Přední Behav Neurosci. 2015 může 19; 9: 128. doi: 10.3389 / fnbeh.2015.00128. eCollection 2015.
Andreatta M1, Pauli P1.
Abstraktní
Při klasickém podmínění se zpočátku neutrální stimul (podmíněný stimul, CS) spojí s biologicky významnou událostí (nepodmíněný stimul, USA), což může být bolest (averzivní podmíněnost) nebo jídlo (apetitivní podmíněnost). Po několika asociacích je CS schopen zahájit obranné nebo dohonivé reakce. Na rozdíl od averzivní kondice je chuťová podmíněnost u lidí zkoumána jen zřídka, i když její význam pro normální a patologické chování (např. Obezita, závislost) je nepopiratelný. Záměrem této studie je přeložit nálezy zvířat o apetitivních úpravách na lidi, kteří používají potraviny jako USA. Třicet tři účastníků bylo vyšetřováno mezi 8 a 10 hodinou ranní bez snídaně, aby bylo zajištěno, že pociťují hlad. Během dvou fází akvizice jeden geometrický tvar (avCS +) předpovídal averzní USA (bolestivý elektrický šok), další tvar (appCS +) předpovídal apetitivní USA (čokoláda nebo slaný preclík podle preference účastníků) a třetí tvar (CS– ) nepředpovídal ani USA. Ve fázi vyhynutí byly tyto tři tvary plus nový tvar (NOVÝ) znovu představeny bez doručení USA. Hodnocení valence a vzrušení, jakož i odezvy polekanosti a vodivosti kůže (SCR) byly shromážděny jako indexy učení. Našli jsme úspěšnou averzivní a chutnou úpravu. Na jedné straně byl avCS + hodnocen jako negativnější a více vzrušující než CS– a vyvolal potlačení úleku a zvýšenou SCR. Na druhé straně byl appCS + hodnocen pozitivněji než CS– a vyvolal útlum úleku a větší SCR. Stručně řečeno, úspěšně jsme potvrdili nálezy na zvířatech u (hladových) lidí tím, že jsme demonstrovali chutné učení a normální averzní učení.
Úvod
Předvídání ohrožení a jídla má zásadní význam pro přežití jakéhokoli organismu. V klasickém kondicionování (Pavlov, 1927), narážka předchází averzivní události, jako je mírný bolestivý elektrický šok (averzní nepodmíněný stimul, USA), nebo apetitivní událost, jako je potravinová peleta (appetitive US), několikrát. Poté je tento samotný stimul (nyní označený CS, CS +) schopen vyvolat buď obranné, nebo konzumní reakce. První druh asociativního učení se nazývá averzivní kondicionování, zatímco druhý se nazývá apetitivní kondicionování.
Ačkoli predikce apetitivních událostí je pro přežití stejně důležitá jako predikce averzivních událostí, apetitivní kondice je pozoruhodně méně zkoumána u zvířat (Bouton a Peck, 1989; Koch a kol., 1996; McDannald a kol., 2011, pro přehled viz Martin-Soelch a kol., 2007) a také u lidí (Klucken a kol., 2009, 2013; Austin a Duka, 2010; Delgado a kol., 2011; Levy a Glimcher, 2011). Tento nedostatek výzkumu může být pravděpodobně způsoben složitostí chutného paradigmatu ve srovnání s averzivním. Například jídlo jako primární posilovač musí být dodáno, když je organismus hladový, aby byl prospěšný (poslední přehled viz Dickinson a Balleine, 1994; Clark a kol., 2012). V lidském výzkumu byl tento problém překonán pomocí peněz (Austin a Duka, 2010; Delgado a kol., 2011; Levy a Glimcher, 2011) nebo erotické obrázky (Klucken a kol., 2009, 2013). Studie neuro-zobrazování však poukázaly na to, že primární (tj. Občerstvení nebo nápoje) a sekundární (tj. Peníze) zesilovače aktivují některé běžné oblasti mozku (např. Striatum), ale také znamenají odlišné vzorce aktivace (Delgado a kol., 2011; Levy a Glimcher, 2011).
Pokud je nám známo, pouze několik studií kondicionování u lidí zkoumalo účinky primárních látek zvyšujících chuť k jídlu, jako je zápach (Gottfried et al., 2002), voda (Kumar a kol., 2008) nebo jídlo (Prévost a kol., 2012). V první studii Gottfried et al. (2002) přidružené neutrální obličeje (CS) s nepříjemným, příjemným nebo neutrálním zápachem. Zajímavé je, že našli větší aktivaci v orbitofrontální kůře (OFC) a ve ventrálním striatu v reakci na chutný CS + ve srovnání s averzivním CS +. Došli k závěru, že OFC zpracovává hodnotu zápachu a podílí se na přenosu afektivní hodnoty z čichové (US) do vizuální (CS) soustavy. (Ventro) striatální aktivace byla interpretována tak, že odráží chutný CR vyvolaný chutný CS +. Ve druhé studii Kumar a kol. (2008) pozvali své účastníky (zdravé kontroly a pacienty s velkou depresí) do laboratoře brzy ráno a požádali je, aby se zdrželi pití během noci, aby se ujistili, že před skenováním žízneli. Fraktální obrázky byly CS a 0.1 ml vody v USA. Je zajímavé, že kromě jiných aktivací vykazovali zdraví účastníci (ale nikoli pacienti s depresí) větší aktivaci ve ventrálním striatu na apetitivní CS +, což naznačuje, že tento stimul byl zpracován jako odměňující. Ve třetí studii Prévost a kol. (2012) také představil fraktální obrázky jako CS a buď sladké nebo slané občerstvení jako USA podle preferencí účastníků. CS byl prezentován po dobu 6 s a během poslední sekundy byl navíc prezentován obraz jídla. Pokaždé, když se objevil obrázek jídla, experimentátor vložil kousek jídla do rukou účastníků, kteří měli dovoleno svačinu okamžitě konzumovat. Autoři bohužel nehlásili aktivaci mozku chutnému CS + během fáze klasické úpravy; pozorovali však menší srdeční zpomalení na odměněnou CS + ve srovnání s neodměněnou CS–, což naznačuje rozdílné kondiční účinky.
Nedostatek studií využívajících překvapivou odezvu jako index pro apetitivní kondici je překvapivý, zejména s ohledem na jeho široké použití v averzivním kondicionování. Úžasná reakce je předková a automatická obranná reakce na náhlé, neočekávané a silné averzní události (Koch, 1999). Tato obranná reakce je zprostředkována relativně jednoduchou neuronální cestou zahrnující kochleární kořenové neurony, jádro kaudálního pontinu retikulární formace (PnC) a spinální motoneurony (Fendt a Fanselow, 1999; Koch, 1999). Studie na zvířatech odhalily, že vyděšená potenciace závisí na projekcích z amygdaly na PnC (Fendt a Fanselow, 1999; Koch, 1999), zatímco útlum překvapení závisí na neporušeném jádru accumbens (NAcc, Koch, 1999). Taková modulace vyděšené odpovědi je užitečným implicitním měřítkem valence prezentovaných popředí. Potenciace tedy ukazuje negativní valenci, zatímco útlum ukazuje pozitivní valenci, aniž by byly výrazně ovlivněny kognitivními procesy (Hamm a Weike, 2005; Andreatta a kol., 2010). Pokud je nám známo, pouze jedna studie na zvířatech zkoumala chuť k jídlu měřením překvapivých odpovědí jako závislého opatření. Zajímavé je, že zkoumané krysy vykazovaly překvapující útlum na CS + (tj. Světlo) spojené s dodáním roztoku sacharózy. Pozoruhodně, takový útlum byl narušen u zvířat s lézemi NAcc, ale ne u zvířat s lézemi amygdaly, což naznačuje, že NAcc hraje specifickou roli při vyvolávání apetitivních CR a při tlumení překvapivých odpovědí (Koch a kol., 1996).
Současná studie je, podle našich znalostí, první, která přeložila tento apetitivní kondiční paradigma na člověka pomocí primárních zesilovačů jako USA, tj. Sladkých (čokoládové smarties)®) nebo slané (malé slané preclíky) a vyděšená modulace jako míra CR. Očekávali jsme, že aplikace appCS + spustí silné apetitivní CR, což se odráží ve vyděšeném útlumu, zvýšeném SCR a pozitivním valenčním hodnocení ve srovnání s ostatními podněty, tj. AvCS + a CS–.
Materiály a metody
Účastníci
Čtyřicet dva dobrovolníků přijalo účast na studiu a získalo kredity za kurz. Devět účastníků bylo z analýzy vyloučeno, dva kvůli technickým problémům, tři proto, že byli kódováni jako neodpovídající (průměrná amplituda vyděšení <5 μV), a čtyři, protože nevykazovali dostatek vyděšených odpovědí na podmínku (minimum = 2; pro podrobnosti viz Materiály a metody). Nakonec bylo do analýzy zahrnuto 33 účastníků (16 mužů; průměrný věk: 22.09 let, SD: 2.84; rozsah: 18 – 29 let). Čtyři účastníci nebyli rodnými Němci a šest z nich byli leváci. Jeden účastník si během experimentu neuvědomoval asociace CSs-USs (viz postup), ale rozhodli jsme se tohoto účastníka nevyloučit, protože jeho reakce byly normální a neovlivnily výsledky.
Materiály
Nepodmíněný Stimuli (USA)
Byly použity dva druhy USA. Jako averzní USA jsme na dominantní předloktí účastníků aplikovali mírně bolestivý elektrický šok. Elektrický šok byl dodán pomocí dvou elektrod o průměru 9 mm a rozteči 30 cm. Elektrický stimul sestával z impulzního stimulu s frekvencí 50 Hz a trvání 200 ms, generovaného proudovým stimulátorem (Digitimer DS7A, Digitimer Ltd, Welwyn Garden City, Velká Británie, 400 V, maximum 9.99 mA). Intenzita elektrického šoku byla stanovena individuálně pomocí postupu prahu popsaného výše (Andreatta a kol., 2010). Stručně řečeno, účastníci podstoupili dvě série vzestupných a sestupných intenzit v intervalech 0.5 mA. Museli hodnotit každý podnět na vizuální stupnici od 0 (vůbec necítí) až po 10 (opravdu intenzivní bolest), přičemž 4 byl kotvou prahu (jen znatelná bolest). Průměrná intenzita elektrického stimulu byla 2.12 mA (SD = 0.56) a byl hodnocen jako bolestivý (M = 6.45, SD = 1.73). Chutný USA se skládal buď z čokolády (Smarties®) nebo malý slaný preclík. Volba apetitivního USA závisela na individuálních preferencích účastníka, jak byly hlášeny během předběžného rozhovoru. Účastníci museli konkrétně hlásit, zda během snídaně obvykle jedí slané nebo sladké jídlo. Mohli si také svobodně vybrat, zda během samotného experimentu upřednostňují čokoládu nebo slaný preclík. Nakonec si 22 účastníků vybralo čokoládu a 11 malý slaný preclík.
Podmíněná stimulace (CS)
Geometrické tvary (modrý čtverec, žlutý kruh, zelený trojúhelník, červený šestiúhelník) s úhlopříčkou 8 cm byly prezentovány jako CS. Tvary byly prezentovány uprostřed černé počítačové obrazovky pro 8. Jeden tvar (avCS +) byl vždy spojen s averzivním USA (bolestivý elektrický šok), jeden tvar (appCS +) byl vždy spojen s chutný USA (čokoláda nebo slaný preclík), jeden tvar (CS–) nebyl nikdy spojen s averzivním nebo chutný USA, a čtvrtý tvar (NOVÝ) byl představen ve vyhynulé fázi, ale ne během akviziční fáze, aby byla zajištěna jeho neutralita.
Počáteční sonda
Jako sdělovací sonda byl použit bílý šum 103 dB s dobou trvání 50 ms. Akustické podněty byly prezentovány binaurálně přes sluchátka a vyskytovaly se náhodně 4–6 s po nástupu tvaru.
Dotazníky
Před a po experimentu museli účastníci vyplnit německé verze soupisu stavu úzkosti státu (STAI, Laux a kol., 1981) a plán pozitivních negativních dopadů (PANAS, Krohne a kol., 1996). STAI je soupis k posouzení zvláštnosti účastníků a / nebo stavové úzkosti a skládá se z 20 položek jak pro zvláštnost, tak pro státní verzi. Úroveň úzkosti účastníků dříve (M = 37.06, SD = 7.80) a poté (M = 39.33, SD = 9.16) experiment se významně nezměnil [t(32) = 1.61, p = 0.117]. Skóre skóre úzkosti v aktuálním vzorku se pohybovalo mezi 20 a 58 (M = 36.6, SD = 8.98), což je srovnatelné s publikovaným normálním rozsahem dospělých (Laux a kol., 1981). PANAS (Krohne a kol., 1996) je index pro pozitivní a negativní náladu. Jedinci s vysokým skóre na škále pozitivních vlivů (PAS) jsou náchylní k emocím, jako je nadšení, zatímco jedinci s vysokým skóre na škále negativních vlivů (NAS), jsou náchylní k emocím, jako je úzkost. Každá položka se skládá z adjektiva a účastníci naznačují na stupnici od 1 (velmi mírně) do 5 (extrémně), do jaké míry adjektivum odráží jejich pocity v daném okamžiku. V důsledku experimentu nebyly zjištěny žádné významné rozdíly v negativním vlivu účastníků [začátek: M = 11.67, SD = 2.29; konec: M = 12.88, SD = 4.69; t(32) = 1.55, p = 0.130]. Účastníci nějak od začátku výrazně změnili svou pozitivní náladu (M = 26.72, SD = 4.70) do konce (M = 23.66, SD = 7.02) experimentu [t(31) = 3.11, p = 0.004]. Toto snížení pozitivní nálady účastníků mohlo souviset s nepříjemností paradigmatu (byly představeny bolestivé elektrické šoky i averzivní bílý šum).
Postup
Po příjezdu do laboratoře si účastníci přečetli a podepsali formulář informovaného souhlasu schválený etickou komisí Katedry psychologie University of Würzburg. Nebyli informováni o nepředvídaných událostech mezi CS a USA. Po vyplnění dotazníků byly připojeny elektrody a byl proveden postup prahu bolesti, jak je popsáno výše.
Během návyková fáze, čtyři geometrické tvary byly prezentovány dvakrát s inter-trial intervalem (ITI) měnícím se mezi 18 a 25 s (průměr: 21.5 s). Během této fáze nebyly dodány žádné americké nebo překvapivé sondy.
Před akviziční fází bylo každých 7 – 15 vysláno sedm záchvatů bílého šumu, aby se snížila počáteční překvapivá reaktivita.
Následující dva fáze akvizice byly identické (obr 1). Každá akviziční fáze spočívala v pokusech 24: 8 CS–, 8 avCS + a 8 appCS +. CS sekvence byla pseudonáhodná, s jediným omezením, že stejný stimul nemohl být prezentován více než dvakrát za sebou. Pozoruhodně, avCS + byl představen spolu s bleskem jako symbol pro elektrický šok a bolestivé USA byly dodány na ofset. AppCS + byl představen ve směsi s obrázkem Smarties nebo soleného preclíka a účastník si mohl vybrat Smarties nebo preclík z jar. CS - byl představen spolu se zákazovým symbolem a žádné USA nebyly dodány. Během tří prezentací 8 CS každého typu byla mezi nástupy 4 a 6 podána překvapivá sonda po nástupu stimulu. Během ITI byly představeny další tři vyděšené sondy, aby se zajistila jejich nepředvídatelnost a snížilo se překvapení. ITI, skládající se z černé obrazovky, se pohyboval mezi 18 a 25 s průměrnou hodnotou 21.5.
OBRÁZEK 1
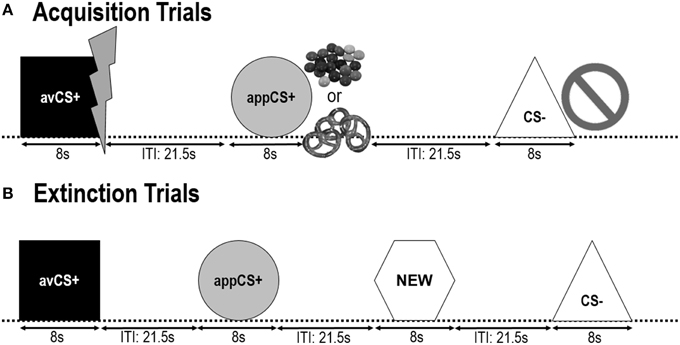
Obrázek 1. Zkoušky během dvou akvizičních fází (A) a extinkční fáze (B). Účastníci se dozvěděli, že jeden tvar (avCS +) předpovídal mírný bolestivý elektrický šok, jeden tvar (appCS +) předpovídal buď kousek čokolády nebo malý slaný preclík (podle jejich preferencí) a třetí tvar (CS–) nepředpovídal žádný biologicky významná událost. Každý tvar byl představen ve spojení s obrázkem znázorňujícím elektrický šok, smarties / slaný preclík nebo nic v závislosti na spojení s USA. Během vyhynulé fáze byly znovu představeny tři geometrické tvary, ale nebyly dodány USA. Kromě toho byl jako neutrální kontrola prezentován čtvrtý geometrický tvar (NOVÝ).
Během zániková fáze, účastníci znovu viděli tři geometrické tvary (tj. avCS +, appCS + a CS–) plus nový neutrální tvar (NOVÝ). Nebyly dodány žádné USA a tvary nebyly prezentovány ve spojení s obrázky záblesků, čokoládových / slaných preclíků nebo zákazů. Každý podnět byl prezentován osmkrát v pseudonáhodném pořadí (tj. Stejný podnět nebyl prezentován více než dvakrát za sebou), čímž se vytvořily studie 32. Podněty s překvapivou sondou byly prezentovány během 4 mimo prezentaci stimulu 8 každého typu CS. Stejně jako ve fázi akvizice a návyku se ITI lišily mezi 18 a 25 s, a 4 další vyděšené sondy byly během ITI dodávány nepředvídatelně.
Po každé fázi účastníci hodnotili valenci (příjemnost) a vzrušení (excitační) CS s vizuální analogovou stupnicí (VAS) v rozsahu od 1 do 9. Stupnice volnosti se pohybovala od „1“ označující „velmi nepříjemné“ po „9“ označující „velmi příjemné“; stupnice vzrušení se pohybovala od „1“ označující „klid“ po „9“ označující „vzrušující“. hodnoceno po dvou fázích akvizice a extinkci. Účastníci viděli geometrický tvar pro 1y a pak museli uvést, zda byl tento tvar spojen s elektrickým proudem, čokoládovým / slaným preclíkem, s ničím, nebo zda nebyli schopni spojit se. Zejména všichni účastníci (s výjimkou jednoho) si byli vědomi nepředvídané události po akvizici 2.
Redukce dat
Fyziologické odpovědi byly zaznamenány pomocí V-Amp 16 zesilovače a softwaru Vision Recorder V-Amp Edition (verze 1.03.0004, BrainProducts Inc., Mnichov, Německo). Byla použita vzorkovací frekvence 1000 Hz a 50 Hz vrubový filtr. Offline analýzy byly provedeny pomocí analyzátoru Brain Vision (verze 2.0; BrainProducts Inc., Mnichov, Německo).
Úžasná odpověď
Překvapivá odpověď byla měřena pomocí elektromyografie (EMG) vlevo orbicularis oculi sval s dvěma 5 mm Ag / AgCl elektrodami. V souladu s pokyny (Blumenthal a kol., 2005), jedna elektroda byla umístěna pod zornici a druhá 1 cm laterálně. Zemní a referenční elektrody byly umístěny na pravý a levý mastoid. Před připojením elektrod byla kůže lehce odřena a očištěna alkoholem, aby se impedance udržela pod 10 kΩ. Elektromyografický signál byl offline filtrován pomocí filtru 28 Hz s nízkým limitem a 500 Hz s filtrem s vysokým limitem. Poté byl signál EMG usměrněn a byl použit klouzavý průměr 50 ms. Jako základní linii jsme použili 50 ms před začátkem překvapivé sondy (Grillon a kol., 2006). Reakce na překvapivé sondy byly hodnoceny ručně a experimenty s nadměrnými posuny základní linie (± 5 μV) nebo artefakty pohybu byly z další analýzy vyloučeny. Úžasné reakce nižší než 5 μV byly kódovány jako nula a byly brány v úvahu pro výpočet velikosti vyděšené (Blumenthal a kol., 2005). Celkem bylo odmítnuto 10.4% pokusů a minimum 2 z 3 překvapujících odpovědí ve fázi akvizice a 4 mimo 8 překvapujících odpovědí ve fázi vymírání pro každou podmínku bylo vyžadováno, aby se účastník udržel v analytické skupině. Z tohoto důvodu byli vyloučeni čtyři účastníci. Amplituda píku byla definována jako maximální pík vztažený k základní linii během časového okna 20 – 120 ms po začátku překvapivé sondy. Nezpracovaná data byla poté normalizována v rámci jedinců pomocí z- hodnotí a poté T-skóre za účelem snížení vlivu individuální variability a lepšího odhalení psychologických procesů. Skóre T byla průměrována pro každou podmínku (avCS +, appCS +, CS–, NEW a ITI). Za účelem zkoumání vyděšené potenciace nebo úlevu od překvapení byla odečtena skóre pro vyděšené reakce ITI od vyděšených odpovědí pro každou podmínku.
Reakce na vodivost kůže (SCR)
Odpověď na vodivost kůže (SCR) byla zaznamenána za použití dvou elektrod 5 mm Ag / AgCl umístěných na dlani dominantní ruky. Galvanická odezva byla offline filtrována pomocí 1 Hz vysokého mezního filtru. SCR byl definován jako rozdíl (v μS) mezi počátkem reakce (1 – 3 s po nástupu stimulu) a vrcholem odpovědi (Tranel a Damasio, 1994; Delgado a kol., 2011). Zkoušky obsahující vyděšené sondy nebyly při analýze SCR brány v úvahu. Reakce pod 0.02 μS byly kódovány jako nula. Pět dalších účastníků bylo vyloučeno z analýzy SCR, protože měli průměrnou SCR nižší než 0.02 μS. Surová data vodivosti kůže byla transformována na druhou odmocninu s cílem normalizovat distribuci a skóre byla zprůměrována pro každou podmínku zvlášť pro dvě akviziční fáze (avCS +, appCS +, CS–) a extinkční fázi (avCS +, appCS +, CS– a NOVÁ) ).
Statistická analýza
Všechna data byla analyzována pomocí SPSS pro Windows (verze 20.0, SPSS Inc.). Pro fyziologické odpovědi byly vypočteny oddělené vícerozměrné analýzy rozptylu (ANOVA) pro dvě akviziční fáze a extinkční fázi. ANOVA pro fáze akvizice měla podnět (avCS +, appCS +, CS–) a fázi (akvizice 1, akvizice 2) jako faktory uvnitř subjektu. ANOVA pro extinkční fázi měla pouze podnět (avCS +, appCS +, CS–, NEW) jako faktor uvnitř subjektu. Hodnoty valence, vzrušení a kontingence byly analyzovány pomocí samostatných ANOVA obsahujících stimul faktorů uvnitř subjektu (avCS +, appCS +, CS– a NOVÝ) a fáze. Tento faktor měl čtyři úrovně pro valenční a vzrušení (T1: po návykové fázi, T2: po první akviziční fázi, T3: po druhé akviziční fázi, T4: po extinkční fázi), ale tři úrovně pro kontingenční hodnocení (T1: po první akviziční fáze, T2: po druhé akviziční fázi, T3: po extinkční fázi).
Hladina alfa (a) byla nastavena na 0.05 pro všechny analýzy. Velikost efektu je uvedena jako částečná η2.
výsledky
Hodnoty valence a vzrušení pro každou fázi jsou znázorněny na obrázku 2; překvapivé reakce a SCR jsou znázorněny na obrázku 3.
OBRÁZEK 2
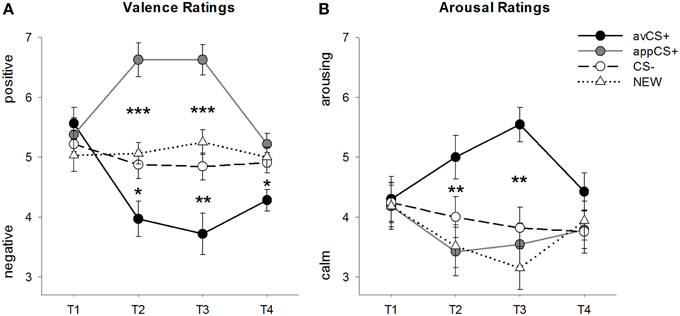
Obrázek 2. Hodnocení valence (A) a vzrušení (B). Řádky (se standardními chybami) zobrazují hodnocení po habituační fázi (T1), akvizici 1 (T2), akvizici 2 (T3) a extinkční fázi (T4). Averzní CS + (černá plná čára) získala zápornou valenci a vysoké vzrušení po dvou fázích akvizice ve srovnání s CS– (černá přerušovaná čára) a NOVINKA (černá tečkovaná čára). Důležité je, že apetitivní CS + (šedá plná čára) získala ve srovnání s CS– a NOVOU pozitivní valenci. *p <0.05, **p > 0.01, ***p <0.001.
OBRÁZEK 3
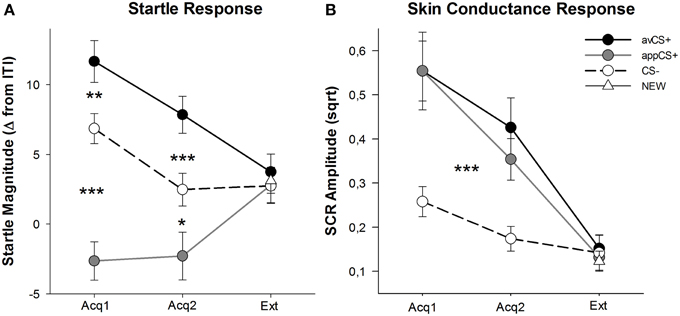
Obrázek 3. Překvapivé reakce (A) a vodivost kůže (B) (se standardními chybami) během první akviziční fáze (Acq1), druhé akviziční fáze (Acq2) a extinkční fáze (Ext). Překvapivé reakce byly během akvizičních fází významně potencovány k averzivní CS + (černá plná čára) a významně utlumeny k apetitivní CS + (šedá plná čára). SCR byla významně větší než avCS + a appCS + ve srovnání s CS–. Během zánikové fáze nebyly zjištěny žádné rozdíly. *p <0.05, **p > 0.01, ***p <0.001.
Hodnocení
ANOVA pro valenční hodnocení během akvizice odhalily významné hlavní účinky podnětu [F(3, 93) = 17.26, GG-ε = 0.801, p <0.001, částečné η2 = 0.358] a fáze [F(3, 93) = 3.30, GG-ε = 0.731, p = 0.039, částečný η2 = 0.096] a významná interakce mezi podnětem a fází [F(9, 279) = 9.54, GG-ε = 0.463, p <0.001, částečné η2 = 0.235]. Následovat t- testy naznačují, že valence čtyř geometrických tvarů na začátku experimentu byly totožné (ps> 0.19), zatímco po akvizici 1 i akvizici 2 byl avCS + hodnocen jako zvlášť negativní a appCS + jako zvláště pozitivní. Konkrétně byl avCS + hodnocen jako negativněji valencovaný ve srovnání s CS– [Acq1: t(31) = 2.34, p = 0.026; Acq2: t(31) = 3.07, p = 0.004], NOVÝ [Acq1: t(31) = 2.70, p = 0.011; Acq2: t(31) = 3.89, p <0.001] a appCS + [Acq1: t(31) = 5.41, p <0.001; Acq2: t(31) = 6.11, p <0.001]. AppCS + byl hodnocen jako výrazně pozitivnější než CS– [Acq1: t(31) = 4.99, p <0.001; Acq2: t(31) = 5.31, p <0.001] a NOVINKA [Acq1: t(31) = 4.92, p <0.001; Acq2: t(31) = 4.14, p <0.001]. Rozdíly mezi CS– a NOVINKOU nebyly nikdy významné (ps> 0.18).
Důležité bylo, že nebyly zjištěny žádné významné rozdíly při porovnání hodnot valence pro appCS + spojené s čokoládovým vs. slaným preclíkem [Acq1: t(30) = 0.03, p = 0.477; Acq2: t(30) = 0.29, p = 0.775].
Po extinkční fázi byl avCS + stále hodnocen jako negativnější ve srovnání s CS– [t(31) = 2.40, p = 0.023], NOVÝ [t(31) = 3.40, p = 0.002] a appCS + [t(31) = 3.35, p = 0.002]. Naopak valence appCS + se nelišila od CS– [t(31) = 1.77, p = 0.086] nebo NOVÝ [t(31) = 1.07, p = 0.293].
ANOVA pro hodnocení vzrušení během akvizice odhalil významný hlavní účinek podnětu [F(3, 96) = 7.07, GG-ε = 0.737, p = 0.001, částečný η2 = 0.181], ale ne fáze [F(3, 96) = 1.27, GG-ε = 0.805, p = 0.289, částečný η2 = 0.038] a významná interakce mezi podnětem a fází [F(9, 288) = 4.53, GG-ε = 0.582, p = 0.001, částečný η2 = 0.124]. Následovat t- testy neukázaly žádné významné rozdíly mezi podněty, pokud jde o jejich počáteční vzrušení (ps> 0.74). Po dvou fázích akvizice však byl avCS + hodnocen více vzrušujícím než NOVINKA [Acq1: t(32) = 2.99, p = 0.005; Acq2: t(32) = 5.97, p <0.001] a appCS + [Acq1: t(32) = 2.62, p = 0.013; Acq2: t(32) = 4.42, p <0.001] a o něco více vzrušující než CS– po první akviziční fázi [t(32) = 1.96, p = 0.058], ale po druhé akviziční fázi výrazně vzbuzující vzrušení [t(32) = 3.65, p = 0.001]. Na rozdíl od valenčních hodnocení se vzrušení appCS + nelišilo od vzrušení CS– a NOVÉ (ps > 0.13) po fázích akvizice.
Podobně jako u valenčních hodnocení se CS– a NEW v jejich vzrušení nelišily (ps> 0.07).
Stejně jako v případě valence nebyly zjištěny žádné rozdíly pro appCS + spojený s čokoládou a appCS + spojený se slaným preclíkem [Acq1: t(31) = 0.26, p = 0.797; Acq2: t(31) = 0.33, p = 0.724].
Po vyhynulé fázi nebyly odhaleny žádné významné rozdíly v hodnocení vzrušení (ps> 0.08).
Pro další post-hoc t-zkoušky porovnávající hodnocení po fázi návyku, první a druhé akviziční fázi a extinkční fázi, viz Dodatkové materiály.
Úžasná odpověď
ANOVA pro akviziční fáze vrátila hlavní účinek stimulu [F(2, 64) = 49.92, GG-ε = 0.964, p <0.001, částečné η2 = 0.609], ale ne fáze [F(1, 32) = 3.16, p = 0.085, částečný η2 = 0.090] a významná interakce Stimulus × Fáze [F(2, 64) = 3.37, GG-ε = 0.875, p = 0.048, částečný η2 = 0.095]. Následovat t- testy odhalily výraznou vyděšenou potenciaci avCS + ve srovnání s CS– během obou prvních [t(32) = 3.27, p = 0.003] a druhý [t(32) = 4.00, p <0.001] fáze akvizice. Vyděšené reakce na avCS + byly také významně potencovány ve srovnání s reakcemi na appCS +, opět po obou akvizicích 1 [t(32) = 8.20, p <0.001] a akvizice 2 [t(32) = 5.74, p <0.001]. Důležité je, že hodnota překvapení vůči appCS + byla významně oslabena ve srovnání s CS– během obou akvizic 1 [t(32) = 6.34, p <0.001] a akvizice 2 [t(32) = 2.91, p = 0.007]. Opět a v souladu s hodnocením nebyly u aplikace appCS + při spojení s čokoládou nebo slaným preclíkem odhaleny žádné rozdílné překvapivé reakce [Acq1: t(31) = 1.04, p = 0.309; Acq2: t(31) = 0.07, p = 0.947]. Během vyhynulé fáze nebyl zjištěn žádný významný účinek [F(3, 96) = 0.26, GG-ε = 0.906, p = 0.833, částečný η2 = 0.008].
Reakce na vodivost kůže (SCR)
Z ANOVA pro SCR během dvou akvizičních fází stimuluje hlavní účinky [F(2, 54) = 18.04, GG-ε = 0.908, p <0.001, částečné η2 = 0.401] a fáze [F(1, 27) = 20.91, p <0.001, částečné η2 = 0.436] se ukázalo být významné, ale ne jejich interakce [F(2, 54) = 0.68, GG-ε = 0.637, p = 0.451, částečný η2 = 0.024]. Post-hoc t- testy naznačily výrazně vyšší SCR než avCS + [t(27) = 6.46, p <0.001] a do appCS + [t(27) = 4.84, p <0.001] ve srovnání s CS–, zatímco účastníci vykazovali srovnatelnou SCR s avCS + a appCS + [t(27) = 0.64, p = 0.527]. Zejména nebyly zjištěny žádné rozdíly v SCR u čokoládové appCS + a slané preclíky appCS + [Acq1: t(26) = 2.55, p = 0.120; Acq2: t(26) = 1.29, p = 0.210]. Jak platilo pro hodnocení a překvapivou odezvu, nebyly zjištěny žádné významné účinky na fázi vymírání [F(3, 81) = 0.28, GG-ε = 0.634, p = 0.743, částečný η2 = 0.010].
Diskuse
Cílem této studie bylo převést nálezy zvířat na člověka pomocí klasického paradigmatu apetitivního kondicionování s primárním posilovačem jako nepodmíněným stimulem (USA). Aby to bylo možné, účastníci přišli do laboratoře brzy ráno bez snídaně, aby se ujistili, že měli hlad, a podle jejich preferencí byly jako chutné USA použity kousky čokolády nebo slané preclíky. Během akviziční fáze se jeden geometrický tvar (avCS +) spojil s mírně bolestivým elektrickým šokem (averzní USA), další tvar (appCS +) s apetitivními USA a třetí tvar (CS–) s apetitivními USA ani s averzní USA. Výsledky naznačují úspěšnou averzivní a apetivní kondici na explicitní slovní úrovni (tj. Hodnocení), na implicitní úrovni chování (tj. Na překvapivou odpověď) a na fyziologické úrovni (tj. SCR). Konkrétně, avCS + ve srovnání s CS– vyvolala negativní hodnocení valence, vyšší hodnocení vzrušení, vyděšený potencionál a větší SCR. Nejdůležitější je, že aplikace appCS + ve srovnání s CS– vyvolala pozitivnější valenční hodnocení, útlum vyděšení a větší SCR. Naše zjištění ohledně averzivního kondicionování byla v souladu s očekáváním, protože předchozí studie ukázaly, že hrozba předpovídající stimul (avCS +) je hodnocena jako averzivní, vyvolává větší reakce na strach a zvyšuje fyziologické vzrušení (Fendt a Fanselow, 1999; Hamm a Weike, 2005; Andreatta a kol., 2010, 2013). Naše výsledky týkající se kondicionování chuti k jídlu byly také v souladu s předchozími studiemi na lidech a zvířatech, které ukazují, že odměna předpovídající stimuly (appCS +) je hodnocena jako pozitivní, inhibuje reakce strachu a zvyšuje fyziologické vzrušení (Koch a kol., 1996; Gottfried et al., 2002; Kumar a kol., 2008; Klucken a kol., 2009, 2013; Austin a Duka, 2010; Prévost a kol., 2012). Pokud je nám známo, jedná se o první studii, která demonstruje podmíněné překvapivé útlum u lidí na stimul předpovídající primární odměnu. Důležité je, že jsme dokázali přenést a potvrdit výsledky studie na zvířatech (Koch a kol., 1996). Tato studie na zvířatech prokázala, že úleka od překvapení u potkanů závisí na projekcích z NAcc (část ventrálního striata) do PnC. Proto by překvapující útlum v naší studii mohl znamenat aktivitu NAcc, což je také v souladu s nálezy fMRI (Gottfried et al., 2002; Kumar a kol., 2008; Klucken a kol., 2009, 2013; Delgado a kol., 2011; Levy a Glimcher, 2011). Proto jsme dospěli k závěru, že naše chutný aparát kondicionování byl úspěšný, jak naznačují jak explicitní (hodnocení), tak implicitní (překvapující útlum) pozitivní valence.
Kromě tohoto nového, ale spíše předvídatelného zjištění si zaslouží zmínku dva další zajímavé výsledky. Nejprve se disociovaly verbální a fyziologické vzrušení na appCS +. Zadruhé jsme zjistili rychlejší zánik apetitivních CR ve srovnání s averzivními CR.
SCR indikuje sympatickou aktivaci, která je zvýšena na stimul spojený s odměnou (appCS +) i stimul spojený s hrozbou (avCS +). Tento výsledek je v souladu s předchozím studiem kondicionování, ve kterém byly erotické obrázky použity jako chutný USA (Klucken a kol., 2013) a studie odhalující srovnatelnou SCR s narážkou na předpovídání peněz a narážkou předpovídající averzní hluk (Austin a Duka, 2010). Zejména SCR je orientační odpověď související s aktivací sympatického systému. Tato reakce byla navržena tak, aby odrážela přípravu behaviorální reakce na motivačně významné události (Bradley, 2009). Na základě toho si myslíme, že jak stimuly spojené s odměnami, tak stimuly spojené s odměnami vyvolaly přípravnou odpověď na následné reakce na chování. Jinými slovy, signály potravy a bolesti zahájily přípravu na chování při přístupu a vyhýbání se. Na rozdíl od vysokého fyziologického vzrušení, slovní odpovědi naznačovaly nízké vzrušení k stimulům spojeným s odměnou. Je možné, že slovní vzrušení může být více ovlivněno vzbuzující povahou USA než fyziologickou aktivací samo o sobě. Ve skutečnosti si myslíme, že aplikace appCS + vs. avCS + byly hodnoceny jako nízko versus vysoce vzbuzující, protože byly spojeny s nízko a vysoce vzbuzujícími USA. Bohužel jsme neshromažďovali hodnocení vzrušení pro USA, a proto bude na budoucích studiích tuto hypotézu výslovně otestovat.
Během vyhaslé fáze nebyly dodány žádné USA. To mohlo vyvolat nové inhibiční učení zvané zánik (viz přehled viz Milad a Quirk, 2012), přičemž v důsledku toho klesá jak averzivní, tak i chutný CR. Je zřejmé, že na behaviorální (tj. Vyděšená odpověď) a na fyziologických (tj. SCR) úrovních už nebyly detekovatelné žádné rozdílné reakce na avCS +, appCS + a CS–. Podobně i explicitní vzrušení podmíněných podnětů hodnocených po extinkční fázi se vyrovnalo na nízké úrovni, což svědčí o úspěšném učení extinkce. AvCS + byl však stále hodnocen výrazně negativněji než CS–, zatímco valence appCS + se od valence CS nelišila. Pomalejší vymírání averzivní explicitní reakce může být způsobeno evolučním konzervatismem, což znamená, že na signály ohrožení je obzvláště těžké zapomenout, protože nereagování na signál ohrožení může ohrozit život.
Zbývá ještě jedna otázka: Proč potom vyděšená reakce (tj. Implicitní valence) úplně zhasla během vyhynulé fáze? Nejprve byly tyto odpovědi vypočteny během celého průběhu fáze. Je proto myslitelné, že diskriminační CR by byly stále detekovatelné během prvních pokusů extinkční fáze. Výzkumným způsobem jsme sledovali tuto hypotézu a zkoumali jsme jak vyděšené reakce, tak SCR během extinkční fáze (viz doplňkový materiál). Ačkoli jsme nenašli významné rozdíly, pozorovali jsme mírně vyšší vyděšenou velikost pro avCS + ve srovnání s CS– a novým kontrolním stimulem. Také jsme pozorovali o něco více vyděšený útlum appCS + ve srovnání s CS– a novým kontrolním stimulem na samém začátku extinkční fáze, která však během několika pokusů zmizela. SCR na appCS + klesla již po 2nd vyhynulém pokusu, zatímco SCR na avCS + zůstala vyšší téměř pro všechny vymírání ve srovnání s SCR na NOVOU. Ačkoli bychom tyto výsledky měli interpretovat s velkou opatrností, zdálo se, že překvapivé reakce paralelují s valenčním hodnocením. Překvapivé reakce i SCR dále podporují myšlenku evolučního konzervatismu vůči hrozebním podnětům.
Nakonec musíme uznat některá omezení této studie. Za prvé, kvůli technickému problému jsme nemohli ohlásit příjemnost (a vzrušení) čokolády a slaného preclíka. Chutný CR však naznačuje, že účastníci skutečně zažili oba USA jako chutný. Za druhé, doba averze a chutného USA se velmi lišila. Bolestivý elektrický šok byl tedy doručen na ofset avCS + a trval přesně 200 ms, zatímco čokoláda a slaný preclík byly účastníkům představeny přibližně 2 s po nástupu appCS + a trvání bylo nedefinovatelné, protože to záleželo na tom, jak rychle jednotlivec je snědl. Volba dodávat chutný američanům tímto způsobem byla založena na předchozí studii na lidech (Prévost a kol., 2012). Bylo by však metodologicky elegantnější dodávat chutný USA, který je srovnatelnější s averzivními USA, jako je například doušek šťávy nebo zázvorové pivo na appCS + offset. Abychom kompenzovali tento velký vnímavý rozdíl, představili jsme geometrické tvary ve spojení s vizuálním stimulem symbolizujícím USA. Zatřetí, nemůžeme definitivně vyloučit možnost, že rychle uhasené fyziologické reakce ve vyhynulé fázi jsou způsobeny metodologickým aspektem. Ve skutečnosti nebyly vizuální podněty v této fázi prezentovány ve spojení se symbolem USA, jako tomu bylo ve fázi akvizice. Je možné, že prezentace samotných CS mohla mít vliv na CR, a proto nebyly během prvních pokusů o vyhynutí patrné žádné významné rozdíly. Naše rychlé uhasené reakce jsou však v souladu s uhasenými odpověďmi v předchozí studii, ve které byl CS + (obličej) prezentován ve směsi s USA (křik) během akvizice, nikoli však při zániku (Lissek a kol., 2008).
Na závěr jsme zjistili úspěšné averzní a chutné podmíněné reakce na podnět spojený s hrozbou a na podnět spojený s odměnou. Je zajímavé, že explicitní (hodnocení) a implicitní (vyděšená reflex) úroveň odpovědí fungovala synergickým způsobem v tom, že avCS + byla hlášena jako negativní a indukovaná vyděšená potenciace a appCS + byla hlášena jako pozitivní a indukovaná úleva při vyděšení. Kromě toho, explicitní (hodnocení) a fyziologické (SCR) vzrušení appCS + disociované, odrážející dva odlišné procesy.
Prohlášení o konfliktu zájmů
Autoři prohlašují, že výzkum byl proveden bez obchodních či finančních vztahů, které by mohly být považovány za potenciální střet zájmů.
Poděkování
Tuto práci podpořilo Centrum pro spolupráci výzkumu „Strach, úzkost a úzkostné poruchy“, SFB-TRR 58, projekt B1.
Doplňkový materiál
Doplňkový materiál k tomuto článku lze nalézt na adrese: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
Reference
Andreatta, M., Mühlberger, A., Glotzbach-Schoon, E., a Pauli, P. (2013). Předvídatelnost bolesti zvrací valenční hodnocení stimulu spojeného s reliéfem. Přední. Syst. Neurosci. 7: 53. doi: 10.3389 / fnsys.2013.00053
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Andreatta, M., Mühlberger, A., Yarali, A., Gerber, B., a Pauli, P. (2010). Rozpor mezi implicitní a explicitně podmíněnou valencí po učení úlevy od bolesti u lidí. Proc. Biol. Sci. 277, 2411 – 2416. doi: 10.1098 / rspb.2010.0103
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Austin, AJ a Duka, T. (2010). Mechanismy pozornosti pro chutný a averzivní výsledek v Pavlovianově kondici. Behav. Brain Res. 213, 19-26. dva: 10.1016 / j.bbr.2010.04.019
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Blumenthal, TD, Cuthbert, BN, Filion, DL, Hackley, S., Lipp, OV a van Boxtel, A. (2005). Zpráva výboru: pokyny pro elektromyografické studie s překvapením očí u lidí. Psychofyziologie 42, 1-15. dva: 10.1111 / j.1469-8986.2005.00271.x
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Bouton, ME a Peck, CA (1989). Kontextové účinky na kondicionování, vyhynutí a obnovení v chutném kondicionačním přípravku. Anim. Učit se. Behav. 17, 188-198. dva: 10.3758 / BF03207634
CrossRef Plný text | Google Scholar
Bradley, MM (2009). Přirozená selektivní pozornost: orientace a emoce. Psychofyziologie 46, 1-11. dva: 10.1111 / j.1469-8986.2008.00702.x
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Clark, JJ, Hollon, NG a Phillips, PEM (2012). Pavlovianovy oceňovací systémy v učení a rozhodování. Curr. Opin. Neurobiol. 22, 1054 – 1061. doi: 10.1016 / j.conb.2012.06.004
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Delgado, MR, Jou, RL a Phelps, EA (2011). Neurální systémy, které jsou základem averzivní kondice u lidí s primárními a sekundárními posilovači. Přední. Neurosci. 5: 71. dva: 10.3389 / fnins.2011.00071
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Dickinson, A., a Balleine, B. (1994). Motivační kontrola cílené akce. Anim. Učit se. Behav. 22, 1-18. dva: 10.3758 / BF03199951
CrossRef Plný text | Google Scholar
Fendt, M., a Fanselow, MS (1999). Neuroanatomický a neurochemický základ podmíněného strachu. Neurosci. Biobehav. Rev. 23, 743 – 760. doi: 10.1016 / j.ijpsycho.2012.09.006
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Gottfried, JA, O'Doherty, J. a Dolan, RJ (2002). Chutné a averzní čichové učení u lidí studované pomocí funkčního zobrazování magnetickou rezonancí souvisejícího s událostmi. J. Neurosci. 22, 10829 – 10837. K dispozici online na adrese: http://www.jneurosci.org/content/22/24/10829.full
Grillon, C., Baas, JM, Cornwell, B., a Johnson, L. (2006). Kondicionování kontextu a vyhýbání se chování v prostředí virtuální reality: účinek předvídatelnosti. Biol. Psychiatrie 60, 752-759. dva: 10.1016 / j.biopsych.2006.03.072
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Hamm, AO a Weike, AI (2005). Neuropsychologie učení strachu a regulace strachu. Int. J. Psychophysiol. 57, 5 – 14. doi: 10.1016 / j.ijpsycho.2005.01.006
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Klucken, T., Schweckendiek, J., Merz, CJ, Tabbert, K., Walter, B., Kagerer, S., a kol. (2009). Neurální aktivace získávání podmíněného sexuálního vzrušení: účinky kontingenčního vědomí a sexu. J. Sex. Med. 6, 3071-3085. dva: 10.1111 / j.1743-6109.2009.01405.x
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Klucken, T., Wehrum, S., Schweckendiek, J., Merz, CJ, Hennig, J., Vaitl, D., a kol. (2013). Polymorfismus 5-HTTLPR je spojen se změněnými hemodynamickými odpověďmi během apetitivního kondicionování. Hučení. Brain Mapp. 34, 2549 – 2560. doi: 10.1002 / hbm.22085
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Koch, M. (1999). Neurobiologie vyplašení. Prog. Neurobiol. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Koch, M., Schmid, A., a Schnitzler, H.-U. (1996). Potěšení z úlevy vyplašení je narušeno lézemi nucleus accumbens. Neuroreport 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Krohne, HW, Egloff, B., Kohmann, C.-W., a Tausch, A. (1996). Untersuchungen mit einer deutschen version der “Pozitivní a negativní dopadový plán” (PANAS). Diagnostica 42, 139-156.
Kumar, P., Waiter, G., Ahearn, T., Milders, M., Reid, I., a Steele, JD (2008). Abnormální časové rozdíly odměn-učení signálů při velké depresi. Mozek 131, 2084-2093. dva: 10.1093 / mozek / awn136
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Laux, L., Glanzmann, P., Schaffner, P., a Spielberger, CD (1981). Das State-Trait Angstinventar. Weinheim: Beltzův test.
Levy, DJ a Glimcher, PW (2011). Porovnání jablek a pomerančů: použití subjektivní hodnotové reprezentace odměny specifické a odměněné v mozku. J. Neurosci. 31, 14693-14707. dva: 10.1523 / JNEUROSCI.2218-11.2011
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Lissek, S., Levenson, J., Biggs, AL, Johnson, LL, Ameli, R., Pine, DS, a kol. (2008). Zvýšená úprava strachu sociálně relevantním nepodmíněným podnětům při sociální úzkostné poruchě. Dopoledne. J. Psychiatry 165, 124-132. dva: 10.1176 / appi.ajp.2007.06091513
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Martin-Soelch, C., Linthicum, J., a Ernst, M. (2007). Chutná kondice: nervové základy a důsledky pro psychopatologii. Neurosci. Biobehav. Rev. 31, 426-440. dva: 10.1016 / j.neubiorev.2006.11.002
PubMed Abstrakt | CrossRef Plný text | Google Scholar
McDannald, MA, Lucantonio, F., Burke, KA, Niv, Y. a Schoenbaum, G. (2011). Ventrální striatum a orbitofrontální kůra jsou vyžadovány pro učení založené na modelu, ale ne bez modelu. J. Neurosci. 31, 2700-2705. dva: 10.1523 / JNEUROSCI.5499-10.2011
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Milad, MR a Quirk, G. (2012). Strach z vyhynutí jako model translační neurovědy: deset let pokroku. Annu. Rev. Psychol. 63, 129-111. dva: 10.1146 / annurev.psych.121208.131631
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Pavlov, IP (1927). Podmíněné reflexy: zkoumání fyziologické aktivity mozkové kůry. Londýn: Oxford University Press.
Prévost, C., Liljeholm, M., Tyszka, JM a O'Doherty, JP (2012). Neurální koreláty specifického a obecného přenosu pavlovianů na instrumenty v lidských podoblastech amygdalaru: studie fMRI s vysokým rozlišením. J. Neurosci. 32, 8383 – 8390. doi: 10.1523 / jneurosci.6237-11.2012
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Tranel, D., a Damasio, H. (1994). Neuroanatomické koreláty elektrodermálních kožních vodivých reakcí. Psychofyziologie 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
PubMed Abstrakt | CrossRef Plný text | Google Scholar
Klíčová slova: klasická kondice, odměna, trest, překvapivý reflex, kožní vodivost
Citace: Andreatta M a Pauli P (2015) Chuť proti averzní kondici u lidí. Přední. Behav. Neurosci. 9: 128. dva: 10.3389 / fnbeh.2015.00128
Přijato: 11 Únor 2015; Přijato: 05 Může 2015;
Publikováno: 19 Může 2015.
Úprava:
Niels Birbaumer, University of Tuebingen, Německo
Recenze:
Tim Klucken, Justus Liebig University Giessen, Německo
Alessandro Angrilli, Univerzita Padova, Itálie
Copyright © 2015 Andreatta a Pauli. Toto je článek s otevřeným přístupem distribuovaný podle podmínek Creative Commons Attribution License (CC BY). Používání, distribuce nebo reprodukce na jiných fórech je přípustné za předpokladu, že jsou připočítáni původní autoři nebo nositelé licence a že je uvedena původní publikace v tomto časopise v souladu s uznávanou akademickou praxí. Není povoleno použití, distribuce nebo reprodukce, která nesplňuje tyto podmínky.
* Korespondence: Marta Andreatta, Katedra psychologie (Biologická psychologie, Klinická psychologie a psychoterapie), Univerzita ve Würzburgu, Marcusstraße 9-11, D-97070 Würzburg, Německo, [chráněno e-mailem]
