Dev Rev. 2008; 28(1): 62-77. doi: 10.1016 / j.dr.2007.08.003
Abstract
Adolescența este o perioadă de dezvoltare caracterizată prin decizii și acțiuni suboptimale care dau naștere unei incidențe sporite de leziuni și violență neintenționate, abuz de alcool și droguri, sarcină neintenționată și boli cu transmitere sexuală. Explicațiile neurobiologice și cognitive tradiționale pentru comportamentul adolescentului nu au reușit să țină seama de modificările neliniare ale comportamentului observate în perioada adolescenței, în raport cu copilăria și vârsta adultă. Această revizuire oferă o conceptualizare biologic plauzibilă a mecanismelor neuronale care stau la baza acestor modificări neliniare ale comportamentului, ca o reacție sporită la stimulente, în timp ce controlul impulsurilor este încă relativ imatur în această perioadă. Studii recente de imagini umane și animale oferă o bază biologică pentru acest punct de vedere, sugerând dezvoltarea diferențială a sistemelor de recompensă limbică în raport cu sistemele de control de sus în jos în timpul adolescenței în raport cu copilăria și vârsta adultă. Acest tipar de dezvoltare poate fi agravat la adolescenții cu predispoziție către asumarea riscurilor, crescând riscul pentru rezultate slabe.
Conform Centrului Național de Statistică în Sănătate, în Statele Unite există peste 13,000 decese de adolescenți în fiecare an. Aproximativ 70% din aceste decese rezultă din prăbușiri ale autovehiculelor, răni neintenționate, omucideri și sinucideri (Eaton și colab., 2006). Rezultatele Studiului Național de Comportament al Riscului pentru Tineret 2005 (YRBS) arată că adolescenții se angajează în comportamente care le crește probabilitatea de deces sau de boală conducând un vehicul după băut sau fără centura de siguranță, purtând arme, folosind substanțe ilegale și angajându-se în relații sexuale neprotejate. rezultând sarcini neintenționate și BTS, inclusiv infecție cu HIV (Eaton și colab., 2006). Aceste statistici evidențiază importanța înțelegerii alegerilor și acțiunilor riscante la adolescenți.
O serie de ipoteze cognitive și neurobiologice au fost postulate de ce adolescenții se angajează în comportament de alegere suboptimală. Într-o recenzie recentă a literaturii despre dezvoltarea creierului uman al adolescenților, Yurgelun-Todd (2007) sugerează că dezvoltarea cognitivă în anii adolescenței este asociată cu o eficiență progresivă mai mare a capacităților de control cognitiv. Această eficiență este descrisă ca dependentă de maturizarea cortexului prefrontal, așa cum este evidențiată de o activitate crescută în regiunile prefrontal focale (Rubia și colab., 2000; Tamm, Menon și Reiss, 2002) și diminuarea activității în regiunile cerebrale irelevante (Brown și colab., 2005; Durston și colab., 2006).
Acest model general, de control cognitiv îmbunătățit cu maturarea cortexului prefrontal, sugerează o creștere liniară a dezvoltării de la copilărie până la vârsta adultă. Cu toate acestea, alegerile și acțiunile suboptimale observate în adolescență reprezintă o schimbare neliniară a comportamentului care poate fi deosebită de copilărie și vârstă adultă, așa cum a demonstrat Centrul Național de Statistică în Sănătate privind comportamentul și mortalitatea adolescenților. Dacă controlul cognitiv și o cortexă prefrontală imatură au stat la baza comportamentului de alegere suboptimală, atunci copiii ar trebui să arate remarcabil de asemănători sau chiar mai răi decât adolescenții, având în vedere cortexul prefrontal mai puțin dezvoltat și abilitățile cognitive. Astfel, funcția prefrontală imatură singură, nu poate contabiliza comportamentul adolescentului.
O conceptualizare precisă a schimbărilor cognitive și neurobiologice în timpul adolescenței trebuie să trateze adolescența ca o perioadă de dezvoltare tranzitorie (Spear, 2000), mai degrabă decât o singură instantanee în timp (Casey, Tottenham, Liston și Durston, 2005). Cu alte cuvinte, pentru a înțelege această perioadă de dezvoltare, tranzițiile în adolescență și în afara acesteia sunt necesare pentru a distinge atribute distincte ale acestei etape de dezvoltare. Stabilirea traiectoriilor de dezvoltare pentru procesele cognitive și neuronale este esențială pentru caracterizarea acestor tranziții și constrângerea interpretărilor despre schimbările de comportament în această perioadă. La nivel cognitiv sau comportamental, adolescenții sunt caracterizați ca impulsivi (adică, lipsiți de control cognitiv) și de asumare a riscurilor cu aceste constructe utilizate sinonim și fără apreciere pentru traiectoriile de dezvoltare distincte ale fiecăruia. La nivel neurobiologic, studiile de imagini umane și animale sugerează baze neurobiologice distincte și traiectorii de dezvoltare pentru sistemele neuronale care stau la baza acestor construcții separate de control al impulsurilor și decizii riscante.
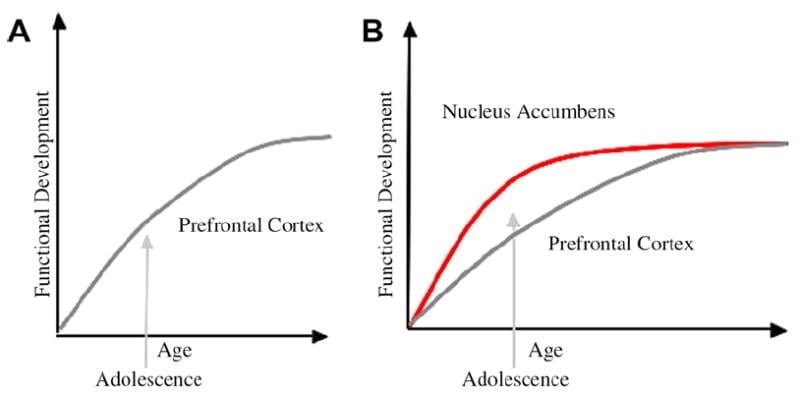
Am dezvoltat un model neurobiologic al dezvoltării adolescenților în acest cadru care se bazează pe modele de rozătoare (Laviola, Adriani, Terranova și Gerra, 1999; Spear, 2000) și studiile recente privind imagistica adolescenței (Ernst și colab., 2005; Galvan, Hare, Voss, Glover și Casey, 2007; Galvan și colab., 2006). Fig. 1 mai jos prezintă acest model. În stânga este caracterizarea tradițională a adolescenței ca fiind legată aproape exclusiv de imaturitatea cortexului prefrontal. În dreapta este modelul nostru neurobiologic propus care ilustrează modul în care regiunile de control subcorticale și prefrontalice limbice trebuie să fie luate în considerare împreună. Caricatura ilustrează diferite traiectorii de dezvoltare pentru aceste sisteme, sistemele limbice dezvoltând mai devreme decât regiunile de control prefrontal. Conform acestui model, individul este părtinitor mai mult de regiunile limbice mature funcțional în perioada adolescenței (adică dezechilibrul limbicului în raport cu controlul prefrontal), în comparație cu copiii, pentru care aceste sisteme (adică, limbice și prefrontal) sunt în curs de dezvoltare; și comparativ cu adulții, pentru care aceste sisteme sunt pe deplin mature. Această perspectivă oferă o bază pentru schimbările neliniare ale comportamentului de-a lungul dezvoltării, datorită maturizării anterioare a acestui limbic în raport cu regiunea prefrontală de control de jos în jos mai puțin matură. Odată cu dezvoltarea și experiența, conectivitatea funcțională dintre aceste regiuni oferă un mecanism de control de sus în jos a acestor regiuni (Hare, Voss, Glover și Casey, 2007a). Mai mult, modelul conciliază contradicția statisticilor de sănătate privind comportamentul riscant în adolescență, cu observația astută de Reyna și Farley (2006) că adolescenții sunt capabili să rezoneze și să înțeleagă riscurile comportamentelor în care se implică. Conform modelului nostru, în situații emițioase din punct de vedere emoțional, sistemul limbic va câștiga peste sistemele de control, având în vedere maturitatea sa în raport cu sistemul de control prefrontal. Dovezi efectuate de studii de comportament și imagistică umană care susțin acest model sunt furnizate în contextul acțiunilor în contexte răsplătitoare și emoționaleGalvan și colab., 2006, 2007; Hare, Voss, Glover și Casey, 2007b; Hare și colab., 2007a). În plus, speculăm de ce creierul se poate dezvolta în acest fel și de ce unii adolescenți pot avea un risc mai mare de a lua decizii suboptimale care să conducă la rezultate mai slabe pe termen lung (Galvan și colab., 2007; Hare și colab., 2007b).
S-a sugerat că explicația tradițională a comportamentului adolescentului se datorează dezvoltării prelungite a cortexului prefrontal (A). Modelul nostru ia în considerare dezvoltarea cortexului prefrontal împreună cu regiunile limbice subcorticale (de exemplu, nucleus accumbens) care au fost implicate în alegeri și acțiuni riscante (B).
Dezvoltarea unui comportament orientat spre obiective
O piatră de temelie a dezvoltării cognitive este capacitatea de a suprima gândurile și acțiunile necorespunzătoare în favoarea celor orientate spre scopuri, în special în prezența stimulentelor convingătoare (Casey, Galvan și Hare, 2005; Casey și colab., 2000b; Casey, Thomas, David-son, Kunz și Franzen, 2002a; Casey, Tottenham și Fossella, 2002b). O serie de studii clasice de dezvoltare au arătat că această abilitate se dezvoltă de-a lungul copilăriei și adolescenței (Caz, 1972; Flavell, Feach și Chinsky, 1966; Keating și Bobbitt, 1978; Pascual-Leone, 1970). Câțiva teoreticieni au susținut că dezvoltarea cognitivă se datorează creșterii vitezei și eficienței procesării și nu datorită creșterii capacității mentale (de ex. Bjorkland, 1985; Bjorkland, 1987; Caz, 1985). Alți teoreticieni au inclus construcția proceselor „inhibitoare” în contul lor de dezvoltare cognitivă (Harnishfeger & Bjorkland, 1993). Conform acestui raport, cogniția imatură este caracterizată de susceptibilitatea la interferențe din surse concurente care trebuie suprimate (de ex. Brainerd & Reyna, 1993; Casey, Thomas, Davidson, Kunz și Franzen, 2002a; Dempster, 1993; Diamond, 1985; Munakata & Yerys, 2001). Astfel, comportamentul orientat spre obiectiv necesită controlul impulsurilor sau întârzierea satisfacției pentru optimizarea rezultatelor și această abilitate pare să se maturizeze de-a lungul copilăriei și adolescenței.
Comportamentul adolescentului a fost descris ca impulsiv și riscant, aproape sinonim, totuși aceste construcții se bazează pe diferite procese cognitive și neuronale, care sugerează construcții distincte cu traiecții de dezvoltare diferite. Mai exact, o revizuire a literaturii sugerează că impulsivitatea scade odată cu vârsta de-a lungul copilăriei și adolescenței (Casey și colab., 2002a; Casey, Galvan și colab., 2005; Galvan și colab., 2007) și este asociat cu dezvoltarea prelungită a cortexului prefrontal (Casey, Galvan și colab., 2005), deși există diferențe în ceea ce privește gradul în care un anumit individ este impulsiv sau nu, indiferent de vârstă.
Spre deosebire de controlul impulsului / cognitiv, asumarea riscurilor pare să crească în perioada adolescenței în raport cu copilăria și vârsta adultă și este asociată cu sisteme subcorticale cunoscute a fi implicate în evaluarea recompenselor. Studiile de imagistică umană care vor fi revizuite sugerează o creștere a activării subcorticale (de exemplu, obișnuiți) atunci când faceți alegeri riscante (Kuhnen și Knutson, 2005; Matthews și colab., 2004; Montague & Berns, 2002) care este exagerat la adolescenți, în raport cu copiii și adulții (Ernst și colab., 2005; Galvan și colab., 2006). Aceste descoperiri sugerează diferite traiectorii pentru comportamentul bazat pe recompense sau stimulente, cu dezvoltarea anterioară a acestor sisteme în raport cu sistemele de control care arată un curs de dezvoltare prelungit și liniar, în ceea ce privește alegerile și acțiunile inadecvate imperative în favoarea celor direcționate prin obiectiv.
Dovezi din studii de neuroimagistică asupra dezvoltării creierului uman
Investigațiile recente ale dezvoltării creierului adolescenților s-au bazat pe progrese în metodologiile de neuroimagistică care pot fi ușor utilizate cu populațiile umane în curs de dezvoltare. Aceste metode se bazează pe metode de imagistica prin rezonanță magnetică (RMN) Fig. 2) și includ: RMN structural, care este utilizat pentru a măsura dimensiunea și forma structurilor; RMN funcțional, care este utilizat pentru a măsura tiparele activității creierului; și imagistica cu tensiune de difuzie (DTI), care este utilizată pentru a indexa conectivitatea traiectelor de fibre de materie albă. Dovada pentru modelul nostru de dezvoltare a competiției între regiunile corticale și subcorticale este susținută de conectivitatea structurală și funcțională imatură, măsurată de DTI și, respectiv, de RMN.
Studii RMN de dezvoltare a creierului uman
Mai multe studii au utilizat RMN-ul structural pentru a cartografia cursul anatomic al dezvoltării normale a creierului (vezi analiza de Durston și colab., 2001). Deși dimensiunea totală a creierului este de aproximativ 90% din dimensiunea adultă până la vârsta de șase ani, subcomponentele de culoare gri și alb ale creierului continuă să sufere modificări dinamice de-a lungul adolescenței. Datele din studiile RMN longitudinale recente indică faptul că volumul de materie cenușie are un model în formă de U inversat, cu o variație regională mai mare decât materia albă (Giedd, 2004; Gogtay și colab., 2004; Sowell și colab., 2003; Sowell, Thompson și Toga, 2004). În general, regiunile care subordonează funcțiile primare, cum ar fi sistemele motorii și senzoriale, se maturizează mai devreme; ariile de asociere mai înalte, care integrează aceste funcții primare, se maturizează mai târziu (Gogtay și colab., 2004; Sowell, Thompson și Toga, 2004). De exemplu, studiile care utilizează MRI bazate pe măsuri arată că pierderea corticală a substanței cenușii apare mai devreme în zonele senzorimotorii primare și mai târziu în cortexul temporal prefrontal dorsolateral și lateral (Gogtay și colab., 2004). Acest tipar este în concordanță cu studiile primare non-umane și postmortem uman care arată că cortexul prefrontal este una din ultimele regiuni ale creierului care trebuie maturizate (Bourgeois, Goldman-Rakic și Rakic, 1994; Huttenlocher, 1979). Spre deosebire de substanța cenușie, volumul materiei albe crește într-un model aproximativ liniar, crescând pe tot parcursul dezvoltării până la vârsta adultă (Gogtay și colab., 2004). Aceste modificări reflectă probabil mielinizarea continuă a axonilor prin oligodendrocite sporind conducerea neuronală și comunicarea.
Deși s-a acordat mai puțină atenție regiunilor subcorticale atunci când se examinează modificările structurale, unele dintre cele mai mari schimbări ale creierului de-a lungul dezvoltării sunt observate în aceste regiuni, în special în ganglionii bazali (Sowell și colab., 1999, A se vedea Fig. 3) și în special la bărbați (Giedd și colab., 1996). Modificările de dezvoltare ale volumului structural în ganglionii bazali și regiunile prefrontal sunt interesante în lumina proceselor de dezvoltare cunoscute (de exemplu, arborizarea dendritică, moartea celulelor, tăierea sinaptică, mielinizarea) care apar în copilărie și adolescență. Aceste procese permit reglarea fină și întărirea conexiunilor între regiunile prefrontal și subcortical cu dezvoltarea și învățarea care pot coincide cu un control cognitiv mai mare. Cum se raportează aceste modificări structurale la modificările cognitive? O serie de studii au legat maturizarea structurală a lobului frontal și funcția cognitivă folosind măsuri neuropsihologice și cognitive (de ex. Sowell și colab., 2003). În mod specific, asociațiile au fost raportate între volumele regionale ale ganglionilor pre-frontali și ganglioni bazali pre-frontali RMN și măsurile de control cognitiv (adică, capacitatea de a trece peste un răspuns necorespunzător în favoarea altuia sau de a suprima atenția asupra atributului de stimulare irelevantă în favoarea atributului stimulativ relevant (Casey, Trainor și colab., 1997). Aceste descoperiri sugerează că modificările cognitive sunt reflectate în modificările structurale ale creierului și subliniază importanța dezvoltării subcorticale (ganglionii bazali), precum și a dezvoltării corticale (de exemplu, cortexul prefrontal).

Studii DTI privind dezvoltarea creierului uman
Studiile de morfometrie pe baza RMN-ului revizuite sugerează că conexiunile corticale sunt adaptate cu eliminarea unei supraabundențe a sinapselor și consolidarea conexiunilor relevante cu dezvoltarea și experiența. Progresele recente în tehnologia RMN, cum ar fi DTI, oferă un instrument potențial pentru examinarea rolului unor tracte specifice de materie albă în dezvoltarea creierului și a comportamentului cu mai multe detalii. Relevante pentru această lucrare sunt studiile de neuroimagistică care au legat dezvoltarea tractului de fibre cu îmbunătățirea capacității cognitive. Mai exact, au fost arătate asocieri între măsurile bazate pe DTI pentru dezvoltarea prefrontală a materiei albe și controlul cognitiv la copii. Într-un studiu, dezvoltarea acestei capacități a fost corelată pozitiv cu traiectele de fibre prefrontal-parietale (Nagy, Westerberg și Klingberg, 2004) în concordanță cu studiile funcționale de neuroimagistică care arată recrutarea diferențială a acestor regiuni la copii în raport cu adulții.
Folosind o abordare similară, Liston și colab. (2005) au arătat că traiectele de substanțe albe dintre ganglionii prefrontal-bazali și traiectele de fibre anterioare continuă să se dezvolte de-a lungul copilăriei până la vârsta adultă, dar numai acele tracturi între cortexul prefrontal și ganglionii bazali sunt corelate cu controlul impulsului, măsurat prin performanță în mers / nogo sarcină. Traiectele pre-frontale de fibre au fost definite de regiuni de interese identificate într-un studiu RMN folosind aceeași sarcină. Pe ambele studii de dezvoltare DTI, măsurile tractului fibros au fost corelate cu dezvoltarea, dar specificitatea tracturilor de fibre particulare cu performanța cognitivă a fost demonstrată prin disocierea tractului particular (Liston și colab., 2005) sau capacitatea cognitivă (Nagy și colab., 2004). Aceste constatări subliniază importanța examinării nu numai a modificărilor regionale, ci a circuitelor atunci când se fac afirmații cu privire la modificările dependente de vârstă în substraturile neuronale ale dezvoltării cognitive.
Studii IRM funcționale de dezvoltare a comportamentului și creierului
Deși schimbările structurale măsurate de RMN și DTI au fost asociate cu modificări de comportament în timpul dezvoltării, o abordare mai directă pentru examinarea asocierii structurii - funcției este de a măsura schimbările în creier și comportament simultan, ca și în cazul RMN-ului. Capacitatea de a măsura schimbările funcționale în creierul în curs de dezvoltare cu RMN are potențial semnificativ în domeniul științei dezvoltării. În contextul lucrării actuale, fMRI oferă un mijloc pentru constrângerea interpretărilor comportamentului adolescentului. După cum s-a spus anterior, dezvoltarea cortexului prefrontal se crede că joacă un rol important în maturizarea abilităților cognitive superioare, cum ar fi luarea deciziilor și controlul cognitiv (Casey, Tottenham și Fossella 2002b; Casey, Trainor și colab., 1997). Multe paradigme au fost utilizate, împreună cu RMN-ul, pentru a evalua baza neurobiologică a acestor abilități, inclusiv sarcinile de flanker, Stroop și go / nogo (Casey, Castellanos și colab., 1997; Casey, Giedd și Thomas, 2000a; Durston și colab., 2003). Colectiv, aceste studii arată că copiii recrutează regiuni prefrontale distincte, dar deseori mai mari, mai difuze atunci când îndeplinesc aceste sarcini decât adulții. Modelul de activitate din cadrul regiunilor creierului central pentru performanța sarcinii (adică, care se corelează cu performanța cognitivă) devine mai focalizat sau mai bine ajustat cu vârsta, în timp ce regiunile care nu sunt corelate cu performanța sarcinii scad în activitate cu vârsta. Acest model a fost observat pe ambele secțiuni transversale (Brown și colab., 2005) și studii longitudinale (Durston și colab., 2006) și peste o varietate de paradigme. Deși studiile de neuroimagistică nu pot caracteriza definitiv mecanismul unor astfel de modificări de dezvoltare (de exemplu, arborizarea dendritică, tăierea sinaptică), rezultatele reflectă dezvoltarea și perfecționarea proiecțiilor către și de la, din regiunile cerebrale activate cu maturizare și sugerează că aceste modificări apar peste o prelungire perioada de timp (Brown și colab., 2005; Bunge, Dudukovic, Thomason, Vaidya și Gabrieli, 2002; Casey, Trainor și colab., 1997; Casey și colab., 2002a; Crone, Donohue, Honomichl, Wendelken și Bunge, 2006; Luna și colab., 2001; Moses și colab., 2002; Schlaggar și colab., 2002; Tamm și colab., 2002; Thomas și colab., 2004; Turkeltaub, Gareau, Flowers, Zeffiro și Eden, 2003).
Cum ne poate informa această metodologie dacă adolescenții nu au într-adevăr un control cognitiv suficient (impulsiv) sau sunt riscanți în alegerile și acțiunile lor? Controlul impulsului măsurat prin sarcini de control cognitiv, cum ar fi sarcina go / nogo, arată un model liniar de dezvoltare în copilărie și adolescență, așa cum este descris mai sus. Cu toate acestea, studii recente de neuroimagistică au început să examineze procesarea referitoare la recompense specifice asumării riscurilor la adolescenți (Bjork și colab., 2004; Ernst și colab., 2005; May și colab., 2004). Aceste studii s-au concentrat în principal pe regiunea accumbens, o porțiune a ganglionilor bazali implicați în prezicerea recompensei, mai degrabă decât caracterizarea dezvoltării acestei regiuni în combinație cu regiunile de control de sus în jos (cortexul prefrontal). Deși a fost arătat un raport recent privind activitatea prefrontală mai puțin ventrală la adolescenți în raport cu adulții în timpul unei sarcini de luare a deciziilor monetare privind comportamentul de asumare a riscurilor (Eshel, Nelson, Blair, Pine și Ernst, 2007).
În general, puține studii au examinat modul în care evoluția circuitelor de recompense în regiunile subcorticale (de exemplu, acumbens) se modifică în legătură cu dezvoltarea regiunilor prefrontal corticale. Mai mult decât atât, modul în care aceste schimbări neuronale coincid cu căutarea recompenselor, impulsivitatea și comportamentele de asumare a riscurilor rămâne relativ necunoscut. Modelul nostru neurobiologic propune că combinația de reacție sporită la recompense și imaturitate în zonele de control comportamental poate prejudicia adolescenții să caute imediat, mai degrabă decât câștiguri pe termen lung, explicând probabil creșterea lor în luarea deciziilor riscante și comportamentele impulsive. Urmărirea dezvoltării decizionale subcorticale (de exemplu, accumbens) și corticală (de exemplu, prefrontală) de-a lungul copilăriei până la vârsta adultă, oferă constrângeri suplimentare dacă modificările raportate la adolescență sunt specifice acestei perioade de dezvoltare sau reflectă maturizarea care se produce constant la o un model oarecum liniar de la copilărie până la vârsta adultă.
Dovezi empirice dintr-un studiu recent RMN ajută la susținerea modelului nostru neurobiologic și adoptă o abordare tranzitorie a înțelegerii adolescenței prin examinarea schimbărilor anterioare și următoare adolescenței. In acest studiu (Galvan și colab., 2006), am examinat răspunsurile comportamentale și neuronale pentru a răsplăti manipulările de-a lungul dezvoltării, punându-ne accent pe regiunile cerebrale implicate în învățarea și comportamentul legate de recompensă la animal (Hikosaka și Watanabe, 2000; Pecina, Cagniard, Berridge, Aldridge și Zhuang, 2003; Schultz, 2006) și studii de imagistică pentru adulți (de ex. Knutson, Adams, Fong și Hommer, 2001; O, Doherty, Kringelbach, Rolls, Hornak, Andrews, 2001; Zald și colab., 2004) și în studii de dependență (Hyman și Malenka, 2001; Volkow și Li, 2004). Pe baza modelelor de rozătoare (Laviola și colab., 1999; Spear, 2000) și imagistica anterioară (Ernst și colab., 2005), am emis ipoteza că, în raport cu copiii și adulții, adolescenții ar manifesta activarea exagerată a persoanelor obișnuite, în concordanță cu recrutarea mai puțin matură a regiunilor de control prefrontal de sus în jos. Lucrările recente care prezintă o conectivitate funcțională întârziată între aceste regiuni subcorticale prefrontală și limbică în adolescență în raport cu adulții, oferă un mecanism pentru lipsa controlului de sus în jos a acestor regiuni (Hare și colab., 2007a).
Rezultatele noastre au fost în concordanță cu modelele de rozătoare (Laviola, Macri, Morley-Fletcher și Adriani, 2003) și studiile anterioare privind imagistica (Ernst și colab., 2005) sugerează o activitate accentuată a recompenselor pentru recompense în timpul adolescenței. Într-adevăr, în raport cu copiii și adulții, adolescenții au arătat un răspuns exagerat de accumbens în așteptarea recompensei. Cu toate acestea, atât copiii, cât și adolescenții au arătat un răspuns mai puțin matur în regiunile de control prefrontal decât la adulți. Aceste descoperiri sugerează diferite trasee de dezvoltare pentru aceste regiuni, care pot sta la baza îmbunătățirii activității obișnuite, în raport cu copiii sau adulții, care, la rândul lor, pot avea legătură cu comportamentele impulsive și riscante crescute observate în această perioadă de dezvoltare (vezi Fig. 4).

Recrutarea diferențială a regiunilor prefrontală și subcorticală a fost raportată la o serie de studii IRM în dezvoltare (Casey și colab., 2002b; Monk și colab., 2003; Thomas și colab., 2004). În mod tipic, aceste constatări au fost interpretate mai degrabă în termeni de regiuni prefrontale imature decât dezechilibre între dezvoltarea regională prefrontală și subcorticală. Având în vedere dovezile regiunilor prefrontale în ghidarea acțiunilor adecvate în diferite contexte (Miller și Cohen, 2001), activitatea prefrontală imatură ar putea împiedica o estimare adecvată a rezultatelor viitoare și o evaluare a alegerilor riscante și, prin urmare, ar putea fi mai puțin influentă în evaluarea recompensei decât accumbens. Acest model este în concordanță cu cercetările anterioare care arată o creștere subcorticală față de activitatea corticală atunci când deciziile sunt părtinitoare prin câștiguri imediate pe termen lung (McClure, Laibson, Loewenstein și Cohen, 2004). Mai mult, s-a demonstrat că activitățile obișnuite cu fMRI se corelează pozitiv cu comportamentele ulterioare de asumare a riscurilor (Kuhnen și Knutson, 2005). În timpul adolescenței, în raport cu copilăria sau vârsta adultă, cortexul prefrontal ventral imatur poate să nu asigure controlul suficient de sus în jos al regiunilor de procesare a recompenselor puternic activate (de exemplu, accumbens), ceea ce duce la o influență mai mică a sistemelor prefrontal (cortexul orbitofrontal) în raport cu obișnuiții recompensați evaluare.
De ce ar fi programat creierul să se dezvolte astfel?
Adolescența este perioada de tranziție dintre copilărie și vârstă adultă care apare adesea cu pubertatea. Pubertatea marchează începuturile maturizării sexuale (Graber & Brooks-Gunn, 1998) și pot fi definite prin markeri biologici. Adolescența poate fi descrisă ca o tranziție progresivă la vârsta adultă cu un curs de timp ontogenetic nebulos (Spear, 2000). Evoluțional vorbind, adolescența este perioada în care sunt dobândite abilități de independență pentru a crește succesul la separarea de protecția familiei, deși crește șansele de circumstanțe dăunătoare (de exemplu, vătămare, depresie, anxietate, consum de droguri și dependență (Kelley, Schochet și Landry, 2004). Comportamentele care doresc independența sunt predominante la specii, cum ar fi creșterile interacțiunilor sociale direcționate de la egal la egal și intensificări în comportamentele în căutare de noutăți și în asumarea riscurilor. Factorii psihosociali au impact asupra înclinației adolescenților pentru un comportament riscant. Cu toate acestea, comportamentul riscant este produsul unui dezechilibru determinat biologic între o noutate sporită și căutarea senzațiilor în combinație cu „competența de autoreglare” imatură (Steinberg, 2004). Datele noastre neurobiologice sugerează că acest lucru se întâmplă prin dezvoltarea diferențială a acestor două sisteme (limbic și control).
Speculațiile ar sugera că acest model de dezvoltare este o caracteristică evolutivă. Trebuie să te angajezi într-un comportament cu risc ridicat pentru a părăsi familia și satul pentru a găsi un partener și asumarea riscurilor în același timp în care hormonii îi determină pe adolescenți să caute parteneri sexuali. În societatea actuală, când adolescența se poate extinde la nesfârșit, copiii trăind cu părinții și au dependență financiară și își aleg partenerii mai târziu în viață, această evoluție poate fi considerată nepotrivită.
Există dovezi în ceea ce privește speciile pentru căutarea de noutăți și pentru asumarea riscurilor în perioada adolescenței. Căutarea unor colegi de aceeași vârstă și lupta cu părinții, care toate ajută la îndepărtarea adolescentului de pe teritoriul natal pentru împerechere se observă în alte specii, inclusiv rozătoare, primate neumane și unele păsări (Spear, 2000). În raport cu adulții, șobolanii periadolescenți prezintă comportamente crescute în căutare de noutăți într-o paradigmă de noutate la alegere liberă (Laviola și colab., 1999). Dovezile neurochimice indică faptul că echilibrul din creierul adolescent între sistemele de dopamină corticală și subcorticală începe să se deplaseze către niveluri mai mari de dopamină corticală în timpul adolescenței (Spear, 2000). S-a arătat și conservarea dopaminergică similară prin adolescență la vârsta adultă și în cortexul prefrontal primat non-uman (Rosenberg și Lewis, 1995). Astfel, această asumare crescută a riscurilor pare să fie pe specii și are scopuri adaptative importante.
Predispoziții biologice, dezvoltare și risc
Diferențele individuale în controlul impulsurilor și în asumarea riscurilor sunt recunoscute în psihologie de ceva timp (Benthin, Slovic și Severson, 1993). Poate unul dintre exemplele clasice ale diferențelor individuale raportate în aceste abilități în literatura de psihologie socială, cognitivă și de dezvoltare este întârzierea satisfacției (Mischel, Shoda și Rodriguez, 1989). Întârzierea mulțumirii este de obicei evaluată la copii mici de 3- la 4-ani. Copilul este întrebat dacă ar prefera o recompensă mică (un cookie) sau o recompensă mare (două fursecuri). Copilului i se spune apoi că experimentatorul va părăsi camera pentru a se pregăti pentru activitățile viitoare și îi explică copilului că, dacă rămâne pe scaunul ei și nu mănâncă o fursecă, va primi recompensa mare. Dacă copilul nu poate sau nu poate aștepta, ar trebui să sune un clopot pentru a chema experimentatorul și, prin urmare, va primi recompensa mai mică. Odată ce este clar, copilul înțelege sarcina, este așezat la masă cu cele două recompense și clopoțelul. Distracțiile în cameră sunt reduse la minimum, fără jucării, cărți sau imagini. Experimentatorul revine după 15 min sau după ce copilul a sunat clopotul, a mâncat recompensele sau a arătat orice semne de suferință. Mischel a arătat că, de obicei, copiii se comportă într-unul din cele două moduri: (1) fie sună clopotul aproape imediat pentru a avea cookie-ul, ceea ce înseamnă că primesc doar unul; sau (2) așteaptă și își optimizează câștigurile și primesc ambele cookie-uri. Această observație sugerează că unii indivizi sunt mai buni decât alții în capacitatea lor de a controla impulsurile în fața stimulentelor extrem de importante și această părtinire poate fi detectată în copilărie timpurie (Mischel și colab., 1989) și par să rămână de-a lungul adolescenței și a vârstei tinere (Eigsti și colab., 2006).
Ce ar putea explica diferențele individuale în luarea deciziilor și comportamentul optim? Unii teoreticieni au postulat că circuitele mezolimbice dopaminergice, implicate în procesarea recompenselor, stau la baza unui comportament riscant. Diferențele individuale în acest circuit, cum ar fi variantele alelice la genele legate de dopamină, care rezultă în prea puțină sau prea multă dopamină în regiunile subcorticale, ar putea avea legătură cu înclinația de a se implica într-un comportament riscant (O'Doherty, 2004). S-a dovedit că nucleul accumbens crește în activitate imediat înainte de a face alegeri riscante pe paradigme de risc monetar (Kuhnen și Knutson, 2005; Matthews și colab., 2004; Montague & Berns, 2002) și așa cum s-a descris anterior, adolescenții arată o activitate exagerată a celor obișnuiți cu rezultate satisfăcătoare în raport cu copiii sau adulții (Ernst și colab., 2005; Galvan și colab., 2006). În mod colectiv, aceste date sugerează că adolescenții pot fi mai predispuși la alegeri riscante ca grup (Gardener & Steinberg, 2005), dar unii adolescenți vor fi mai predispuși decât alții să se angajeze în comportamente riscante, punându-le într-un risc potențial mai mare pentru rezultate negative. Prin urmare, este important să luăm în considerare variabilitatea individuală atunci când examinăm relații complexe de comportament cerebral legate de preluarea de riscuri și procesarea recompenselor în populațiile în curs de dezvoltare.
Pentru a explora diferențele individuale în comportamentul de asumare a riscurilor, Galvan și colab. (2007) recent a examinat asocierea dintre activitatea în circuitele neuronale legate de recompense, în așteptarea unei mari recompense monetare, cu măsuri de caracteristică ale asumării riscurilor și impulsivității în adolescență. Imagistica prin rezonanță magnetică funcțională și scale de evaluare anonime ale auto-raportului de comportament riscant, percepție a riscului și impulsivitate au fost dobândite la indivizii cu vârsta între 7 și 29 ani. A existat o asociere pozitivă între activitatea celor obișnuiți și probabilitatea implicării în comportament riscant de-a lungul dezvoltării. Această activitate a variat în funcție de evaluările indivizilor privind consecințele pozitive sau negative anticipate ale unui astfel de comportament. Acei indivizi care au perceput comportamente riscante ca ducând la consecințe grave, au activat persoanele fizice mai puțin recompensate. Această asociere a fost condusă în mare parte de copii, adulții evaluând consecințele unui astfel de comportament posibil. Evaluările de impulsuri nu au fost asociate cu activitatea obișnuită, ci mai degrabă cu vârsta. Aceste descoperiri sugerează că, în adolescență, unii indivizi pot fi mai predispuși să se angajeze în comportamente riscante din cauza schimbărilor de dezvoltare în concordanță cu variabilitatea în predispoziția unui individ dat să se angajeze într-un comportament riscant, mai degrabă decât la simple schimbări în impulsivitate (vezi Fig. 5).

Comportamentul adolescentului a fost în mod repetat caracterizat ca impulsiv și riscant (Steinberg, 2004, 2007), cu toate acestea, această revizuire a literaturii imagistice sugerează diferite substraturi neurobiologice și diferite traiectorii de dezvoltare pentru aceste comportamente. Mai exact, impulsivitatea este asociată cu dezvoltarea prefrontală ventrală imatură și scade treptat de la copilărie la vârsta adultă (Casey, Galvan și colab., 2005). Corelația negativă între cotele de impulsivitate și vârstă din studiu Galvan și colab. (2007) susține în continuare această noțiune. În schimb, asumarea riscurilor este asociată cu o creștere a activității obișnuite (Kuhnen și Knutson, 2005; Matthews și colab., 2004; Montague & Berns, 2002), care este exagerat la adolescenți, în raport cu copiii și adulții (Ernst și colab., 2005; Galvan și colab., 2006). Astfel, alegerile și comportamentul adolescenților nu pot fi explicate doar prin impulsivitate sau prin dezvoltarea lungă a cortexului prefrontal, deoarece copiii ar putea fi preconizați mai mari de riscuri. Rezultatele oferă o bază neurală pentru motivul pentru care unii adolescenți prezintă un risc mai mare decât alții, dar oferă o bază pentru modul în care comportamentul adolescentului este diferit de copiii și adulții în asumarea riscurilor.
Colectiv, aceste date sugerează că, deși adolescenții ca grup sunt considerați consumatori de risc (Gardener & Steinberg, 2005), unii adolescenți vor fi mai predispuși decât alții să se angajeze în comportamente riscante, punându-le într-un risc potențial mai mare pentru rezultate negative. Aceste constatări subliniază importanța luării în considerare a variabilității individuale atunci când examinăm relații complexe de comportament cerebral legate de preluarea riscurilor și procesarea recompenselor în populațiile în curs de dezvoltare. Mai mult, aceste diferențe individuale și de dezvoltare pot ajuta la explicarea vulnerabilității la unii indivizi la asumarea riscurilor asociate cu consumul de substanțe și, în final, dependența.
Concluzii
Studiile de imagistică umană arată modificări structurale și funcționale în regiunile frontostriatale (Giedd și colab., 1996, 1999; Jernigan și colab., 1991; Sowell și colab., 1999; pentru revizuire, Casey, Galvan și colab., 2005) care par paralele cu creșterea controlului cognitiv și a auto-reglării (Casey, Trainor și colab., 1997; Luna & Sweeney, 2004; Luna și colab., 2001; Rubia și colab., 2000; Steinberg, 2004, A se vedea, de asemenea, Steinberg, 2008, această problemă). Aceste schimbări par să arate o schimbare în activarea regiunilor prefrontal de la recrutarea difuză la mai focală în timp (Brown și colab., 2005; Bunge și colab., 2002; Casey, Trainor și colab., 1997; Durston și colab., 2006; Moses și colab., 2002) și recrutarea crescută a regiunilor subcortice în timpul adolescenței (Casey și colab., 2002a; Durston și colab., 2006; Luna și colab., 2001). Deși studiile de neuroimagistică nu pot caracteriza definitiv mecanismul unor astfel de schimbări de dezvoltare, aceste modificări ale volumului și structurii pot reflecta dezvoltarea în interiorul și rafinarea proiecțiilor către și dinspre aceste regiuni ale creierului în timpul maturizării sugestive pentru reglarea fină a sistemului cu dezvoltarea.
Luate împreună, concluziile sintetizate aici indică faptul că un comportament crescut de asumare a riscurilor în adolescență este asociat cu traiectorii diferite de dezvoltare a plăcerii subcorticale și a regiunilor de control cortical. Aceste schimbări de dezvoltare pot fi agravate de diferențele individuale în activitatea sistemelor de recompense. Deși adolescența a fost distinsă ca o perioadă caracterizată prin comportamente de căutare a recompenselor și de asumare a riscurilor (Gardener & Steinberg, 2005; Spear, 2000) diferențele individuale în răspunsurile neuronale la recompensă, predispune unii adolescenți să-și asume mai multe riscuri decât alții, punându-i un risc mai mare pentru rezultate negative. Aceste constatări oferă fundamentul crucial prin sinteza diferitelor constatări legate de asumarea riscurilor în adolescență și prin înțelegerea diferențelor individuale și a markerilor de dezvoltare pentru înclinația de a se implica într-un comportament negativ.
recunoasteri
Această lucrare a fost susținută în parte de subvenții de la Institutul Național de Abuz de Droguri R01 DA18879 și Institutul Național de Sănătate Mintală 1P50 MH62196.
Referinte
- Benthin A, Slovic P, Severson H. Un studiu psihometric al percepției asupra riscului adolescent. Jurnalul adolescenței. 1993;16: 153-168. [PubMed]
- Bjork JM, Knutson B, Fong GW, Caggiano DM, Bennett SM, Hommer DW. Stimularea activării creierului la adolescenți: asemănări și diferențe față de adulții tineri. Revista de Neuroștiințe. 2004;24: 1793-1802. [PubMed]
- Bjorkland DF. Rolul cunoștințelor conceptuale în dezvoltarea organizației în memoria copiilor. În: Brainerd CJ, Pressley M, editori. Procesele de bază în dezvoltarea memoriei: Progresul în cercetarea dezvoltării cognitive. New York: Springer-Verlag; 1985. pp. 103-142.
- Bjorkland DF. Modul în care schimbările de vârstă contribuie la dezvoltarea memoriei copiilor: o revizuire interpretativă. Analiza dezvoltării. 1987;7: 93-130.
- Bourgeois JP, PS Goldman-Rakic, Rakic P. Synaptogenesis în cortexul prefrontal al maimuțelor rhesus. Cortex cerebral. 1994;4: 78-96. [PubMed]
- Brainerd CJ, Reyna VF. Independența memoriei și interferența memoriei în dezvoltarea cognitivă. Revizuirea psihologică. 1993;100: 42-67. [PubMed]
- Brown TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. Modificări evolutive în organizarea funcțională cerebrală umană pentru generarea de cuvinte. Cortex cerebral. 2005;15: 275-290. [PubMed]
- Bunge SA, Dudukovic NM, Thomason ME, Vaidya CJ, Gabrieli JD. Contribuții ale lobului frontal imatur la controlul cognitiv la copii: dovezi din RMN. Neuron. 2002;33: 301-311. [PubMed]
- Cazul R. Validarea unei construcții de capacități neo-piagetiene. Jurnalul Psihologiei Experimentale a Copilului. 1972;14: 287-302.
- Cazul R. Dezvoltare intelectuală: naștere până la vârsta adultă. New York: Academic Press; 1985.
- Casey BJ, Castellanos FX, Giedd JN, Marsh WL, Hamburger SD, Schubert AB. și colab. Implicarea circuitului frontostriatal drept în inhibarea răspunsului și tulburare de deficit de atenție / hiperactivitate. Jurnalul Academiei Americane de Psihiatrie a Copilului și Adolescentului. 1997;36: 374-383. [PubMed]
- Casey BJ, Galvan A, Hare TA. Schimbări în organizarea funcțională cerebrală în timpul dezvoltării cognitive. Opinia curentă în neurobiologie. 2005;15: 239-244. [PubMed]
- Casey BJ, Giedd JN, Thomas KM. Dezvoltarea creierului structural și funcțional și relația sa cu dezvoltarea cognitivă. Psihologie biologică. 2000a;54: 241-257. [PubMed]
- Casey BJ, Thomas KM, Davidson MC, Kunz K, Franzen PL. Disocierea funcției striatale și a hipocampului în mod evolutiv cu o sarcină de compatibilitate a răspunsului stimulului. Revista de Neuroștiințe. 2002a;22: 8647-8652. [PubMed]
- Casey BJ, Thomas KM, Welsh TF, Badgaiyan RD, Eccard CH, Jennings JR, și colab. Disocierea conflictului de răspuns, selecția atențională și speranța cu imagistică prin rezonanță magnetică funcțională. Proceedings of the National Academy of Science. 2000b;97: 8728-8733.
- Casey BJ, Tottenham N, Fossella J. Abordări clinice, imagistice, leziunii și genetice spre un model de control cognitiv. Psihologia dezvoltării. 2002b;40: 237-254. [PubMed]
- Casey BJ, Tottenham N, Liston C, Durston S. Imaginarea creierului în curs de dezvoltare: Ce am învățat despre dezvoltarea cognitivă? Tendințe în știința cognitivă. 2005;9: 104-110.
- Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN, și colab. Un studiu funcțional de IRM funcțional de activare prefrontală în timpul desfășurării unei sarcini go-no-go. Revista de Neuroștiințe cognitive. 1997;9: 835-847.
- Crone E, Donohue S, Honomichl R, Wendelken C, Bunge S. Regiunile cerebrale care mediază utilizarea flexibilă a regulilor în timpul dezvoltării. Revista de Neuroștiințe. 2006;26: 11239-11247. [PubMed]
- Dempster FN. Rezistența la interferențe: Schimbări de dezvoltare într-un mecanism de procesare de bază. În: Howe ML, Pasnak R, editori. Teme emergente în dezvoltarea cognitivă Volumul 1: Fundații. New York: Springer; 1993. pp. 3 – 27.
- Diamant A. Dezvoltarea abilității de a folosi rechemarea pentru a ghida acțiunea, așa cum este indicat de performanțele sugarilor în AB. Dezvoltarea copilului. 1985;56: 868-883. [PubMed]
- Durston S, Davidson MC, Thomas KM, Worden MS, Tottenham N, Martinez A, și colab. Manipularea parametrică a competiției de conflict și de răspuns, utilizând fMRI în legătură cu evenimente mixte rapide. Neuroimage. 2003;20: 2135-2141. [PubMed]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella J, și colab. Trecerea de la activitatea corticală difuză la focală cu dezvoltarea. Dezvoltare Știință. 2006;1: 18-20. [PubMed]
- Durston S, Hulshoff Pol HE, Casey BJ, Giedd JN, Buitelaar JK, van Engeland H. RMN anatomic al creierului uman în curs de dezvoltare: Ce am învățat? Journal of American Academy of Child Adolescent Psychiatry. 2001;40: 1012-1020.
- Eaton LK, Kinchen S, Ross J, Hawkins J, Harris WA, Lowry R și colab. Supravegherea comportamentului de risc pentru tineri - Statele Unite, 2005, rezumate ale supravegherii. Raportul săptămânal privind morbiditatea și mortalitatea. 2006;55: 1-108. [PubMed]
- Eigsti IM, Zayas V, Mischel W, Shoda Y, Ayduk O, Dadlani MB, și colab. Predicarea controlului cognitiv de la preșcolari până la adolescență târzie și la vârsta adultă. Știință psihologică. 2006;17: 478-484. [PubMed]
- Ernst M, Nelson EE, Jazbec S, McClure EB, Monk CS, Leibenluft E, și colab. Amygdala și nucleul accumbens în răspunsurile la primirea și omisiunea de câștiguri la adulți și adolescenți. Neuroimage. 2005;25: 1279-1291. [PubMed]
- Eshel N, Nelson EE, Blair RJ, Pine DS, Ernst M. Substraturi neurale de selecție la adulți și adolescenți: Dezvoltarea cortexului ventricular prefrontal și cingular anterior. Neuropsychologia. 2007;45: 1270-1279. [Articol gratuit PMC] [PubMed]
- Flavell JH, Feach DR, Chinsky JM. Repetare verbală spontană într-o sarcină de memorie în funcție de vârstă. Dezvoltarea copilului. 1966;37: 283-299. [PubMed]
- Galvan A, Hare TA, Parra CE, Penn J, Voss H, Glover G și colab. Dezvoltarea anterioară a accumbens în raport cu cortexul orbitofrontal ar putea sta la baza comportamentului de asumare a riscului la adolescenți. Revista de Neuroștiințe. 2006;26: 6885-6892. [PubMed]
- Galvan A, Hare T, Voss H, Glover G, Casey BJ. Asumarea riscului și creierul adolescentului: cine este în pericol? Dezvoltare Știință. 2007;10: F8-F14. [PubMed]
- Gardener M, Steinberg L. Influența de la egal la egal cu asumarea riscurilor, preferința riscului și luarea deciziilor riscante la adolescență și vârstă adultă: Un studiu experimental. Psihologia dezvoltării. 2005;41: 625-635. [PubMed]
- Giedd JN. Imagistica prin rezonanță magnetică structurală a creierului adolescent. Analele Academiei de Științe din New York. 2004;1021: 77-85. [PubMed]
- Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A, și colab. Dezvoltarea creierului în timpul copilăriei și adolescenței: studiu longitudinal RMN. Natură Neuroștiință. 1999;2: 861-863.
- Giedd JN, Snell JW, Lange N, Rajapakse JC, Casey BJ, Kozuch PL, și colab. Imagistica prin rezonanță magnetică cantitativă a dezvoltării creierului uman: vârsta 4 – 18. Cortex cerebral. 1996;6: 551-560. [PubMed]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, și colab. Cartografierea dinamică a dezvoltării corticale umane în timpul copilăriei până la vârsta adultă. Proceedings of the National Academy of Sciences din Statele Unite ale Americii. 2004;101: 8174-8179. [Articol gratuit PMC] [PubMed]
- Graber JA, Brooks-Gunn J. Puberty. În: Blechman EA, Brownell KD, editori. Medicină comportamentală și femei un manual cuprinzător. New York, NY: Guilford Press; 1998. pp. 51 – 58.
- Hare TA, Voss HU, Glover GH, Casey BJ. Creierul adolescentului și riscul potențial de anxietate și depresie. 2007a Trimis spre publicare.
- Hare TA, Voss HU, Glover GH, Casey BJ. Concurența între sistemele limbice prefrontal și subcorticale stau la baza reactivității emoționale în timpul adolescenței. 2007b Trimis spre publicare.
- Harnishfeger KK, Bjorkland F. Ontogenia mecanismelor de inhibare: o abordare reînnoită a dezvoltării cognitive. În: Howe ML, Pasnek R, editori. Teme emergente în dezvoltarea cognitivă. Voi. 1. New York: Springer-Verlag; 1993. pp. 28 – 49.
- Hikosaka K, Watanabe M. Activitatea de întârziere a neuronilor prefrontali orbitali și laterali ai maimuței variază cu recompense diferite. Cortex cerebral. 2000;10: 263-271. [PubMed]
- Huttenlocher PR. Densitatea sinaptică în cortexul frontal uman - modificări de dezvoltare și efecte ale îmbătrânirii. Cercetarea creierului. 1979;163: 195-205. [PubMed]
- Hyman SE, Malenka RC. Dependența și creierul: neurobiologia compulsiei și persistența acesteia. Nature Review Neuroscience. 2001;2: 695-703.
- Jernigan TL, Zisook S, Heaton RK, Moranville JT, Hesselink JR, Braff DL. Anomalii imagistice de rezonanță magnetică în nucleele lenticulare și cortexul cerebral în schizofrenie. Arhivele de psihiatrie generală. 1991;48: 811-823.
- Keating DP, Bobbitt BL. Diferențe individuale și de dezvoltare în componentele de procesare cognitivă ale abilităților mentale. Dezvoltarea copilului. 1978;49: 155-167.
- Kelley AE, Schochet T, Landry C. Analele Academiei de Științe din New York. 2004;1021: 27-32. [PubMed]
- Knutson B, Adams CM, Fong GW, Hommer D. Anticiparea unei recompense monetare crescătoare selectiv recrutează nucleul accumbens. Revista de Neuroștiințe. 2001;21: RC159. [PubMed]
- Kuhnen CM, Knutson B. Bazele neuronale ale riscului financiar. Neuron. 2005;47: 763-770. [PubMed]
- Laviola G, Adriani W, Terranova ML, Gerra G. Factori psiobiologici de risc pentru vulnerabilitatea la psihostimulanți la adolescenții umani și la modelele animale. Neuroștiințe și recenzii biobehaviorale. 1999;23: 993-1010. [PubMed]
- Laviola G, Macri S, Morley-Fletcher S, Adriani W. Comportamentul abstract de asumare a riscurilor la șoarecii adolescenți: determinanți psiobiologici și influență epigenetică timpurie. Neuroștiințe și recenzii biobehaviorale. 2003;27: 19-31. [PubMed]
- Liston C, Watts R, Tottenham N, Davidson MC, Niogi S, Ulug AM, și colab. Microstructura frontostriatală modulează recrutarea eficientă a controlului cognitiv. Cortex cerebral. 2005;16: 553-560. [PubMed]
- Luna B, Sweeney JA. Apariția funcției creierului colaborativ: studiile FMRI privind dezvoltarea inhibării răspunsului. Analele Academiei de Științe din New York. 2004;1021: 296-309. [PubMed]
- Luna B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ și colab. Maturarea funcției cerebrale distribuite pe scară largă susține dezvoltarea cognitivă. Neuroimage. 2001;13: 786-793. [PubMed]
- Matthews SC și colab. Activarea selectivă a nucleului accumbens în procesul de luare a deciziilor. Neuroreport. 2004;15: 2123-2127. [PubMed]
- May JC, Delgado MR, Dahl RE, Stenger VA, Ryan ND, Fiez JA, și colab. Imagistica prin rezonanță magnetică funcțională legată de eveniment a circuitelor cerebrale legate de recompensă la copii și adolescenți. Biologie psihiatrie. 2004;55: 359-366. [PubMed]
- McClure SM, Laibson DI, Loewenstein G, Cohen JD. Sistemele neuronale separate valorizează recompensele monetare imediate. Știință. 2004;306: 503-507. [PubMed]
- Miller EK, Cohen JD. O teorie integrativă a funcției cortexului prefrontal. Revizuirea anuală a neuroștiinței. 2001;24: 167-202.
- Mischel W, Shoda Y, Rodriguez MI. Întârzierea satisfacției la copii. Știință. 1989;244: 933-938. [PubMed]
- Monk CS, McClure EB, Nelson EE, Zarahn E, Bilder RM, Leibenluft E și colab. Imaturitatea adolescenților în implicarea creierului legată de atenție la expresiile faciale emoționale. Neuroimage. 2003;20: 420-428. [PubMed]
- Montague PR, Berns GS. Economia neuronală și substraturile biologice ale evaluării. Neuron. 2002;36: 265-284. [PubMed]
- Moses P, Roe K, Buxton RB, Wong EC, Frank LR, Stiles J. RMN funcțional de procesare globală și locală la copii. Neuroimage. 2002;16: 415-424. [PubMed]
- Munakata Y, Yerys BE. Toate împreună acum: Când disocierile dintre cunoaștere și acțiune dispar. Știința psihologică. 2001;12: 335-337.
- Nagy Z, Westerberg H, Klingberg T. Maturarea materiei albe este asociată cu dezvoltarea funcțiilor cognitive în timpul copilăriei. Revista de Neuroștiințe cognitive. 2004;16: 1227-1233. [PubMed]
- O'Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Reprezentări de recompensă și pedeapsă în cortexul orbitofrontal uman. Nature Neurosci. 2001;4: 95-102. [PubMed]
- O'Doherty JP. Reprezentări de recompensă și învățare legate de recompense în creierul uman: Perspective din neuroimagistică. Opinii actuale în neurobiologie. 2004;14: 769-776.
- Pascual-Leone JA. Un model matematic pentru tranziția în etapele de dezvoltare ale lui Piaget. Acta Psychologica. 1970;32: 301-345.
- Pecina S, Cagniard B, Berridge KC, Aldridge JW, Zhuang X. Șoarecii mutanți hiperdopaminergici au o "dorință" mai mare, dar nu "plac" pentru recompense dulci. Revista de Neuroștiințe. 2003;23: 9395-9402. [PubMed]
- Reyna VF, Farley F. Risc și raționalitate în procesul de luare a deciziilor adolescenților: implicații pentru teorie, practică și politici publice. Știința psihologică în interes public. 2006;7: 1-44.
- Rosenberg DR, Lewis DA. Maturarea postnatală a inervației dopaminergice a corticalelor prefrontale și motorii de maimuță: Analiza imunohistochimică a hidroxilazei tirozinei. Jurnalul de Neurologie Comparativa. 1995;358: 383-400. [PubMed]
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A și colab. Frontalizare funcțională cu vârsta: cartografierea traiectoriilor neurodezvoltate cu fMRI. Neuroștiințe și recenzii biobehaviorale. 2000;24: 13-19. [PubMed]
- Schlaggar BL, Brown TT, Lugar HM, Visscher KM, Miezin FM, Petersen SE. Diferențe neuroanatomice funcționale între adulți și copii de vârstă școlară în procesarea cuvintelor singulare. Știință. 2002;296: 1476-1479. [PubMed]
- Schultz W. Teoriile comportamentale și neurofiziologia recompensei. Recenzii anuale de psihologie. 2006;57: 87-115.
- Sowell ER, Peterson BS, Thompson PM, Bine ati venit SE, Henkenius AL, Toga AW. Cartografierea schimbărilor corticale pe toată durata vieții umane. Natură Neuroștiință. 2003;6: 309-315.
- Sowell ER, Thompson PM, Holmes CJ, Jernigan TL, Toga AW. Dovezi in vivo pentru maturizarea creierului post-adolescent în regiunile frontale și striatale. Natură Neuroștiință. 1999;2: 859-861.
- Sowell ER, Thompson, Toga AW. Maparea schimbărilor în cortexul uman pe toată durata vieții. Neurolog. 2004;10: 372-392. [PubMed]
- Spear LP. Creierul adolescent și manifestările comportamentale legate de vârstă. Neuroștiințe și recenzii biobehaviorale. 2000;24: 417-463. [PubMed]
- Steinberg L. Asumarea riscurilor în adolescență: Ce se schimbă și de ce? Analele Academiei de Științe din New York. 2004;1021: 51-58. [PubMed]
- Steinberg L. Asumarea riscurilor în adolescență: Noi perspective din știința creierului și a comportamentului. Direcții curente în știința psihologică. 2007;16: 55-59.
- Steinberg L. O perspectivă a neuroștiințelor sociale asupra riscului adolescentului. Analiza dezvoltării. 2008;28: 78-106. [Articol gratuit PMC] [PubMed]
- Tamm L, Menon V, Reiss AL. Maturarea funcției cerebrale asociată cu inhibarea răspunsului. Jurnalul Academiei Americane de Psihiatrie a Copilului și Adolescentului. 2002;41: 1231-1238. [PubMed]
- Thomas KM, Hunt RH, Vizueta N, Sommer T, Durston S, Yang Y, și colab. Dovada diferențelor de dezvoltare în învățarea secvenței implicite: Un studiu FMRI la copii și adulți. Revista de Neuroștiințe cognitive. 2004;16: 1339-1351. [PubMed]
- Turkeltaub PE, Gareau L, Florile DL, Zeffiro TA, Eden GF. Dezvoltarea mecanismelor neuronale pentru lectură. Natură Neuroștiință. 2003;6: 767-773.
- Volkow ND, Li TK. Dependența de droguri: neurobiologia comportamentului s-a deranjat. Nature Review Neuroscience. 2004;5: 963-970.
- Yurgelun-Todd D. Schimbări emoționale și cognitive în timpul adolescenței. Opinia curentă în neurobiologie. 2007;17: 251-257. [PubMed]
- Zald DH, Boileau I, El-Dearedy W, Gunn R, McGlone F, Dichter GS, și colab. Transmiterea dopaminei în striatum uman în timpul sarcinilor de recompensă monetară. Revista de Neuroștiințe. 2004;24: 4105-4112. [PubMed]
