Față. Comportament. Neurosci., 23 martie 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2,
Masroor R. Shariff1, Arnauld Belmer1, Matthew J. Fogarty2, Erica WH Mu2, 

- 1Institutul de Cercetare Translațională și Institutul pentru Sănătate și Inovare Biomedicală, Queensland University of Technology, Brisbane, QLD, Australia
- 2Școala de Științe Biomedicale, Universitatea Queensland, Brisbane, QLD, Australia
Dieta modernă a devenit foarte îndulcită, ducând la niveluri fără precedent de consum de zahăr, în special în rândul adolescenților. În timp ce se știe că aportul cronic de zahăr pe termen lung contribuie la dezvoltarea tulburărilor metabolice, inclusiv obezitatea și diabetul de tip II, se știe puține cu privire la consecințele directe ale consumului de zahăr pe termen lung, asemănător binge-ului, asupra creierului. BDeoarece zahărul poate provoca eliberarea de dopamină în nucleul accumbens (NAc) în mod similar cu drogurile de abuz, am investigat modificările morfologiei neuronilor din această regiune a creierului în urma unei abundențe pe termen scurt (4 săptămâni) și pe termen lung (12 săptămâni). cum ar fi consumul de zaharoză folosind o paradigmă intermitentă de alegere a două sticle. Am folosit colorarea Golgi-Cox pentru a impregna neuronii spinoși medii (MSN) din miezul NAc și coaja de șobolani consumatoare de zaharoză pe termen scurt și lung și i-am comparat cu controalele de apă potrivite în funcție de vârstă. Arătăm că consumul prelungit de zaharoză a scăzut semnificativ lungimea dendritică totală a MSN-urilor cu coajă NAc în comparație cu șobolanii de control potriviți în funcție de vârstă. De asemenea, am constatat că restructurarea acestor neuroni a rezultat în principal din complexitatea dendritică distală redusă. Dimpotrivă, am observat densități crescute ale coloanei vertebrale la ordinele de ramuri distale ale MSN-urilor cu coajă NAc de la șobolanii consumatoare de zaharoză pe termen lung. Combinate, aceste rezultate evidențiază efectele neuronale ale aportului prelungit de zaharoză, asemănător unui binge, asupra morfologiei MSN a cochiliei NAc.
Introducere
În ultimii 40 de ani, a existat o creștere documentată a consumului de băuturi îndulcite cu zahăr și de alimente care conțin zaharuri adăugate (Nielsen și colab., 2002; Popkin, 2010; Ng și colab., 2012), cu rapoarte care estimează că până la 75% din toate alimentele și băuturile conțin cantități mari de zaharuri adăugate (Ford și Dietz, 2013; Bray și Popkin, 2014). În această perioadă, a existat și o creștere concomitentă a prevalenței obezității și a diabetului de tip II, în special în rândul adolescenților (Arslanian, 2002; Reinehr, 2013; Dabelea et al., 2014; Fryar și colab., 2014). Studii recente au arătat că copiii supraponderali și obezi consumă adesea cantități mari de zahăr adăugat, totuși contribuția dietelor cu conținut ridicat de zahăr la creșterea incidenței copiilor supraponderali și obezi rămâne controversată (Hu, 2013; Bray și Popkin, 2014; Bucher Della Torre și colab., 2015).
În timp ce un număr tot mai mare de dovezi indică faptul că consumul de diete bogate în zahăr poate contribui, parțial, la creșterea în greutate în rândul copiilor și adolescenților (Malik și colab., 2010; Te Morenga et al., 2013; Bray și Popkin, 2014), s-a acordat mai puțină atenție consecințelor nemetabolice adverse care decurg din consumul excesiv de zahăr. Interesant este că unele modele comportamentale și psihologice comune apar adesea în rândul unui subgrup de cei care mănâncă în exces și mențin diete bogate în zahăr. Cele mai notabile sunt dezvoltarea tulburărilor de alimentație, inclusiv alimentația excesivă, combinată cu apariția concomitentă a simptomelor psihologice, inclusiv lipsa de motivație și depresia (revizuită în Sheehan și Herman, 2015). În plus, deoarece persoanele care mănâncă în exces manifestă adesea o pierdere a controlului și o incapacitate de a-și auto-limita aportul de zahăr, este probabil ca aceste comportamente să apară ca urmare a adaptărilor neurologice în regiunile creierului care evaluează valoarea hedonică a alimentelor foarte gustoase. (Saper și colab., 2002; Lutter și Nestler, 2009; Kenny, 2011). Această rațiune este susținută și de dovezi la oameni care demonstrează că zahărul și dulceața pot provoca pofte similare cu cele induse de drogurile care creează dependență, cum ar fi alcoolul și nicotina (Volkow și colab., 2012).
Deși proprietățile de dependență ale zahărului sunt încă speculative, aceste observații s-au combinat cu studii care demonstrează contribuția consumului excesiv de zahăr la modificările circuitelor de recompensă și la dezvoltarea unor comportamente și stări emoționale asemănătoare dependenței la modelele animale. (Avena și colab., 2008; Benton, 2010; Ventura și colab., 2014), justifică necesitatea unei investigații suplimentare. Studii anterioare la rozătoare au arătat că accesul intermitent la zaharoză modifică activitatea mai multor neurotransmițători din sistemul mezolimbic, inclusiv dopamina, opioidele și acetilcolina (revizuit în Avena și colab., 2008). S-a demonstrat că consumul de zaharoză în mod similar facilitează eliberarea de dopamină în nucleul accumbens (NAc), în mod similar cu drogurile de abuz (Avena și colab., 2008). În plus, am arătat că consumul pe termen lung de zaharoză utilizând o paradigmă de alegere a două sticle cu acces intermitent de 24 de ore (Simms et al., 2008) modulează expresia receptorului nicotinic de acetilcolină (nAChR) în NAc (Shariff et al., în presă). Interesant, am observat, de asemenea, că compușii nAChR cunoscuți că modulează activitatea dopaminei și acetilcolinei în NAc, au efecte diferite asupra consumului de zaharoză după aportul pe termen scurt și lung (Shariff et al., în presă).
Deși aceste studii au demonstrat asemănări în ceea ce privește modificările comportamentale și neurochimice cauzate de accesul intermitent la zahăr și droguri de abuz, nu se știe dacă aceste efecte facilitează modificările morfologiei neuronale în NAc. Teste în contrast cu substanțele de abuz, inclusiv cocaina, amfetamina și nicotina, care produc modificări bine caracterizate în morfologia neuronilor spinoși medii (MSN) în NAc, inclusiv densitatea crescută a coloanei vertebrale și complexitatea dendritică modificată. (Robinson și Kolb, 1999, 2004; Li și colab., 2003; Crombag și colab., 2005). Deoarece am arătat anterior că expunerea pe termen lung (12 săptămâni) la alcool și zaharoză folosind paradigma de alegere intermitentă a două sticle produce un răspuns diferențial la intervențiile farmacoterapeutice în comparație cu aportul pe termen scurt (4 săptămâni; Steensland și colab., 2007; Shariff et al., în presă), am evaluat efectele consumului de zaharoză pe termen scurt și lung asupra morfologiei MSN în NAc. Am permis șobolanilor adolescenți să consume zaharoză într-un mod asemănător cu binge timp de 4 (pe termen scurt) sau 12 (pe termen lung) săptămâni și apoi am analizat morfologia MSN-urilor NAc de la șobolanii care consumă zaharoză pe termen scurt și lung și am comparat aceasta cu controale potrivite în funcție de vârstă cărora li sa dat acces numai la apă. Rezultatele noastre arată că MSN-urile din învelișul NAc sunt modificate în urma consumului de zaharoză pe termen lung, dar nu pe termen scurt, având lungimea dendritică redusă, dar densitatea coloanei dendritice distale crescute. În plus, am constatat că morfologia MSN-urilor din miezul NAc a rămas relativ intactă în urma consumului de zaharoză pe termen scurt și lung. Aceste rezultate evidențiază o consecință neurologică directă a consumului de zaharoză pe termen lung într-o manieră asemănătoare. În plus, aceste date demonstrează necesitatea unor studii ulterioare care să vizeze elucidarea modificărilor moleculare și neurochimice care însoțesc restructurarea morfologică a MSN-urilor de coajă NAc indusă de aportul prelungit de zaharoză, asemănător binge.
Materiale și metode
Declarație de etică
Toate procedurile experimentale au fost efectuate în conformitate cu Codul australian pentru îngrijirea și utilizarea animalelor în scopuri științifice, ediția a 8-a (National Health and Medical Research Council, 2013). Protocoalele au fost aprobate de Comitetul de etică a animalelor de la Universitatea de Tehnologie din Queensland și de Comitetul de etică a animalelor de la Universitatea din Queensland.
Animale și locuințe
Șobolani Wistar masculi (adolescenti) în vârstă de cinci săptămâni (control: 176.4 ± 4.8 g; zaharoză: 178.3 ± 5.0 g) (ARC, WA, Australia), au fost găzduiți individual în plexiglas ventilat cu două niveluri® cuști. Șobolanii au fost aclimatizați la condițiile individuale de adăpostire, manipulare și ciclu de lumină inversă cu 5 zile înainte de începerea experimentelor. Toți șobolanii au fost găzduiți într-o cameră cu climă controlată de 12 ore inversă lumină/întuneric (luminile stinse la ora 9 dimineața), cu mâncare standard pentru șobolani și apă disponibilă ad libitum.
Paradigma de băutură cu acces intermitent în două sticle
Accesul intermitent 5% zaharoză în două sticle paradigma de băut la alegere (Simms et al., 2008) a fost adaptat din Înțelept (1973). Toate fluidele au fost prezentate în sticle de plastic gradate de 300 ml cu duze de băut din oțel inoxidabil introduse prin două coloane din partea din față a cuștii după începerea ciclului de lumină întunecată. Greutățile fiecărei sticle au fost înregistrate înainte de prezentarea sticlei. Au fost prezentate simultan două sticle: o sticlă cu apă; a doua sticlă conţinând 5% (g/v) zaharoză. Amplasarea sticlei de zaharoză 5% (g/v) a fost alternată cu fiecare expunere la control pentru preferințele secundare. Sticlele au fost cântărite la 24 de ore după ce fluidele au fost prezentate, iar măsurătorile au fost efectuate cu cel mai apropiat 0.1 g. Greutatea fiecărui șobolan a fost, de asemenea, măsurată pentru a calcula gramele de aport de zaharoză per kilogram de greutate corporală. În ziua 1 a perioadei de băut, șobolanii (n = 6–9) au primit acces la o sticlă de zaharoză 5% (g/v) și o sticlă de apă. După 24 de ore, sticla de zaharoză a fost înlocuită cu o a doua sticlă de apă care a fost disponibilă pentru următoarele 24 de ore. Acest model se repeta miercurea și vineri. Sobolanii au avut acces nelimitat la apa in toate celelalte zile. Consumul de zaharoză asemănător cu binge a dus la o escaladare a aportului total de zaharoză (ml) în timp (Figura suplimentară 1) și a fost însoțită de niveluri inițiale stabile de băutură bazate pe greutatea corporală [20 ± 5 g/kg din 5% (g/v)] pe termen scurt [~4 săptămâni (13 sesiuni de băut)] și pe termen lung [ ~12 săptămâni (37 sesiuni de băutură)] perioade de băut. Un grup separat de șobolani martor (n = 6–9) au primit acces la apă în ambele sticle (adică, fără zaharoză) în aceleași condiții descrise mai sus. Greutatea corporală medie a șobolanilor martor și consumatoare de zaharoză la sfârșitul expunerii pe termen scurt a fost de 405.7 ± 40.8 g și, respectiv, 426.4 ± 31.2 g. La sfârșitul expunerii pe termen lung, greutatea corporală medie pentru grupele martor și zaharoză a fost de 578.8 ± 53.4 g și 600.2 ± 45.2 g.
Colorație Golgi-Cox
După ultima sesiune de băut, șobolanii au fost transferați de la unitatea de animale pentru a permite procesarea probelor de creier la unitatea de histologie de la Școala de Științe Biomedicale, Universitatea din Queensland (Sf. Lucia, Australia). Au fost luate toate măsurile aprobate pentru a reduce stresul în timpul transportului, după care șobolanii au fost lăsați să-și revină peste noapte. A doua zi, șobolanii au fost sacrificați prin supradozaj cu pentobarbital de sodiu (60–80 mg/kg, ip Vetcare, Brisbane, Australia) și perfuzați intracardic cu ~300 ml lichid cefalorahidian artificial care conținea, (în mM): 130 NaCl, 3 KCI, 26 NaHCXNUMX3, 1.25 NaH2PO4, 5 MgCl2, 1 CaCl2și 10D-glucoză. Fiecare animal a fost apoi decapitat, iar creierul a fost îndepărtat și incubat la întuneric în soluție Golgi-Cox care conținea 5% dicromat de potasiu, 5% cromat de potasiu și 5% clorură de mercur (toate substanțele chimice de la Sigma-Aldrich) care a fost preparată proaspătă timp de 3 zile. înainte de sacrificiu așa cum a fost descris anterior (Rutledge și colab., 1969). Metodele de incubare și post-procesare a colorației Golgi-Cox au fost modificate din Ranjan și Mallick (2010). Creierele de la animalele consumatoare de zaharoză pe termen scurt au fost incubate timp de 6 zile la 37°C, în timp ce creierele de la animalele consumatoare de zaharoză pe termen lung au fost incubate timp de 10 zile, cu o schimbare la soluție proaspătă Golgi-Cox după 4 zile de incubare.
După incubare, secțiunile coronale de 300 μm au fost tăiate folosind un microtom vibrant Zeiss Hyrax V50 (Carl Zeiss, Germania). Feliile au fost apoi plasate secvenţial în plăci cu 24 de godeuri umplute cu zaharoză 30% (g/v) în soluţie salină tamponată cu fosfat 0.1 M şi procesate după cum este descris în (Ranjan și Mallick, 2010). Pe scurt, secțiunile au fost deshidratate în etanol 50% timp de 5 minute, apoi plasate în 0.1 M NH4Soluție de OH timp de 30 de minute, clătită de două ori cu apă distilată timp de 5 minute și plasată în fixator de film Fujihunt (Fujifilm, Singapore) timp de 30 de minute la întuneric. Feliile au fost apoi clătite de două ori în apă distilată timp de 2 minute fiecare și deshidratate în 70, 90, 95 și 100% etanol de două ori timp de 5 minute fiecare. Secțiunile au fost apoi curățate în soluție de CXA (1:1:1 cloroform:xilen:alcool) timp de 10 minute și montate în DPX (Sigma-Aldrich) pe lame Superfrost Plus (Menzel-Glaser, Lomb Scientific, Australia) și acoperite cu lame. (Menzel-Glaser, Germania). Lamelele au fost lăsate la întuneric să se usuce la temperatura camerei peste noapte.
Selecția neuronală și urmărirea în cadrul nucleului accumbens
Secțiunile coronale dintre bregma +2.8 și +1.7 au fost cercetate pentru MSN în miezul și învelișul NAc, folosind ventriculul lateral și comisura anterioară ca repere cu ajutorul unui atlas al creierului de șobolan (Paxinos și Watson, 2007) (Figura 1). Funcția de contur din Neurolucida 7 (MBF Bioscience, VT, SUA) a fost utilizată pentru a delimita miezul NAc și învelișul NAc în fiecare felie (Figura 2). Între 2 și 9 neuroni per regiune per animal au fost urmăriți pentru parametrii de lungime dendritică folosind un obiectiv de 63x sau pentru densitățile coloanei vertebrale (raportate ca țepi pe 100 μm) folosind un obiectiv de 100x pe un Zeiss Axioskop II (Carl Zeiss, Germania) folosind un dispozitiv automat xyz etapă condusă de Neurolucida® 7 software (MBF Biosciences, VT, SUA). Toate urmărirea au fost efectuate într-un mod orb în ceea ce privește tratamentul. Parametrii morfologici ai neuronilor impregnați cu Golgi-Cox au fost analizați într-un mod similar cu rapoartele anterioare (Klenowski și colab., 2015).
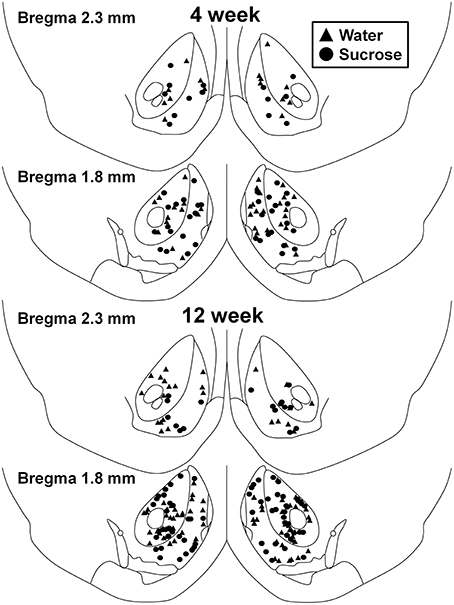
Figura 1. Hartă care arată locațiile neuronilor spinoși medii prelevați din miezul și învelișul nucleului accumbens ale șobolanilor care consumă zaharoză timp de 4 și 12 săptămâni și martori potriviți în funcție de vârstă. Cele două panouri de sus arată locațiile neuronilor prelevați din miezul nucleului accumbens și învelișul animalelor de control de 4 săptămâni (triunghiuri) și zaharoză (cercuri). Cele două panouri de jos arată pozițiile neuronilor prelevați de la animalele de control (triunghiuri) și zaharoză (cercuri) de 12 săptămâni.
Analiza statistică
Media și eroarea standard a mediei (SEM) au fost calculate pentru fiecare set de date cu animalul ca n, folosind datele de morfometrie medie de la toate MSN-urile NAc de bază sau de înveliș (n = 7 pentru învelișul NAc și n = 6 pentru miezul NAc de 4 săptămâni, n = 9 pentru grupuri de 12 săptămâni). Acolo unde este indicat, student neperecheat cu două cozi t-s-au efectuat teste sau ANOVA bidirecționale cu post-testele Bonferroni pentru toate analizele care implică compararea mediilor de grup, folosind GraphPad Prism versiunea 6.02 (GraphPad Software, San Diego, CA). Semnificația statistică a fost acceptată la P < 0.05. Toate datele din secțiunea cu rezultate sunt prezentate ca medii ± SEM. Modificările procentuale sunt calculate în raport cu valoarea de control.
REZULTATE
Neuronii spinoși medii din învelișul Nucleus Accumbens au o lungime dendritică scăzută, o complexitate dendritică scăzută, dar o densitate medie a coloanei vertebrale crescută la ordinele ramurilor distale în urma consumului de zaharoză pe termen lung, dar nu pe termen scurt.
În urma consumului de zaharoză pe termen scurt (4 săptămâni), nu au existat diferențe semnificative în parametrii morfometrici MSN înveliș NAc (Tabel 1). De asemenea, nu au existat diferențe semnificative între consumul de zaharoză pe termen scurt și MSN-urile de coajă NAc pentru controlul apei în analizele legate de ordinea ramurilor centrifuge. Și anume, segmentele dendritice pe ordin de ramificație (P = 0.4111), lungimea medie dendritică pe ordin de ramificare (P = 0.5581) și densitatea medie a coloanei vertebrale pe ordin de ramuri (P = 0.2977, ANOVA bidirecționale) nu au fost semnificativ diferite între grupuri. O hartă a locației care arată pozițiile aproximative ale neuronilor prelevați este prezentată în figură 1.
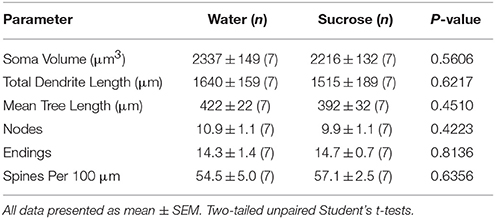
Tabelul 1. Parametrii morfologici generali ai neuronilor spinoși medii din învelișul nucleului accumbens al șobolanilor care consumă zaharoză pe termen scurt și controalelor de apă potrivite în funcție de vârstă.
După consumul de zaharoză pe termen lung (12 săptămâni), lungimea totală a arborelui dendritic a MSN-urilor cu coajă NAc a fost scăzută cu 21% în comparație cu martorii consumatoare de apă (Apă: 1827 ± 148 μm, n = 9; Zaharoză 1449 ± 78 μm, n = 9, *P = 0.0384, Student nepereche cu două cozi t-test, Figura 2, Masa 2). Comparația numărului mediu de bifurcații dendritice (noduri) și terminații dendritice dintre grupurile de apă și zaharoză a evidențiat un nivel redus (deși nu semnificativ) de complexitate dendritică în MSN-urile cu coajă NAc (noduri: apă 12.9 ± 1.4). n = 9, zaharoză 10.1 ± 0.8 n = 9, P = 0.0879; terminații: Apă 17.9 ± 1.4 n = 9, zaharoză 14.8 ± 0.7 n = 9, P = 0.0657, Student nepereche cu două cozi t-test, Tabel 2). Nu a existat nicio modificare a volumului somei (P = 0.9400), lungimea medie a arborelui dendritic (P = 0.1646) sau densitatea totală a coloanei vertebrale (P = 0.3662) în MSN-uri cu coajă NAc de la șobolani consumatoare de zaharoză pe termen lung, în comparație cu martorii de apă. Acești parametri morfometrici sunt detaliați în tabel 2.
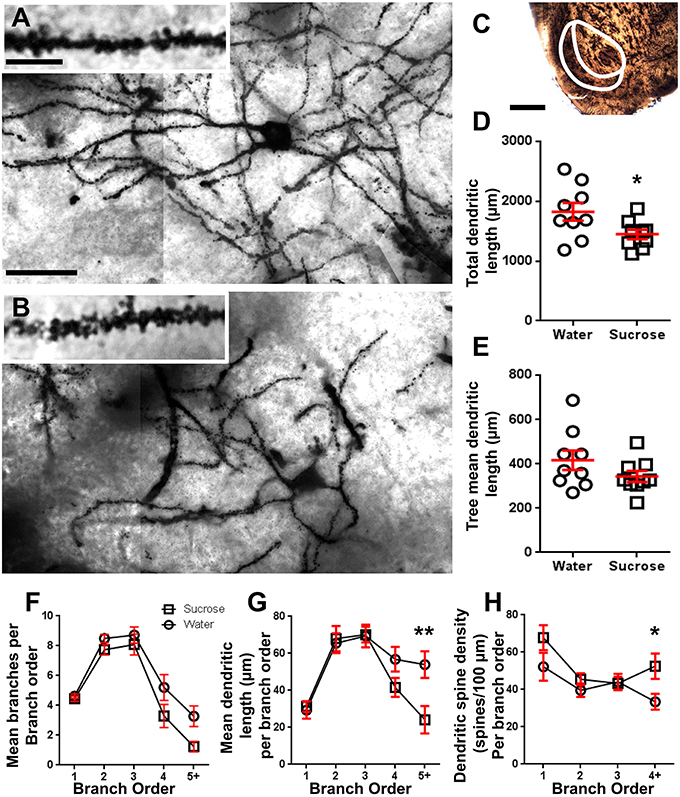 Figura 2. Scăderea lungimii arborelui dendritic și creșterea densității coloanei dendritice distale a neuronilor spinoși medii (MSN) din coaja nucleus accumbens (NAc) a șobolanilor tratați pe termen lung cu zaharoză în comparație cu șobolanii martor. (A,B) arată reprezentări ale câmpului luminos tratat cu zaharoză (de jos) de control (sus) și pe termen lung (12 săptămâni) z-stivuiți mozaicuri de MSN impregnate cu Golgi din carcasa NAc (mărire de 63x). Inset de (A, B) prezintă imagini de control și de câmp luminos tratate cu zaharoză pe termen lung ale dendritelor MSN impregnate cu Golgi și ale spinilor dendritici din învelișul NAc (mărire de 100x). (C) arată regiunile anatomice din care au fost prelevate MSN-urile în acest studiu. (D) arată o diagramă de împrăștiere a arborelui dendritic MSN total scăzut (media ± SEM) din coaja NAc la animalele de zaharoză pe termen lung (pătrate) în comparație cu martori (cercuri), studenți nepereche. t-Test, *P <0.05, n = 9; control şi n = 9; 12 săptămâni zaharoză. (E) arată o diagramă de dispersie a lungimii medii neschimbate a arborelui dendritic MSN (medie ± SEM) din coaja NAc la animalele de zaharoză pe termen lung (pătrate) în comparație cu martori (cercuri), studenți nepereche. t-Test, P > 0.05, n = 9; control şi n = 9; 12 săptămâni zaharoză. Analiza ordinii de ramificație (media ± SEM) a numărului de segmente dendritice pe ordin de ramificație (F), lungimea medie a dendritice pe ordinea ramurilor (G) și densitatea coloanei dendritice pe ordinea ramurilor (H). Consumul de zaharoză pe termen lung a scăzut lungimea dendritică la ordinele de ramuri distale (5+) și a crescut densitatea coloanei dendritice la ordinele de ramuri distale (4+) în comparație cu martorii (G,H), ANOVA bidirecționale cu post-testele Bonferroni, *P <0.05, **P <0.01, n = 9; control şi n = 9; zaharoza pe termen lung. Bare de scară: (A, B) = 20 μm; inserție de (A, B) = 10 μm; (C) = 1 mm.
Figura 2. Scăderea lungimii arborelui dendritic și creșterea densității coloanei dendritice distale a neuronilor spinoși medii (MSN) din coaja nucleus accumbens (NAc) a șobolanilor tratați pe termen lung cu zaharoză în comparație cu șobolanii martor. (A,B) arată reprezentări ale câmpului luminos tratat cu zaharoză (de jos) de control (sus) și pe termen lung (12 săptămâni) z-stivuiți mozaicuri de MSN impregnate cu Golgi din carcasa NAc (mărire de 63x). Inset de (A, B) prezintă imagini de control și de câmp luminos tratate cu zaharoză pe termen lung ale dendritelor MSN impregnate cu Golgi și ale spinilor dendritici din învelișul NAc (mărire de 100x). (C) arată regiunile anatomice din care au fost prelevate MSN-urile în acest studiu. (D) arată o diagramă de împrăștiere a arborelui dendritic MSN total scăzut (media ± SEM) din coaja NAc la animalele de zaharoză pe termen lung (pătrate) în comparație cu martori (cercuri), studenți nepereche. t-Test, *P <0.05, n = 9; control şi n = 9; 12 săptămâni zaharoză. (E) arată o diagramă de dispersie a lungimii medii neschimbate a arborelui dendritic MSN (medie ± SEM) din coaja NAc la animalele de zaharoză pe termen lung (pătrate) în comparație cu martori (cercuri), studenți nepereche. t-Test, P > 0.05, n = 9; control şi n = 9; 12 săptămâni zaharoză. Analiza ordinii de ramificație (media ± SEM) a numărului de segmente dendritice pe ordin de ramificație (F), lungimea medie a dendritice pe ordinea ramurilor (G) și densitatea coloanei dendritice pe ordinea ramurilor (H). Consumul de zaharoză pe termen lung a scăzut lungimea dendritică la ordinele de ramuri distale (5+) și a crescut densitatea coloanei dendritice la ordinele de ramuri distale (4+) în comparație cu martorii (G,H), ANOVA bidirecționale cu post-testele Bonferroni, *P <0.05, **P <0.01, n = 9; control şi n = 9; zaharoza pe termen lung. Bare de scară: (A, B) = 20 μm; inserție de (A, B) = 10 μm; (C) = 1 mm.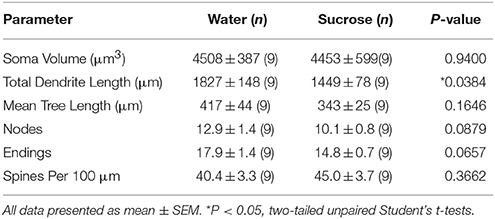
Tabelul 2. Parametrii morfologici generali ai neuronilor spinoși medii din învelișul nucleului accumbens al șobolanilor consumatoare de zaharoză pe termen lung și controalelor de apă potrivite în funcție de vârstă.
În urma caracterizării morfologiei dendritice generale a MSN-urilor de înveliș NAc consumatoare de zaharoză pe termen lung, am analizat arborizările dendritice și densitățile coloanei vertebrale în ceea ce privește caracteristicile ordinului lor de ramuri. Evaluarea noastră cuprinzătoare a copacilor dendritici a cuantificat numărul de segmente dendritice pe ordinea de ramuri, lungimea medie a segmentelor dendritice pe ordinea ramurilor și densitatea medie a coloanei vertebrale pe ordinea de ramuri a MSN-urilor de control al apei cu coajă NAc și șobolanii consumatoare de zaharoză pe termen lung. Un rezumat al datelor și analizei comenzilor de ramuri este prezentat în tabel 3.
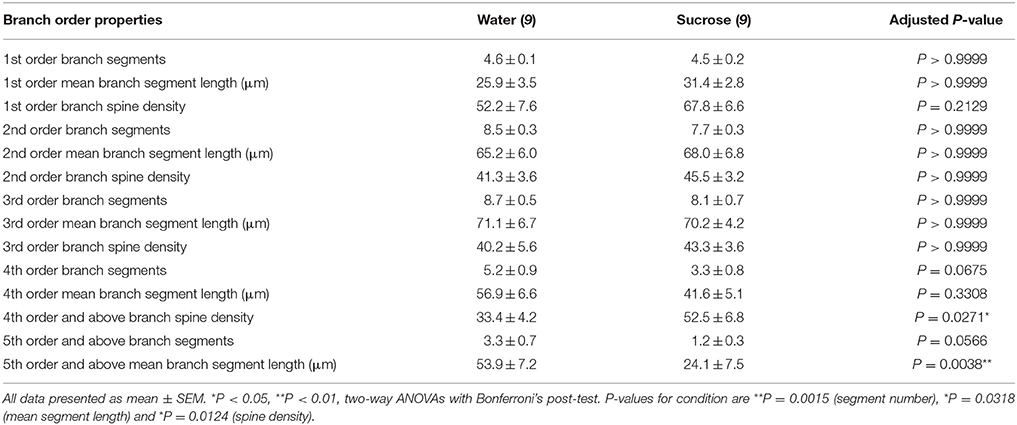 Tabelul 3. Caracteristicile ordinii de ramuri ale neuronilor spinoși medii de la șobolani pe termen lung de zaharoză și apă potabilă.
Tabelul 3. Caracteristicile ordinii de ramuri ale neuronilor spinoși medii de la șobolani pe termen lung de zaharoză și apă potabilă.Numărul mediu de segmente de ramuri dendritice pe ordinul de ramuri ale MSN-urilor cu coajă NAc a fost redus semnificativ la șobolanii consumatoare de zaharoză pe termen lung, comparativ cu martorii de apă (**P = 0.0015, ANOVA în două sensuri). Post-testele Bonferroni au evidențiat o tendință către un număr redus de segmente de ramuri la locul 4 (Apă: 5.2 ± 0.9, n = 9; Zaharoză 3.3 ± 0.8, n = 9, P = 0.0675, Figura 2F, Masa 3), și ordinele de ramuri de ordinul 5 și mai sus (Apă: 3.3 ± 0.7, n = 9; Zaharoză 1.2 ± 0.3, n = 9, P = 0.0566, Figura 2F, Masa 3). Lungimea medie a segmentului dendritic pe ordinul de ramificare a MSN-urilor cu coajă NAc a fost, de asemenea, redusă semnificativ la șobolanii consumatoare de zaharoză pe termen lung, comparativ cu martorii de apă (*P = 0.0444, ANOVA în două sensuri). Post-testele Bonferroni au arătat o reducere de 55% la ramurile de ordinul 5 și mai departe (Apă: 53.9 ± 7.2 μm, n = 9; Zaharoză 24.1 ± 7.5 μm, n = 9, **P = 0.0038, Figura 2G, Masa 3).
Analiza ordinii ramurilor a arătat o creștere semnificativă a densității coloanei dendritice a MSN-urilor de coajă NAc ale șobolanilor consumatoare de zaharoză pe termen lung, comparativ cu martorii (*P = 0.0124, ANOVA în două sensuri). Post-testele Bonferroni au arătat o creștere a densității coloanei vertebrale cu 57% la ramurile distale de ordinul 4 și mai departe (Apă: 33.4 ± 4.2, n = 9; Zaharoză 52.5 ± 6.8, n = 9, P = 0.0271*, inserție de cifre 2A,B,H, Masa 3). Imagini reprezentative ale arhitecturii generale MSN și densității coloanei vertebrale distale (inserție) sunt prezentate în figuri 2A, B.
Luate împreună, aceste rezultate indică faptul că consumul de zaharoză pe termen scurt are un efect redus asupra parametrilor morfologici ai MSN-urilor din învelișul NAc. Cu toate acestea, în urma consumului prelungit, există o scădere semnificativă a lungimii și complexității arborelui neuronal, în special în ramurile dendritice distale. Creșterile concomitente ale densității coloanei vertebrale distale sunt, de asemenea, evidente în MSN-urile de coajă NAc ale șobolanilor care consumă zaharoză pe termen lung.
Neuronii spinoși medii din miezul Nucleus Accumbens au redus complexitatea ramificației după consumul de zaharoză pe termen lung, dar nu pe termen scurt
După consumul de zaharoză pe termen scurt, nu au existat diferențe semnificative în parametrii morfometrici de bază ai NAc MSN (Tabel 4). De asemenea, nu au existat diferențe semnificative între consumul de zaharoză pe 4 săptămâni și MSN-urile centrale de control al apei în analizele legate de ordinea ramurilor centrifuge. Și anume, segmentele dendritice pe ordin de ramificație (P = 0.7717), lungimea medie dendritică pe ordin de ramificare (P = 0.2096) și densitatea medie a coloanei vertebrale pe ordin de ramuri (P = 0.3521, ANOVA bidirecționale) nu au fost diferite între grupuri.
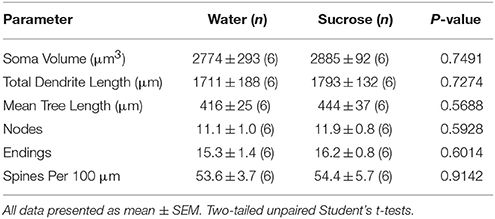 Tabelul 4. Parametrii morfologici generali ai neuronilor spinoși medii din miezul nucleului accumbens al șobolanilor care consumă zaharoză pe termen scurt și controalele de apă potrivite cu vârsta.
Tabelul 4. Parametrii morfologici generali ai neuronilor spinoși medii din miezul nucleului accumbens al șobolanilor care consumă zaharoză pe termen scurt și controalele de apă potrivite cu vârsta.Consumul prelungit de zaharoză nu a avut nicio semnificație asupra parametrilor morfometrici MSN de bază NAc (Tabel 5). Numărul mediu de segmente de ramuri dendritice pe ordinul de ramuri ale MSN-urilor de bază NAc a fost redus semnificativ la șobolanii care consumă zaharoză pe termen lung în comparație cu martorii de apă (*P = 0.0416, ANOVA bidirecțional), cu toate acestea nu au existat diferențe semnificative în lungimea medie a dendritice pe ordinea de ramuri (P = 0.0995) și densitatea medie a coloanei vertebrale pe ordin de ramuri (P = 0.4888, ANOVA în două sensuri) între MSN-urile din miezul NAc al șobolanilor consumatoare de zaharoză pe termen lung în comparație cu martorii de apă. Luate împreună, datele noastre arată că miezul NAc nu este la fel de sensibil la consumul de zaharoză pe termen lung în comparație cu MSN-urile din regiunea coajă NAc.
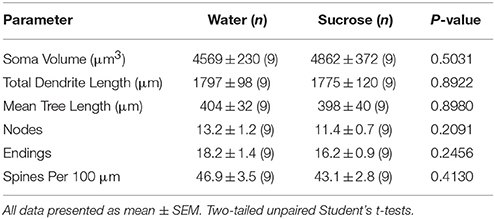 Tabelul 5. Parametrii morfologici generali ai neuronilor spinoși medii din miezul nucleului accumbens al șobolanilor care consumă zaharoză pe termen lung și controalele de apă potrivite cu vârsta.
Tabelul 5. Parametrii morfologici generali ai neuronilor spinoși medii din miezul nucleului accumbens al șobolanilor care consumă zaharoză pe termen lung și controalele de apă potrivite cu vârsta.Discuție
Disponibilitatea crescută a alimentelor foarte îndulcite în dieta occidentală nu numai că a contribuit la creșterea prevalenței și a poverii economice a obezității și a diabetului de tip II, ci a condus și la apariția tulburărilor de alimentație, cum ar fi alimentația excesivă.Swanson și colab., 2011; Kessler și colab., 2013; Davis, 2015). Deși proprietățile de dependență ale zaharurilor, inclusiv fructoza și zaharoza, rămân speculative, există o similitudine izbitoare în corelațiile comportamentale și neuronale care se manifestă ca urmare a supraalimentării și consumului prelungit de droguri. (Avena și colab., 2008, 2011). În plus, zahărul activează circuitele de recompensă ale creierului într-un mod similar cu drogurile de abuz (Volkow și colab., 2012), iar rezultatele din studiile pe oameni sugerează că zahărul și dulceața pot induce pofte care sunt comparabile ca magnitudine cu cele induse de drogurile care creează dependență, cum ar fi alcoolul și nicotina. (Volkow și colab., 2012). Prin urmare, am folosit un model de consum excesiv de zaharoză la șobolani pentru a determina efectele consumului de zaharoză pe termen scurt (4 săptămâni) și pe termen lung (12 săptămâni) asupra morfologiei neuronale a MSN-urilor în NAc, o componentă cheie a circuitelor de recompensă suprapuse. care este modulată de zahăr și droguri care creează dependență. Arătăm că MSN-urile din învelișul NAc a șobolanilor cronici care consumă zaharoză pe termen lung au scăzut semnificativ lungimea și complexitatea dendritice, dar au crescut densitatea coloanei dendritice distale. Consumul de zaharoză pe termen lung nu a avut niciun efect asupra morfologiei MSN din miezul NAc, în timp ce consumul de zaharoză pe termen scurt nu a avut nici un efect semnificativ asupra morfologiei MSN din miezul sau învelișul NAc. Aceste rezultate nu numai că demonstrează un efect direct al aportului prelungit de zaharoză asemănătoare binge-like asupra morfologiei neuronale a MSN-urilor cu coajă NAc, dar evidențiază și consecințele potențial dăunătoare ale consumului prelungit de diete cu conținut ridicat de zahăr.
NAc, care face parte din striatul ventral, este format în principal din MSN, care sunt caracterizați morfologic ca neuroni de dimensiuni medii cu arborizări dendritice extinse și densitate mare a coloanei vertebrale (Kemp și Powell, 1971; Graveland și DiFiglia, 1985; Rafols și colab., 1989; Kawaguchi și colab., 1990). Neuronii glutamatergici și dopaminergici sunt cele două intrări aferente primare la NAc, contactând în primul rând arborii dendritici și coloanele MSN. (Groves, 1980; Kaiya și Namba, 1981; Groves și colab., 1994). În mod specific, învelișul și miezul NAc primesc input glutamatergic din zone corticale distincte funcțional (Brog și colab., 1993). Învelișul NAc este, de asemenea, inervat de aferente excitatorii din regiunile subcorticale, cum ar fi hipocampul, talamusul și amigdala bazolaterală. (Brog și colab., 1993; Wright și Groenewegen, 1995). Studiile anterioare au demonstrat că aceste aporturi glutamatergice joacă un rol esențial în motivație și comportamente direcționate către obiective, cum ar fi căutarea hranei și a recompenselor. (Maldonado-Irizarry și colab., 1995; Kelley și Swanson, 1997; Reynolds și Berridge, 2003; Richard și Berridge, 2011). Cealaltă intrare predominantă asupra MSN-urilor NAc este din aferentele dopaminergice care se proiectează din zona tegmentală ventrală (Lindvall și Björklund, 1978; Veening și colab., 1980; Kalivas și Miller, 1984). În mod interesant, studiile anterioare care utilizează modele similare de acces intermitent la zahăr au arătat că consumul similar rezultat are ca rezultat o creștere a dopaminei extracelulare în NAc, în mod similar (deși într-o măsură mai mică) cu drogurile de abuz. (Rada și colab., 2005; Avena și colab., 2006), și poate modula expresia receptorului de dopamină (Colantuoni și colab., 2001, 2002) în miezul și învelișul NAc. În mod interesant, consumul de zaharoză în mod asemănător provoacă o escaladare a aportului în timp, similar cu autoadministrarea de droguri de abuz, cum ar fi cocaina și eroul.n (Ahmed și Koob, 1998; Ahmed și colab., 2000, 2003) care este asociat cu dezvoltarea unei stări „de dependență asemănătoare”.
Analiza noastră a morfometriei ordinii de ramificație arată că reducerea globală a lungimii dendritice a MSN-urilor învelișului NAc cauzată de aportul de zaharoză pe termen lung, rezultă în principal din reducerile complexității ordinelor de ramificație distale. Am observat o ramificare distală redusă (ordinele 4 și 5 și mai sus) și lungimea medie redusă semnificativ la dendrite de ordinul 5 și mai sus, combinată cu densități crescute ale coloanei vertebrale la aceste ordine de ramuri. Un factor comun care poate influența acest tip de restructurare dendritică include modificări ale conectivității și/sau funcției sinaptice (Russo și colab., 2010). Studiile anterioare au arătat că sinapsele glutamatergice de pe MSN se formează în principal pe coloane, în special la dendritele distale. (Groenewegen și colab., 1999). În plus, co-localizarea dopaminei și a aporturilor glutamatergice din cortexul prefrontal (Sesack și Pickel, 1992), cal de mare (Totterdell și Smith, 1989; Sesack și Pickel, 1990), si amigdala (Johnson și colab., 1994) au fost observate pe coloanele dendritice ale MSN. Aceste observații, combinate cu densitatea crescută a coloanei vertebrale în urma consumului de zaharoză pe termen lung, observată în studiul nostru, susțin formarea unor intrări excitatorii crescute. Prin urmare, apare posibilitatea în cazul în care efectele persistente cauzate de aportul prelungit de zaharoză asemănătoare binge-ului ar putea facilita creșterea activității sinaptice excitatorii la dendritele distale ale MSN-urilor din învelișul NAc. În consecință, reducerea și/sau retragerea dendritelor distale poate rezulta printr-un mecanism homeostatic sinaptic. (Reissner și Kalivas, 2010), cu toate acestea, aceasta rămâne de stabilit.
Este interesant de remarcat faptul că Crombag și colegii săi au arătat că nu a existat o creștere a densității coloanei vertebrale în învelișul NAc după consumul de zaharoză timp de 4 săptămâni prin paradigma de autoadministrare cu nasul, în ciuda unei achiziții mai robuste și a unei rate de răspuns mai ridicate la zaharoză în comparație. cu amfetamina (Crombag și colab., 2005). Observarea lor a unei absențe a modificării densității coloanei vertebrale la 4 săptămâni oglindește constatările noastre. Prin contrast, totuși, studiul nostru demonstrează că în urma expunerii pe termen lung (12 săptămâni) la consumul cronic de zaharoză, există o creștere semnificativă a densității coloanei vertebrale distale pe MSN-urile șobolanilor cu experiențe cu zaharoză. Mai mult, laboratorul nostru a arătat anterior că consumul de zaharoză pe termen lung (12 săptămâni) facilitează un răspuns farmacologic diferențial la medicamentele farmacoterapeutice care s-au dovedit că modulează răspunsurile la dopamină și acetilcolină la nivelul NAc (Shariff et al., în presă). Luat împreună, acest lucru sugerează că expunerea la zaharoză pe termen lung (12 săptămâni și mai mult), care reflectă mai exact scenariile din lumea reală, are ca rezultat adaptări morfologice la nivelul NAc.
În ceea ce privește drogurile de abuz, expunerea repetată la diferite medicamente produce modificări de lungă durată în structura dendritelor și a coloanelor dendritice. De exemplu, amfetaminele și cocaina cresc ambele densitatea coloanei vertebrale în NAc atât în coajă, cât și în miez (Robinson și Kolb, 2004). De asemenea, sa demonstrat că expunerea la nicotină crește densitatea coloanei vertebrale în învelișul NAc. În schimb, expunerea la morfină duce la o scădere a densității coloanei vertebrale și a complexității ramurilor dendritice. (Robinson și Kolb, 2004). În ceea ce privește consumul de zaharoză pe termen lung, am observat o creștere a densității coloanei vertebrale similară cu amfetaminei, cocainei și nicotinei și opus efectului morfinei. Cu toate acestea, spre deosebire de amfetamină și cocaină, dar similar cu nicotina, creșterea densității coloanei vertebrale la expunerea pe termen lung la zaharoză este limitată la învelișul NAc. De asemenea, este interesant faptul că modificările atât în ramificarea dendritică (Robinson și Kolb, 1999) și densitatea coloanei vertebrale (Li și colab., 2003) produse de amfetamină sau cocaină sunt limitate la dendritele distale ale MSN din NAc, ceea ce reflectă concluziile studiului nostru. În plus, și coroborând modificările descrise mai sus, sa demonstrat anterior că consumul de zaharoză îmbunătățește puterea sinaptică excitatoare asupra neuronilor dopaminergici acumulatori (Stuber și colab., 2008b), precum și alte componente ale căii recompensei mezolimbice (Stuber și colab., 2008a; Chen și colab., 2010). Luate împreună, aceasta prezintă zaharoza ca un modulator puternic al morfologiei neuronilor în urma utilizării intense prelungite, ceea ce este asemănător cu efectele observate în urma abuzului de droguri.
Deși sunt necesare investigații suplimentare pentru a descoperi mecanismele celulare și sinaptice care contribuie la modificările morfologice observate în acest studiu, rezultatele noastre demonstrează efecte neuronale semnificative generate de consumul de zaharoză pe termen lung. În special, o considerație care nu a fost examinată în studiul nostru este dacă efectele morfologice observate ale zaharozei pot fi provocate și cu îndulcitori necalorici, cum ar fi zaharina. În acest sens, este important de menționat că Lenoir și colegii săi au arătat că dulceața intensă depășește recompensa cocainei, fie ea generată de zaharină sau zaharoză (Lenoir și colab., 2007). Mai mult, un studiu recent publicat de laboratorul nostru (Shariff et al., în presă) demonstrează că vareniclina, un agonist parțial al receptorului de acetilcolină nicotinic, a redus atât aportul de zaharoză, cât și de zaharină la rozătoare, urmând același regim de acces intermitent pe termen lung utilizat în prezentul studiu. Interesant, studiile anterioare au arătat asemănări în efectele acute ale îndulcitorilor necalorici, cum ar fi zaharina și zaharoza, la nivelul NAc (Scheggi și colab., 2013; Tukey și colab., 2013; Carelli și West, 2014). Cu toate acestea, sunt necesare studii suplimentare pentru a determina dacă îndulcitorii necalorici pot induce efecte pe termen lung similare cu modificările morfologiei MSN-urilor învelișului NAc cauzate de consumul de zaharoză pe termen lung raportat aici.
Lipsa efectului asupra morfologiei NAc MSN în urma consumului de zaharoză pe termen scurt, evidențiază importanța implementării unor studii pe termen lung pentru a evalua impactul abuzului prelungit de droguri sau recompense naturale precum zaharoza. În ceea ce privește dependența, nu numai că ciclurile repetate de consum excesiv și abstinența sunt componente cheie ale ciclului de dependență, un număr tot mai mare de dovezi au dezvăluit că tranziția la dependență este un proces progresiv care are loc adesea pe o perioadă lungă de timp. Deși proprietățile de dependență ale zaharurilor rămân incerte, plauzibilitatea dependenței de alte recompense non-drog, cum ar fi sexul, jocurile de noroc și mâncarea este din ce în ce mai investigată. Rezultatele acestui studiu adaugă merit ipotezei conform căreia zaharurile, cum ar fi zaharoza, au potențial proprietăți de dependență în urma consumului pe termen lung, asemănător cu excesul. Rezultatele noastre au, de asemenea, implicații pentru numărul tot mai mare de copii și adolescenți care mențin obiceiuri alimentare nesănătoase (consum mare de zahăr și alimentație excesivă) până la vârsta adultă. În concordanță cu riscul crescut de a dezvolta efecte metabolice, este, de asemenea, posibil ca și consecințe neurologice și psihiatrice care afectează starea de spirit și motivația să rezulte din aceste comportamente.
Contribuțiile autorului
A participat la proiectarea cercetării: PK, SB. Experimente efectuate: PK, MS, AB, MF, EM. Analiza datelor: PK, MF, MS. A interpretat datele și a contribuit la scrierea manuscrisului: PK, MS, MF, EM, MB, SB. Toți autorii au citit și au aprobat manuscrisul final pentru depunere.
Declarația privind conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Revisorii SC, SA și Handling Editor și-au declarat afilierea comună, iar Handling Editor afirmă că procesul a îndeplinit totuși standardele unei revizuiri corecte și obiective.
recunoasteri
Aceasta lucrare a fost sustinuta de finantare din granturi de la Australian Research Council (FT1110884) catre SB si National Health and Medical Research Council (1061979) catre SB si MB.
Material suplimentar
Materialul suplimentar pentru acest articol poate fi găsit online la adresa: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
Figura suplimentară 1. Aportul de zaharoză și preferința de la șobolanii care consumă zaharoză de 4 și 12 săptămâni. (A,B) arată o creștere a aportului total de zaharoză (ml) pe parcursul a 4 și 12 săptămâni de expunere. (CD) arată o preferință ridicată pentru zaharoză față de apă în perioadele de prezentare a zaharozei.
Referinte
Ahmed, SH și Koob, GF (1998). Tranziția de la aportul moderat la consumul excesiv de medicamente: modificarea punctului de referință hedonic. Ştiinţă 282, 298-300. doi: 10.1126 / science.282.5387.298
Ahmed, SH, Lin, D., Koob, GF și Parsons, LH (2003). Escaladarea autoadministrarii cocainei nu depinde de alterarea nivelului de dopamină indus de cocaină nucleus accumbens. J. Neurochem. 86, 102-113. doi: 10.1046 / j.1471-4159.2003.01833.x
Ahmed, SH, Walker, JR și Koob, GF (2000). Creșterea persistentă a motivației de a lua heroină la șobolanii cu antecedente de escaladare a drogurilor. Neuropsychopharmacology 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
Arslanian, S. (2002). Diabetul de tip 2 la copii: aspecte clinice și factori de risc. Horm Res 57 (Supl. 1), 19–28. doi: 10.1159/000053308
Avena, NM, Bocarsly, ME, Hoebel, BG și Gold, MS (2011). Suprapuneri în nosologia abuzului de substanțe și a supraalimentării: implicațiile translaționale ale „dependenței alimentare”. Curr. Abuzul de droguri Rev. 4, 133-139. doi: 10.2174 / 1874473711104030133
Avena, NM, Rada, P. și Hoebel, BG (2008). Dovezi privind dependența de zahăr: efectele comportamentale și neurochimice ale aportului intermitent, excesiv de zahăr. Neurosci. Biobehav. Rev. 32, 20-39. doi: 10.1016 / j.neubiorev.2007.04.019
Avena, NM, Rada, P., Moise, N. și Hoebel, BG (2006). Hrănirea simulată de zaharoză după un program de exces eliberează dopamina accumbens în mod repetat și elimină răspunsul de sațietate cu acetilcolină. Neuroştiinţe 139, 813-820. doi: 10.1016 / j.neuroscience.2005.12.037
Benton, D. (2010). Plauzibilitatea dependenței de zahăr și rolul acesteia în obezitate și tulburări de alimentație. Clin. Nutr. 29, 288–303. doi: 10.1016/j.clnu.2009.12.001
Bray, GA și Popkin, BM (2014). Zahărul alimentar și greutatea corporală: am ajuns la o criză în epidemia de obezitate și diabet?: sănătatea să fie dracu’! Se toarnă zahărul. Îngrijirea diabetului 37, 950–956. doi: 10.2337/dc13-2085
Brog, JS, Salyapongse, A., Deutch, AY și Zahm, DS (1993). Modelele de inervație aferentă a miezului și a învelișului în partea „accumbens” a striatului ventral de șobolan: detectarea imunohistochimică a fluoro-aurului transportat retrograd. J. Comp. Neural. 338, 255–278. doi: 10.1002/cne.903380209
Bucher Della Torre, S., Keller, A., Laure Depeyre, J. și Kruseman, M. (2015). Băuturile îndulcite cu zahăr și riscul de obezitate la copii și adolescenți: o analiză sistematică a modului în care calitatea metodologică poate influența concluziile. J. Acad. Nutr. Dietă. [Epub înainte de tipărire]. doi: 10.1016/j.jand.2015.05.020
Carelli, RM și West, EA (2014). Când un gust bun devine rău: mecanismele neuronale care stau la baza apariției afectului negativ și devalorizarea recompensei naturale asociate de către cocaină. Neuropharmacology 76 (Pt B), 360–369. doi: 10.1016/j.neuropharm.2013.04.025
Chen, BT, Hopf, FW și Bonci, A. (2010). Plasticitatea sinaptică în sistemul mezolimbic: implicații terapeutice pentru abuzul de substanțe. Ann. NY Acad. Sci. 1187, 129-139. doi: 10.1111 / j.1749-6632.2009.05154.x
Colantuoni, C., Rada, P., McCarthy, J., Patten, C., Avena, NM, Chadeayne, A., și colab. (2002). Dovezi că aportul intermitent și excesiv de zahăr cauzează dependență endogenă de opioide. Obes. Res. 10, 478-488. doi: 10.1038 / oby.2002.66
Colantuoni, C., Schwenker, J., McCarthy, J., Rada, P., Ladenheim, B., Cadet, JL, et al. (2001). Aportul excesiv de zahăr modifică legarea de receptorii dopamină și mu-opioizi din creier. Neuroreport 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
Crombag, HS, Gorny, G., Li, Y., Kolb, B. și Robinson, TE (2005). Efecte opuse ale experienței de autoadministrare a amfetaminei asupra coloanelor dendritice din cortexul prefrontal medial și orbital. Cereb. cortex 15, 341-348. doi: 10.1093 / cercor / bhh136
Dabelea, D., Mayer-Davis, EJ, Saydah, S., Imperatore, G., Linder, B., Divers, J., et al. (2014). Prevalența diabetului de tip 1 și tip 2 în rândul copiilor și adolescenților din 2001 până în 2009. JAMA 311, 1778-1786. doi: 10.1001 / jama.2014.3201
Davis, C. (2015). Epidemiologia și genetica tulburării de alimentație excesivă (BED). CNS Spectr. 20, 522-529. doi: 10.1017 / s1092852915000462
Ford, ES și Dietz, WH (2013). Tendințe în consumul de energie în rândul adulților din Statele Unite: constatări de la NHANES. A.m. J. Clin. Nutr. 97, 848–853. doi: 10.3945/ajcn.112.052662
Fryar, CD, Carroll, MD și Ogden, CL (2014). Prevalența excesului de greutate, a obezității și a obezității extreme în rândul adulților: Statele Unite, 1960–1962 până în 2011–2012. Atlanta, GA: CDC.
Graveland, GA şi DiFiglia, M. (1985). Frecvența și distribuția neuronilor de dimensiuni medii cu nuclee indentate în neostriatul de primate și rozătoare. Brain Res. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
Groenewegen, HJ, Wright, CI, Beijer, AV și Voorn, P. (1999). Convergența și segregarea intrărilor și ieșirilor striatale ventrale. Ann. NY Acad. Sci. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
Groves, PM (1980). Terminațiile sinaptice și țintele lor postsinaptice în neostriat: specializări sinaptice relevate din analiza secțiunilor în serie. Proc. Natl. Acad. Sci. Statele Unite ale Americii 77, 6926-6929. doi: 10.1073 / pnas.77.11.6926
Groves, PM, Linder, JC și Young, SJ (1994). Axoni dopaminergici marcați cu 5-hidroxidopamină: reconstrucții tridimensionale ale axonilor, sinapselor și țintelor postsinaptice în neostriatul de șobolan. Neuroştiinţe 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
Hu, FB (2013). Rezolvat: există suficiente dovezi științifice că scăderea consumului de băuturi îndulcite cu zahăr va reduce prevalența obezității și a bolilor legate de obezitate. Obes. Rev. 14, 606–619. doi: 10.1111/obr.12040
Johnson, LR, Aylward, RL, Hussain, Z. şi Totterdell, S. (1994). Intrarea de la amigdala la nucleul accumbens de șobolan: relația sa cu imunoreactivitatea tirozin-hidroxilazei și neuronii identificați. Neuroştiinţe 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
Kaiya, H. și Namba, M. (1981). Două tipuri de terminale nervoase dopaminergice în neostriatul șobolanului. Un studiu ultrastructural. Neurosci. Lett. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
Kalivas, PW și Miller, JS (1984). Neuronii neurotensinei din zona tegmentală ventrală se proiectează către nucleul medial accumbens. Brain Res. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
Kawaguchi, Y., Wilson, CJ și Emson, PC (1990). Subtipuri de proiecție ale celulelor matricei neostriatale de șobolan dezvăluite prin injectarea intracelulară de biocitină. J. Neurosci. 10, 3421-3438.
Kelley, AE și Swanson, CJ (1997). Hrănirea indusă de blocarea receptorilor AMPA și kainați în striatul ventral: un studiu de cartografiere a microinfuziei. Behav. Brain Res. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
Kemp, JM și Powell, TP (1971). Organizarea sinaptică a nucleului caudat. Philos. Trans. R. Soc. Lond. B Biol. Sci. 262, 403-412. doi: 10.1098 / rstb.1971.0103
Kenny, PJ (2011). Mecanisme de recompensare în obezitate: noi perspective și direcții viitoare. Neuron 69, 664-679. doi: 10.1016 / j.neuron.2011.02.016
Kessler, RC, Berglund, PA, Chiu, WT, Deitz, AC, Hudson, JI, Shahly, V., et al. (2013). Prevalența și corelațiile tulburării de alimentație excesivă în Sondajele Mondiale de Sănătate Mintală ale Organizației Mondiale a Sănătății. Biol. Psihiatrie 73, 904-914. doi: 10.1016 / j.biopsych.2012.11.020
Klenowski, PM, Fogarty, MJ, Belmer, A., Noakes, PG, Bellingham, MC și Bartlett, SE (2015). Caracterizarea structurală și funcțională a arborelor dendritice și a intrărilor sinaptice GABAergice pe interneuroni și celulele principale din amigdala bazolaterală de șobolan. J. Neurophysiol. 114, 942-957. doi: 10.1152 / jn.00824.2014
Lenoir, M., Serre, F., Cantin, L. și Ahmed, SH (2007). Dulceața intensă depășește recompensa pentru cocaină. PLoS ONE 2: e698. doi: 10.1371 / journal.pone.0000698
Li, Y., Kolb, B. și Robinson, TE (2003). Localizarea modificărilor persistente induse de amfetamine în densitatea spinilor dendritici pe neuronii spinoși medii din nucleul accumbens și caudat-putamen. Neuropsychopharmacology 28, 1082-1085. doi: 10.1038 / sj.npp.1300115
Lindvall, O. și Björklund, A. (1978). Anatomia sistemelor neuronale dopaminergice din creierul de șobolan. Adv. Biochim. Psihofarmacol. 19, 1-23.
Lutter, M., și Nestler, EJ (2009). Semnalele homeostatice și hedonice interacționează în reglarea aportului alimentar. J. Nutr. 139, 629-632. doi: 10.3945 / jn.108.097618
Maldonado-Irizarry, CS, Swanson, CJ și Kelley, AE (1995). Receptorii de glutamat din coaja nucleului accumbens controlează comportamentul de hrănire prin intermediul hipotalamusului lateral. J. Neurosci. 15, 6779-6788.
Malik, VS, Popkin, BM, Bray, GA, Després, JP și Hu, FB (2010). Băuturi îndulcite cu zahăr, obezitate, diabet zaharat de tip 2 și risc de boli cardiovasculare. Circulație 121, 1356–1364. doi: 10.1161/CIRCULATIONAHA.109.876185
Ng, SW, Slining, MM și Popkin, BM (2012). Utilizarea îndulcitorilor calorici și necalorici în alimentele ambalate pentru consumatori din SUA, 2005-2009. J. Acad. Nutr. Dietă. 112, 1828–1834 e1821–e1826. doi: 10.1016/j.jand.2012.07.009
Nielsen, SJ, Siega-Riz, AM și Popkin, BM (2002). Tendințe în consumul de energie în SUA între 1977 și 1996: schimbări similare observate pe grupe de vârstă. Obes. Res. 10, 370-378. doi: 10.1038 / oby.2002.51
Paxinos, G. și Watson, C. (2007). Brainul de șobolan în coordonatele stereotaxice. Amsterdam; Boston, MA: Academic Press/Elsevier.
Popkin, BM (2010). Ce este în neregulă cu abordarea obezității în SUA? Virtual Mentor 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
Rada, P., Avena, NM și Hoebel, BG (2005). Consumul zilnic de zahăr eliberează în mod repetat dopamină în coaja accumbens. Neuroştiinţe 134, 737-744. doi: 10.1016 / j.neuroscience.2005.04.043
Rafols, JA, Cheng, HW și McNeill, TH (1989). Studiul Golgi al striatului de șoarece: modificări dendritice legate de vârstă în diferite populații neuronale. J. Comp. Neural. 279, 212–227. doi: 10.1002/cne.902790205
Ranjan, A. și Mallick, BN (2010). O metodă modificată pentru colorarea Golgi-cox consistentă și fiabilă într-un timp semnificativ redus. Față. Neural. 1:157. doi: 10.3389/fneur.2010.00157
Reinehr, T. (2013). Diabet zaharat de tip 2 la copii și adolescenți. World J. Diabet 4, 270–281. doi: 10.4239/wjd.v4.i6.270
Reissner, KJ și Kalivas, PW (2010). Folosirea homeostaziei glutamatului ca țintă pentru tratarea tulburărilor de dependență. Behav. Pharmacol. 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
Reynolds, SM și Berridge, KC (2003). Ansambluri motivaționale de glutamat în nucleus accumbens: gradienți rostrocaudal de frică și hrănire. EURO. J. Neurosci. 17, 2187-2200. doi: 10.1046 / j.1460-9568.2003.02642.x
Richard, JM și Berridge, KC (2011). Interacțiunea Nucleus accumbens dopamină/glutamat schimbă modurile pentru a genera dorință versus groază: D(1) singur pentru mâncare apetisantă, dar D(1) și D(2) împreună de frică. J. Neurosci. 31, 12866-12879. doi: 10.1523 / JNEUROSCI.1339-11.2011
Robinson, TE şi Kolb, B. (1999). Modificări ale morfologiei dendritelor și spinilor dendritici în nucleul accumbens și cortexul prefrontal în urma tratamentului repetat cu amfetamină sau cocaină. EURO. J. Neurosci. 11, 1598-1604. doi: 10.1046 / j.1460-9568.1999.00576.x
Robinson, TE și Kolb, B. (2004). Plasticitatea structurală asociată cu expunerea la droguri de abuz. Neuropharmacology 47(Suppl. 1), 33–46. doi: 10.1016/j.neuropharm.2004.06.025
Russo, SJ, Dietz, DM, Dumitriu, D., Morrison, JH, Malenka, RC și Nestler, EJ (2010). Sinapsei dependente: mecanismele plasticității sinaptice și structurale în nucleul accumbens. Tendințe Neurosci. 33, 267-276. doi: 10.1016 / j.tins.2010.02.002
Rutledge, LT, Duncan, J. și Beatty, N. (1969). Un studiu al colateralelor axonilor celulelor piramidale în cortexul cerebral adult intact și parțial izolat. Brain Res. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper, CB, Chou, TC și Elmquist, JK (2002). Nevoia de hrănire: controlul homeostatic și hedonic al consumului. Neuron 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
Scheggi, S., Secci, ME, Marchese, G., De Montis, MG și Gambarana, C. (2013). Influența palatabilității asupra motivației de a opera pentru hrana calorică și necalorică la șobolanii care nu sunt lipsiți de alimente și lipsiți de hrană. Neuroştiinţe 236, 320-331. doi: 10.1016 / j.neuroscience.2013.01.027
Sesack, SR și Pickel, VM (1990). În nucleul accumbens medial de șobolan, terminalele hipocampale și catecolaminergice converg spre neuronii spinoși și sunt în apoziție unul cu celălalt. Brain Res. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
Sesack, SR și Pickel, VM (1992). Eferente corticale prefrontale în sinapsa șobolanului pe ținte neuronale neetichetate ale terminalelor catecolaminei din nucleul accumbens septus și pe neuronii dopaminergici din zona tegmentală ventrală. J. Comp. Neural. 320, 145–160. doi: 10.1002/cne.903200202
Shariff, M., Quik, M., Holgate, JY, Morgan, M., Patkar, OL, Tam, V., și colab. (in presa). Modulatorii neuronali ai receptorilor nicotinici de acetilcolină reduc aportul de zahăr. PLoS ONE.
Sheehan, DV și Herman, BK (2015). Factorii psihologici și medicali asociați cu tulburarea de alimentație excesivă netratată. Prim. Îngrijitorul însoțitor CNS Disord. 17. doi: 10.4088/PCC.14r01732
Simms, JA, Steensland, P., Medina, B., Abernathy, KE, Chandler, LJ, Wise, R., et al. (2008). Accesul intermitent la 20% etanol induce un consum mare de etanol la șobolanii Long-Evans și Wistar. Alcool. Clin. Exp. Res. 32, 1816-1823. doi: 10.1111 / j.1530-0277.2008.00753.x
Steensland, P., Simms, JA, Holgate, J., Richards, JK și Bartlett, SE (2007). Vareniclina, un agonist parțial al receptorului de acetilcolină nicotinic alfa4beta2, scade selectiv consumul și căutarea de etanol. Proc. Natl. Acad. Sci. Statele Unite ale Americii 104, 12518-12523. doi: 10.1073 / pnas.0705368104
Stuber, GD, Hopf, FW, Hahn, J., Cho, SL, Guillory, A. și Bonci, A. (2008a). Aportul voluntar de etanol mărește puterea sinaptică excitatoare în zona tegmentală ventrală. Alcool. Clin. Exp. Res. 32, 1714-1720. doi: 10.1111 / j.1530-0277.2008.00749.x
Stuber, GD, Klanker, M., de Ridder, B., Bowers, MS, Joosten, RN, Feenstra, MG și colab. (2008b). Indiciile predictive de recompensă îmbunătățesc puterea sinaptică excitatoare asupra neuronilor dopaminergici din creierul mijlociu. Ştiinţă 321, 1690-1692. doi: 10.1126 / science.1160873
Swanson, SA, Crow, SJ, Le Grange, D., Swendsen, J. și Merikangas, KR (2011). Prevalența și corelațiile tulburărilor de alimentație la adolescenți. Rezultatele studiului național de comorbiditate replicare supliment pentru adolescenți. Arc. Gen. Psihiatrie 68, 714-723. doi: 10.1001 / archgenpsychiatry.2011.22
Te Morenga, L., Mallard, S. și Mann, J. (2013). Zaharuri alimentare și greutate corporală: revizuire sistematică și meta-analize ale studiilor controlate randomizate și ale studiilor de cohortă. BMJ 346:e7492. doi: 10.1136/bmj.e7492
Totterdell, S. și Smith, AD (1989). Convergența aportului hipocampal și dopaminergic asupra neuronilor identificați în nucleul accumbens al șobolanului. J. Chem. Neuroanat. 2, 285-298.
Tukey, DS, Ferreira, JM, Antoine, SO, D'Amour, JA, Ninan, I., Cabeza de Vaca, S., et al. (2013). Ingestia de zaharoză induce traficul rapid de receptori AMPA. J. Neurosci. 33, 6123-6132. doi: 10.1523 / JNEUROSCI.4806-12.2013
Veening, JG, Cornelissen, FM și Lieven, PA (1980). Organizarea topică a aferentelor la caudatoputamenul șobolanului. Un studiu de peroxidază de hrean. Neuroştiinţe 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
Ventura, T., Santander, J., Torres, R. și Contreras, AM (2014). Bazele neurobiologice ale poftei de carbohidrați. Nutriţie 30, 252-256. doi: 10.1016 / j.nut.2013.06.010
Volkow, ND, Wang, GJ, Fowler, JS, Tomasi, D. și Baler, R. (2012). Recompensa pentru alimente și medicamente: circuite suprapuse în obezitatea și dependența umană. Curr. Top. Behav. Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
Wise, RA (1973). Aportul voluntar de etanol la șobolani după expunerea la etanol la diferite programe. Psychopharmacologia 29, 203-210. doi: 10.1007 / BF00414034
Wright, CI și Groenewegen, HJ (1995). Modele de convergență și segregare în nucleul medial accumbens al șobolanului: relațiile dintre corticala prefrontală, linia mediană talamică și aferentele amigdaloide bazale. J. Comp. Neural. 361, 383–403. doi: 10.1002/cne.903610304
Cuvinte cheie: consum de tip binge-like, pe termen lung, neuron spinos mediu, nucleus accumbens, zaharoză
Referire: Klenowski PM, Shariff MR, Belmer A, Fogarty MJ, Mu EWH, Bellingham MC și Bartlett SE (2016) Consumul prelungit de zaharoză într-o manieră de binge-Like, Alters the Morphology of Medium Spiny Neurons in the Nucleus Accumbens Shell. Față. Behav. Neurosci. 10: 54. doi: 10.3389 / fnbeh.2016.00054
Primit: 03 decembrie 2015; Acceptat: 07 martie 2016;
Publicat: 23 martie 2016.
Editat de:
Djoher Nora Abrous, Institut des Neurosciences din Bordeaux, Franța
Revizuite de:
Serge H. Ahmed, Centrul Național de Recherche Scientifique, Franța
Stéphanie Caille, Centrul Național de Recherche Scientifique, Franța
Copyright © 2016 Klenowski, Shariff, Belmer, Fogarty, Mu, Bellingham și Bartlett. Acesta este un articol cu acces deschis distribuit în conformitate cu termenii Creative Commons Attribution License (CC BY). Utilizarea, distribuirea sau reproducerea în alte forumuri este permisă, cu condiția ca autorii sau licențiatorii originali să fie creditați și să fie citată publicația originală din acest jurnal, în conformitate cu practica academică acceptată. Nu este permisă utilizarea, distribuirea sau reproducerea care nu respectă acești termeni.
* Corespondență: Selena E. Bartlett, [e-mail protejat]