studenti t-test, *P <0.05, n = 9; ovládání a n = 9; 12 týdně sacharóza. (E) ukazuje rozptýlený graf nezměněné střední délky dendritického stromu MSN (průměr ± SEM) od skořápky NAc u dlouhodobých zvířat se sacharózou (čtverce) ve srovnání s kontrolami (kruhy), nespárovaní studenti t-test, P > 0.05, n = 9; ovládání a n = 9; 12 týdně sacharóza. Analýza větveného řádu (průměr ± SEM) čísla dendritického segmentu na pořadí větví (F), střední dendritická délka na větev (G) a dendritická hustota páteře na pořadí větví (H). Dlouhodobá spotřeba sacharózy snížila dendritickou délku při distálních větvích (5 +) a zvýšila hustotu dendritické páteře při distálních větvích (4 +) ve srovnání s kontrolami (G, H), dvousměrné ANOVA s Bonferroni po testech, *P <0.05, **P <0.01, n = 9; ovládání a n = 9; dlouhodobá sacharóza. Měřítko: (A, B) = 20 μm; vložený (A, B) = 10 μm; (C) = 1 mm.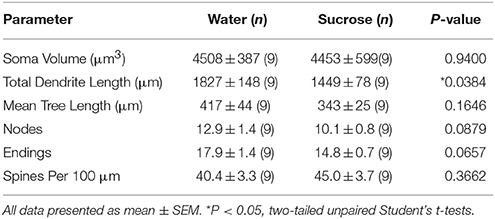
Tabulka 2. Obecné morfologické parametry středně ostnatých neuronů z jádra accumbens shellů dlouhodobých potkanů spotřebovávajících sacharózu a věkově sladěných kontrol.
Po charakterizaci obecné dendritické morfologie dlouhodobě spotřebovaných NAN skořápek konzumujících sacharózu jsme analyzovali dendritické arborizace a hustoty páteře s ohledem na jejich charakteristiky větvení. Naše komplexní hodnocení dendritických stromů kvantifikovalo počet dendritických segmentů na větev, střední délku dendritických segmentů na větev a střední hustotu páteře na větev pořadí NAN skořápek MSN kontroly vody a dlouhodobě potkany konzumující sacharózu. Souhrn údajů o větev a analýza je uveden v tabulce 3.
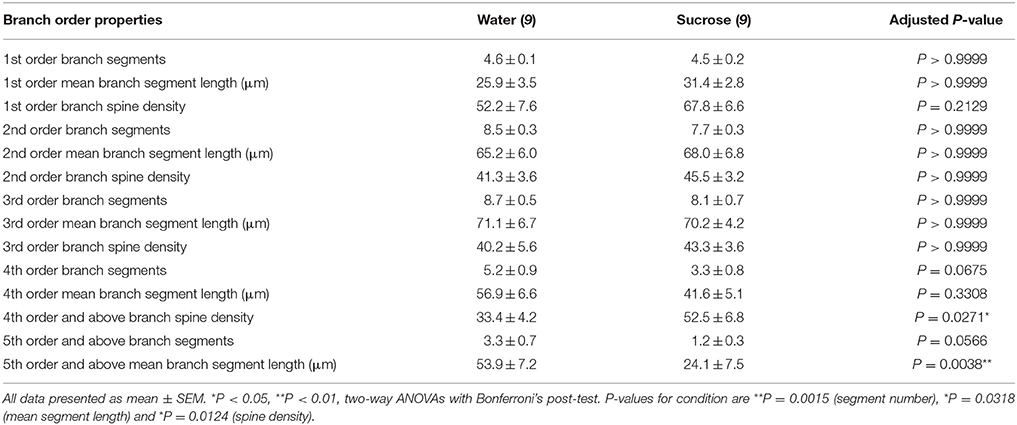 Tabulka 3. Charakteristiky větvích středně ostnatých neuronů u potkanů s dlouhodobou sacharózou a vodou.
Tabulka 3. Charakteristiky větvích středně ostnatých neuronů u potkanů s dlouhodobou sacharózou a vodou.Průměrné číslo segmentu dendritického větve na pořadí větví skořápek MSN NAc bylo významně sníženo u potkanů s dlouhodobou spotřebou sacharózy ve srovnání s kontrolami s vodou (**)P = 0.0015, obousměrná ANOVA). Bonferroni následné testy odhalily trend ke snížení počtu větvených segmentů v 4th (Voda: 5.2 ± 0.9, n = 9; Sacharóza 3.3 ± 0.8, n = 9, P = 0.0675, obrázek 2F, Stůl 3) a 5th a nad větvemi (Water: 3.3 ± 0.7, n = 9; Sacharóza 1.2 ± 0.3, n = 9, P = 0.0566, obrázek 2F, Stůl 3). Průměrná délka dendritického segmentu na pořadí větví skořápky MSN NAc byla také významně snížena u potkanů s dlouhodobou spotřebou sacharózy ve srovnání s kontrolami s vodou (*)P = 0.0444, obousměrná ANOVA). Bonferroni následné testy prokázaly snížení 55% u větví 5th a dále (voda: 53.9 ± 7.2 μm, n = 9; Sacharóza 24.1 ± 7.5 μm, n = 9, **P = 0.0038, obrázek 2G, Stůl 3).
Analýza větveného pořadí ukázala významné zvýšení hustoty dendritické páteře u MSN lastur NAc u potkanů konzumujících dlouhodobě sacharózu ve srovnání s kontrolami (*)P = 0.0124, obousměrná ANOVA). Bonferroni následné testy prokázaly nárůst hustoty páteře o 57% na distálních větvích 4th a dále (voda: 33.4 ± 4.2, n = 9; Sacharóza 52.5 ± 6.8, n = 9, P = 0.0271 *, vložení obrázků 2A, B, H, Stůl 3). Reprezentativní obrazy celkové architektury MSN a hustoty distální páteře (vsazení) jsou znázorněny na obrázcích 2A, B.
Dohromady tyto výsledky naznačují, že krátkodobá spotřeba sacharózy má malý vliv na morfologické parametry MSN ve skořápce NAc. Po dlouhodobé spotřebě však dochází k významnému snížení délky a složitosti neuronového trnu, zejména u distálních dendritických větví. Souběžné zvýšení hustoty distální páteře je také patrné u NAN skořápek MSN dlouhodobě potkanů konzumujících sacharózu.
Středně ostnaté neurony z jádra Nucleus Accumbens snížily složitost větvení po dlouhodobé, ale nikoli krátkodobé spotřebě sacharózy
Po krátkodobé spotřebě sacharosy nebyly žádné významné rozdíly v morfometrických parametrech NAc jádra MSN (tabulka 4). V analýzách týkajících se pořadí odstředivých větví nebyly také žádné významné rozdíly mezi 4-týdenní spotřebou sacharózy a MSN jádra pro regulaci vody. Konkrétně dendritické segmenty na pobočkovou objednávku (P = 0.7717), střední dendritická délka na větev (P = 0.2096) a průměrná hustota páteře na pořadí větví (P = 0.3521, dvousměrné ANOVA) se mezi skupinami nelišily.
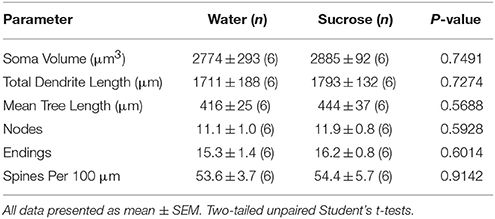 Tabulka 4. Obecné morfologické parametry středně ostnatých neuronů z jádra accumbens jádra potkanů s krátkodobou konzumací sacharózy a kontrolních vod odpovídajících věku.
Tabulka 4. Obecné morfologické parametry středně ostnatých neuronů z jádra accumbens jádra potkanů s krátkodobou konzumací sacharózy a kontrolních vod odpovídajících věku.Prodloužená spotřeba sacharózy také neměla významný morfometrický parametr jádra MSN NAc (tabulka 1) 5). Průměrné číslo segmentu dendritického větve na pořadí větví jádra MSN NAc bylo významně sníženo u potkanů s dlouhodobou spotřebou sacharózy ve srovnání s kontrolami s vodou (*)P = 0.0416, obousměrná ANOVA), nebyly však zaznamenány žádné významné rozdíly v průměrné dendritické délce na pořadí větví (P = 0.0995) a průměrná hustota páteře na pořadí větví (P = 0.4888, dvousměrné ANOVA) mezi MSN v jádru NAc u potkanů s dlouhodobou spotřebou sacharózy ve srovnání s kontrolami s vodou. Naše data dohromady ukazují, že jádro NAc není tak citlivé na dlouhodobou spotřebu sacharózy ve srovnání s MSN z oblasti skořápky NAc.
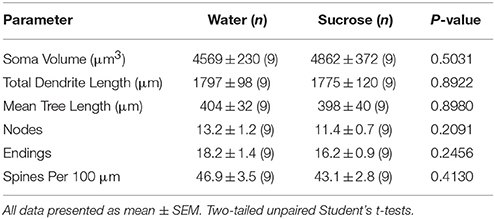 Tabulka 5. Obecné morfologické parametry středně ostnatých neuronů z jádra accumbens jádra dlouhodobě potkanů konzumujících sacharózu a věkově odpovídajících vodních kontrol.
Tabulka 5. Obecné morfologické parametry středně ostnatých neuronů z jádra accumbens jádra dlouhodobě potkanů konzumujících sacharózu a věkově odpovídajících vodních kontrol.Diskuse
Zvýšená dostupnost vysoce oslazených potravin v západní stravě nejenže přispěla ke zvýšené prevalenci a ekonomické zátěži spojené s obezitou a cukrovkou typu II, ale také vedla k nástupu poruch příjmu potravy, jako je přejídání (Swanson a kol., 2011; Kessler a kol., 2013; Davis, 2015). Přestože návykové vlastnosti cukrů včetně fruktózy a sacharózy zůstávají spekulativní, existuje výrazná podobnost v behaviorálních a nervových korelacích, které se projevují v důsledku nadměrného příjmu potravy a dlouhodobého užívání drog (Avena a kol., 2008, 2011). Cukr navíc aktivuje mozkové obvody odměn podobným způsobem jako zneužívané drogy (Volkow a kol., 2012) a výsledky studií na lidech naznačují, že cukr a sladkost mohou vyvolat touhu srovnatelnou s tou, kterou vyvolávají návykové drogy, jako je alkohol a nikotin. (Volkow a kol., 2012). Proto jsme použili model spotřeby binge-sacharózy u potkanů pro stanovení účinků krátkodobé (4 týdny) a dlouhodobé (12 týdny) spotřeby sacharózy na neuronální morfologii MSN v NAc, klíčové součásti překrývající se soustavy odměn to je modulováno cukrem a návykovými drogami. Ukazujeme, že MSN ze skořápky NAc u chronicky dlouhodobých potkanů konzumujících sacharózu výrazně snížily dendritickou délku a složitost, ale zvýšily hustotu distální dendritické páteře. Dlouhodobá spotřeba sacharózy neměla žádný vliv na morfologii MSN z jádra NAc, zatímco krátkodobá spotřeba sacharózy neměla významný vliv na morfologii MSN z jádra nebo obalu NAc. Tyto výsledky nejen prokazují přímý účinek prodlouženého binge-like sacharózy na neuronální morfologii NAN skořápek MSN, ale také zdůrazňují potenciálně škodlivé důsledky dlouhodobé konzumace diet s vysokým obsahem cukru.
NAc, který tvoří část ventrálního striata, je složen primárně z MSN, které jsou morfologicky charakterizovány jako středně velké neurony s rozsáhlými dendritickými arborizacemi a vysokou hustotou páteře (Kemp a Powell, 1971; Graveland a DiFiglia, 1985; Rafols a kol., 1989; Kawaguchi a kol., 1990). Glutamatergické a dopaminergní neurony jsou dva primární aferentní vstupy do NAc, primárně kontaktující dendritické hřídele a páteře MSN (Groves, 1980; Kaiya a Namba, 1981; Groves a kol., 1994). Konkrétně, NAc shell a jádro přijímají glutamatergický vstup z funkčně odlišných kortikálních oblastí (Brog a kol., 1993). Obal NAc je také inervován excitačními aferenty ze subkortikálních oblastí, jako je hippocampus, thalamus a basolaterální amygdala (Brog a kol., 1993; Wright a Groenewegen, 1995). Předchozí studie prokázaly, že tyto glutamatergické vstupy hrají klíčovou roli v motivaci a cílovém chování, jako je hledání potravy a odměn (Maldonado-Irizarry a kol., 1995; Kelley a Swanson, 1997; Reynolds a Berridge, 2003; Richard a Berridge, 2011). Dalším převládajícím vstupem do NAc MSN je od dopaminergních aferentů, které vyčnívají z ventrální tegmentální oblasti (Lindvall a Björklund, 1978; Veening a kol., 1980; Kalivas a Miller, 1984). Je zajímavé, že předchozí studie využívající podobné modely přerušovaného přístupu k cukru ukázaly, že výsledná spotřeba podobná binge vede k nárůstu extracelulárního dopaminu v NAc podobně (i když v menší míře) k zneužívání drog (Rada a kol., 2005; Avena a kol., 2006) a může modulovat expresi receptoru dopaminu (Colantuoni a kol., 2001, 2002) v jádru a shellu NAc. Zajímavé je, že bush-like konzumace sacharózy způsobuje eskalaci příjmu v čase podobně jako při samoléčení drog zneužívaných jako je kokain a heroin (Ahmed a Koob, 1998; Ahmed a kol., 2000, 2003), která je spojena s vývojem stavu „návykové“.
Naše analýza morfometrie větvích řádů ukazuje, že celkové snížení dendritické délky MSN skořápky NAc způsobené dlouhodobým příjmem sacharózy je primárně důsledkem snížení složitosti distálních větví. Pozorovali jsme zmenšené distální větvení (4th a 5th a nad větvemi) a významně sníženou střední délku v 5tém a nad dendrity v kombinaci se zvýšenou hustotou páteře v těchto větvích. Společným faktorem, který pravděpodobně ovlivní tento typ dendritické restrukturalizace, jsou změny synaptické konektivity a / nebo funkce (Russo a kol., 2010). Předchozí studie ukázaly, že glutamatergické synapsie na MSN se tvoří především na páteřích, zejména na distálních dendritech (Groenewegen a kol., 1999). Kromě toho, co-lokalizace dopaminu a glutamatergických vstupů z prefrontální kůry (Sesack a Pickel, 1992), hippocampus (Totterdell a Smith, 1989; Sesack a Pickel, 1990), a amygdala (Johnson a kol., 1994) byly pozorovány na dendritických páteřích MSN. Tato pozorování spojená se zvýšenou hustotou páteře po dlouhodobé konzumaci sacharózy pozorované v naší studii podporují tvorbu zvýšených excitačních vstupů. Proto existuje možnost, kdy by přetrvávající účinky způsobené prodlouženým binge-like sacharózou mohly usnadnit zvýšenou excitační synaptickou aktivitu na distálních dendritech MSN ve skořápce NAc. Následkem toho může prostřednictvím synaptického homeostatického mechanismu dojít ke zmenšení a / nebo zatažení distálních dendritů (Reissner a Kalivas, 2010), to však musí být stanoveno.
Je zajímavé poznamenat, že Crombag a jeho kolegové ukázali, že nedošlo ke zvýšení hustoty páteře v NAc shell po 4-týdenní spotřebě sacharózy prostřednictvím paradigmatu nose-poke self-Administration, a to navzdory robustnějšímu získání a vyšší míře odezvy na sacharózu ve srovnání s amfetaminem (Crombag a kol., 2005). Jejich pozorování nepřítomnosti změny hustoty páteře v 4 týdnech odráží naše zjištění. Naproti tomu naše studie ukazuje, že po dlouhodobé expozici chronické konzumaci sacharózy (týden 12) dochází k významnému zvýšení hustoty distální páteře na MSN potkanů trpících sacharózou. Naše laboratoř dále již dříve ukázala, že dlouhodobá konzumace sacharózy (12 týden) usnadňuje diferenciální farmakologickou odpověď na farmakoterapeutika, u nichž bylo prokázáno, že modulují odpovědi dopaminu a acetylcholinu na úrovni NAc (Shariff a kol., V tisku). Dohromady to naznačuje, že dlouhodobá (12 týdny a déle) expozice sacharosy, která přesněji odráží scénáře reálného světa, vede k úpravám morfologie na úrovni NAc.
Pokud jde o zneužívání drog, opakovaná expozice různým drogám způsobuje dlouhodobé změny ve struktuře dendritů a dendritických páteří. Například amfetaminy i kokain zvyšují hustotu páteře v NAc ve skořápce i jádru (Robinson a Kolb, 2004). Ukázalo se také, že expozice nikotinu zvyšuje hustotu páteře ve skořápce NAc. Naopak expozice morfinu vede ke snížení hustoty páteře a složitosti dendritických větví (Robinson a Kolb, 2004). Pokud jde o dlouhodobou spotřebu sacharózy, pozorovali jsme nárůst hustoty páteře podobný amfetaminu, kokainu a nikotinu a na rozdíl od účinku morfinu. Na rozdíl od amfetaminu a kokainu, ale podobného nikotinu, je však nárůst hustoty páteře při dlouhodobé expozici sacharóze omezen na skořápku NAc. Je také zajímavé, že změny v obou dendritických větvích (Robinson a Kolb, 1999) a hustota páteře (Li et al., 2003) produkované amfetaminem nebo kokainem jsou omezeny na distální dendrity MSN v NAc, což odráží zjištění v naší studii. Kromě toho a v souladu s výše uvedenými změnami bylo také ukázáno, že spotřeba sacharózy zvyšuje excitační synaptickou sílu na neurony dopaminu (Stuber a kol., 2008b) a další složky mezolimbické odměny (Stuber a kol., 2008a; Chen a kol., 2010). Dohromady to představuje sacharózu jako silný modulátor morfologie neuronů po dlouhodobém intenzivním užívání, což je podobné účinkům pozorovaným u drog zneužívání.
Ačkoliv jsou zapotřebí další výzkumy k odhalení buněčných a synaptických mechanismů přispívajících k morfologickým změnám pozorovaným v této studii, naše výsledky ukazují významné neuronální účinky vyvolané dlouhodobou konzumací sacharózy. Zejména není v naší studii zkoumáno, zda pozorované morfologické účinky sacharózy mohou být vyvolány také nekalorickými sladidly, jako je sacharin. V tomto ohledu je důležité poznamenat, že Lenoir a jeho kolegové prokázali, že intenzivní sladkost převyšuje odměnu za kokain, ať už je produkována sacharinem nebo sacharózou (Lenoir a kol., 2007). Navíc nedávná studie publikovaná naší laboratoří (Shariff a kol., V tisku) ukazuje, že vareniklin, parciální agonista nikotinového acetylcholinového receptoru, snižoval příjem sacharózy i sacharinu u hlodavců po stejném režimu dlouhodobého přerušovaného přístupu, jaký byl použit v této studii. Je zajímavé, že předchozí studie prokázaly podobnost akutních účinků nekalorických sladidel, jako je sacharin a sacharóza, na úrovni NAc (Scheggi a kol., 2013; Tukey a kol., 2013; Carelli a West, 2014). Další studie však musí stanovit, zda nekalorická sladidla mohou vyvolat dlouhodobé účinky podobné změnám v morfologii MSN skořápky NAc způsobené dlouhodobou konzumací sacharózy.
Nedostatek účinku na morfologii NAc MSN po krátkodobé konzumaci sacharózy zdůrazňuje význam provádění dlouhodobých studií pro posouzení dopadu dlouhodobého zneužívání drog nebo přírodních výhod, jako je sacharóza. Pokud jde o závislost, nejen opakované cykly příjmu a abstinence, klíčové složky závislostního cyklu, rostoucí množství důkazů odhalilo, že přechod na závislost je progresivní proces, ke kterému často dochází po delší časové období. Ačkoli návykové vlastnosti cukrů zůstávají nejisté, hodnověrnost závislosti na jiných nrogových přínosech, jako je sex, hazard a jídlo, se stále více zkoumá. Výsledky této studie přispívají k hypotéze, že cukry, jako je sacharóza, mají potenciálně návykové vlastnosti po dlouhodobé spotřebě podobné nárazům. Naše výsledky mají také dopad na rostoucí počet dětí a dospívajících, kteří udržují nezdravé stravovací návyky (vysoká spotřeba cukru a nadužívání) do dospělosti. V souladu se zvýšeným rizikem rozvoje metabolických účinků je také možné, že z tohoto chování mohou vyplývat také neurologické a psychiatrické důsledky ovlivňující náladu a motivaci.
Autorské příspěvky
Podílel se na výzkumu: PK, SB. Provedené experimenty: PK, MS, AB, MF, EM. Analýza dat: PK, MF, MS. Interpretoval data a přispěl k psaní rukopisu: PK, MS, MF, EM, MB, SB. Všichni autoři přečetli a schválili konečný rukopis k podání.
Prohlášení o konfliktu zájmů
Autoři prohlašují, že výzkum byl proveden bez obchodních či finančních vztahů, které by mohly být považovány za potenciální střet zájmů.
Recenzenti SC, SA a editor zpracování deklarovali své společné přidružení a editor zpracování uvádí, že tento proces přesto splnil standardy spravedlivého a objektivního přezkumu.
Poděkování
Tato práce byla podporována financováním z grantů Australské rady pro výzkum (FT1110884) SB a Národní rady pro zdraví a lékařský výzkum (1061979) SB a MB.
Doplňkový materiál
Doplňkový materiál k tomuto článku lze nalézt na adrese: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
Doplňkový obrázek 1. Příjem sacharózy a preference od krys konzumujících sacharózu 4 a 12 týdně. (A, B) ukazují eskalaci celkového příjmu sacharózy (ml) během týdnů expozice 4 a 12. (CD) vykazují vysokou preferenci pro sacharózu před vodou během období prezentace sacharózy.
Reference
Ahmed, SH, a Koob, GF (1998). Přechod z mírného na nadměrný příjem léčiv: změna hedonické nastavené hodnoty. Věda 282, 298-300. dva: 10.1126 / science.282.5387.298
Ahmed, SH, Lin, D., Koob, GF a Parsons, LH (2003). Eskalace samopodávání kokainu nezávisí na změněném jádru indukovaném kokainem accumbens hladině dopaminu. J. Neurochem. 86, 102-113. dva: 10.1046 / j.1471-4159.2003.01833.x
Ahmed, SH, Walker, JR, a Koob, GF (2000). Trvalé zvyšování motivace k užívání heroinu u krys s anamnézou eskalace drog. Neuropsychopharmacology 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
Arslanian, S. (2002). Diabetes typu 2 u dětí: klinické aspekty a rizikové faktory. Horm Res 57 (doplněk 1), 19 – 28. doi: 10.1159 / 000053308
Avena, NM, Bocarsly, ME, Hoebel, BG a Gold, MS (2011). Překrývání se v nosologii zneužívání návykových látek a přejídání se: translační důsledky „závislosti na potravě“. Měna. Zneužívání drog Rev. 4, 133-139. dva: 10.2174 / 1874473711104030133
Avena, NM, Rada, P. a Hoebel, BG (2008). Důkazy pro závislost na cukru: behaviorální a neurochemické účinky přerušovaného, nadměrného příjmu cukru. Neurosci. Biobehav. Rev. 32, 20-39. dva: 10.1016 / j.neubiorev.2007.04.019
Avena, NM, Rada, P., Moise, N., a Hoebel, BG (2006). Sacharóza falešného krmení podle časového harmonogramu uvolňuje opakovaně dopamin a vylučuje saturaci acetylcholinu. Neurovědy 139, 813-820. dva: 10.1016 / j.neuroscience.2005.12.037
Benton, D. (2010). Věrohodnost závislosti na cukru a její úloha při obezitě a poruchách příjmu potravy. Clin. Nutr. 29, 288 – 303. doi: 10.1016 / j.clnu.2009.12.001
Bray, GA a Popkin, BM (2014). Dietní cukr a tělesná hmotnost: dosáhli jsme v epidemii obezity a cukrovky krizi? Nalijte cukr. Diabetes Care 37, 950 – 956. doi: 10.2337 / dc13-2085
Brog, JS, Salyapongse, A., Deutch, AY a Zahm, DS (1993). Vzory aferentní inervace jádra a skořápky v „accumbens“ části ventrálního striata potkana: imunohistochemická detekce retrográdně transportovaného fluoro-zlata. J. Comp. Neurol. 338, 255 – 278. doi: 10.1002 / cne.903380209
Bucher Della Torre, S., Keller, A., Laure Depeyre, J., a Kruseman, M. (2015). Nápoje slazené cukrem a riziko obezity u dětí a dospívajících: systematická analýza toho, jak může metodická kvalita ovlivnit závěry. J. Acad. Nutr. Strava. [EPUB před tiskem]. doi: 10.1016 / j.jand.2015.05.020
Carelli, RM a West, EA (2014). Když se špatná chuť zlepší: nervové mechanismy, které jsou základem vzniku negativního vlivu a související devalvace přirozené odměny kokainem. Neurofarmakologie 76 (Pt B), 360 – 369. doi: 10.1016 / j.neuropharm.2013.04.025
Chen, BT, Hopf, FW a Bonci, A. (2010). Synaptická plasticita mezolimbického systému: terapeutické důsledky pro zneužívání návykových látek. Ann. NY Acad. Sci. 1187, 129-139. dva: 10.1111 / j.1749-6632.2009.05154.x
Colantuoni, C., Rada, P., McCarthy, J., Patten, C., Avena, NM, Chadeayne, A., a kol. (2002). Důkazy, že přerušovaný nadměrný příjem cukru způsobuje endogenní závislost na opioidech. Obes. Res. 10, 478 – 488. doi: 10.1038 / oby.2002.66
Colantuoni, C., Schwenker, J., McCarthy, J., Rada, P., Ladenheim, B., Cadet, JL, a kol. (2001). Nadměrný příjem cukru mění vazbu na dopaminové a mu-opioidní receptory v mozku. Neuroreport 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
Crombag, HS, Gorny, G., Li, Y., Kolb, B., a Robinson, TE (2005). Opačné účinky zkušenosti se samopodáváním amfetaminu na dendritické páteře ve střední a orbitální prefrontální kůře. Cereb. Kůra 15, 341-348. dva: 10.1093 / cercor / bhh136
Dabelea, D., Mayer-Davis, EJ, Saydah, S., Imperatore, G., Linder, B., Divers, J., a kol. (2014). Prevalence diabetu typu 1 a typu 2 u dětí a dospívajících od 2001 do 2009. JAMA 311, 1778 – 1786. doi: 10.1001 / jama.2014.3201
Davis, C. (2015). Epidemiologie a genetika poruchy příjmu potravy (BED). CNS Spectr. 20, 522-529. dva: 10.1017 / s1092852915000462
Ford, ES a Dietz, WH (2013). Trendy v příjmu energie mezi dospělými ve Spojených státech: zjištění z NHANES. Dopoledne. J. Clin. Nutr. 97, 848 – 853. doi: 10.3945 / ajcn.112.052662
Fryar, CD, Carroll, MD a Ogden, CL (2014). Prevalence nadváhy, obezity a extrémní obezity u dospělých: USA, 1960 – 1962 až 2011 – 2012. Atlanta, GA: CDC.
Graveland, GA, a DiFiglia, M. (1985). Frekvence a distribuce středně velkých neuronů s odsazenými jádry u primátů a hlodavců. Brain Res. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
Groenewegen, HJ, Wright, CI, Beijer, AV, a Voorn, P. (1999). Konvergence a segregace ventrálních striatálních vstupů a výstupů. Ann. NY Acad. Sci. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
Groves, PM (1980). Synaptické zakončení a jejich postsynaptické cíle v neostrii: synaptické specializace odhalené z analýzy sériových řezů. Proc. Natl. Acad. Sci. USA 77, 6926 – 6929. doi: 10.1073 / pnas.77.11.6926
Groves, PM, Linder, JC, a Young, SJ (1994). Dopaminergní axony značené 5-hydroxydopaminem: trojrozměrné rekonstrukce axonů, synapsí a postsynaptických cílů u potkana neostriatum. Neurovědy 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
Hu, FB (2013). Vyřešeno: existuje dostatek vědeckých důkazů, že snižující se konzumace nápojů slazených cukrem sníží výskyt obezity a nemocí souvisejících s obezitou. Obes. Rev. 14, 606 – 619. doi: 10.1111 / obr.12040
Johnson, LR, Aylward, RL, Hussain, Z., a Totterdell, S. (1994). Vstup z amygdaly do jádra krysy accumbens: její vztah k imunoreaktivitě tyrosinhydroxylázy a identifikovaných neuronů. Neurovědy 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
Kaiya, H. a Namba, M. (1981). Dva typy dopaminergních nervových terminálů v krysím neostrii. Ultrastrukturální studie. Neurosci. Lett. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
Kalivas, PW a Miller, JS (1984). Neurotensinové neurony ve ventrální tegmentální oblasti se promítají do mediálního jádra accumbens. Brain Res. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
Kawaguchi, Y., Wilson, CJ, a Emson, PC (1990). Projekční subtypy krysích neostriatálních matricových buněk odhalené intracelulární injekcí biocytinu. J. Neurosci. 10, 3421-3438.
Kelley, AE a Swanson, CJ (1997). Krmení vyvolané blokádou AMPA a kainátových receptorů uvnitř ventrálního striata: studie mikroinfúzního mapování. Behav. Brain Res. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
Kemp, JM a Powell, TP (1971). Synaptická organizace jádra caudate. Philos. Trans. R. Soc. Lond. B Biol. Sci. 262, 403 – 412. doi: 10.1098 / rstb.1971.0103
Kenny, PJ (2011). Mechanismy odměňování v obezitě: nové pohledy a budoucí směry. Neuron 69, 664-679. doi: 10.1016 / j.neuron.2011.02.016
Kessler, RC, Berglund, PA, Chiu, WT, Deitz, AC, Hudson, JI, Shahly, V., et al. (2013). Prevalence a korelace poruch příjmu potravy ve Světových zdravotnických organizacích Světové průzkumy duševního zdraví. Biol. Psychiatrie 73, 904-914. dva: 10.1016 / j.biopsych.2012.11.020
Klenowski, PM, Fogarty, MJ, Belmer, A., Noakes, PG, Bellingham, MC, a Bartlett, SE (2015). Strukturální a funkční charakterizace dendritických trnů a GABAergických synaptických vstupů na interneuronech a hlavních buňkách krysí bazolaterální amygdaly. J. Neurophysiol. 114, 942 – 957. doi: 10.1152 / jn.00824.2014
Lenoir, M., Serre, F., Cantin, L., a Ahmed, SH (2007). Intenzivní sladkost převyšuje odměnu za kokain. PLoS ONE 2: e698. dva: 10.1371 / journal.pone.0000698
Li, Y., Kolb, B., a Robinson, TE (2003). Místo přetrvávajících změn hustoty dendritických páteř vyvolaných amfetaminem na středních ostnatých neuronech v nucleus accumbens a caudate-putamen. Neuropsychopharmacology 28, 1082-1085. dva: 10.1038 / sj.npp.1300115
Lindvall, O. a Björklund, A. (1978). Anatomie dopaminergních neuronových systémů v mozku krysy. Adv. Biochem. Psychopharmacol. 19, 1-23.
Lutter, M. a Nestler, EJ (2009). Homeostatické a hedonické signály interagují při regulaci příjmu potravy. J. Nutr. 139, 629 – 632. doi: 10.3945 / jn.108.097618
Maldonado-Irizarry, CS, Swanson, CJ, a Kelley, AE (1995). Glutamátové receptory v jádře accumbens shell řídí chování při krmení prostřednictvím laterálního hypotalamu. J. Neurosci. 15, 6779-6788.
Malik, VS, Popkin, BM, Bray, GA, Després, JP a Hu, FB (2010). Nápoje slazené cukrem, obezita, diabetes mellitus typu 2 a riziko kardiovaskulárních chorob. Oběh 121, 1356 – 1364. doi: 10.1161 / CIRCULATIONAHA.109.876185
Ng, SW, Slining, MM a Popkin, BM (2012). Použití kalorických a nekalorických sladidel v USA spotřebitelských balených potravinách, 2005-2009. J. Acad. Nutr. Strava. 112, 1828 – 1834 e1821 – e1826. doi: 10.1016 / j.jand.2012.07.009
Nielsen, SJ, Siega-Riz, AM a Popkin, BM (2002). Trendy v příjmu energie v USA mezi 1977 a 1996: podobné posuny pozorované napříč věkovými skupinami. Obes. Res. 10, 370 – 378. doi: 10.1038 / oby.2002.51
Paxinos, G. a Watson, C. (2007). Krysa mozku ve stereotaxických souřadnicích. Amsterdam; Boston, MA: Academic Press / Elsevier.
Popkin, BM (2010). Co je špatného na přístupu USA k obezitě? Virtuální mentor 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
Rada, P., Avena, NM, a Hoebel, BG (2005). Každodenní flákání cukru opakovaně uvolňuje dopamin ve skořápce accumbens. Neurovědy 134, 737-744. dva: 10.1016 / j.neuroscience.2005.04.043
Rafols, JA, Cheng, HW a McNeill, TH (1989). Golgiho studie myšího striata: dendritické změny související s věkem v různých populacích neuronů. J. Comp. Neurol. 279, 212 – 227. doi: 10.1002 / cne.902790205
Ranjan, A., a Mallick, BN (2010). Upravená metoda pro konzistentní a spolehlivé barvení Golgi-coxu ve významně zkrácené době. Přední. Neurol. 1: 157. doi: 10.3389 / fneur.2010.00157
Reinehr, T. (2013). Typ 2 diabetes mellitus u dětí a dospívajících. World J. Diabetes 4, 270 – 281. doi: 10.4239 / wjd.v4.i6.270
Reissner, KJ a Kalivas, PW (2010). Použití glutamátové homeostázy jako cíle pro léčbu návykových poruch. Behav. Pharmacol. 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
Reynolds, SM a Berridge, KC (2003). Glutamátové motivační soubory v nucleus accumbens: rostrocaudální skořápkové gradienty strachu a krmení. Eur. J. Neurosci. 17, 2187-2200. dva: 10.1046 / j.1460-9568.2003.02642.x
Richard, JM a Berridge, KC (2011). Nucleus accumbens interakce mezi dopaminem a glutamátem přepínají režimy a vytvářejí touhu versus strach: D (1) sám pro chuť k jídlu, ale D (1) a D (2) společně ze strachu. J. Neurosci. 31, 12866-12879. dva: 10.1523 / JNEUROSCI.1339-11.2011
Robinson, TE, a Kolb, B. (1999). Změny v morfologii dendritů a dendritických páteří v nucleus accumbens a prefrontální kůře po opakovaném ošetření amfetaminem nebo kokainem. Eur. J. Neurosci. 11, 1598-1604. dva: 10.1046 / j.1460-9568.1999.00576.x
Robinson, TE, a Kolb, B. (2004). Strukturální plasticita spojená s vystavením zneužívání drog. Neurofarmakologie 47 (doplněk 1), 33 – 46. doi: 10.1016 / j.neuropharm.2004.06.025
Russo, SJ, Dietz, DM, Dumitriu, D., Morrison, JH, Malenka, RC a Nestler, EJ (2010). Závislost synapse: mechanismy synaptické a strukturální plasticity v nucleus accumbens. Trendy Neurosci. 33, 267 – 276. doi: 10.1016 / j.tins.2010.02.002
Rutledge, LT, Duncan, J., a Beatty, N. (1969). Studie kolagenů axonů pyramidálních buněk v intaktním a částečně izolovaném mozkovém kortexu dospělých. Brain Res. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper, CB, Chou, TC a Elmquist, JK (2002). Nutnost krmení: homeostatická a hedonická kontrola stravování. Neuron 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
Scheggi, S., Secci, ME, Marchese, G., De Montis, MG, a Gambarana, C. (2013). Vliv chutnosti na motivaci k provozování kalorických a nekalorických potravin u potkanů bez příjmu potravy a bez příjmu potravy. Neurovědy 236, 320-331. dva: 10.1016 / j.neuroscience.2013.01.027
Sesack, SR a Pickel, VM (1990). V mediánovém jádru potkana se hippocampální a katecholaminergní terminály sbíhají na ostnatých neuronech a navzájem se přibližují. Brain Res. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
Sesack, SR a Pickel, VM (1992). Prefrontální kortikální efektory v synapsích potkanů na neznačených neuronálních cílech katecholaminových terminálů v jádře accumbens septi a na dopaminových neuronech ve ventrální tegmentální oblasti. J. Comp. Neurol. 320, 145 – 160. doi: 10.1002 / cne.903200202
Shariff, M., Quik, M., Holgate, JY, Morgan, M., Patkar, OL, Tam, V., a kol. (v tisku). Neuronální modulátory nikotinového acetylcholinového receptoru snižují příjem cukru. PLoS ONE.
Sheehan, DV a Herman, BK (2015). Psychologické a lékařské faktory spojené s neléčenou poruchou příjmu potravy. Prim. Care Companion CNS Disord. 17. doi: 10.4088 / PCC.14r01732
Simms, JA, Steensland, P., Medina, B., Abernathy, KE, Chandler, LJ, Wise, R., et al. (2008). Přerušovaný přístup k 20% ethanolu indukuje vysokou spotřebu etanolu u potkanů Long-Evans a Wistar. Alkohol. Clin. Exp. Res. 32, 1816-1823. dva: 10.1111 / j.1530-0277.2008.00753.x
Steensland, P., Simms, JA, Holgate, J., Richards, JK, a Bartlett, SE (2007). Vareniklin, parciální agonista nikotinového acetylcholinového receptoru alfa4beta2, selektivně snižuje spotřebu a hledání ethanolu. Proc. Natl. Acad. Sci. USA 104, 12518 – 12523. doi: 10.1073 / pnas.0705368104
Stuber, GD, Hopf, FW, Hahn, J., Cho, SL, Guillory, A., a Bonci, A. (2008a). Dobrovolný příjem ethanolu zvyšuje excitační synaptickou sílu ve ventrální tegmentální oblasti. Alkohol. Clin. Exp. Res. 32, 1714-1720. dva: 10.1111 / j.1530-0277.2008.00749.x
Stuber, GD, Klanker, M., de Ridder, B., Bowers, MS, Joosten, RN, Feenstra, MG, a kol. (2008b). Odměny prediktivní zvyšují excitační synaptickou sílu na dopaminové neurony midbrainu. Věda 321, 1690-1692. dva: 10.1126 / science.1160873
Swanson, SA, Crow, SJ, Le Grange, D., Swendsen, J., a Merikangas, KR (2011). Prevalence a korelace poruch příjmu potravy u dospívajících. Výsledky z replikace adolescentů z národního průzkumu komorbidity. Oblouk. Gen. Psychiatrie 68, 714-723. dva: 10.1001 / archgenpsychiatry.2011.22
Te Morenga, L., Mallard, S., a Mann, J. (2013). Dietní cukry a tělesná hmotnost: systematický přehled a metaanalýzy randomizovaných kontrolovaných studií a kohortových studií. BMJ 346: e7492. doi: 10.1136 / bmj.e7492
Totterdell, S. a Smith, AD (1989). Konvergence hippocampálního a dopaminergního vstupu na identifikované neurony v jádru accumbens krysy. J. Chem. Neuroanat. 2, 285-298.
Tukey, DS, Ferreira, JM, Antoine, SO, D'Amour, JA, Ninan, I., Cabeza de Vaca, S. a kol. (2013). Požití sacharózy indukuje rychlé obchodování s AMPA receptory. J. Neurosci. 33, 6123-6132. dva: 10.1523 / JNEUROSCI.4806-12.2013
Veening, JG, Cornelissen, FM a Lieven, PA (1980). Aktuální organizace aferentů k caudatoputamenům krysy. Studie křenové peroxidázy. Neurovědy 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
Ventura, T., Santander, J., Torres, R., a Contreras, AM (2014). Neurobiologické základy touhy po uhlohydrátech. Výživa 30, 252 – 256. doi: 10.1016 / j.nut.2013.06.010
Volkow, ND, Wang, GJ, Fowler, JS, Tomasi, D., a Baler, R. (2012). Odměna za jídlo a drogy: překrývající se okruhy lidské obezity a závislosti. Curr. Horní. Behav. Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
Wise, RA (1973). Dobrovolný příjem ethanolu u potkanů po expozici ethanolu v různých režimech. Psychopharmacologia 29, 203 – 210. doi: 10.1007 / BF00414034
Wright, CI a Groenewegen, HJ (1995). Vzory konvergence a segregace ve středním jádru accumbens krysy: vztahy prefrontálních kortikálních, středních talamických a bazálních amygdaloidních aferentů. J. Comp. Neurol. 361, 383 – 403. doi: 10.1002 / cne.903610304
Klíčová slova: binge-like konzum, dlouhodobý, středně ostnatý neuron, nucleus accumbens, sacharóza
Citace: Klenowski PM, Shariff MR, Belmer A, Fogarty MJ, Mu EWH, Bellingham MC a Bartlett SE (2016) Prodloužená spotřeba sacharózy v Binge-like způsobu, mění morfologii středních ostnatých neuronů v Nucleus Accumbens Shell. Přední. Behav. Neurosci. 10: 54. dva: 10.3389 / fnbeh.2016.00054
Přijato: 03 prosinec 2015; Přijato: 07 March 2016;
Publikováno: 23 březen 2016.
Úprava:
Djoher Nora Abrous, Institut des Neurosciences de Bordeaux, Francie
Recenze:
Serge H. Ahmed, Centre National de la Recherche Scientifique, Francie
Stéphanie Caille, Centre National de la Recherche Scientifique, Francie
Copyright © 2016 Klenowski, Shariff, Belmer, Fogarty, Mu, Bellingham a Bartlett. Toto je článek s otevřeným přístupem distribuovaný podle podmínek Creative Commons Attribution License (CC BY). Používání, distribuce nebo reprodukce na jiných fórech je přípustné za předpokladu, že jsou připočítáni původní autoři nebo nositelé licence a že je uvedena původní publikace v tomto časopise v souladu s uznávanou akademickou praxí. Není povoleno použití, distribuce nebo reprodukce, která nesplňuje tyto podmínky.
* Korespondence: Selena E. Bartlett, [chráněno e-mailem]