Dev Rev. 2008; 28(1): 62-77. doi: 10.1016 / j.dr.2007.08.003
Abstract
Adolescentie is een ontwikkelingsperiode die wordt gekenmerkt door suboptimale beslissingen en acties die leiden tot een verhoogde incidentie van onopzettelijke verwondingen en geweld, alcohol- en drugsmisbruik, onbedoelde zwangerschap en seksueel overdraagbare aandoeningen. Traditionele neurobiologische en cognitieve verklaringen voor gedrag van adolescenten hebben geen verantwoording afgelegd voor de niet-lineaire veranderingen in gedrag waargenomen tijdens de adolescentie, ten opzichte van kindertijd en volwassenheid. Deze beoordeling biedt een biologisch plausibele beeldvorming van de neurale mechanismen die ten grondslag liggen aan deze niet-lineaire gedragsveranderingen, als een verhoogde gevoeligheid voor prikkels, terwijl impulscontrole nog relatief onrijp is gedurende deze periode. Recente humane beeldvorming en dierstudies vormen een biologische basis voor dit beeld, wat wijst op een differentiële ontwikkeling van limbische beloningssystemen ten opzichte van top-down controlesystemen tijdens de adolescentie in verhouding tot de kindertijd en de volwassenheid. Dit ontwikkelingspatroon kan verergeren bij adolescenten met een aanleg voor het nemen van risico's, waardoor het risico op slechte resultaten toeneemt.
Volgens het National Center for Health Statistics zijn er elk jaar meer dan 13,000 sterfgevallen door adolescenten in de Verenigde Staten. Ongeveer 70% van deze sterfgevallen is het gevolg van ongevallen met motorvoertuigen, onopzettelijke verwondingen, moord en zelfmoord (Eaton et al., 2006). Resultaten van de 2005 Nationale Jeugd Risico Gedragsonderzoek (YRBS) laten zien dat adolescenten gedrag vertonen dat hun kans op dood of ziekte vergroot door een voertuig te besturen na het drinken of zonder een veiligheidsgordel, het dragen van wapens, het gebruiken van illegale stoffen en het deelnemen aan onbeschermde seks. resulterend in onbedoelde zwangerschappen en SOA's, waaronder HIV-infectie (Eaton et al., 2006). Deze statistieken onderstrepen het belang van het begrijpen van risicovolle keuzes en acties bij adolescenten.
Een aantal cognitieve en neurobiologische hypothesen zijn gepostuleerd voor de reden waarom adolescenten zich bezighouden met suboptimaal keuzegedrag. In een recent overzicht van de literatuur over de ontwikkeling van de menselijke adolescente hersenen, Yurgelun-Todd (2007) suggereert dat cognitieve ontwikkeling gedurende de adolescente jaren geassocieerd is met een steeds grotere efficiëntie van cognitieve controle-capaciteiten. Deze efficiëntie wordt beschreven als afhankelijk van rijping van de prefrontale cortex, zoals blijkt uit verhoogde activiteit in focale prefrontale gebieden (Rubia et al., 2000; Tamm, Menon en Reiss, 2002) en verminderde activiteit in irrelevante hersengebieden (Brown et al., 2005; Durston et al., 2006).
Dit algemene patroon, van verbeterde cognitieve controle met rijping van de prefrontale cortex, suggereert een lineaire toename in ontwikkeling van kindertijd tot volwassenheid. Suboptimale keuzes en acties die tijdens de adolescentie zijn waargenomen, vertegenwoordigen echter een niet-lineaire gedragsverandering die kan worden onderscheiden van kindertijd en volwassenheid, zoals wordt aangetoond door het National Center for Health Statistics over gedrag en mortaliteit bij adolescenten. Als cognitieve controle en een onvolgroeide prefrontale cortex de basis waren voor suboptimaal keuzegedrag, dan zouden kinderen er opmerkelijk gelijk of zelfs slechter uit moeten zien dan adolescenten, gezien hun minder ontwikkelde prefrontale cortex en cognitieve vaardigheden. Onrijpe prefrontale functie alleen kan dus geen verklaring bieden voor het gedrag van adolescenten.
Een accurate conceptualisatie van cognitieve en neurobiologische veranderingen tijdens de adolescentie moet adolescentie behandelen als een overgangsperiode van ontwikkeling (Speer, 2000), in plaats van een enkele momentopname in de tijd (Casey, Tottenham, Liston en Durston, 2005). Met andere woorden, om deze ontwikkelingsperiode te begrijpen, zijn overgangen in en uit de adolescentie noodzakelijk om onderscheid te maken tussen verschillende eigenschappen van deze ontwikkelingsfase. Het ontwikkelen van ontwikkelingsroutes voor cognitieve en neurale processen is essentieel voor het karakteriseren van deze overgangen en beperkende interpretaties over gedragsveranderingen gedurende deze periode. Op cognitief of gedragsniveau worden adolescenten gekenmerkt als impulsief (dat wil zeggen, zonder cognitieve controle) en het nemen van risico's met deze constructies die als synoniemen en zonder waardering voor verschillende ontwikkelingsroutes van elk worden gebruikt. Op neurobiologisch niveau suggereren menselijke beeldvorming en dierproeven verschillende neurobiologische basissen en ontwikkelingsroutes voor de neurale systemen die ten grondslag liggen aan deze afzonderlijke constructies van impulsbeheersing en risicovolle beslissingen.
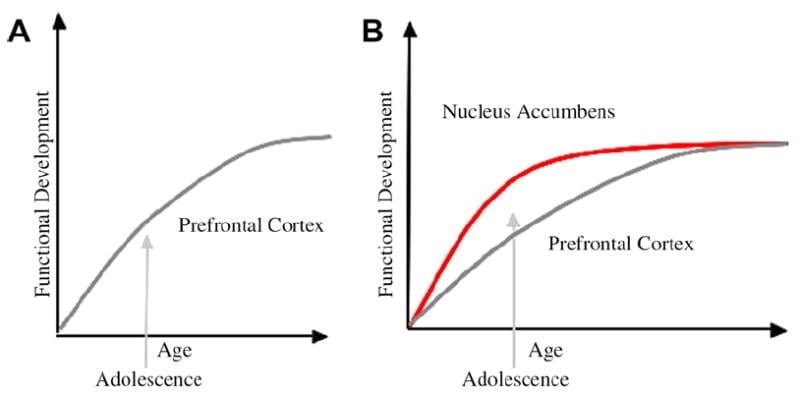
We hebben een neurobiologisch model ontwikkeld voor de ontwikkeling van adolescenten in dit kader dat voortbouwt op knaagdiermodellen (Laviola, Adriani, Terranova en Gerra, 1999; Speer, 2000) en recente beeldvormende onderzoeken naar adolescentie (Ernst et al., 2005; Galvan, Hare, Voss, Glover en Casey, 2007; Galvan et al., 2006). Fig 1 hieronder toont dit model. Aan de linkerkant is de traditionele karakterisering van de adolescentie bijna uitsluitend gerelateerd aan de onvolwassenheid van de prefrontale cortex. Aan de rechterkant is ons voorgestelde neurobiologische model dat illustreert hoe limbische subcorticale en prefrontale top-down controlegebieden samen moeten worden beschouwd. De cartoon illustreert verschillende ontwikkelingsroutes voor deze systemen, waarbij limbische systemen eerder ontwikkelen dan prefrontale controlegebieden. Volgens dit model is het individu meer bevooroordeeld door functioneel volwassen limbische regio's tijdens de adolescentie (dwz onbalans van limbisch ten opzichte van prefrontale controle) in vergelijking met kinderen, voor wie deze systemen (dat wil zeggen limbisch en prefrontaal) zich nog steeds aan het ontwikkelen zijn; en vergeleken met volwassenen, voor wie deze systemen volledig volgroeid zijn. Dit perspectief biedt een basis voor niet-lineaire verschuivingen in gedrag tijdens de ontwikkeling, als gevolg van eerdere rijping van dit limbische relatief ten opzichte van minder volwassen top-down prefrontale controleregio. Met ontwikkeling en ervaring biedt de functionele connectiviteit tussen deze regio's een mechanisme voor top-down controle van deze regio's (Hare, Voss, Glover, & Casey, 2007a). Verder verzoent het model de contradictie van gezondheidsstatistieken van risicovol gedrag tijdens de adolescentie, met de scherpzinnige observatie door Reyna en Farley (2006) dat adolescenten in staat zijn te redeneren en de risico's van gedrag te begrijpen waarin zij zich bezighouden. Volgens ons model zal het limbische systeem in emotioneel opzicht opvallende situaties controlesystemen overwinnen, gezien zijn volwassenheid ten opzichte van het prefrontale controlesysteem. Bewijsmateriaal uit gedrags- en humane beeldvormingsstudies ter ondersteuning van dit model wordt gegeven in de context van acties in belonende en emotionele contexten (Galvan et al., 2006, 2007; Hare, Voss, Glover, & Casey, 2007b; Hare et al., 2007a). Daarnaast speculeren we waarom de hersenen zich op deze manier kunnen ontwikkelen en waarom sommige tieners een groter risico lopen op het nemen van suboptimale beslissingen die leiden tot slechtere langetermijnresultaten (Galvan et al., 2007; Hare et al., 2007b).
De traditionele verklaring voor het gedrag van adolescenten is gesuggereerd als gevolg van de langdurige ontwikkeling van de prefrontale cortex (A). Ons model houdt rekening met de ontwikkeling van de prefrontale cortex samen met subcorticale limbische gebieden (bijv. Nucleus accumbens) die zijn betrokken bij risicovolle keuzes en acties (B).
Ontwikkeling van doelgericht gedrag
Een hoeksteen van cognitieve ontwikkeling is het vermogen om ongepaste gedachten en acties te onderdrukken in het voordeel van doelgerichte, vooral in de aanwezigheid van overtuigende prikkels (Casey, Galvan, & Hare, 2005; Casey et al., 2000b; Casey, Thomas, David-son, Kunz, & Franzen, 2002a; Casey, Tottenham en Fossella, 2002b). Een aantal klassieke ontwikkelingsstudies hebben aangetoond dat dit vermogen zich gedurende de kindertijd en adolescentie ontwikkelt (Case, 1972; Flavell, Feach en Chinsky, 1966; Keating & Bobbitt, 1978; Pascual-Leone, 1970). Verschillende theoretici hebben betoogd dat cognitieve ontwikkeling het gevolg is van een hogere verwerkingssnelheid en -efficiëntie en niet van een toename van de mentale capaciteit (bijv. Bjorkland, 1985; Bjorkland, 1987; Case, 1985). Andere theoretici hebben het construct van 'remmende' processen opgenomen in hun beschrijving van cognitieve ontwikkeling (Harnishfeger & Bjorkland, 1993). Volgens dit rapport wordt onrijpe cognitie gekenmerkt door gevoeligheid voor interferentie van concurrerende bronnen die onderdrukt moeten worden (bijv. Brainerd en Reyna, 1993; Casey, Thomas, Davidson, Kunz en Franzen, 2002a; Dempster, 1993; Diamond, 1985; Munakata & Yerys, 2001). Dus doelgericht gedrag vereist de beheersing van impulsen of uitstel van bevrediging voor het optimaliseren van resultaten en dit vermogen lijkt volwassen te worden in de kindertijd en de adolescentie.
Het gedrag van adolescenten is beschreven als impulsief en riskant, bijna synoniem, maar deze constructies vertrouwen op verschillende cognitieve en neurale processen, die verschillende constructies met verschillende ontwikkelingsroutes suggereren. Specifiek suggereert een literatuuroverzicht dat impulsiviteit afneemt met de leeftijd in de kindertijd en adolescentie (Casey et al., 2002a; Casey, Galvan et al., 2005; Galvan et al., 2007) en is geassocieerd met langdurige ontwikkeling van de prefrontale cortex (Casey, Galvan et al., 2005), hoewel er verschillen zijn in de mate waarin een bepaald individu impulsief is of niet, ongeacht de leeftijd.
In tegenstelling tot impuls / cognitieve controle lijkt het nemen van risico's te stijgen tijdens de adolescentie in verhouding tot de kindertijd en volwassenheid en wordt geassocieerd met subcorticale systemen waarvan bekend is dat ze betrokken zijn bij de evaluatie van beloningen. Humane beeldvormende onderzoeken die worden beoordeeld, suggereren een toename in subcorticale activering (bijv. Accumbens) bij het maken van risicovolle keuzes (Kuhnen & Knutson, 2005; Matthews & et al., 2004; Montague & Berns, 2002) dat overdreven is bij adolescenten, in verhouding tot kinderen en volwassenen (Ernst et al., 2005; Galvan et al., 2006). Deze bevindingen suggereren verschillende trajecten voor belonings- of incentive-gebaseerd gedrag, waarbij systemen eerder werden ontwikkeld ten opzichte van controlesystemen die een langdurige en lineaire ontwikkelingskoers vertonen, in termen van het negeren van ongepaste keuzes en acties ten gunste van doelgerichte.
Bewijsmateriaal van neuroimaging-onderzoeken van de ontwikkeling van de menselijke hersenen
Recent onderzoek naar de ontwikkeling van adolescente hersenen is gebaseerd op vooruitgang in neuroimaging-methodologieën die gemakkelijk kunnen worden gebruikt bij de ontwikkeling van menselijke populaties. Deze methoden zijn gebaseerd op MRI-methoden (Magnetic Resonance Imaging) (zie Fig 2) en omvatten: structurele MRI, die wordt gebruikt om de grootte en de vorm van structuren te meten; functionele MRI die wordt gebruikt om patronen van hersenactiviteit te meten; en diffusie tensor imaging (DTI) die wordt gebruikt voor het indexeren van de connectiviteit van witte vezel traktaten. Bewijs voor ons ontwikkelingsmodel van competitie tussen corticale en subcorticale regio's wordt ondersteund door onrijpe structurele en functionele connectiviteit zoals gemeten door respectievelijk DTI en fMRI.
MRI-onderzoeken naar ontwikkeling van de menselijke hersenen
Verschillende studies hebben structurele MRI gebruikt om het anatomische verloop van normale hersenontwikkeling in kaart te brengen (zie beoordeling door Durston et al., 2001). Hoewel de totale hersengrootte op zesjarige leeftijd ongeveer 90% van de volwassen grootte is, ondergaan de grijze en witte materiecomponenten van de hersenen gedurende de adolescentie nog steeds dynamische veranderingen. Gegevens uit recente longitudinale MRI-onderzoeken geven aan dat het volume grijze stof een omgekeerd U-vormig patroon heeft, met een grotere regionale variatie dan witte stof (Giedd, 2004; Gogtay et al., 2004; Sowell et al, 2003; Sowell, Thompson en Toga, 2004). In het algemeen vervallen regio's die primaire functies bedienen, zoals motorische en sensorische systemen, het vroegst; hogere orde associatiegebieden, die deze primaire functies integreren, rijpen later (Gogtay et al., 2004; Sowell, Thompson en Toga, 2004). Studies die bijvoorbeeld MRI-gebaseerde metingen gebruiken, tonen aan dat corticale grijsstofverlies het vroegst voorkomt in de primaire sensorimotorische gebieden en recent in de dorsolaterale prefrontale en laterale temporale cortex (Gogtay et al., 2004). Dit patroon komt overeen met niet-humane primaat en menselijke postmortemstudies die aantonen dat de prefrontale cortex een van de laatste hersenregio's is die volwassen is (Bourgeois, Goldman-Rakic en Rakic, 1994; Huttenlocher, 1979). In tegenstelling tot grijze stof neemt het witte stofvolume in een ruwweg lineair patroon toe, terwijl het gedurende de gehele ontwikkeling tot in de volwassenheid toeneemt (Gogtay et al., 2004). Deze veranderingen weerspiegelen vermoedelijk de aanhoudende myelinisatie van axonen door oligodendrocyten die de neuronale geleiding en communicatie versterken.
Hoewel er minder aandacht is besteed aan subcorticale regio's bij het onderzoeken van structurele veranderingen, worden enkele van de grootste veranderingen in de hersenen over ontwikkeling waargenomen in deze regio's, met name in de basale ganglia (Sowell et al., 1999, Zie Fig 3) en vooral bij mannen (Giedd et al., 1996). Ontwikkelingsveranderingen in structureel volume in basale ganglia en prefrontale regio's zijn interessant in het licht van bekende ontwikkelingsprocessen (bijv. Dendritische arborisatie, celdood, synaptische snoei, myelinisatie) die optreden tijdens de kindertijd en adolescentie. Deze processen maken fijnafstemming en versterking mogelijk van verbindingen tussen prefrontale en subcorticale gebieden met ontwikkeling en leren die kunnen samenvallen met een grotere cognitieve controle. Hoe verhouden deze structurele veranderingen zich tot cognitieve veranderingen? Een aantal studies hebben de structurele rijping van de frontaalkwab en de cognitieve functie gerelateerd aan neuropsychologische en cognitieve metingen (bijv. Sowell et al., 2003). Meer in het bijzonder zijn associaties gerapporteerd tussen MRI-gebaseerde prefrontale corticale en basale ganglia regionale volumes en metingen van cognitieve controle (dwz het vermogen om een ongepaste respons ten gunste van een andere te vervangen of om de aandacht voor irrelevant stimulusattribuut te onderdrukken in het voordeel van relevant stimulusattribuut (Casey, Trainor et al., 1997). Deze bevindingen suggereren dat cognitieve veranderingen worden weerspiegeld in structurele hersenveranderingen en het belang onderstrepen van subcorticale (basale ganglia) en corticale (bijv. Prefrontale cortex) ontwikkeling.

DTI-onderzoeken naar de ontwikkeling van de menselijke hersenen
De MRI-gebaseerde onderzochte morfometriestudies suggereren dat corticale verbindingen worden verfijnd met de eliminatie van een overvloed aan synapsen en het versterken van relevante verbindingen met ontwikkeling en ervaring. Recente ontwikkelingen op het gebied van MRI-technologie, zoals DTI, bieden een potentieel hulpmiddel om de rol van specifieke witte stofkanalen in de ontwikkeling van de hersenen en het gedrag gedetailleerder te onderzoeken. Relevant voor dit artikel zijn de neuroimaging-onderzoeken die de ontwikkeling van fibre tracts hebben gekoppeld aan verbeteringen in cognitieve vaardigheden. In het bijzonder zijn associaties tussen op DTI-gebaseerde metingen van de ontwikkeling van de witte stof van de prefrontale zijde en cognitieve controle bij kinderen aangetoond. In één studie was de ontwikkeling van deze capaciteit positief gecorreleerd met prefrontale en pariëtale fibre tracts (Nagy, Westerberg en Klingberg, 2004) in overeenstemming met functionele neuroimaging-onderzoeken die differentiële rekrutering van deze regio's bij kinderen ten opzichte van volwassenen aantonen.
Met een vergelijkbare aanpak, Liston et al. (2005) hebben aangetoond dat witte stofkanalen tussen prefrontaal-basale ganglia en -posterior fibre tracts zich vanaf de kindertijd tot in de volwassenheid blijven ontwikkelen, maar alleen die trajecten tussen de prefrontale cortex en basale ganglia zijn gecorreleerd met impulscontrole, zoals gemeten door prestaties onderweg / nogo taak. De prefrontale fibre tracts werden gedefinieerd door regio's van belangen geïdentificeerd in een fMRI-onderzoek met dezelfde taak. Over beide ontwikkelings-DTI-onderzoeken waren metingen van het vezeltracé gecorreleerd met de ontwikkeling, maar de specificiteit van bepaalde vezelbanen met cognitieve prestaties werd aangetoond door het specifieke kanaal te dissociëren (Liston et al., 2005) of cognitieve vaardigheden (Nagy et al., 2004). Deze bevindingen onderstrepen het belang van het onderzoeken van niet alleen regionale, maar circuitgerelateerde veranderingen bij het doen van beweringen over leeftijdsafhankelijke veranderingen in neurale substraten van cognitieve ontwikkeling.
Functionele MRI-onderzoeken naar gedrags- en hersenontwikkeling
Hoewel structurele veranderingen gemeten met MRI en DTI in verband zijn gebracht met gedragsveranderingen tijdens de ontwikkeling, is een directere benadering voor het onderzoeken van structuur-functie-associatie het gelijktijdig meten van veranderingen in de hersenen en het gedrag, zoals bij fMRI. Het vermogen om functionele veranderingen in de zich ontwikkelende hersenen te meten met MRI heeft een aanzienlijk potentieel voor het gebied van de ontwikkelingswetenschap. In de context van de huidige paper biedt fMRI een manier om interpretaties van adolescentengedrag te beperken. Zoals eerder vermeld, wordt aangenomen dat de ontwikkeling van de prefrontale cortex een belangrijke rol speelt in de rijping van hogere cognitieve vaardigheden zoals besluitvorming en cognitieve controle (Casey, Tottenham en Fossella 2002b; Casey, Trainor et al., 1997). Veel paradigma's zijn gebruikt, samen met fMRI, om de neurobiologische basis van deze vermogens te beoordelen, waaronder flanker, Stroop en go / nogo-taken (Casey, Castellanos et al., 1997; Casey, Giedd, & Thomas, 2000a; Durston et al., 2003). Gezamenlijk tonen deze studies aan dat kinderen bij het uitvoeren van deze taken afzonderlijke maar vaak grotere, meer diffuse prefrontale gebieden aanwerven dan volwassenen. Het patroon van activiteit in hersengebieden centraal in taakprestaties (dat wil zeggen, dat correleert met cognitieve prestaties) wordt meer focus of verfijnd met de leeftijd, terwijl regio's die niet gecorreleerd zijn met taakprestaties afnemen in activiteit met de leeftijd. Dit patroon is waargenomen in beide dwarsdoorsneden (Brown et al., 2005) en longitudinale studies (Durston et al., 2006) en over verschillende paradigma's. Hoewel neuroimaging-onderzoeken het mechanisme van dergelijke ontwikkelingsveranderingen (bijv. Dendritische arborisatie, synaptische snoei) niet definitief kunnen karakteriseren, weerspiegelen de bevindingen de ontwikkeling binnen en verfijning van projecties van en naar geactiveerde hersenregio's met rijping en suggereren dat deze veranderingen gedurende een langdurige periode optreden. periode (Brown et al., 2005; Bunge, Dudukovic, Thomason, Vaidya en Gabrieli, 2002; Casey, Trainor et al., 1997; Casey et al., 2002a; Crone, Donohue, Honomichl, Wendelken en Bunge, 2006; Luna et al., 2001; Moses et al., 2002; Schlaggar et al., 2002; Tamm et al., 2002; Thomas et al., 2004; Turkeltaub, Gareau, Flowers, Zeffiro, & Eden, 2003).
Hoe kan deze methodiek ons informeren over de vraag of adolescenten inderdaad onvoldoende cognitieve controle hebben (impulsief) of risicovol zijn in hun keuzes en acties? Impulscontrole zoals gemeten door cognitieve beheertaken zoals de go / nogo-taak, toont een lineair ontwikkelingspatroon over de kindertijd en adolescentie zoals hierboven beschreven. Recente neuroimaging-onderzoeken zijn echter begonnen om beloningsgerelateerde verwerking specifiek voor het nemen van risico's bij adolescenten te onderzoeken (Bjork et al., 2004; Ernst et al., 2005; May et al., 2004). Deze studies hebben zich primair gericht op de regio van de accumbens, een deel van de basale ganglia die betrokken zijn bij het voorspellen van beloning, in plaats van karakterisering van de ontwikkeling van deze regio in combinatie met top-down controle regio's (prefrontale cortex). Hoewel een recent rapport van minder ventrale prefrontale activiteit bij adolescenten ten opzichte van volwassenen tijdens een monetaire besluitvormingstaak over risicovol gedrag is aangetoond (Eshel, Nelson, Blair, Pine en Ernst, 2007).
Over het algemeen hebben weinig studies onderzocht hoe de ontwikkeling van beloningscircuits in subcorticale regio's (bijv. Accumbens) verandert in samenhang met de ontwikkeling van corticale prefrontale regio's. Bovendien, hoe deze neurale veranderingen samenvallen met beloning zoeken, impulsiviteit en het nemen van risico's blijft relatief onbekend. Ons neurobiologisch model stelt voor dat de combinatie van verhoogde responsiviteit op beloningen en onvolwassenheid in gedragscontrolegebieden adolescenten kan beïnvloeden om directe, in plaats van langetermijnwinsten te zoeken, misschien als verklaring voor hun toename in risicovolle besluitvorming en impulsief gedrag. Het volgen van subcorticale (bijv. Accumbens) en corticale (bijv. Prefrontale) ontwikkeling van besluitvorming over de kindertijd tot in de volwassenheid, biedt extra beperkingen of de in de adolescentie gemelde veranderingen specifiek zijn voor deze periode van ontwikkeling, of weerspiegelen de rijping die zich gestaag voordoet in een periode van ontwikkeling. enigszins lineair patroon van kindertijd tot volwassenheid.
Empirisch bewijs van een recente fMRI-studie helpt ons neurobiologische model te ondersteunen en neemt een overgangsbenadering in het begrijpen van de adolescentie door de veranderingen voorafgaand aan en volgend op de adolescentie te onderzoeken. In dit onderzoek (Galvan et al., 2006), onderzochten we gedrags- en neurale reacties om manipulaties te belonen tijdens de ontwikkeling, met de nadruk op hersengebieden die betrokken zijn bij beloningsgerelateerd leren en gedrag bij dieren (Hikosaka en Watanabe, 2000; Pecina, Cagniard, Berridge, Aldridge en Zhuang, 2003; Schultz, 2006) en beeldvormende onderzoeken bij volwassenen (bijv. Knutson, Adams, Fong en Hommer, 2001; O, Doherty, Kringelbach, Rolls, Hornak, Andrews, 2001; Zald et al., 2004) en in onderzoeken naar verslaving (Hyman & Malenka, 2001; Volkow & Li, 2004). Gebaseerd op knaagdiermodellen (Laviola et al., 1999; Speer, 2000) en eerdere beeldvorming (Ernst et al., 2005), veronderstelden we dat adolescenten ten opzichte van kinderen en volwassenen een overdreven activering van de accumbens zouden vertonen, in combinatie met minder rijpe rekrutering van top-down prefrontale controlegebieden. Recent werk met vertraagde functionele connectiviteit tussen deze prefrontale en limbische subcorticale regio's in de adolescentie ten opzichte van volwassenen, biedt een mechanisme voor het ontbreken van top-down controle van deze regio's (Hare et al., 2007a).
Onze bevindingen kwamen overeen met knaagdiermodellen (Laviola, Macri, Morley-Fletcher en Adriani, 2003) en eerdere beeldvormende onderzoeken (Ernst et al., 2005) wat wijst op verbeterde accumbens-activiteit voor beloningen tijdens de adolescentie. Inderdaad toonden kinderen in vergelijking met kinderen en volwassenen een overdreven accumbensrespons in afwachting van een beloning. Zowel kinderen als adolescenten toonden echter een minder volwassen respons in prefrontale controlegebieden dan volwassenen. Deze bevindingen suggereren dat verschillende ontwikkelingsstrategieën voor deze regio's ten grondslag kunnen liggen aan de verbetering van de accumbens-activiteit ten opzichte van kinderen of volwassenen, wat op zijn beurt weer verband kan houden met het verhoogde impulsieve en risicovolle gedrag dat tijdens deze periode van ontwikkeling werd waargenomen (zie Fig 4).

Differentiële rekrutering van prefrontale en subcorticale regio's is gerapporteerd in een aantal ontwikkelingsfMRI-onderzoeken (Casey et al., 2002b; Monk et al., 2003; Thomas et al., 2004). Deze bevindingen zijn typisch geïnterpreteerd in termen van onvolgroeide prefrontale regio's in plaats van een disbalans tussen prefrontale en subcorticale regionale ontwikkeling. Gezien het bewijs van prefrontale regio's bij het begeleiden van passende acties in verschillende contexten (Miller & Cohen, 2001) onvolgroeide prefrontale activiteit kan een juiste inschatting van toekomstige uitkomsten en beoordeling van risicovolle keuzes belemmeren, en zou dus minder invloed kunnen hebben op de waardering van beloningen dan de accumbens. Dit patroon komt overeen met eerder onderzoek dat een verhoogde subcorticale, in verhouding tot corticale activiteit vertoont, wanneer beslissingen worden beïnvloed door directe langetermijnwinsten (McClure, Laibson, Loewenstein en Cohen, 2004). Verder is accumbens-activiteit aangetoond met fMRI om positief te correleren met daaropvolgend risicomijdend gedrag (Kuhnen & Knutson, 2005). Tijdens de adolescentie, in relatie tot de kindertijd of volwassenheid, kan een onvolgroeide ventrale prefrontale cortex mogelijk niet voldoende top-down controle bieden over robuust geactiveerde beloningsregio's (bijv. Accumbens), wat resulteert in minder invloed van prefrontale systemen (orbitofrontale cortex) ten opzichte van de accumbens in beloning waardering.
Waarom zouden de hersenen geprogrammeerd zijn om op deze manier te ontwikkelen?
Adolescentie is de overgangsperiode tussen kindertijd en volwassenheid, vaak gelijktijdig met de puberteit. De puberteit markeert het begin van seksuele rijping (Graber en Brooks-Gunn, 1998) en kan worden gedefinieerd door biologische markers. Adolescentie kan worden omschreven als een progressieve overgang naar volwassenheid met een vage ontogenetische tijdsverloop (Speer, 2000). Evolutionair gezien is de adolescentie de periode waarin onafhankelijkheidsvaardigheden worden verworven om het succes te vergroten bij de scheiding van de bescherming van het gezin, hoewel de kansen op schadelijke omstandigheden (bijvoorbeeld letsel, depressie, angst, drugsgebruik en verslaving) toenemen (Kelley, Schochet en Landry, 2004). Onafhankelijkheidszoekende gedragingen komen veel voor bij soorten, zoals toenamen in peer-directed sociale interacties en intensificaties in nieuwheidszoekende en risicovol gedrag. Psychosociale factoren beïnvloeden de neiging van adolescenten om riskant gedrag te vertonen. Riskant gedrag is echter het product van een biologisch gedreven onevenwichtigheid tussen toegenomen nieuwheid en sensatiezoekend in combinatie met onrijpe "zelfregulerende competentie" (Steinberg, 2004). Onze neurobiologische gegevens suggereren dat dit gebeurt door differentiële ontwikkeling van deze twee systemen (limbisch en controle).
Speculatie suggereert dat dit ontwikkelingspatroon een evolutionair kenmerk is. Je moet risicovol gedrag vertonen om je familie en dorp te verlaten om een partner te vinden en risico's te nemen op het moment dat hormonen adolescenten ertoe aanzetten seksuele partners te zoeken. In de huidige maatschappij waarin de adolescentie zich oneindig kan uitstrekken, met kinderen die bij hun ouders wonen en financiële afhankelijkheid hebben en die partners later in hun leven kiezen, kan deze evolutie als ongepast worden beschouwd.
Over de soorten heen is er bewijs voor verhoogde nieuwigheid en het nemen van risico's tijdens de adolescente jaren. Oudere leeftijdsgenoten opzoeken en met ouders vechten, die allemaal helpen om de adolescent weg te krijgen van het thuisgebied om te paren, is te zien bij andere soorten, inclusief knaagdieren, niet-menselijke primaten en sommige vogels (Speer, 2000). Ten opzichte van volwassenen vertonen periadolescente ratten verhoogde nieuwheidszoekende gedragingen in een vrije keuze-nieuwigheidsparadigma (Laviola et al., 1999). Neurochemisch bewijs geeft aan dat het evenwicht in het brein van adolescenten tussen corticale en subcorticale dopaminesystemen tijdens de adolescentie begint te verschuiven naar grotere corticale dopaminegehalten (Speer, 2000). Vergelijkbare langdurige dopaminerge enervatie door de adolescentie tot in de volwassenheid is ook aangetoond in de prefrontale cortex van de niet-menselijke primaat (Rosenberg en Lewis, 1995). Dus lijkt dit verhoogde schijnbare nemen van risico's over alle soorten heen te gaan en belangrijke adaptieve doelen te hebben.
Biologische predisposities, ontwikkeling en risico's
Individuele verschillen in impulsbeheersing en het nemen van risico's wordt al enige tijd erkend in de psychologie (Benthin, Slovic en Severson, 1993). Misschien is een van de klassieke voorbeelden van individuele verschillen die in deze vaardigheden worden gerapporteerd in de sociale, cognitieve en ontwikkelingspsychologische literatuur uitstel van bevrediging (Mischel, Shoda en Rodriguez, 1989). Vertraging van bevrediging wordt meestal beoordeeld in 3- tot 4-jarige peuters. De peuter wordt gevraagd of ze een kleine beloning (één cookie) of een grote beloning (twee cookies) prefereren. Het kind krijgt vervolgens te horen dat de onderzoeker de kamer zal verlaten om zich voor te bereiden op aanstaande activiteiten en legt aan het kind uit dat als zij op haar stoel blijft en geen koekje eet, zij de grote beloning zal ontvangen. Als het kind niet of niet kan wachten, moet ze een belletje rinkelen om de onderzoeker op te roepen en daardoor de kleinere beloning te ontvangen. Zodra het duidelijk is begrijpt het kind de taak, zit ze aan de tafel met de twee beloningen en de bel. Afleiding in de kamer wordt geminimaliseerd, zonder speelgoed, boeken of foto's. De onderzoeker komt terug na 15 min. Of nadat het kind de bel heeft gebeld, de beloningen heeft genuttigd of tekenen van angst heeft getoond. Mischel toonde aan dat kinderen zich meestal op twee manieren gedragen: (1) of ze bellen bijna onmiddellijk om de cookie te hebben, wat betekent dat ze er slechts één krijgen; of (2) ze wachten en optimaliseren hun winst, en ontvangen beide cookies. Deze observatie suggereert dat sommige individuen beter zijn dan anderen in hun vermogen om impulsen te beheersen in het licht van zeer opvallende prikkels en deze afwijking kan worden gedetecteerd in de vroege kinderjaren (Mischel et al., 1989) en ze lijken te blijven gedurende de adolescentie en jonge volwassenheid (Eigsti et al., 2006).
Wat zou individuele verschillen in optimale besluitvorming en gedrag kunnen verklaren? Sommige theoretici hebben gepostuleerd dat dopaminerge mesolimbische circuits, betrokken bij beloningsverwerking, ten grondslag liggen aan risicogedrag. Individuele verschillen in dit circuit, zoals allelische varianten in dopamine-gerelateerde genen, resulterend in te weinig of te veel dopamine in subcorticale regio's, kunnen verband houden met de neiging om risicovol gedrag aan te gaan (O'Doherty, 2004). Van de nucleus accumbens is aangetoond dat deze direct vóór het maken van risicovolle keuzes op monetair-risic paradigma's in activiteit toeneemt (Kuhnen & Knutson, 2005; Matthews et al., 2004; Montague & Berns, 2002) en zoals eerder beschreven, laten adolescenten overdreven accumbens-activiteit zien om resultaten te belonen ten opzichte van kinderen of volwassenen (Ernst et al., 2005; Galvan et al., 2006). Samengevat suggereren deze gegevens dat adolescenten mogelijk meer geneigd zijn tot risicovolle keuzes als groep (Tuinman & Steinberg, 2005), maar sommige adolescenten zijn meer geneigd dan anderen om risicovol gedrag aan te gaan, waardoor ze mogelijk een groter risico lopen op negatieve uitkomsten. Daarom is het belangrijk om rekening te houden met individuele variabiliteit bij het onderzoeken van complexe hersen-gedragsrelaties gerelateerd aan het nemen van risico's en het verwerken van beloningen in ontwikkelingspopulaties.
Individuele verschillen in risicogedrag onderzoeken, Galvan et al. (2007) Onlangs onderzocht de associatie tussen activiteit in beloningsgerelateerde neurale circuits in afwachting van een grote monetaire beloning met persoonlijkheidskenmerken van het nemen van risico's en impulsiviteit in de adolescentie. Functionele magnetische resonantie beeldvorming en anonieme zelfbeoordelingsbeoordelingsschalen van risicogedrag, risicoperceptie en impulsiviteit werden verkregen bij personen in de leeftijd van 7- en 29-jaren. Er was een positieve associatie tussen Accumbens-activiteit en de waarschijnlijkheid om risicovol gedrag aan te gaan tijdens de ontwikkeling. Deze activiteit varieerde als een functie van de individuele beoordelingen van verwachte positieve of negatieve gevolgen van dergelijk gedrag. Degenen die risicovol gedrag ervoeren als leidend tot ernstige gevolgen, activeerden de accumbens minder om te belonen. Deze associatie werd grotendeels aangedreven door de kinderen, waarbij de volwassenen de consequenties van dergelijk gedrag beoordeelden. Impulsiviteitsclassificaties waren niet geassocieerd met accumbens-activiteit, maar eerder met de leeftijd. Deze bevindingen suggereren dat tijdens de adolescentie sommige individuen meer risico lopen om risicovol gedrag aan te nemen als gevolg van veranderingen in de ontwikkeling in combinatie met variabiliteit in de aanleg van een bepaald individu om risicovol gedrag aan te gaan, in plaats van eenvoudige veranderingen in impulsiviteit (zie Fig 5).

Het gedrag van adolescenten is herhaaldelijk gekarakteriseerd als impulsief en risicovol (Steinberg, 2004, 2007), maar dit overzicht van de literatuur over beeldvorming suggereert verschillende neurobiologische substraten en verschillende ontwikkelingsroutes voor deze gedragingen. In het bijzonder wordt impulsiviteit geassocieerd met onrijpe ventrale prefrontale ontwikkeling en neemt deze geleidelijk af van de kindertijd tot de volwassenheid (Casey, Galvan et al., 2005). De negatieve correlatie tussen impulsiviteitsbeoordelingen en leeftijd in de studie door Galvan et al. (2007) ondersteunt dit begrip verder. Het nemen van risico's daarentegen is geassocieerd met een toename van accumbens-activiteit (Kuhnen & Knutson, 2005; Matthews et al., 2004; Montague & Berns, 2002), dat overdreven is bij adolescenten, in verhouding tot kinderen en volwassenen (Ernst et al., 2005; Galvan et al., 2006). Dus adolescente keuzes en gedrag kunnen niet worden verklaard door impulsiviteit of langdurige ontwikkeling van de prefrontale cortex alleen, omdat dan wordt voorspeld dat kinderen grotere risiconemers zijn. De bevindingen verschaffen een neurale basis voor de reden waarom sommige adolescenten een groter risico lopen dan anderen, maar bieden verder een basis voor hoe adolescentiegedrag verschilt van kinderen en volwassenen bij het nemen van risico's.
Samengevat suggereren deze gegevens dat hoewel adolescenten als een groep worden beschouwd als risiconemers (Tuinman & Steinberg, 2005), zullen sommige adolescenten meer geneigd zijn dan anderen om risicovol gedrag aan te gaan, waardoor ze mogelijk een groter risico lopen op negatieve uitkomsten. Deze bevindingen onderstrepen het belang van het beschouwen van individuele variabiliteit bij het onderzoeken van complexe hersen-gedragsrelaties gerelateerd aan het nemen van risico's en het verwerken van beloningen in ontwikkelingspopulaties. Verder kunnen deze individuele en ontwikkelingsverschillen helpen bij het verklaren van de kwetsbaarheid van sommige individuen voor het nemen van risico's in verband met middelengebruik en uiteindelijk voor verslaving.
Conclusies
Humane beeldvormende onderzoeken laten structurele en functionele veranderingen zien in frontostriatale regio's (Giedd et al., 1996, 1999; Jernigan et al., 1991; Sowell et al., 1999; ter beoordeling, Casey, Galvan et al., 2005) die parallel lijken te wijzen op een toename van cognitieve controle en zelfregulering (Casey, Trainor et al., 1997; Luna en Sweeney, 2004; Luna et al., 2001; Rubia et al., 2000; Steinberg, 2004Zie ook Steinberg, 2008, deze kwestie). Deze veranderingen lijken een verschuiving te vertonen in de activatie van prefrontale regio's van diffuse naar meer focale rekrutering in de tijd (Brown et al., 2005; Bunge et al., 2002; Casey, Trainor et al., 1997; Durston et al., 2006; Moses et al., 2002) en verhoogde rekrutering van subcorticale regio's tijdens de adolescentie (Casey et al., 2002a; Durston et al., 2006; Luna et al., 2001). Hoewel neuroimaging-onderzoeken het mechanisme van dergelijke ontwikkelingsveranderingen niet definitief kunnen karakteriseren, kunnen deze veranderingen in volume en structuur een weerspiegeling zijn van de ontwikkeling en verfijning van projecties naar en van deze hersenregio's tijdens rijping, wat erop duidt dat het systeem zich verder moet ontwikkelen.
Alles bij elkaar geven de hier gesynthetiseerde bevindingen aan dat verhoogd risicogedrag in de adolescentie geassocieerd is met verschillende ontwikkelingsroutes van subcorticaal genot en corticale controlegebieden. Deze ontwikkelingsveranderingen kunnen nog worden verergerd door individuele verschillen in activiteit van beloningssystemen. Hoewel de adolescentie is onderscheiden als een periode die gekenmerkt wordt door beloningzoekende en risicovolle gedragingen (Tuinman & Steinberg, 2005; Speer, 2000) individuele verschillen in neurale respons op beloning, predisponeren sommige adolescenten om meer risico te nemen dan anderen, waardoor ze een groter risico lopen op negatieve uitkomsten. Deze bevindingen bieden cruciale basis door de verschillende bevindingen met betrekking tot risicogedrag in de adolescentie samen te vatten en individuele verschillen en ontwikkelingsmarkeringen te begrijpen voor geneigdheid tot negatief gedrag.
Dankwoord
Dit werk werd gedeeltelijk ondersteund door subsidies van het National Institute of Drug Abuse R01 DA18879 en het National Institute of Mental Health 1P50 MH62196.
Referenties
- Benthin A, Slovic P, Severson H. Een psychometrische studie van de risicoperceptie bij adolescenten. Journal of Adolescence. 1993;16: 153-168. [PubMed]
- Bjork JM, Knutson B, Fong GW, Caggiano DM, Bennett SM, Hommer DW. Door incentives opgewekte hersenactivatie bij adolescenten: overeenkomsten en verschillen tussen jongvolwassenen. Journal of Neuroscience. 2004;24: 1793-1802. [PubMed]
- Bjorkland DF. De rol van conceptuele kennis bij de ontwikkeling van organisaties in het geheugen van kinderen. In: Brainerd CJ, Pressley M, redacteuren. Basisprocessen in geheugenontwikkeling: vooruitgang in onderzoek naar cognitieve ontwikkeling. New York: Springer-Verlag; 1985. pp. 103-142.
- Bjorkland DF. Hoe leeftijdsveranderingen in kennisbasis bijdragen aan de ontwikkeling van kindergeheugen: een interpretatieve beoordeling. Developmental Review. 1987;7: 93-130.
- Bourgeois JP, Goldman-Rakic PS, Rakic P. Synaptogenese in de prefrontale cortex van rhesusapen. Cerebrale cortex. 1994;4: 78-96. [PubMed]
- Brainerd CJ, Reyna VF. Geheugenonafhankelijkheid en geheugeninterferentie bij cognitieve ontwikkeling. Psychologisch overzicht. 1993;100: 42-67. [PubMed]
- Brown TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. Ontwikkelingsveranderingen in menselijke cerebrale functionele organisatie voor het genereren van woorden. Cerebrale cortex. 2005;15: 275-290. [PubMed]
- Bunge SA, Dudukovic NM, Thomason ME, Vaidya CJ, Gabrieli JD. Onvolwassen frontale kwab bijdragen aan cognitieve controle bij kinderen: aanwijzingen van fMRI. Neuron. 2002;33: 301-311. [PubMed]
- Case R. Validatie van een neo-Piagetiaans capaciteitsconstruct. Journal of Experimental Child Psychology. 1972;14: 287-302.
- Case R. Intellectuele ontwikkeling: geboorte tot volwassenheid. New York: Academic Press; 1985.
- Casey BJ, Castellanos FX, Giedd JN, Marsh WL, Hamburger SD, Schubert AB. et al. Implicatie van rechter frontostriatale circuits bij responsremming en aandachtstekortstoornis / hyperactiviteit. Tijdschrift van de American Academy of Child and Adolescent Psychiatry. 1997;36: 374-383. [PubMed]
- Casey BJ, Galvan A, Hare TA. Veranderingen in cerebrale functionele organisatie tijdens cognitieve ontwikkeling. Huidige mening in de neurobiologie. 2005;15: 239-244. [PubMed]
- Casey BJ, Giedd JN, Thomas KM. Structurele en functionele hersenontwikkeling en de relatie tot cognitieve ontwikkeling. Biologische psychologie. 2000a;54: 241-257. [PubMed]
- Casey BJ, Thomas KM, Davidson MC, Kunz K, Franzen PL. Dissociëren van striatale en hippocampusfunctie in ontwikkeling met een compatibiliteitstaak voor stimulusrespons. Journal of Neuroscience. 2002a;22: 8647-8652. [PubMed]
- Casey BJ, Thomas KM, Welsh TF, Badgaiyan RD, Eccard CH, Jennings JR, et al. Dissociatie van responsconflict, aandachtsselectie en verwachting met functionele magnetische resonantiebeeldvorming. Proceedings van de National Academy of Science. 2000b;97: 8728-8733.
- Casey BJ, Tottenham N, Fossella J. Klinisch onderzoek, beeldvorming, laesie en genetische benaderingen in de richting van een model van cognitieve controle. Ontwikkelingspsychobiologie. 2002b;40: 237-254. [PubMed]
- Casey BJ, Tottenham N, Liston C, Durston S. Beeldvorming van de ontwikkelende hersenen: wat hebben we geleerd over cognitieve ontwikkeling? Trends in cognitieve wetenschap. 2005;9: 104-110.
- Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN, et al. Een ontwikkelingsfunctioneel MRI-onderzoek naar prefrontale activering tijdens de uitvoering van een go-no-go-taak. Journal of Cognitive Neuroscience. 1997;9: 835-847.
- Crone E, Donohue S, Honomichl R, Wendelken C, Bunge S. Hersengebieden die flexibel regelgebruik bemiddelen tijdens de ontwikkeling. Journal of Neuroscience. 2006;26: 11239-11247. [PubMed]
- Dempster FN. Weerstand tegen interferentie: Ontwikkelingsveranderingen in een basisverwerkingsmechanisme. In: Howe ML, Pasnak R, redacteuren. Opkomende thema's in cognitieve ontwikkeling Volume 1: Foundations. New York: Springer; 1993. pp. 3-27.
- Diamond A. Ontwikkeling van de vaardigheid om recall te gebruiken om actie te geleiden, zoals aangegeven door de prestaties van baby's op AB. Kinder ontwikkeling. 1985;56: 868-883. [PubMed]
- Durston S, Davidson MC, Thomas KM, Worden MS, Tottenham N, Martinez A, et al. Parametrische manipulatie van conflict- en responsconcurrentie met behulp van snelle mixed-trial event-gerelateerde fMRI. Neuroimage. 2003;20: 2135-2141. [PubMed]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella J, et al. Een verschuiving van diffuse naar focale corticale activiteit met ontwikkeling. Ontwikkelingswetenschap. 2006;1: 18-20. [PubMed]
- Durston S, Hulshoff Pol HE, Casey BJ, Giedd JN, Buitelaar JK, van Engeland H. Anatomische MRI van het ontwikkelende menselijke brein: wat hebben we geleerd? Journal of American Academy of Child Adolescent Psychiatry. 2001;40: 1012-1020.
- Eaton LK, Kinchen S, Ross J, Hawkins J, Harris WA, Lowry R, et al. Jeugdgedrag risicogedrag-Verenigde Staten, 2005, bewakingssamenvattingen. Weekrapport voor morbiditeit en mortaliteit. 2006;55: 1-108. [PubMed]
- Eigsti IM, Zayas V, Mischel W, Shoda Y, Ayduk O, Dadlani MB, et al. Het voorspellen van cognitieve controle van kleuterschool tot late adolescentie en jongvolwassenheid. Psychological Science. 2006;17: 478-484. [PubMed]
- Ernst M, Nelson EE, Jazbec S, McClure EB, Monk CS, Leibenluft E, et al. Amygdala en nucleus accumbens in reacties op ontvangst en weglating van winst bij volwassenen en adolescenten. Neuroimage. 2005;25: 1279-1291. [PubMed]
- Eshel N, Nelson EE, Blair RJ, Pine DS, Ernst M. Neurale substraten van de selectie van de keuze bij volwassenen en adolescenten: ontwikkeling van de ventrolaterale prefrontale en anterior cingulate cortices. Neuropsychologia. 2007;45: 1270-1279. [PMC gratis artikel] [PubMed]
- Flavell JH, Feach DR, Chinsky JM. Spontane verbale repetitie in een geheugentaak als functie van de leeftijd. Kinder ontwikkeling. 1966;37: 283-299. [PubMed]
- Galvan A, Hare TA, Parra CE, Penn J, Voss H, Glover G, et al. Vroegere ontwikkeling van de accumbens ten opzichte van de orbitofrontale cortex zou ten grondslag kunnen liggen aan risicogedrag bij adolescenten. Journal of Neuroscience. 2006;26: 6885-6892. [PubMed]
- Galvan A, Hare T, Voss H, Glover G, Casey BJ. Risicobereidheid en het brein van de adolescent: wie loopt er risico? Ontwikkelingswetenschap. 2007;10: F8-F14. [PubMed]
- Gardener M, Steinberg L. Peer-invloed op het nemen van risico's, risicoformatie en risicovolle beslissingen in adolescentie en volwassenheid: een experimenteel onderzoek. Ontwikkelingspsychologie. 2005;41: 625-635. [PubMed]
- Giedd JN. Structurele magnetische resonantie beeldvorming van de hersenen van de adolescent. Annalen van de New York Academy of Sciences. 2004;1021: 77-85. [PubMed]
- Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A, et al. Hersenontwikkeling tijdens de kindertijd en adolescentie: een longitudinale MRI-studie. Nature Neuroscience. 1999;2: 861-863.
- Giedd JN, Snell JW, Lange N, Rajapakse JC, Casey BJ, Kozuch PL, et al. Kwantitatieve magnetische resonantie beeldvorming van de ontwikkeling van de menselijke hersenen: Leeftijden 4-18. Cerebrale cortex. 1996;6: 551-560. [PubMed]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, et al. Dynamische mapping van menselijke corticale ontwikkeling tijdens de kindertijd tot de vroege volwassenheid. Proceedings van de National Academy of Sciences in de Verenigde Staten van Amerika. 2004;101: 8174-8179. [PMC gratis artikel] [PubMed]
- Graber JA, Brooks-Gunn J. Puberty. In: Blechman EA, Brownell KD, redacteuren. Gedragsgeneeskunde en vrouwen een uitgebreid handboek. New York, NY: Guilford Press; 1998. pp. 51-58.
- Hare TA, Voss HU, Glover GH, Casey BJ. De hersenen van de adolescent en het potentiële risico op angst en depressie. 2007 Ingediend voor publicatie.
- Hare TA, Voss HU, Glover GH, Casey BJ. Concurrentie tussen prefrontale en subcorticale limbische systemen ten grondslag liggen aan emotionele reactiviteit tijdens de adolescentie. 2007b Ingediend voor publicatie.
- Harnishfeger KK, Bjorkland F. De ontogenese van remmingsmechanismen: een vernieuwde benadering van cognitieve ontwikkeling. In: Howe ML, Pasnek R, redacteuren. Opkomende thema's in cognitieve ontwikkeling. Vol. 1. New York: Springer-Verlag; 1993. pp. 28-49.
- Hikosaka K, Watanabe M. Vertragingsactiviteit van orbitale en laterale prefrontale neuronen van de aap, variërend met verschillende beloningen. Cerebrale cortex. 2000;10: 263-271. [PubMed]
- Huttenlocher PR. Synaptische dichtheid in menselijke frontale cortex-Ontwikkelingsveranderingen en effecten van veroudering. Hersenenonderzoek. 1979;163: 195-205. [PubMed]
- Hyman SE, Malenka RC. Verslaving en de hersenen: de neurobiologie van dwang en de persistentie ervan. Nature Beoordelingen Neuroscience. 2001;2: 695-703.
- Jernigan TL, Zisook S, Heaton RK, Moranville JT, Hesselink JR, Braff DL. Magnetische resonantie beeldvormingsafwijkingen in lenticulaire kernen en hersenschors bij schizofrenie. Archives of General Psychiatry. 1991;48: 811-823.
- Keating DP, Bobbitt BL. Individuele en ontwikkelingsverschillen in componenten voor cognitieve verwerking van mentale vaardigheden. Kinder ontwikkeling. 1978;49: 155-167.
- Kelley AE, Schochet T, Landry C. Annalen van de New York Academy of Sciences. 2004;1021: 27-32. [PubMed]
- Knutson B, Adams CM, Fong GW, Hommer D. Anticiperen op het verhogen van de geldbeloning trekt selectief nucleus accumbens aan. Journal of Neuroscience. 2001;21: RC159. [PubMed]
- Kuhnen CM, Knutson B. De neurale basis van het nemen van financiële risico's. Neuron. 2005;47: 763-770. [PubMed]
- Laviola G, Adriani W, Terranova ML, Gerra G. Psychobiologische risicofactoren voor de kwetsbaarheid voor psychostimulantia bij adolescenten en proefdiermodellen. Neuroscience and Biobehavioral Reviews. 1999;23: 993-1010. [PubMed]
- Laviola G, Macri S, Morley-Fletcher S, Adriani W. Abstract risicogedrag bij adolescente muizen: psychobiologische determinanten en vroege epigenetische invloed. Neuroscience and Biobehavioral Reviews. 2003;27: 19-31. [PubMed]
- Liston C, Watts R, Tottenham N, Davidson MC, Niogi S, Ulug AM, et al. Frontostriatale microstructuur moduleert efficiënte rekrutering van cognitieve controle. Cerebrale cortex. 2005;16: 553-560. [PubMed]
- Luna B, Sweeney JA. De opkomst van collaboratieve hersenfunctie: FMRI-onderzoeken naar de ontwikkeling van responsremming. Annalen van de New York Academy of Sciences. 2004;1021: 296-309. [PubMed]
- Luna B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ, et al. Rijping van wijdverspreide hersenfuncties dient voor de cognitieve ontwikkeling. Neuroimage. 2001;13: 786-793. [PubMed]
- Matthews SC, et al. Selectieve activering van de nucleus accumbens tijdens het nemen van risico's. Neuroreport. 2004;15: 2123-2127. [PubMed]
- May JC, Delgado MR, Dahl RE, Stenger VA, Ryan ND, Fiez JA, et al. Gebeurtenis-gerelateerde functionele magnetische resonantie beeldvorming van beloningsgerelateerde hersencircuits bij kinderen en adolescenten. Biologische psychiatrie. 2004;55: 359-366. [PubMed]
- McClure SM, Laibson DI, Loewenstein G, Cohen JD. Afzonderlijke neurale systemen waarderen onmiddellijke geldelijke beloningen. Science. 2004;306: 503-507. [PubMed]
- Miller EK, Cohen JD. Een integratieve theorie van de prefrontale cortexfunctie. Jaaroverzicht van neurowetenschap. 2001;24: 167-202.
- Mischel W, Shoda Y, Rodriguez MI. Vertraging van bevrediging bij kinderen. Science. 1989;244: 933-938. [PubMed]
- Monk CS, McClure EB, Nelson EE, Zarahn E, Bilder RM, Leibenluft E, et al. Onvolwassenheid van adolescenten in aandachtgerelateerde hersenbetrokkenheid bij emotionele gezichtsuitdrukkingen. Neuroimage. 2003;20: 420-428. [PubMed]
- Montague PR, Berns GS. Neurale economie en de biologische substraten van waardering. Neuron. 2002;36: 265-284. [PubMed]
- Moses P, Roe K, Buxton RB, Wong EC, Frank LR, Stiles J. Functionele MRI van globale en lokale verwerking bij kinderen. Neuroimage. 2002;16: 415-424. [PubMed]
- Munakata Y, Yerys BE. Alles bij elkaar nu: wanneer dissociaties tussen kennis en actie verdwijnen. Pscychologische wetenschap. 2001;12: 335-337.
- Nagy Z, Westerberg H, Klingberg T. Maturatie van witte stof is geassocieerd met de ontwikkeling van cognitieve functies tijdens de kindertijd. Journal of Cognitive Neuroscience. 2004;16: 1227-1233. [PubMed]
- O'Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Abstracte belonings- en strafrepresentaties in de menselijke orbitofrontale cortex. Nature Neurosci. 2001;4: 95-102. [PubMed]
- O'Doherty JP. Beloningsrepresentaties en beloningsgerelateerd leren in het menselijk brein: inzichten van neuroimaging. Huidige meningen in Neurobiology. 2004;14: 769-776.
- Pascual-Leone JA. Een wiskundig model voor overgang in de ontwikkelingsstadia van Piaget. Acta Psychologica. 1970;32: 301-345.
- Pecina S, Cagniard B, Berridge KC, Aldridge JW, Zhuang X. Hyperdopaminergische mutante muizen hebben een hogere "willen", maar niet "willen" voor zoete beloningen. Journal of Neuroscience. 2003;23: 9395-9402. [PubMed]
- Reyna VF, Farley F. Risico en rationaliteit bij de besluitvorming van adolescenten: implicaties voor theorie, praktijk en openbaar beleid. Psychologische wetenschap in het algemeen belang. 2006;7: 1-44.
- Rosenberg DR, Lewis DA. Postnatale rijping van de dopaminerge innervatie van prefrontale en motorische cortex van de aap: een immunohistochemische analyse van tyrosinehydroxylase. The Journal of Comparative Neurology. 1995;358: 383-400. [PubMed]
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A, et al. Functionele frontalisatie met leeftijd: Mapping van neurologische ontwikkelingsbanen met fMRI. Neuroscience and Biobehavioral Reviews. 2000;24: 13-19. [PubMed]
- Schlaggar BL, Brown TT, Lugar HM, Visscher KM, Miezin FM, Petersen SE. Functionele neuro-anatomische verschillen tussen volwassenen en kinderen van schoolgaande leeftijd in de verwerking van afzonderlijke woorden. Science. 2002;296: 1476-1479. [PubMed]
- Schultz W. Gedragstheorieën en de neurofysiologie van beloning. Jaarlijkse beoordelingen van psychologie. 2006;57: 87-115.
- Sowell ER, Peterson BS, Thompson PM, Welcome SE, Henkenius AL, Toga AW. Het in kaart brengen van de corticale verandering in de menselijke levensduur. Nature Neuroscience. 2003;6: 309-315.
- Sowell ER, Thompson PM, Holmes CJ, Jernigan TL, Toga AW. In vivo bewijs voor post-adolescente hersenrijping in frontale en striatale regio's. Nature Neuroscience. 1999;2: 859-861.
- Sowell ER, Thompson PM, Toga AW. Het in kaart brengen van veranderingen in de menselijke cortex gedurende de hele levensduur. Neuroloog. 2004;10: 372-392. [PubMed]
- Speer LP. De adolescente hersenen en aan leeftijd gerelateerde gedragsuitingen. Neuroscience and Biobehavioral Reviews. 2000;24: 417-463. [PubMed]
- Steinberg L. Risico's tijdens de adolescentie: welke veranderingen en waarom? Annalen van de New York Academy of Sciences. 2004;1021: 51-58. [PubMed]
- Steinberg L. Risico's tijdens de adolescentie: nieuwe perspectieven vanuit de hersen- en gedragswetenschappen. Huidige richtingen in Psychological Science. 2007;16: 55-59.
- Steinberg L. Een perspectief van sociale neurowetenschap op het nemen van risico's voor adolescenten. Developmental Review. 2008;28: 78-106. [PMC gratis artikel] [PubMed]
- Tamm L, Menon V, Reiss AL. Rijping van de hersenfunctie geassocieerd met responsremming. Tijdschrift van de American Academy of Child and Adolescent Psychiatry. 2002;41: 1231-1238. [PubMed]
- Thomas KM, Hunt RH, Vizueta N, Sommer T, Durston S, Yang Y, et al. Bewijs van ontwikkelingsverschillen in impliciet sequentieleren: een FMRI-onderzoek van kinderen en volwassenen. Journal of Cognitive Neuroscience. 2004;16: 1339-1351. [PubMed]
- Turkeltaub PE, Gareau L, Flowers DL, Zeffiro TA, Eden GF. Ontwikkeling van neurale mechanismen voor lezen. Nature Neuroscience. 2003;6: 767-773.
- Volkow ND, Li TK. Drugsverslaving: de neurobiologie van verkeerd gedrag. Nature Beoordelingen Neuroscience. 2004;5: 963-970.
- Yurgelun-Todd D. Emotionele en cognitieve veranderingen tijdens de adolescentie. Huidige mening in de neurobiologie. 2007;17: 251-257. [PubMed]
- Zald DH, Boileau I, El-Dearedy W, Gunn R, McGlone F, Dichter GS, et al. Dopamine-overdracht in het menselijk striatum tijdens geldbeloningstaken. Journal of Neuroscience. 2004;24: 4105-4112. [PubMed]