Frontul Behav Neurosci. 2015 Mai 19; 9: 128. doi: 10.3389 / fnbeh.2015.00128. eCollecție 2015.
Andreatta M1, Pauli P1.
Abstract
În condiționarea clasică, un stimul inițial neutru (stimul condiționat, CS) devine asociat cu un eveniment biologic important (stimul necondiționat, SUA), care ar putea fi durere (condiționare aversivă) sau mâncare (condiționare apetisantă). După câteva asociații, CS este capabil să inițieze fie răspunsuri defensive, fie consumative. Contrar condiționării aversive, condiționarea apetitivă este rar investigată la om, deși importanța sa pentru comportamentele normale și patologice (de exemplu, obezitatea, dependența) este incontestabilă. Prezentul studiu intenționează să traducă descoperirile animalelor cu privire la condiționarea apetitivă la oameni care folosesc alimente ca SUA. Treizeci și trei de participanți au fost anchetați între orele 8 și 10 dimineața, fără mic dejun, pentru a se asigura că le este foame. În timpul a două faze de achiziție, o formă geometrică (avCS +) a prezis un SU aversiv (șoc electric dureros), o altă formă (appCS +) a prezis un SU apetisant (ciocolată sau covrig sărat în funcție de preferința participanților) și o a treia formă (CS- ) nu a prezis nici SUA. Într-o fază de dispariție, aceste trei forme plus o formă nouă (NOU) au fost prezentate din nou fără livrare în SUA. Evaluările valenței și excitării, precum și răspunsurile la tresărire și conductanța pielii (SCR) au fost colectate ca indici de învățare. Am găsit condiționări aversive și apetisante de succes. Pe de o parte, avCS + a fost evaluat ca fiind mai negativ și mai excitant decât CS- și a indus potențarea de tresărire și SCR îmbunătățit. Pe de altă parte, appCS + a fost evaluat mai pozitiv decât CS- și a indus atenuarea tresăririi și SCR mai mare. Pe scurt, am confirmat cu succes descoperirile animalelor la oameni (flămânzi), demonstrând învățarea apetitivă și învățarea aversivă normală.
Introducere
Prezicerea amenințărilor și a alimentelor este de o importanță crucială pentru supraviețuirea oricărui organism. În condiționarea clasică (Pavlov, 1927), un indiciu precede un eveniment aversiv, cum ar fi un șoc electric dureros (stimul stimulativ necondiționat, SUA) sau un eveniment apetit, cum ar fi un pelet alimentar (apetitul american), de mai multe ori. Ulterior, acest stimul singur (acum etichetat CS, CS +) este capabil să provoace fie răspunsuri defensive, fie consumatoare. Fostul tip de învățare asociativă se numește condiționarea aversivă, în timp ce acesta din urmă este numit condiționarea apetitului.
Deși predicția evenimentelor apetisante este la fel de importantă pentru supraviețuire ca și prezicerea evenimentelor aversive, condiționarea apetitului este remarcabil mai puțin investigată la animale (Bouton și Peck, 1989; Koch și colab., 1996; McDannald și colab., 2011, pentru o revizuire a se vedea Martin-Soelch și colab., 2007), precum și la om (Klucken și colab., 2009, 2013; Austin și Duka, 2010; Delgado și colab., 2011; Levy și Glimcher, 2011). Această lipsă de cercetare s-ar putea datora complexității paradigmei apetisante comparativ cu cea aversitivă. De exemplu, alimentele ca un prim întăritor trebuie livrate atunci când organismul este înfometat pentru a fi recompensat (pentru o recapitulare recentă vezi Dickinson și Balleine, 1994; Clark și colab., 2012). În cercetarea umană, această dificultate a fost depășită prin utilizarea banilor (Austin și Duka, 2010; Delgado și colab., 2011; Levy și Glimcher, 2011) sau imagini erotice (Klucken și colab., 2009, 2013). Cu toate acestea, studiile neuro-imagistice au arătat că activitățile primare (de exemplu, gustări sau băuturi) și secundar (adică bani) întăresc anumite regiuni ale creierului (de exemplu, striatum), dar implică și moduri distincte de activareDelgado și colab., 2011; Levy și Glimcher, 2011).
Din cunoștințele noastre, numai câteva studii de condiționare a oamenilor au examinat efectele substanțelor primare de întărire a apetitului precum mirosul (Gottfried și colab., 2002), apă (Kumar și colab., 2008) sau alimente (Prévost și colab., 2012). În primul studiu, Gottfried și colab. (2002) fețele neutre asociate (CS) cu miros neplăcut, plăcut sau neutru. Interesant, ei au descoperit o mai mare actiune in cortexul orbitofrontal (OFC) si in striatum ventral ca raspuns la apetitul CS + fata de CS + aversiv. Ei au concluzionat că OFC procesează valoarea mirosului și implică transferul valorii afective din sistemul olfactiv (US) către sistemul vizual (CS). Activarea (ventro) striatală a fost interpretată ca reflectând CR apetită provocată de apetitul CS +. În al doilea studiu, Kumar și colab. (2008) a invitat participanții (controale sănătoase și pacienți cu depresie majoră) la laborator devreme dimineața și le-a cerut să se abțină de la băut în timpul nopții, pentru a se asigura că au fost sete înainte de scanare. Imaginile fractale au fost CS și 0.1 ml apă din SUA. Interesant este faptul că, printre alte activări, participanții sănătoși (dar nu pacienții cu depresie) au prezentat o mai mare activare a striatumului ventral la CS + apetit, ceea ce sugerează că acest stimul a fost prelucrat ca o recompensă. În al treilea studiu, Prévost și colab. (2012) au prezentat, de asemenea, imagini fractale ca CS și gustări dulci sau sărate ca SUA în funcție de preferința participanților. CS a fost prezentat timp de 6 secunde și în ultima secundă a fost prezentată suplimentar o imagine a alimentelor. De fiecare dată când a apărut imaginea alimentară, experimentatorul a pus o bucată de mâncare în mâinile participanților, cărora li s-a permis să consume gustarea imediat. Din păcate, autorii nu au raportat activarea creierului către CS + apetisant în timpul fazei de condiționare clasică; cu toate acestea, au observat o decelerare cardiacă mai mică la CS + recompensat comparativ cu CS nerecompensat, indicând efecte de condiționare diferențiale.
Lipsa de studii care utilizează răspunsul surprinzător ca indice pentru condiționarea apetitului este surprinzătoare, în special având în vedere utilizarea sa largă în condiționarea aversivă. Răspunsul inițial este un răspuns defensiv, ancestral și automat, la evenimente bruște, neașteptate și puternice aversive (Koch, 1999). Acest răspuns defensiv este mediat de o cale neuronală relativ simplă care implică neuronii rădăcinii cohleare, nucleul pontinei caudale a formării reticulare (PnC) și motoneuronii spinării (Fendt și Fanselow, 1999; Koch, 1999). Studiile pe animale au arătat că potențarea de stingere depinde de proiecțiile de la amigdală la PnC (Fendt și Fanselow, 1999; Koch, 1999), în timp ce atenuarea șocului depinde de nucleul accumbens intact (NAcc, Koch, 1999). Această modulare a răspunsului surprinzător este o măsură utilă implicită pentru valența stimulilor prezentați în prim plan. Astfel, potențarea indică valența negativă, în timp ce atenuarea indică valența pozitivă, ambele fără a fi puternic influențate de procesele cognitive (Hamm și Weike, 2005; Andreatta și colab., 2010). Din câte știm, un singur studiu pe animale a investigat condiționarea apetitului prin măsurarea răspunsurilor surprinzătoare ca măsură dependentă. Interesant, șobolanii examinați au arătat o atenuare a uimii la un CS + (adică la lumină) asociat cu eliberarea unei soluții de zaharoză. În mod special, o astfel de atenuare a fost afectată la acele animale cu leziuni NAcc, dar nu la cei cu leziuni amigdale, sugerând că NAcc joacă un rol specific în apariția CR apetit și în atenuarea răspunsurilor surprinzătoare (Koch și colab., 1996).
Studiul actual este, după cunoștințele noastre, primul care traduce această paradigmă de condiționare a apetitului la oameni, prin utilizarea armatorilor primari ca SUA, adică dulce (Smarties de ciocolată®) sau mâncare sărată (covrigi mici) și modulația surprinzătoare ca măsură a CR-urilor. Ne-am așteptat ca aplicația appCS + să declanșeze CR puternice apetisante, așa cum se reflectă în atenuarea amortizării, SCR îmbunătățită și evaluarea pozitivă a valenței în comparație cu ceilalți stimuli, adică avCS + și CS-.
Materiale și metode
Participanții
Patruzeci și doi de voluntari au acceptat să participe la studiu și au primit credite de curs. Nouă participanți au fost excluși din analiză, doi din cauza problemelor tehnice, trei pentru că au fost codificați ca non-respondenți (amplitudine medie de tresărire <5 μV) și patru pentru că nu au prezentat suficiente răspunsuri de tresărire pe afecțiune (minim = 2; pentru detalii, a se vedea Materiale și metodă). În cele din urmă, 33 de participanți au fost luați în considerare în analiză (16 bărbați; vârsta medie: 22.09 ani, SD: 2.84; interval: 18-29 ani). Patru participanți nu erau germani nativi, iar șase erau stângaci. Un participant a rămas fără conștiință de asociațiile CS-SUA în timpul experimentului (a se vedea procedura), dar am decis să nu excludem acest participant deoarece răspunsurile sale au fost normale și nu au afectat rezultatele.
materiale
Stimuli necondiționate (SUA)
Au fost folosite două tipuri de SUA. Ca și SUA aversivă, am aplicat un șoc electric ușor dureros asupra antebralei non-dominante a participanților. Șocul electric a fost livrat cu ajutorul a doi electrozi cu diametrul 9 mm și distanța 30 cm. Stimularea electrică a constat într-un stimulent de impulsuri cu o frecvență de 50 Hz și durata 200 ms, generată de un stimulator curent (Digitimer DS7A, Digitimer Ltd, Welwyn Garden City, Marea Britanie, 400 V, maxim 9.99 mA). Intensitatea șocului electric a fost determinată individual printr-o procedură de prag descrisă anterior (Andreatta și colab., 2010). Pe scurt, participanții au suferit două serii de intensități ascendente și descendente în intervale de 0.5 mA. Ei trebuiau să evalueze fiecare stimul pe o scală vizuală, variind de la 0 (simțind nimic) la 10 (durere foarte intensă), cu 4 ca o ancoră pentru prag (durere simplă). Intensitatea medie a stimulului electric a fost 2.12 mA (SD = 0.56) și a fost evaluat ca dureros (M = 6.45, SD = 1.73). Apetitul american a constat din ciocolată (Smarties®) sau covrig mic sărat. Alegerea apetitivului SUA a depins de preferința individuală a participantului, așa cum a fost raportat în timpul interviului preliminar. Anume, participanții au trebuit să raporteze dacă în mod normal mănâncă mâncare sărată sau dulce în timpul micului dejun. De asemenea, ar putea alege liber dacă preferă ciocolata sau covrigul sărat în timpul experimentului în sine. La final, 22 de participanți au ales ciocolata și 11 covrigul mic sărat.
Stimulări condiționate (CS)
Forme geometrice (pătrat albastru, cerc galben, triunghi verde, hexagon roșu) cu o diagonală de 8 cm au fost prezentate ca CS. Formele au fost prezentate în mijlocul unui ecran de calculator negru pentru 8 s. O formă (avCS +) a fost întotdeauna asociată cu SUA aversivă (șoc electric dureros), o formă (appCS +) a fost întotdeauna asociată cu apetitul american (ciocolată sau covrigul sărat), o formă (CS-) sau US apetitul, iar a patra formă (NEW) a fost prezentată în faza de extincție, dar nu și în faza de achiziție, pentru a asigura neutralitatea acesteia.
Startle Probe
Zgomotul alb de 103 dB cu durata de 50 ms a fost folosit ca sondă de tresărire. Stimulii acustici au fost prezentați binaural pe căști și au apărut aleatoriu la 4-6 secunde după apariția formei.
chestionare
Înainte și după experiment, participanții au trebuit să completeze versiunile germane de inventar de anxietate de stat (STAI, Laux și colab., 1981) și programul pozitiv negativ afectat (PANAS, Krohne și colab., 1996). STAI este un inventar pentru evaluarea trăsăturii participanților și / sau anxietății stării și constă din 20 de articole atât pentru trăsătură, cât și pentru versiunile de stare. Nivelul de anxietate al participanților înainte (M = 37.06, SD = 7.80) și după (M = 39.33, SD = 9.16) experimentul nu sa schimbat semnificativ [t(32) = 1.61, p = 0.117]. Trasaturile de anxietate din probă curentă au fost cuprinse între 20 și 58 (M = 36.6, SD = 8.98), care este comparabil cu intervalul normal publicat de adulți (Laux și colab., 1981). Panas (Krohne și colab., 1996) este un indice pentru starea de spirit pozitivă și cea negativă. Persoanele cu scoruri ridicate la scara afectelor pozitive (PAS) sunt predispuse la emoții precum entuziasmul, în timp ce indivizii cu scoruri ridicate la scara afectelor negative (NAS) sunt predispuse la emoții precum stresul. Fiecare articol este format dintr-un adjectiv, iar participanții indică pe o scară cuprinsă între 1 (foarte ușor) și 5 (extrem) în ce măsură adjectivul reflectă sentimentele lor în acel moment particular. Nu s-au găsit diferențe semnificative în efectul negativ al participanților ca urmare a experimentului [începe: M = 11.67, SD = 2.29; Sfârșit: M = 12.88, SD = 4.69; t(32) = 1.55, p = 0.130]. Cumva, participanții și-au schimbat în mod semnificativ starea de spirit pozitivă încă de la început (M = 26.72, SD = 4.70) până la sfârșit (M = 23.66, SD = 7.02) din experiment [t(31) = 3.11, p = 0.004]. Această scădere a dispoziției pozitive a participanților ar fi putut fi legată de neplăcerea paradigmei (au fost prezentate șocuri electrice dureroase, precum și zgomot alb aversiv).
Procedură
După sosirea în laborator, participanții au citit și au semnat un formular de consimțământ informat, aprobat de comitetul de etică al Departamentului de Psihologie al Universității din Würzburg. Nu au fost informați despre situația de urgență dintre CS și SUA. După ce au fost completate chestionarele, electrozii au fost atașați și procedura pragului de durere a fost efectuată așa cum s-a descris mai sus.
În timpul faza de obișnuință, cele patru forme geometrice au fost prezentate de două ori cu un interval între probe (ITI) care variază între 18 și 25 s (media: 21.5 s). În timpul acestei faze nu au fost furnizate sonde americane sau surprinzătoare.
Înainte de faza de achiziție, s-au livrat șapte perioade de zgomot alb la fiecare 7-15 s, pentru a scădea reactivitatea inițială.
Următoarele două fazele de achiziție au fost identice (Figura 1). Fiecare fază de achiziție a constat în studiile 24: 8 CS-trials, 8 avCS + trialuri și 8 appCS +. Secvența CS a fost pseudorandom, singura restricție fiind aceea că același stimul nu a putut fi prezentat mai mult de două ori la rând. În mod special, avCS + a fost prezentat împreună cu un fulger ca un simbol al șocului electric, iar SUA dureroasă a fost livrată la offset. ApCS + a fost prezentată în compoziție cu o imagine a Smarties sau a unui covrigi sărate, iar participantul putea alege un Smarties sau un covrig de la un borcan. CS-a fost prezentată împreună cu un simbol de interdicție și nu a fost livrată nicio țară americană. Pe parcursul a trei dintre prezentările 8 CS ale fiecărui tip, s-a furnizat o probă de șoc între 4 și 6 s după debutul stimulului. Trei sonde de surprindere suplimentare au fost prezentate în timpul ITI-urilor pentru a asigura imprevizibilitatea acestora și pentru a reduce habituarea surprinzătoare. ITI, alcătuită dintr-un ecran negru, a variat între 18 și 25 s cu o medie de 21.5 s.
FIGURA 1
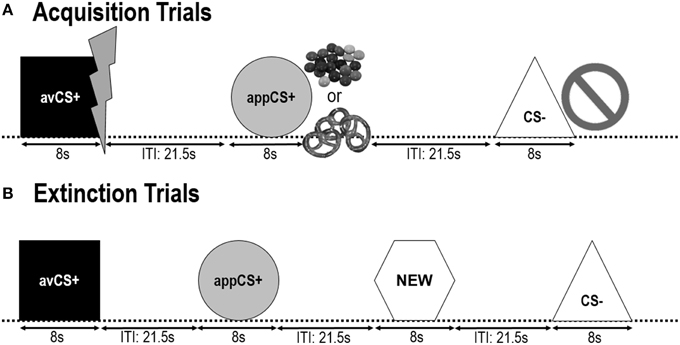
Figura 1. Încercările în timpul celor două faze de achiziție (A) și a fazei de extincție (B). Participantii au aflat ca o forma (avCS +) a prezis un soc electric dureros, o forma (appCS +) a prezis fie o bucata de ciocolata sau un covrig mica (conform preferintei), iar oa treia forma (CS-) din punct de vedere biologic. Fiecare formă a fost prezentată în legătură cu o imagine care prezintă un șoc electric, smarties / covrigei sărate sau nimic în funcție de asocierea cu SUA. În timpul fazei de extincție, cele trei forme geometrice au fost prezentate încă o dată, dar nu au fost livrate SUA. În plus, o a patra formă geometrică (NEW) a fost prezentată ca un control neutru.
În timpul faza de extincție, participanții au văzut din nou cele trei forme geometrice (adică avCS +, appCS + și CS-), plus o nouă formă neutră (NEW). Nu a fost livrată nicio țară americană, iar formele nu au fost prezentate împreună cu imagini ale flash-urilor, ciocolată / covrigi sărate sau interdicții. Fiecare stimul a fost prezentat de opt ori într-o ordine pseudorandomică (adică, același stimul nu a fost prezentat mai mult de două ori la rând), creând studii 32. Stimulele sondei initiale au fost prezentate in timpul 4 din prezentari de stimuli 8 pentru fiecare tip de CS. Ca și în faza de achiziție și de obișnuință, ITI-urile au variat între 18 și 25 s, iar sonde suplimentare de suprimare 4 au fost livrate imprevizibil în timpul ITI-urilor.
După fiecare fază, participanții au evaluat valența (plăcerea) și excitarea (excitația) CS cu scale analoage vizuale (VAS) variind de la 1 la 9. Scala de valtare a variat de la "1" indicând "foarte neplăcut" la "9" indicând "foarte plăcut"; scara excitării a variat de la "1" indicând "calm" la "9" evaluată după cele două faze de achiziție și faza de extincție. Participanții au văzut o formă geometrică pentru 1 s și apoi au trebuit să indice dacă această formă a fost asociată cu șocul electric, cu covriful de ciocolată / sărată, fără nimic sau dacă nu au putut face nici o asociere. În mod deosebit, toți participanții (cu excepția unuia) au fost conștienți de situația de urgență după achiziționarea 2.
Reducerea datelor
Răspunsurile fiziologice au fost înregistrate cu un amplificator V-Amp 16 și Vision Recorder V-Amp Edition Software (versiunea 1.03.0004, BrainProducts Inc., München, Germania). Sa aplicat o rată de eșantionare de 1000 Hz și un filtru cu crestătură 50 Hz. Analizele offline au fost efectuate cu ajutorul analizorului Brain Vision Analyzer (versiunea 2.0, BrainProducts Inc., München, Germania).
Răspunsul inițial
Răspunsul inițial a fost măsurat prin intermediul electromiografiei (EMG) la stânga orbicularis oculi mușchi cu doi electrozi 5 mm Ag / AgCl. În conformitate cu orientările (Blumenthal și colab., 2005), un electrod a fost poziționat sub pupilă, iar al doilea 1 cm lateral. Electrozii de sol și de referință au fost plasați pe mastoidele din dreapta și respectiv stânga. Înainte de atașarea electrozilor, pielea a fost ușor abrazită și curățată cu alcool pentru a păstra impedanța sub 10 kΩ. Semnalul electromiografic a fost filtrat offline, cu un filtru de cutoff low 28 Hz și un filtru de cutoff de înaltă frecvență 500 Hz. Apoi, semnalul EMG a fost rectificat și sa aplicat o medie mobilă de 50 ms. Am folosit 50 ms înainte de debutul sondei de start ca o linie de bază (Grillon și colab., 2006). Răspunsurile la sonde de surprindere au fost marcate manual, iar încercările cu schimbări excesive de bază (± 5 μV) sau artefacte de circulație au fost excluse din analiza ulterioară. Răspunsurile Startle mai mici decât 5 μV au fost codificate ca zero și luate în considerare pentru calcularea amplitudinii de strigare (Blumenthal și colab., 2005). În total, au fost respinse 10.4% din studii și un număr minim de 2 din răspunsurile surprinzătoare 3 în fazele de achiziție și 4 din răspunsurile surprinzătoare 8 în faza de extincție pentru fiecare condiție au fost necesare pentru a menține participantul în grupul de analiză. Din acest motiv, patru participanți au fost excluși. Amplitudinea de vârf a fost definită ca vârful maxim față de linia de bază în timpul ferestrei de timp 20-120 ms după debutul sondei de start. Datele brute au fost apoi normalizate la subiecți folosind z- scoruri și apoi scoruri T pentru a reduce influența variabilității individuale și pentru a detecta mai bine procesele psihologice. Scorurile T au fost medii pentru fiecare condiție (avCS +, appCS +, CS-, NEW și ITI). Pentru a investiga potentierea surprinderii sau atenuarea amortizarii, scorurile pentru raspunsurile ITI au fost scazute din raspunsurile surprinse pentru fiecare conditie.
Reacția de conduită a pielii (SCR)
Răspunsul de conductivitate a pielii (SCR) a fost înregistrat utilizând doi electrozi 5 mm Ag / AgCl plasați pe palma mâinii non-dominante. Răspunsul galvanic a fost filtrat offline cu un filtru de cutoff de înaltă frecvență 1 Hz. SCR a fost definită ca diferența (în μS) între debutul răspunsului (1-3 s după debutul stimulului) și vârful de răspuns (Tranel și Damasio, 1994; Delgado și colab., 2011). În analiza SCR nu au fost luate în considerare încercările care conțin sonde de șoc. Răspunsurile de mai jos 0.02 μS au fost codificate ca fiind zero. Cinci participanți au fost excluși din analiza SCR deoarece au avut un SCR mediu mai mic decât 0.02 μS. Datele de conductanță a pielii brute au fost transformate în rădăcină pătrată pentru a normaliza distribuția, iar scorurile au fost medii pentru fiecare condiție separat pentru cele două faze de achiziție (avCS +, appCS +, CS-) și faza de extincție (avCS +, appCS +, CS- și NEW ).
Analiza statistică
Toate datele au fost analizate cu SPSS pentru Windows (versiunea 20.0, SPSS Inc.). Pentru răspunsurile fiziologice s-au calculat analizele multivariate de variație separate (ANOVA) pentru cele două faze de achiziție și faza de extincție. ANOVA pentru fazele de achiziție a avut stimuli (avCS +, appCS +, CS-) și fază (Acquisition 1, Acquisition 2) ca factori de subiecți. ANOVA pentru faza de extincție a avut doar un stimulent (avCS +, appCS +, CS-, NEW) ca un factor în subiecți. Evaluările privind valența, excitația și situația de urgență au fost analizate cu ANOVA separate, conținând stimulul factorilor intra-subiecți (avCS +, appCS +, CS- și NEW) și faza. Acest factor a avut patru nivele pentru evaluarea valenței și excitației (T1: după faza de obișnuință, T2: după prima fază de achiziție, T3: după a doua fază de achiziție, T4: după faza de extincție), dar trei niveluri pentru evaluările de urgență prima fază de achiziție, T1: după a doua fază de achiziție, T2: după faza de extincție).
Nivelul alfa (α) a fost setat la 0.05 pentru toate analizele. Dimensiunea efectului este raportată ca fiind parțial η2.
REZULTATE
Valorile valenței și excitării pentru fiecare fază sunt prezentate în figura 2; răspunsurile surprinzătoare și SCR-urile sunt prezentate în Figura 3.
FIGURA 2
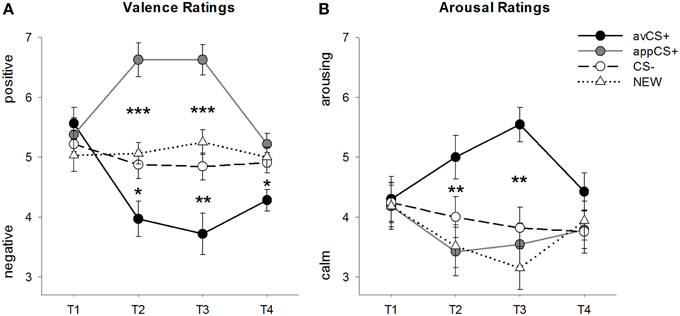
Figura 2. Calificări pentru valență (A) și excitare (B). Liniile (cu erori standard) descriu ratingurile după faza de obișnuință (T1), Achiziția 1 (T2), Achiziția 2 (T3) și faza de extincție (T4). Aversiunea CS + (linia solidă neagră) a obținut valență negativă și excitare ridicată după cele două faze de achiziție, comparativ cu CS- (linia punctată negru) și NEW (linia neagră punctată). Este important faptul că apetitul CS + (linie solidă gri) a dobândit o valență pozitivă în comparație cu CS- și NEW. *p <0.05, **p > 0.01, ***p <0.001.
FIGURA 3
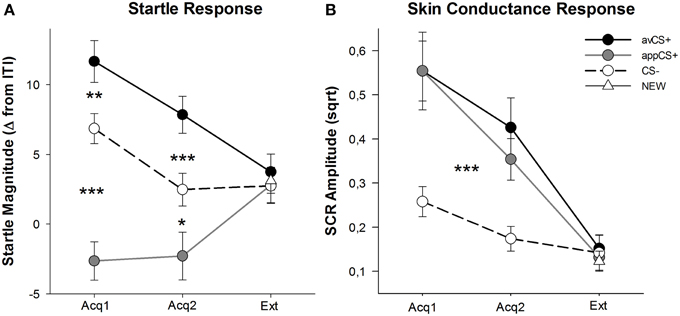
Figura 3. (Acq1), cea de-a doua fază de achiziție (Acq2) și faza de extincție (Ext). Răspunsurile Startle au fost puternic potențate față de linia CS + aversivă (linia solidă neagră) și au fost atenuate în mod semnificativ față de CS + (linia solidă gri) în comparație cu linia CS (linie neagră punctată) în timpul fazelor de achiziție. SCR a fost semnificativ mai mare pentru avCS + și appCS + comparativ cu CS-. Nu s-au descoperit diferențe în timpul fazei de dispariție. *p <0.05, **p > 0.01, ***p <0.001.
Calificative
ANOVA pentru valența în cursul achiziției au fost constatate efecte principale semnificative ale stimulului [F(3, 93) = 17.26, GG-ε = 0.801, p <0.001, parțial η2 = 0.358] și faza [F(3, 93) = 3.30, GG-ε = 0.731, p = 0.039, parțial η2 = 0.096], precum și o interacțiune semnificativă între stimul și faza [F(9, 279) = 9.54, GG-ε = 0.463, p <0.001, parțial η2 = 0.235]. Urmare t- testele indică faptul că valențele celor patru forme geometrice de la începutul experimentului au fost identice (ps> 0.19), în timp ce după achiziția 1 și achiziția 2, avCS + a fost evaluat ca fiind deosebit de negativ și appCS + ca deosebit de pozitiv. Mai exact, avCS + a fost evaluat ca fiind mai valențiat negativ comparativ cu CS- [Acq1: t(31) = 2.34, p = 0.026; Acq2: t(31) = 3.07, p = 0.004], NOUL [Acq1: t(31) = 2.70, p = 0.011; Acq2: t(31) = 3.89, p <0.001] și aplicația CS + [Acq1: t(31) = 5.41, p <0.001; Acq2: t(31) = 6.11, p <0.001]. AppCS + a fost evaluat ca semnificativ mai pozitiv decât CS- [Acq1: t(31) = 4.99, p <0.001; Acq2: t(31) = 5.31, p <0.001] și NOUUL [Acq1: t(31) = 4.92, p <0.001; Acq2: t(31) = 4.14, p <0.001]. Diferențele dintre CS– și NOU nu au fost niciodată semnificative (ps> 0.18).
Este important faptul că nu s-au găsit diferențe semnificative atunci când se compară valorile valențelor pentru appCS + asociate cu ciocolată și covrigei sărată [Acq1: t(30) = 0.03, p = 0.477; Acq2: t(30) = 0.29, p = 0.775].
După faza de extincție, avCS + a fost încă evaluat ca fiind mai negativ în comparație cu CS- [t(31) = 2.40, p = 0.023], modelul NEW [t(31) = 3.40, p = 0.002] și aplicațiile appCS + [t(31) = 3.35, p = 0.002]. Dimpotrivă, valența aplicației appCS + nu s-a mai diferențiat de CS- [t(31) = 1.77, p = 0.086] sau NEW [t(31) = 1.07, p = 0.293].
ANOVA pentru evaluări ale excitării în timpul achiziției a relevat un efect principal semnificativ al stimulului [F(3, 96) = 7.07, GG-ε = 0.737, p = 0.001, parțial η2 = 0.181], dar nu din faza [F(3, 96) = 1.27, GG-ε = 0.805, p = 0.289, parțial η2 = 0.038] și o interacțiune semnificativă între stimul și faza [F(9, 288) = 4.53, GG-ε = 0.582, p = 0.001, parțial η2 = 0.124]. Urmare t- testele nu au indicat diferențe semnificative între stimulii privind excitația inițială (ps> 0.74). Cu toate acestea, după cele două faze de achiziție, avCS + a fost evaluat mai excitant decât NOUUL [Acq1: t(32) = 2.99, p = 0.005; Acq2: t(32) = 5.97, p <0.001] și aplicația CS + [Acq1: t(32) = 2.62, p = 0.013; Acq2: t(32) = 4.42, p <0.001] și marginal mai excitant decât CS - după prima fază de achiziție [t(32) = 1.96, p = 0.058], dar în mod semnificativ mai excitant după a doua fază de achiziție [t(32) = 3.65, p = 0.001]. Spre deosebire de evaluările de valență, excitarea aplicației appCS + nu a fost diferită de excitarea CS și NOU (ps > 0.13) după fazele de achiziție.
La fel ca și în cazul ratingurilor de valență, CS și NEW nu diferă în ceea ce privește excitarea lor (ps> 0.07).
Așa cum a fost cazul valenței, nu s-au evidențiat diferențe pentru appCS + asociat cu ciocolata și appCS + asociate cu covrigul sărat [Acq1: t(31) = 0.26, p = 0.797; Acq2: t(31) = 0.33, p = 0.724].
După faza de extincție, nu s-au evidențiat diferențe semnificative în evaluările de excitație (ps> 0.08).
Pentru mai multe post-hoc t- teste care compară evaluările după faza de obișnuință, prima și cea de-a doua fază de achiziție și faza de extincție, vezi Materialele Suplimentare.
Răspunsul inițial
ANOVA pentru fazele de achiziție a returnat un efect principal al stimulului [F(2, 64) = 49.92, GG-ε = 0.964, p <0.001, parțial η2 = 0.609], dar nu faza [F(1, 32) = 3.16, p = 0.085, parțial η2 = 0.090] și o interacțiune semnificativă Stimulus × Fază [F(2, 64) = 3.37, GG-ε = 0.875, p = 0.048, parțial η2 = 0.095]. Urmare t- testele au arătat o potențială semnificativă de apariție a avCS + în comparație cu CS - atât în timpul primei [t(32) = 3.27, p = 0.003] și al doilea [t(32) = 4.00, p <0.001] faze de achiziție. Răspunsurile inițiale la avCS + au fost, de asemenea, potențiate semnificativ în comparație cu cele la appCS +, din nou după ambele achiziții 1t(32) = 8.20, p <0.001] și achiziția 2 [t(32) = 5.74, p <0.001]. Foarte important, magnitudinea tresăririi față de appCS + a fost semnificativ atenuată în comparație cu CS - atât în timpul achiziției 1 [t(32) = 6.34, p <0.001] și achiziția 2 [t(32) = 2.91, p = 0.007]. Din nou și în concordanță cu evaluările, nu s-au dezvăluit răspunsuri de alarmă pentru appCS + atunci când sunt asociate cu ciocolata sau cu covrigul sărat [Acq1: t(31) = 1.04, p = 0.309; Acq2: t(31) = 0.07, p = 0.947]. În timpul fazei de extincție, nu sa constatat un efect semnificativ [F(3, 96) = 0.26, GG-ε = 0.906, p = 0.833, parțial η2 = 0.008].
Reacția de conduită a pielii (SCR)
Din ANOVA pentru SCR în timpul celor două faze de achiziție, stimulul principal al efectelor [F(2, 54) = 18.04, GG-ε = 0.908, p <0.001, parțial η2 = 0.401] și faza [F(1, 27) = 20.91, p <0.001, parțial η2 = 0.436] s-au dovedit a fi semnificative, dar nu interacțiunea lor [F(2, 54) = 0.68, GG-ε = 0.637, p = 0.451, parțial η2 = 0.024]. Post-hoc t-testurile au indicat SCR semnificativ mai mare pentru avCS + [t(27) = 6.46, p <0.001] și la aplicația CS + [t(27) = 4.84, p <0.001] comparativ cu CS-, în timp ce participanții au prezentat SCR comparabil cu avCS + și appCS + [t(27) = 0.64, p = 0.527]. În mod deosebit, nu au fost găsite diferențe între SCR-urile pentru appCS + ciocolată și pentru apps-ul covrigei sărate (appCS +) [Acq1: t(26) = 2.55, p = 0.120; Acq2: t(26) = 1.29, p = 0.210]. Așa cum sa întâmplat și în ceea ce privește evaluările și răspunsul surprinzător, nu s-au constatat efecte semnificative pentru faza de extincție [F(3, 81) = 0.28, GG-ε = 0.634, p = 0.743, parțial η2 = 0.010].
Discuție
Scopul acestui studiu a fost de a traduce concluziile animalelor la om prin folosirea unei paradigme clasice de condiționare a apetitului cu un prim întăritor ca stimul necondiționat (SUA). Pentru a face acest lucru, participanții au venit la laborator devreme dimineața, fără micul dejun, pentru a se asigura că ei erau înfometați și, în funcție de preferința lor, bucăți de ciocolată sau covrigi sărate au fost folosite ca apetisante din SUA. În timpul fazei de achiziție, o formă geometrică (avCS +) a devenit asociată cu un șoc electric ușor dureros (SUA aversiv), cu o altă formă (appCS +) cu apetitul american și cu o a treia formă (CS) aversiv SUA. Rezultatele indică condiționarea cu succes aversivă și apetită la nivelul verbal explicit (adică ratingurile), la nivelul implicit al comportamentului (adică răspunsul surprinzător) și la nivelul fiziologic (adică SCR). În mod specific, analiza avCS + comparativ cu CS-a obținut mai multe valori negative de valență, un grad mai ridicat de excitație, o potențare de spargere și o SCR mai mare. Cel mai important, aplicația appCS + în comparație cu CS-a declanșat ratinguri de valență mai pozitive, atenuare de șoc și o SCR mai mare. Constatările noastre privind condiționarea aversivă au fost în concordanță cu așteptările, deoarece studiile anterioare au constatat că un stimul care prezice amenințarea (avCS +) este evaluat ca aversiv, provoacă răspunsuri mai mari la frică și crește excitatia fiziologicăFendt și Fanselow, 1999; Hamm și Weike, 2005; Andreatta și colab., 2010, 2013). Rezultatele noastre privind condiționarea apetitului au fost, de asemenea, în concordanță cu studiile anterioare pe animale și pe animale, care arată că un stimulent care prezice recompensa (appCS +) este evaluat ca pozitiv, inhibă răspunsurile la frică și crește excitația fiziologicăKoch și colab., 1996; Gottfried și colab., 2002; Kumar și colab., 2008; Klucken și colab., 2009, 2013; Austin și Duka, 2010; Prévost și colab., 2012). Din cunoștințele noastre, acesta este primul studiu care demonstrează atenuarea condiționată condiționată la om la un stimul care prezice o recompensă primară. Foarte important, am reușit să transferăm și să confirmăm rezultatele unui studiu pe animale (Koch și colab., 1996). Acest studiu pe animale a demonstrat că atenuarea de șoc la șobolani depinde de proiecțiile de la NAcc (parte a striatumului ventral) la PnC. Prin urmare, atenuarea stricătoare în studiul nostru ar putea implica activitate NAcc, care este, de asemenea, în conformitate cu constatările fMRI (Gottfried și colab., 2002; Kumar și colab., 2008; Klucken și colab., 2009, 2013; Delgado și colab., 2011; Levy și Glimcher, 2011). Prin urmare, concluzionăm că paradigma noastră de aranjare apetitoare a avut succes, deoarece au indicat atât valența explicită (evaluarea), cât și implicit (atenuarea stricăciunii) pozitivă.
În plus față de această constatare nouă, dar mai degrabă previzibilă, merită menționat încă două rezultate interesante. Mai întâi, răspunsurile de excitare verbală și fiziologică la appCS + disociate. În al doilea rând, am descoperit o dispariție mai rapidă a apetitului CR comparativ cu CR-urile aversive.
SCR indică activarea simpatică, care este mărită atât la stimulul asociat cu recompensa (appCS +), cât și la stimulul asociat amenințării (avCS +). Acest rezultat este în concordanță cu un studiu de condiționare anterioară, în care imaginile erotice au fost folosite ca apetisante din SUA (Klucken și colab., 2013), precum și un studiu care relevă o SCR comparabilă cu o estimare a banilor și o estimare a zgomotului aversiv (Austin și Duka, 2010). În mod special, SCR este un răspuns orientativ referitor la activarea sistemului simpatic. Acest răspuns a fost sugerat pentru a reflecta pregătirea unei reacții comportamentale la evenimente importante (Bradley, 2009). Pe baza acestor considerente, credem că atât stimulul asociat cu amenințarea, cât și stimulul asociat recompensei au generat un răspuns pregătitor pentru răspunsurile comportamentale succesive. Cu alte cuvinte, semnalele de alimente și durere au inițiat pregătirea pentru comportamentul de abordare și evitare, respectiv. Spre deosebire de excitația fiziologică ridicată, răspunsurile verbale au indicat o excitare scăzută la stimulul asociat recompenselor. Este posibil ca excitația verbală să fie mai mult influențată de natura excitantă a SUA decât de activarea fiziologică în sine. De fapt, considerăm că aplicațiile appCS + vs. avCS + au fost evaluate ca fiind scăzute și foarte excitante, deoarece acestea au fost legate de niște americani cu nivel scăzut și extrem de excitant. Din păcate, nu am colectat ratinguri de excitație pentru SUA și, prin urmare, va fi vorba de studiile viitoare pentru a testa această ipoteză în mod explicit.
În timpul fazei de extincție, nu a fost livrată nicio țară americană. Acest lucru poate să fi indus o nouă învățătură inhibitoare numită extincție (pentru o revizuire a se vedea Milad și Quirk, 2012), atât în ceea ce privește CR-urile aversive, cât și cele apetisante, ca o consecință. Evident, în ceea ce privește răspunsurile comportamentale (cum ar fi răspunsul la surprindere) și pe nivelurile fiziologice (SCR), nu mai puteau fi detectate reacții diferențiale la avCS +, appCS + și CS. În mod similar, excitarea explicită a stimulilor condiționați evaluați după faza de extincție a egalat la un nivel scăzut, sugerând o învățare reușită de dispariție. Totuși, avCS + a fost încă evaluat semnificativ mai negativ decât CS-, în timp ce vC-ul appCS + nu mai diferă de valența CS-ului. O dispariție mai lentă a răspunsului explicit aversiv ar putea fi datorată unui conservatorism evolutiv, ceea ce înseamnă că semnalele de amenințare sunt deosebit de greu de uitat deoarece un non-răspuns la un semnal de amenințare ar putea pune viața în pericol.
O întrebare încă mai rămâne: de ce a stins răspunsul surprinzător (adică valența implicită) în timpul fazei de dispariție? În primul rând, aceste răspunsuri au fost calculate pe întreaga durată a fazei. Prin urmare, este posibil ca CR-urile discriminatorii să fie încă detectabile în primele încercări ale fazei de extincție. Într-o manieră exploratorie, am urmat această ipoteză și ne-am uitat atât la răspunsurile surprinzătoare, cât și la SCR în timpul fazei de extincție (a se vedea Materialul suplimentar). Deși nu am găsit diferențe semnificative, am observat o magnitudine de surprindere puțin mai ridicată pentru avCS + în comparație cu CS și noul stimulent de control. De asemenea, am observat o atenuare puțin mai puternică a aplicației appCS + în comparație cu CS-ul și cu noul stimulent de control la începutul fazei de extincție, care totuși a dispărut în câteva studii. SCR la aplicația appCS + a scăzut deja după încercarea de extincție 2nd, în timp ce SCR la avCS + a rămas mai mare pentru aproape toate extincțiile, comparativ cu SCR cu NOUL. Deși ar trebui să interpretăm aceste rezultate cu mare prudență, răspunsurile surprinzătoare par paralel cu evaluările valenței. În plus, ambele răspunsuri surprinzătoare și SCR-urile susțin în continuare ideea unui conservatorism evolutiv față de stimulii de amenințare.
În cele din urmă, trebuie să recunoaștem unele limitări ale acestui studiu. În primul rând, din cauza unei probleme tehnice nu am putut raporta ratinguri de plăcere (și excitare) pentru ciocolată și covrigei sărate. Cu toate acestea, apetitul CR sugerează că participanții au experimentat cu adevărat cele două SUA ca apetisante. În al doilea rând, durata aversiunii și apetitului american a fost foarte diferită. Astfel, șocul electric dureros a fost livrat la offsetul avCS + și a durat exact 200 ms, în timp ce ciocolata și covriful sărat au fost prezentate participanților circa 2 s după instalarea appCS +, iar durata a fost nedefinabilă deoarece depindea de cât de repede individul le-a mâncat. Alegerea de a furniza US apetitul în acest mod sa bazat pe un studiu anterior la om (Prévost și colab., 2012). Cu toate acestea, ar fi mult mai elegant din punct de vedere metodologic să furnizăm o apetită americană mai asemănătoare cu SUA aversivă, cum ar fi o gustare de suc sau de ghimbir la apCS + offset. Pentru a compensa această mare diferență perceptuală, am prezentat forme geometrice împreună cu un stimul vizual simbolizând Statele Unite. În al treilea rând, nu putem exclude cu siguranță posibilitatea ca răspunsurile fiziologice rapide stinse în faza de extincție să se datoreze unui aspect metodologic. De fapt, stimulii vizuale în această fază nu au fost prezentați împreună cu simbolul SUA, așa cum a fost cazul în fazele de achiziție. Este posibil ca prezentarea CS în monoterapie să fi influențat CR și, din acest motiv, nu au fost observate diferențe semnificative în timpul primelor încercări de extincție. Cu toate acestea, răspunsurile noastre stingate rapid sunt în concordanță cu răspunsurile stinse într-un studiu anterior, în care CS + (față) a fost prezentat în combinație cu SUA (țipa) în timpul achiziției, dar nu în timpul dispariției (Lissek și colab., 2008).
În concluzie, am găsit reacții aversive și apetisante condiționate cu succes la un stimul asociat cu amenințarea și un stimulent asociat, respectiv, cu recompensa. Interesant este faptul că nivelul explicit al răspunsurilor (implicit) și răspunsul implicit (reflex de startle) au acționat într-o manieră sinergică, deoarece avCS + a fost raportat ca potențare negativă și indusă, iar appCS + a fost raportată ca atenuare pozitivă și inductivă. În plus, excitarea explicită (evaluări) și fiziologică (SCR) a appCS + disociată, reflectând două procese distincte.
Declarația privind conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
recunoasteri
Această lucrare a fost susținută de Centrul de cercetare colaborativă "Frica, anxietate și tulburări de anxietate", SFB-TRR 58, proiectul B1.
Material suplimentar
Materialul suplimentar pentru acest articol poate fi găsit online la adresa: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
Referinte
Andreatta, M., Mühlberger, A., Glotzbach-Schoon, E. și Pauli, P. (2013). Previzibilitatea durerii inversează evaluările de valență ale stimulului asociat reliefului. Față. Syst. Neurosci. 7: 53. doi: 10.3389 / fnsys.2013.00053
PubMed Rezumat | CrossRef Full Text | Google Academic
Andreatta, M., Mühlberger, A., Yarali, A., Gerber, B. și Pauli, P. (2010). O ruptură între valența implicită și explicită, după învățarea durerii-relief la om. Proc. Biol. Sci. 277, 2411-2416. doi: 10.1098 / rspb.2010.0103
PubMed Rezumat | CrossRef Full Text | Google Academic
Austin, AJ și Duka, T. (2010). Mecanisme de atenție pentru apetitul și rezultatele aversive în condiționarea Pavlovian. Behav. Brain Res. 213, 19-26. doi: 10.1016 / j.bbr.2010.04.019
PubMed Rezumat | CrossRef Full Text | Google Academic
Blumenthal, TD, Cuthbert, BN, Filion, DL, Hackley, S., Lipp, OV și van Boxtel, A. (2005). Raportul comitetului: orientări pentru studiile electromiografice cu ochi bruiați. Psihofiziologie 42, 1-15. doi: 10.1111 / j.1469-8986.2005.00271.x
PubMed Rezumat | CrossRef Full Text | Google Academic
Bouton, ME și Peck, CA (1989). Efecte de context asupra condiționării, extincției și reintegrării într-un preparat de condiționare a apetitului. Anim. Învăța. Behav. 17, 188-198. doi: 10.3758 / BF03207634
CrossRef Full Text | Google Academic
Bradley, MM (2009). Atenție selectivă naturală: orientare și emoție. Psihofiziologie 46, 1-11. doi: 10.1111 / j.1469-8986.2008.00702.x
PubMed Rezumat | CrossRef Full Text | Google Academic
Clark, JJ, Hollon, NG și Phillips, PEM (2012). Sisteme de evaluare Pavlovian în procesul de învățare și de luare a deciziilor. Curr. Opin. Neurobial. 22, 1054-1061. doi: 10.1016 / j.conb.2012.06.004
PubMed Rezumat | CrossRef Full Text | Google Academic
Delgado, MR, Jou, RL și Phelps, EA (2011). Sistemele neuronale care stau la baza condiționării aversive la om cu elemente de întărire primare și secundare. Față. Neurosci. 5: 71. doi: 10.3389 / fnins.2011.00071
PubMed Rezumat | CrossRef Full Text | Google Academic
Dickinson, A. și Balleine, B. (1994). Controlul motivațional al acțiunii direcționate spre obiectiv. Anim. Învăța. Behav. 22, 1-18. doi: 10.3758 / BF03199951
CrossRef Full Text | Google Academic
Fendt, M., și Fanselow, MS (1999). Baza neuroanatomică și neurochimică a fricii condiționate. Neurosci. Biobehav. Rev. 23, 743-760. doi: 10.1016 / j.ijpsycho.2012.09.006
PubMed Rezumat | CrossRef Full Text | Google Academic
Gottfried, JA, O'Doherty, J. și Dolan, RJ (2002). Învățarea olfactivă apetitivă și aversivă la oameni a studiat utilizând imagistica prin rezonanță magnetică funcțională legată de evenimente. J. Neurosci. 22, 10829-10837. Disponibil online la: http://www.jneurosci.org/content/22/24/10829.full
Grillon, C., Baas, JM, Cornwell, B. și Johnson, L. (2006). Condiționarea condițiilor și evitarea comportamentului într-un mediu al realității virtuale: efectul previzibilității. Biol. Psihiatrie 60, 752-759. doi: 10.1016 / j.biopsych.2006.03.072
PubMed Rezumat | CrossRef Full Text | Google Academic
Hamm, AO și Weike, AI (2005). Neuropsihologia învățării fricii și a reglementării fricii. Int. J. Psychophysiol. 57, 5-14. doi: 10.1016 / j.ijpsycho.2005.01.006
PubMed Rezumat | CrossRef Full Text | Google Academic
Klucken, T., Schweckendiek, J., Merz, CJ, Tabbert, K., Walter, B., Kagerer, S., și colab. (2009). Activarea neuronală a dobândirii excitării sexuale condiționate: efectele conștiinței de urgență și a sexului. J. Sex. Med. 6, 3071-3085. doi: 10.1111 / j.1743-6109.2009.01405.x
PubMed Rezumat | CrossRef Full Text | Google Academic
Klucken, T., Wehrum, S., Schweckendiek, J., Merz, CJ, Hennig, J., Vaitl, D. și colab. (2013). Polimorfismul 5-HTTLPR este asociat cu răspunsuri hemodinamice modificate în timpul condiționării apetitului. Zumzet. Brain Mapp. 34, 2549-2560. doi: 10.1002 / hbm.22085
PubMed Rezumat | CrossRef Full Text | Google Academic
Koch, M. (1999). Neurobiologia uimii. Prog. Neurobial. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
PubMed Rezumat | CrossRef Full Text | Google Academic
Koch, M., Schmid, A. și Schnitzler, H.-U. (1996). Atenuarea plăcerii stupefiantului este perturbată de leziunile nucleului accumbens. Neuroreport 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
PubMed Rezumat | CrossRef Full Text | Google Academic
Krohne, HW, Egloff, B., Kohmann, C.-W. și Tausch, A. (1996). Untersuchungen mit einer deutschen version der "Program pozitiv și negativ afectat" (PANAS). Diagnostics 42, 139-156.
Kumar, P., Waiter, G., Ahearn, T., Milders, M., Reid, I. și Steele, JD (2008). Distorsiuni temporale anormale ale diferențelor de învățare în depresie majoră. Creier 131, 2084-2093. doi: 10.1093 / creier / awn136
PubMed Rezumat | CrossRef Full Text | Google Academic
Laux, L., Glanzmann, P., Schaffner, P. și Spielberger, CD (1981). Das State-Trait Angstinventar. Weinheim: testul Beltz.
Levy, DJ și Glimcher, PW (2011). Compararea merelor și a portocalelor: folosirea unei reprezentări de valoare subiectivă și recompensă-valoare generală subiectivă în creier. J. Neurosci. 31, 14693-14707. doi: 10.1523 / JNEUROSCI.2218-11.2011
PubMed Rezumat | CrossRef Full Text | Google Academic
Lissek, S., Levenson, J., Biggs, AL, Johnson, LL, Ameli, R., Pine, DS și colab. (2008). Cresterea fricii crescute a stimulilor neconditionat relevanti din punct de vedere social in tulburarea de anxietate sociala. A.m. J. Psychiatry 165, 124-132. doi: 10.1176 / appi.ajp.2007.06091513
PubMed Rezumat | CrossRef Full Text | Google Academic
Martin-Soelch, C., Linthicum, J. și Ernst, M. (2007). Reglarea apetitului: baze neuronale și implicații pentru psihopatologie. Neurosci. Biobehav. Rev. 31, 426-440. doi: 10.1016 / j.neubiorev.2006.11.002
PubMed Rezumat | CrossRef Full Text | Google Academic
McDannald, MA, Lucantonio, F., Burke, KA, Niv, Y., și Schoenbaum, G. (2011). Striatura ventrală și cortexul orbitofrontal sunt ambele necesare pentru învățarea de întărire bazată pe model, dar fără model. J. Neurosci. 31, 2700-2705. doi: 10.1523 / JNEUROSCI.5499-10.2011
PubMed Rezumat | CrossRef Full Text | Google Academic
Milad, MR, și Quirk, G. (2012). Frica de dispariție ca model pentru neuroștiința translațională: zece ani de progres. Annu. Rev. Psychol. 63, 129-111. doi: 10.1146 / anurev.psych.121208.131631
PubMed Rezumat | CrossRef Full Text | Google Academic
Pavlov, IP (1927). Reflexe condiționate: o investigație a activității fiziologice a cortexului cerebral. Londra: Oxford University Press.
Prévost, C., Liljeholm, M., Tyszka, JM și O'Doherty, JP (2012). Corelate neuronale ale transferului Pavlovian-instrumental specific și general în subregiunile amigdalare umane: un studiu fMRI de înaltă rezoluție. J. Neurosci. 32, 8383-8390. doi: 10.1523 / jneurosci.6237-11.2012
PubMed Rezumat | CrossRef Full Text | Google Academic
Tranel, D. și Damasio, H. (1994). Corelațiile neuroanatomice ale răspunsurilor de conductivitate ale pielii electrodermice. Psihofiziologie 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
PubMed Rezumat | CrossRef Full Text | Google Academic
Cuvinte cheie: condiționarea clasică, recompensă, pedeapsă, reflex de suspans, răspunsul la conductivitatea pielii
Referință: Andreatta M și Pauli P (2015) Apetisant vs. Avertisment condiționat la om. Față. Behav. Neurosci. 9: 128. doi: 10.3389 / fnbeh.2015.00128
Primit: 11 Februarie 2015; Acceptat: 05 Mai 2015;
Publicat: 19 Mai 2015.
Editat de:
Niels Birbaumer, Universitatea din Tuebingen, Germania
Revizuite de:
Tim Klucken, Universitatea Justus Liebig Giessen, Germania
Alessandro Angrilli, Universitatea din Padova, Italia
Copyright © 2015 Andreatta și Pauli. Acesta este un articol cu acces deschis, distribuit în termenii lui Creative Commons Attribution License (CC BY). Utilizarea, distribuirea sau reproducerea în alte forumuri este permisă, cu condiția ca autorii sau licențiatorii originali să fie creditați și să fie citată publicația originală din acest jurnal, în conformitate cu practica academică acceptată. Nu este permisă utilizarea, distribuirea sau reproducerea care nu respectă acești termeni.
* Corespondență: Marta Andreatta, Departamentul de Psihologie (Psihologie Biologică, Psihologie Clinică și Psihoterapie), Universitatea din Würzburg, Marcusstraße 9-11, D-97070 Würzburg, Germania, [e-mail protejat]
