- 1Centro Interdisciplinario de Neurociențe de Valparaíso, Facultatea de Științe, Universitatea de Valparaíso, Valparaíso, Chile
- 2Departamentul de Neurociencia, Facultatea de Medicină, Universitatea din Chile, Santiago, Chile
- 3Núcleo Milenio NUMIND Biologia tulburărilor neuropsihiatrice, Universitatea de Valparaíso, Valparaíso, Chile
- 4Departamentul de Biologie Celulară și Moleculară, Facultatea de Științe Biologice, Universitatea Católica Pontificia de Chile, Santiago, Chile
- 5Departamentul de Farmacie și Centrul Interdisciplinar de Neuroștiință, Facultatea de Chimie, Pontificia Universidad Católica de Chile, Santiago, Chile
Puterea comportamentelor orientate spre țintă este reglată de neuronii dopamină din creierul mijlociu. Disfuncțiile circuitelor dopaminergice sunt observate în dependența de droguri și tulburarea obsesiv-compulsivă. Comportamentul compulsiv este o caracteristică pe care ambele tulburări o împărtășesc, care este asociată cu o neurotransmisie crescută a dopaminei. Activitatea neuronilor dopaminei cerebrale mijlocii este reglementată în principal de acțiunea homeostatică a dopaminei prin intermediul receptorilor D2 (D2R) care scad arderea neuronilor, precum și sinteza și eliberarea dopaminei. Transmisia dopaminei este, de asemenea, reglată de sisteme de neurotransmițător heterolog, cum ar fi sistemul kappa opioid, printre altele. O mare parte din cunoștințele noastre actuale despre sistemul kappa opioid și influența sa asupra transmiterii dopaminei provin din modele preclinice de boli ale creierului. În 1988, folosind microdialysis cerebral, s-a arătat că activarea acută a Receptorilor de opioizi Kappa (KOR) scade nivelul sinaptic de dopamină în striat. Acest efect inhibitor al KOR se opune influențării facilitatoare a medicamentelor de abuz asupra eliberării dopaminei, ceea ce duce la propunerea utilizării agoniștilor KOR ca terapie farmacologică pentru aportul compulsiv de medicamente. Surprinzător, 30 de ani mai târziu, antagoniștii KOR sunt propuși în schimb să trateze dependența de droguri. Ce s-a putut întâmpla în acești ani care au generat această schimbare drastică a paradigmei? Dovada colectată a sugerat că efectul KOR asupra nivelurilor de dopamină sinaptică este complex, în funcție de frecvența de activare a KOR și de sincronizare cu alți stimuli de intrare la neuronii dopaminei, precum și de diferențele de sex și specii. Spre deosebire de efectul său acut, activarea cronică a KOR pare să faciliteze neurotransmisia dopaminei și comportamentele mediate de dopamină. Acțiunile opuse exercitate de activarea KOR acută versus cronică au fost asociate cu un efect aversiv inițial și cu o întârziere recompensantă, în timpul expunerii la droguri de abuz. Comportamentele compulsive induse de activarea repetată a D2R sunt, de asemenea, potențate de co-activarea susținută a KOR, care se corelează cu scăderea nivelurilor sinaptice de dopamină și D2R sensibilizată. Astfel, activarea KOR dependentă de timp are impact direct asupra nivelurilor de dopamină care afectează reglarea comportamentelor motivate. Această revizuire analizează contribuția sistemului kappa opioid la corelațiile dopaminergice ale comportamentelor compulsive.
Introducere
Sistemul dopaminergic în comportamentele compulsive
Compulsia este imposibilitatea de auto-oprire pentru a executa o acțiune obișnuită cu rezultat cunoscut, în ciuda consecințelor adverse (Robbins și colab., 2012). Comportamentele compulsive sunt repere ale tulburării obsesiv-compulsive (TOC) și dependenței de droguri, printre alte boli psihiatrice. Verificarea comportamentului este foarte frecventă în tulburările de spectru obsesiv-compulsive fiind caracterizată prin repetarea constantă a unei anumite rutine, într-un mod stereotip sau ritualic (Williams și colab., 2013). O gamă largă de comportamente normale (de exemplu, verificarea, curățarea, spălarea mâinilor etc.) se poate transforma în compulsiv la pacienții cu TOC și, în general, apare ca răspuns la gânduri obsesive și înfricoșătoare care induc anxietate. În mod similar, căutarea și consumul de droguri de abuz devin compulsive în dependenții de droguri La fel ca în TOC, anxietatea joacă un rol cheie care declanșează consumul compulsiv de droguri la consumatorii de droguri cu experiență. În ambele condiții se observă aceleași deficiențe în procesarea recompenselor și a pedepselor (Figee și colab., 2016), ceea ce a determinat unii autori să discute despre TOC ca o dependență comportamentală (Holden, 2001).
Un posibil mecanism care conduce la un comportament compulsiv este încadrat în teoria stimulării sensibilizării la dependență, care este aceea că o motivație amplificată („dorință”) pentru medicament se dezvoltă în timpul dependenței, fără a dezvolta un efect plăcut amplificat („plăcere”) (Berridge și colab., 1989; Berridge și Robinson, 2016). Sensibilizarea durabilă a circuitului de recompensare / motivație este implicată în inducerea stimulării-sensibilizării asociate cu consumul de droguri. Circuitul de recompensare / motivație este compus din neuroni de dopamină ale creierului mijlociu din substantia nigra (SN) și zona tegmentală ventrală (VTA), care vizează nivelurile dorsale și, respectiv, ventrale ale striatului. Neuronii Dopamina care se proiectează spre striatul ventral sau nucleul accumbens (NAc) au fost în mod tradițional legate de comportamentele orientate spre scopuri, în timp ce neuronii dopaminici care se proiectează spre striatul dorsal au fost asociați cu dobândirea obiceiurilor (Everitt și Robbins, 2005; Înțelept, 2009; Yager și colab., 2015; Volkow și colab., 2017).
Sensibilizarea circuitului de recompensare / motivație este observată la rozătoare ca creșterea treptată a activității locomotorii indusă de administrarea repetată a unei doze fixate cu un potențial dependență (Pierce și Kalivas, 1997; Robinson și Berridge, 2001). Sensibilizarea locomotorie este un fenomen durabil, deoarece se observă după săptămâni, luni și chiar un an după extragerea drogurilor (Robinson și Berridge, 1993). S-a sugerat din timp că sensibilizarea circuitului de recompensare / motivație contribuie la căutarea compulsivă a drogurilor (Robinson și Berridge, 1993). În consecință, sensibilizarea locomotorie facilitează auto-administrarea cocainei în căutarea reintegrării (De Vries și colab., 2002). Mai mult, șobolanii cu acces extins la auto-administrare de cocaină prezintă un răspuns locomotor mai mare la cocaină decât șobolanii cu acces limitat (Ferrario și colab., 2005). În plus, modificările neurochimice care stau la baza sensibilizării locomotorii la psiostimulanți sunt observate și în căutarea compulsivă a medicamentului (Steketee și Kalivas, 2011; Giuliano și colab., 2019). Aceste date susțin corespondența propusă timpuriu între sensibilizarea locomotorie și căutarea compulsivă a medicamentului observată la om (Robinson și Berridge, 1993; Vanderschuren și Kalivas, 2000). Mecanic, administrarea repetată a medicamentelor de abuz sensibilizează circuitele de dopamină mezolimbică crescând neurotransmisia dopaminergică. Psihostimulanții, cum ar fi cocaina sau amfetaminele, care blochează transportorul de dopamină cu membrană plasmatică (DAT), induc o creștere mare de dopamină în spațiul sinaptic din striatum și NAc, activând astfel locomoția (Steketee și Kalivas, 2011). Ca și în dependența de droguri, sensibilizarea circuitului de recompensare / motivație a dopaminei contribuie la comportamente compulsive observate în TOC. Într-adevăr, activarea repetată a receptorilor dopaminei D2 (D2Rs) este suficientă pentru a induce sensibilizarea locomotorie și verificarea comportamentului atât la șobolani, cât și la șoareci (Szechtman și colab., 1998; Szechtman și colab., 1999; Sun și colab., 2019). Administrarea repetată a chinpirolului, un agonist D2R / D3R, este un model acceptat de TOC, deoarece recapitulează valabilitatea feței, printr-un creștere a verificării compulsive și a unui comportament stereotip, valabilitate predictivă, așa cum este observată de o scădere a comportamentelor compulsive după tratamentul cronic cu recaptarea serotoninei inhibitorii (SRI) și validitatea construirii ca structuri cerebrale implicate în acest model sunt împărtășite cu cei din patologie (Stuchlik și colab., 2016; Szechtman și colab., 2017). În rezumat, activarea repetată a transmisiei dopaminei, fie prin pre-sinaptice (eliberare de dopamină), fie post-sinaptice (activarea D2R), conduc la sensibilizarea locomotorie și comportamente compulsive.
Sistemul opioid kappa este unul dintre cele mai preponderent sisteme care controlează transmisia dopaminei în circuitul de recompensare / motivație. Dovada arată că transmiterea kappa-opioidului se opune efectelor dopaminei; activarea acută a receptorilor opioizi kappa (KORs) contracarează activitatea locomotorie indusă de psihostimulanți (Gray și colab., 1999). În schimb, activarea repetată a KOR menține și îmbunătățește căutarea compulsivă și obișnuită a medicamentelor (Koob, 2013). Consumul de droguri de abuz induce o transmisie homeostatică cappa opioidă, contribuind probabil la stările emoționale negative ale disforiei (Koob, 2013) declanșarea consumului compulsiv de droguri (Chavkin și Koob, 2016). De fapt, blocarea KOR a prevenit reîncadrarea nicotinei indusă de stres, dar nu a fost determinată de medicamente (Jackson și colab., 2013), cocaină (Beardsley și colab., 2005) și etanol (Sperling și colab., 2010). În conformitate cu această constatare, blocajul KOR revine modificărilor dopaminergice în striatul dorsolateral al șobolanilor sensibilizați la amfetamină, fără a modifica răspunsul locomotor îmbunătățit la medicament (Azocar și colab., 2019). Astfel, sistemul KOR pare să îmbunătățească armatura negativă crescând valoarea medicamentului. În TOC, întărirea negativă este declanșată de obsesii, care întăresc o anumită compulsie pentru a evita acea obsesie. Deși nu a fost testat direct, consolidarea negativă ar putea juca un rol în sensibilizarea chinpirolului. Într-adevăr, D2R sunt implicați în generarea de întărire negativă. De exemplu, evitarea locului la o zonă pereche de retragere morfină nu a fost dezvoltată la șoarecii care nu au izoformă lungă de D2R (Smith și colab., 2002) și tratamentul repetat cu quinpirol în perioada de abstinență reinstituie căutarea cu heroină și heroină într-o paradigmă de auto-administrare, efect legat de locomoția sensibilizată la chinpirol (De Vries și colab., 2002), sugerând mecanisme partajate între sensibilizarea indusă de psiostimulant și chinpirol. Mai mult, introducerea cuștii de acasă, dar nu o cușcă inedită, pe arena de câmp deschis, reduce sensibilizarea locomotorie și comportamentul de verificare compulsivă (Szechtman și colab., 2001), indicând faptul că semne de siguranță / familiare ar putea concura cu indicii de mediu negative care favorizează sensibilizarea. În mod similar cu sensibilizarea indusă de psiostimulant, activarea repetată a KOR facilitează sensibilizarea locomotorie (Escobar și colab., 2017) și comportament de verificare compulsivă (Perreault și colab., 2007) indusă prin administrarea repetată de chinpirol. Rămâne de elucidat dacă acest efect potențator este o consecință a consolidării negative îmbunătățite.
Analiza minuțioasă efectuată recent arată că efectul sistemului kappa-opioid asupra transmisiei dopaminergice este complex: depinde de calea dopamină implicată (Margolis și colab., 2006; Margolis și colab., 2008) și cu privire la momentul dintre activarea receptorului KOR și activarea receptorului dopaminei (Chartoff și colab., 2016). În concordanță cu această complexitate, utilizarea terapeutică potențială a liganzilor KOR a fost discutată pe scară largă. S-a propus ca agonistul KOR să fie util din punct de vedere clinic în faza de consum de droguri, atenuând hiperdopaminergia indusă de medicament (Shippenberg și colab., 2007). Pe de altă parte, un antagonist KOR poate fi util în tratarea sindromului de retragere indus de o creștere a expresiei de dinorfină după consumul repetat de medicamente (Wee și Koob, 2010). În consecință, sa propus ca agonistul parțial al KOR (Béguin și colab., 2012) ar putea fi o opțiune terapeutică pentru a trata atât consumul compulsiv de droguri, cât și simptomele de retragere la persoanele dependente (Chartoff și colab., 2016; Callaghan și colab., 2018). În această revizuire, analizăm modularea dependentă de timp / context a corelațiilor dopaminergice ale sensibilizării și compulsivității comportamentale.
Întâlnirea anatomică și funcțională între sistemele opioide Kappa și dopaminergice în regiunile Striatale și ale creierului mijlociu
Regiuni Striatale
KOR-urile sunt receptori cuplați de proteine Gi / o foarte exprimați în sistemul dopaminei cerebrale (Mansour și colab., 1996). Acești receptori aparțin familiei receptorilor opioizi, compuși din mu (MOR), delta (DOR) și kappa (KORs). Agoniștii endogeni pentru acești receptori sunt endorfine, encefalină și, respectiv, dinorfină. În striatum, dinorfină este sintetizată de receptorii dopaminici D1 (D1R) care conțin neuroni de dimensiuni medii (MSN) care au axoni recurenti care activează KOR-uri din aceleași nuclee (Mansour și colab., 1995). Imaginile la microscopie electronică a NAc de șobolan arată că KOR-urile se găsesc cu precădere în structurile presinaptice care conțin DAT, în timp ce o proporție minoră de KOR localizează pe dendrite în apunerea la DAT (Svingos și colab., 2001; Kivell și colab., 2014). Studii imunofluorescente care caracterizează preparate presinaptic-sinaptosomale de la NAc arată că KOR și D2R coexistă în mod preferențial în sinaptozomi care conțin enzima de sinteză dopamină, tirosină hidroxilază (TH) (Escobar și colab., 2017). Mai mult decât atât, KOR-urile sunt abundente în corpurile celulare ale NAc și striatum, și colocalizează cu D2Rs într-o subpopulare celulară (Escobar și colab., 2017). Cu idei genetice și moleculare, s-a sugerat că un procent de 20% din KOR-ul obligatoriu în striatum este observat în terminalele DA (Van't Veer și colab., 2013). În plus, Tejeda și colab. (2017) au arătat că MSN-urile D1R și D2R exprimă KOR cu o preferință mai mare pentru MSN-uri care conțin D1R (Tejeda și colab., 2017). Aceste date anatomice indică faptul că KOR sunt prezente și post sinaptic, reglând neurotransmisia dopaminei în circuitul de recompensare / motivație.
Mai multe abordări experimentale arată că activarea KORs inhibă eliberarea dopamineie. Activarea acută a KORs printr-o injecție sistemică sau perfuzia locală de agoniști scade nivelul extracelular de dopamină în NAc (Di Chiara și Imperato, 1988; Spanagel și colab., 1992; Fuentealba și colab., 2006) și striat dorsal (Gehrke și colab., 2008). Susținerea unei acțiuni tonice de inhibare a KOR asupra neurotransmisiei dopaminei, perfuzia directă a antagonistului KOR de lungă și selectivă nor-binaltorfina (nor-BNI) (Broadbear și colab., 1994) crește nivelul bazal de dopamină în NAc (Spanagel și colab., 1992) și eliberarea de dopamină în striatul dorsal (Azocar și colab., 2019). Dovada finală a inhibării tonice KOR a dopaminei a fost arătată la șoarecii knock-out KOR, care au prezentat creșterea nivelului extracelular de dopamină în striatum și NAc (Chefer și colab., 2005). Mecanismele responsabile de inhibarea KOR a eliberării de dopamină nu sunt complet elucidate. Cu toate acestea, este bine cunoscut faptul că activarea KOR duce la creșterea K + și la scăderea conductanțelor Ca2 +, inducând astfel hiperpolarizarea celulară și blocarea eliberării neurotransmițătorului vezicular (Bruchas și Chavkin, 2010; Margolis și Karkhanis, 2019).
În plus,, in vitro și in vivo datele funcționale sugerează că KOR-urile modifică nivelurile extracelulare de dopamină prin modularea activității DAT. De exemplu, activarea KOR în celulele EM4 care co-exprimă KOR și DAT, duce la o absorbție crescută de dopamină măsurată prin voltammetrie (Kivell și colab., 2014). Un ex vivo Analiza folosind și voltammetrie în țesuturile dezagregate, a arătat că o injecție sistemică de agonist KOR U-69593 a crescut absorbția de dopamină în NAc (Thompson și colab., 2000). Un articol similar similar arată că nor-BNI blochează creșterea absorbției de dopamină în striatul ventral și dorsal, indusă de o injecție sistemică acută de MP1104, un agonist al receptorului opioid Kappa / Delta mixt (Atigari și colab., 2019). Cu toate acestea, efectul activării KOR asupra absorbției de dopamină nu a fost încă complet elucidat. Administrarea sistemică a nalmefenului agonist parțial KOR a scăzut în mod dependent doza de administrare a dopaminei striatale, cuantificată prin voltammetrie ciclică cu scanare rapidă (FSCV) (Rose și colab., 2016). Folosind o microdializă fără flux net la șobolani masculi adulți, blocarea KOR a fost însoțită de o creștere a fracției de extracție (Ed), care este o măsură indirectă a absorbției de dopamină (Chefer și colab., 2006; Azocar și colab., 2019), sugerând că activarea tonică a KOR exercită un control inhibitor asupra activității DAT (absorbția dopaminei). Aceste rezultate evidențiază rolul complex al activității KOR endogene asupra absorbției de dopamină pentru a controla nivelurile extracelulare de dopamină. Abordări mai mari de rezoluție temporală, precum FSCV, nu au reușit să arate un efect al KOR asupra absorbției de dopamină (Ebner și colab., 2010; Ehrich și colab., 2015; Hoffman și colab., 2016), sugerând că KOR care îmbunătățește activitatea DAT în regiunile striatale are nevoie de o perioadă de incubație., îmbunătățirea mediată de KOR a activității DAT ar putea fi explicată printr-o creștere a numărului de DAT pe membranele celulare induse de activarea KOR, așa cum este raportat în sinaptozomii striatali și celulă linii (Kivell și colab., 2014).
Regiunile Midbrain
Testele autoradiografice efectuate la nivelul creierului mijlociu la șobolan arată o legătură semnificativă pentru KOR pe axa rostrocaudală a SN și VTA (Speciale și colab., 1993). Pe de altă parte, datele de microscopie electronică arată că terminalele care conțin dinorfină sinapsează direct pe dendritele TH pozitive în SN și VTA (Sesack și Pickel, 1992), sugerând că KORs se localizează în compartimentele somatodendritice ale neuronilor dopaminici. MSN-urile care conțin D1R sunt unul dintre aporturile din dinorfină pentru neuronii dopaminei cerebrale. Interesant este faptul că blocarea KOR-urilor nu modifică efectul inhibitor al D1R-MSN-urilor la neuronii VTA dopamină, ceea ce indică faptul că această inhibare este mediată de GABA (Edwards și colab., 2017). KOR-urile modulează răspunsurile somatodendritice ale neuronilor de dopamină. Studiile electrofiziologice arată că activarea KOR în VTA hiperpolarizează și scade rata de ardere a spontaneului neuronilor dopaminici (Margolis și colab., 2003). În consecință, infuzia de agoniști KOR scade efluxul de dopamină somatodendritică (Smith și colab., 1992; Dalman și O'Malley, 1999). Cu toate acestea, acest efect inhibitor al KOR asupra neuronilor dopaminici pare să fie dependent de circuit. Infuzia de agoniști kappa-opioizi în VTA scade eliberarea de dopamină în cortexul prefrontal medial (mPFC) (Margolis și colab., 2006) dar nu în NAc (Devine și colab., 1993; Margolis și colab., 2006). În plus, Margolis și colab. (2006) a descoperit că KOR-urile inhibă neuronii dopamina VTA care se proiectează către mPFC și amigdala bazolaterală, dar nu și pe cei care se proiectează către NAc. În același an, Ford și colab. (2006) a arătat că aplicarea în baie a agoniștilor KOR în felii de VTA de șoarece a indus un curent exterior mai ridicat în neuronii dopaminici care se proiectează către NAc în comparație cu cei care se proiectează în amigdala bazolaterală, indicând că KOR-urile exercită o inhibare mai mare a neuronilor dopaminici care se proiectează la NAc decât la amigdala. Mai mult, activarea KOR scade amplitudinea excitatorilor (Margolis și colab., 2005) și inhibitor (Ford și colab., 2007) curenți postsinaptici în neuronii dopaminei cerebrale mijlocii. Diferențele dintre specii și proiecțiile eferente complexe ale VTA la mPFC și NAc (Van Bockstaele și Pickel, 1995; Carr și Sesack, 2000) face dificil să se stabilească dacă KOR-urile inhibă selectiv unele dintre populațiile neuronale de dopamină din VTA. Cu toate acestea, datele rezumate aici indică faptul că KOR-urile sunt în soma și terminalele neuronilor dopaminei, precum și în input-urile care le reglează, astfel poziționate în mod deosebit pentru a controla activitatea sinaptică a neuronilor dopaminei din creierul mijlociu.
Rolul KOR care controlează neurotransmisia dopaminei în sensibilizarea indusă de psihostimulanți și în comportamentele compulsive
Dependența de droguri este un proces care implică căutarea inițială de droguri impulsivă asociată cu efectele lor de consolidare pozitivă. Pe de altă parte, compulsivitatea este o trăsătură de personalitate observabilă la dependenții de droguri. Mai multe neuroadaptări pe căi dopaminergice au fost propuse pentru a ține cont de căutarea și consumul compulsiv de droguri în urma expunerii repetate la droguri de abuz (Everitt și Robbins, 2005; Koob și Volkow, 2016). Una dintre ipotezele propuse care determină consumul compulsiv de droguri este sensibilizarea efectelor sale de consolidare negativă (Koob, 2013). Controlul inhibitor al sistemului kappa opioid la eliberarea de dopamină ar putea contribui la proprietățile de consolidare negativă a medicamentelor de abuz. Cu toate acestea, consecințele activării KOR asupra neurotransmisiei dopaminei și a căutării medicamentelor compulsive par a fi complexe și aparent contradictorii. Într-adevăr, eliberarea de dopamină indusă de amfetamină și cocaină este atenuată prin administrarea concomitentă de agoniști KOR (Heidbreder și Shippenberg, 1994; Maisonneuve și colab., 1994; Thompson și colab., 2000) și chiar scade auto-administrarea cocainei (Negus și colab., 1997). Mai mult decât atât, KOR-urile exercită un feedback inhibitor asupra eliberării dopaminei căii mezolimbice ca răspuns la activarea susținută a D1R post-sinaptic, așa cum apare cu expunerea repetată la psiostimulatori (Cole și colab., 1995; Nestler, 2001). Paradoxal, activarea KOR poate facilita, de asemenea, eliberarea de dopamină pe calea de recompensare / motivație (Fuentealba și colab., 2006; Fuentealba și colab., 2007) și consumul de psiostimulanți (Wee și colab., 2009). Fuentealba și colab. (2007) a arătat că după patru zile administrând U69593, un agonist KOR, a crescut eliberarea de dopamină indusă de amfetamină în NAc. Recent, s-a demonstrat că blocarea KOR-urilor inversează schimbările în eliberarea și absorbția dopaminei în striatul dorsal care are loc în timpul sensibilizării locomotorii indusă de amfetamină (Azocar și colab., 2019). În total, aceste date sugerează că activarea KOR-urilor ar putea contribui și la proprietățile de consolidare pozitivă a drogurilor de abuz (Chartoff și colab., 2016).
În plus, activarea KORs pare să contribuie și la căutarea compulsivă a medicamentelor; Blocajul KORs reduce cocaina (Wee și colab., 2009), heroină (Schlosburg și colab., 2013) și metamfetamină (Whitfield și colab., 2015) aportul la șobolani cu acces nelimitat la medicament (Wee și colab., 2009). Acest efect este evidențiat și în căutarea medicamentelor induse de stres. De exemplu, șoarecii knock-out KOR nu au arătat preferința locului la cocaină după stresul înotat forțat (McLaughlin și colab., 2006a). Blocarea KOR-urilor atenuează preferința locului nicotinei indusă de expunerea forțată la înot (Smith și colab., 2012). Interesant este că blocarea KOR atenuează căutarea cu cocaină și nicotină indusă de stres, dar nu a afectat căutarea indusă de o provocare de droguri (Beardsley și colab., 2005; Jackson și colab., 2013). Acest efect KOR facilitator indus de stres pare a fi mediat de circuitul de recompensare / motivație (Shippenberg și colab., 2007; Wee și Koob, 2010). Într-un studiu elegant realizat de Dr. Kauer și grupul ei, s-a demonstrat că blocarea KOR-urilor în VTA, anterior sau după un stres acut, inhibă reinstalarea căutării cocainei, un efect asociat cu salvarea pe termen lung a potențarea sinapselor inhibitoare în neuronii dopaminici (Graziane și colab., 2013; Polter și colab., 2014).
Facilitarea aportului de psiostimulatori exercitat de KOR pare să depindă de un interval de timp în ceea ce privește expunerea la medicamente. Administrarea agonistului KOR U50488 cu 1 h înainte de expunerea la cocaină potențează atât preferința locului la cocaină, cât și eliberarea relativă de dopamină evocată de cocaină în NAc, în timp ce efectele opuse sunt observate atunci când sunt administrate cu 15 min înainte (McLaughlin și colab., 2006a; Ehrich și colab., 2014). Utilizarea auto-stimulării intracraniene Chartoff și colab. (2016) a observat că agonistul KOR Salvinorină A, are un efect aversiv inițial și un efect satisfăcător întârziat, însoțit de o scădere și, respectiv, de o creștere a eliberarii de dopamină stimulată în NAc. Toate aceste date indică un efect dependent de timp al activării KOR asupra proprietăților plină de satisfacție ale cocainei și indică activarea KOR mediată de stres ca un actor cheie pentru dezvoltarea căutării compulsive a medicamentelor.
Sensibilizare locomotorie indusă de quinpirol și comportament compulsiv
Faptele că sistemul dopaminelor este implicat în generarea de sensibilizare și compulsivitate este consolidat de comportamentul observat la rozătoarele tratate cu agonistul D2R, chinpirol. Pe scurt, D2R-urile sunt receptori cuplat Gi exprimat pe scară largă în circuitul de recompensare / motivație; ele sunt exprimate somatodendritic și pe terminalele axonice ale neuronilor dopaminici (Sesack și colab., 1994), iar activarea acesteia scade nivelurile extracelulare de dopamină (Imperato și Di Chiara, 1988). În striat, D2R-urile sunt, de asemenea, localizate post sinaptic pe neuronii spinoși medieni (Sesack și colab., 1994) și activarea sa inhibă calea indirectă permițând activitatea locomotorie.
Dr. Henry Szechtman a început să studieze efectele chinpirolului asupra comportamentului șobolanilor care se încheie în deceniul din 1980. Constatările lor inițiale au arătat că administrarea acută de quinpirol are efect dependent de doză asupra activității locomotorii. La doze mici (0.03 mg / kg) scade activitatea locomotorie, în timp ce la doze mai mari (> 0.5 mg / kg), crește. (Eilam și Szechtman, 1989). Aceste efecte sunt asociate cu activarea D2R-uri presinaptice cu afinitate înaltă și respectiv D2R-uri postsinaptice cu afinitate scăzută (Usiello și colab., 2000). În mod neașteptat, administrarea repetată (la fiecare două zile) de chinpirol induce o creștere treptată și susținută a locomoției, asemănătoare cu sensibilizarea locomotorie indusă de psihostimulanți (Szechtman și colab., 1993; Szechtman și colab., 1994). Sa dovedit că efectul de sensibilizare locomotor depinde de D2R, deoarece șoarecii deficienți pentru acest receptor nu dezvoltă sensibilizare locomotorie la chinpirol (Escobar și colab., 2015).
La începutul anilor '90, Szechtman și Eilam au raportat că, împreună cu sensibilizarea locomotorie, șobolanii au dezvoltat un comportament stereotip, care este întărit cu fiecare administrare de quinpirol (Eilam și Szechtman, 1989; Szechtman și colab., 1993). Astăzi, administrarea repetată de quinpirol este un model validat pentru TOC (Szechtman și colab., 1999; Szechtman și colab., 2001; Eilam și Szechtman, 2005; Stuchlik și colab., 2016; Szechtman și colab., 2017), pe baza observației că comportamentul șobolanilor devine din ce în ce mai structurat și inflexibil, reminiscența comportamentului ritual caracteristic comportamentului de verificare compulsivă (Szechtman și colab., 1998; Szechtman și colab., 2017). Studii recente arată că chinpirolul repetat induce, de asemenea, comportamente compulsive la șoareci, cum ar fi verificarea compulsivă (Sun și colab., 2019), inflexibilitate comportamentală și mestecare compulsivă (Asaoka și colab., 2019), aceasta din urmă a revenit prin blocarea D2Rs în striatum, susținând în continuare că este necesară activarea repetată a D2Rs pentru a induce comportamente compulsive. Împreună, datele indică un rol crucial al D2R în căile de dopamină ale creierului mediu pentru a induce sensibilizarea și compulsivitatea locomotorie. Administrarea repetată a chinpirolului determină un comportament stereotipizat indus de cocaină (Thompson și colab., 2010) și efectele locomotorii ale amfetaminei (Cope și colab., 2010), consolidând ideea că activarea D2Rs stă la baza sensibilizării induse de psiostimulant și sugerează un mecanism comun între sensibilizarea indusă de chinpirol și psiostimulatori. Interesant este că efectul de sensibilizare al activării repetate a D2Rs pare să fie mai puternic decât cel indus de psihostimulanți, deoarece fiecare șobolan tratat cu quinpirol dezvoltă sensibilizare locomotorie (Escobar și colab., 2015), în timp ce aproximativ șaizeci la sută dintre șobolani sensibilizează la amfetamină (Escobar și colab., 2012; Casanova și colab., 2013).
Sensibilizarea comportamentală indusă de activarea repetată a D2Rs este însoțită de adaptări în circuitul de recompensare / motivație. Șobolanii sensibilizați cu chinpirol au un ton dopaminergic mai scăzut în NAc, observat ca scăzut bazal (Koeltzow și colab., 2003) și eliberarea de dopamină tonică și fazică stimulată (Escobar și colab., 2015), ceea ce indică scăderea capacității de eliberare a dopaminei a circuitului de dopamină. Nivelurile de dopamină sinaptică din NAc sunt controlate de activitatea neuronilor DAT și a dopaminei (Goto și Grace, 2008), care in vivo constă în tragere tonică și explozie (Wilson și colab., 1977; Grace și Bunney, 1980). Rapoartele anterioare arată că șobolanii sensibilizați cu quinpirol afișează un număr mai mic de neuroni dopaminici în arsuri tonice și explozii în VTA (Sesia și colab., 2013). Împreună, aceste date indică faptul că scăderea eliberării de dopamină observată după sensibilizarea la quinpirol este rezultatul scăderii activității generale a neuronilor dopaminici. Comportamentul compulsiv și activitatea locomotorie sensibilizată indusă de tratamentul repetat cu chinpirol ar putea fi o consecință a sensibilizării D2Rs, datorită scăderii tonusului dopaminergic în NAc. Într-adevăr, șobolanii sensibilizați la quinpirol arată o creștere a legării dopaminei D2R (Culver și colab., 2008) și o creștere a stării de afinitate a acestor receptori (Perreault și colab., 2007), susținând această ipoteză.
Interacțiunile KOR-Dopaminei în comportamente compulsive induse de quinpirol
Studiile inițiale privind rolul KOR în comportamentele compulsive induse de D2R au venit, de asemenea, din laboratorul lui Szechtman. Acest grup a examinat administrarea concomitentă a agonistului KOR U69593 cu chinpirol pe activitatea locomotorie. Mai precis, autorii au administrat injecții subcutanate șobolanilor cu un amestec U69593 și chinpirol, până la finalizarea a 8-10 injecții. Spre deosebire de efectul hipolocomotor al U69593 în monoterapie, s-a observat hiperlocomotie la administrarea concomitentă cu doze mici (presinaptice) și mari (postsinaptice) de chinpirol. U69593 a modificat efectul hipolocomotor al unei doze presinaptice de chinpirol în hiperlocomocie și a îmbunătățit efectul hiperlocomotor al unei doze postsinaptice de chinpirol (Perreault și colab., 2006). Co-activarea KOR-urilor a accelerat, de asemenea, inducerea sensibilizării locomotorii și a potențat efectul activării D2Rs, deoarece locomoția maximă obținută prin tratamentul dublu duplică efectul locomotor indus de quinpirol singur (Perreault și colab., 2006; Escobar și colab., 2017). Co-activarea KOR-urilor accelerează, de asemenea, dobândirea unui comportament de verificare compulsivă (Perreault și colab., 2007). Aceste efecte potențante ale KOR asupra comportamentelor induse de chinpirole necesită activarea repetată a KORs. De fapt, injecția acută a agonistului KOR U69593 nu a modificat în continuare activitatea locomotorie la șobolani sensibilizați cu chinpirol (Escobar și colab., 2017). Mecanismul de sensibilizare indus de D2R potentat de KOR nu este cunoscut. O posibilitate este că sistemul opioid cappa endogen însuși mediază sensibilizarea dependentă de D2R. Cu toate acestea, această posibilitate a fost eliminată arătând că pre-administrarea norBNI nu a modificat sensibilizarea locomotorie la chinpirol, ceea ce sugerează că dinorfină nu este eliberată în activarea D2R în aval (Escobar și colab., 2017). Aceste date nu exclud că dinorfină ar putea avea un rol în sensibilizarea comportamentelor compulsive, de exemplu, stresul produce eliberarea de dinorfină și activarea KOR-urilor care facilitează comportamentele compulsive (McLaughlin și colab., 2003; McLaughlin și colab., 2006a; McLaughlin și colab., 2006b).
Crucea între D2R și KOR este complexă și pare să depindă dacă activarea ambilor receptori este coincidentă sau separată temporal. Datele anatomice indică faptul că intersecția dintre D2R și KOR poate apărea presinaptic în axoni și soma neuronilor dopaminei, precum și postinaptic în MSN-urile striatului. Deși nu exclude un rol pentru KOR-urile localizate pe axonii altor sisteme neurochimice, datele anatomice indică puternic un rol direct al KOR-urilor care reglementează D2R-urile. Acut sau repetat, activarea KOR scade funcția inhibitoare D2Rs asupra neuronilor dopaminici. Studiile de electrofiziologie au arătat că activarea acută a KOR în neuronii dopaminici ai VTA și SN inhibă curentul inhibitorii postsinaptice mediate de D2R, efect mediat de mecanisme pre și post-sinynaptice, deoarece KOR scade eliberarea de dopamină și dinorfină blochează efectul inhibitor al dopaminei aplicate în baie (Ford și colab., 2007). Studiile neurochimice au arătat că activarea repetată a KORs blochează inhibarea eliberarii de dopamină indusă de D2R în NAc (Fuentealba și colab., 2006). Mai mult decât atât, activarea acută a D2R-urilor și a KOR-urilor scade inhibarea eliberării de dopamină în NAc în comparație cu efectul fiecărui receptor singur (Escobar și colab., 2017). Astfel, KOR-urile presinaptice nu acționează în mod aditiv sau în sinergie cu D2R-urile presinaptice, invers, KOR-urile inhibă efectul inhibitor D2R sau ocluzează. Acest mecanism ar putea explica efectul de activare locomotor al unei doze acute de agoniști KOR concomitent cu o doză mică de chinpirol (Perreault și colab., 2006).
Un studiu recent arată că activarea KOR în VTA mediază comportamentul compulsiv măsurat ca inhibiție comportamentală și îngroparea marmurei (Abraham și colab., 2017), consolidând ideea că activarea KORs este într-adevăr un declanșator al compulsivității. Date publicate de Margolis și colab. (2006; 2008) indică faptul că interacțiunea KOR și D2R ar trebui să aibă loc pe neuronii dopaminei care vizează mPFC (Margolis și colab., 2006; Margolis și colab., 2008). În ciuda, Ford și colab. (2006; 2007) a constatat că inhibarea KORs a IPSC mediată de D2R are loc pe neuronii dopaminei care vizează NAc (Ford și colab., 2006; Ford și colab., 2007). Împreună, aceste date arată că interacțiunea KOR cu D2R la compartimentul somatodendritic al neuronilor dopaminici ar putea apărea ca urmare a unei creșteri în același neuron dopaminic. Dacă acest lucru se întâmplă în proiecțiile mesolimbice sau mezocorticale este încă controversat.
În mod remarcabil, KOR a fost găsită în MSN-urile NAc (Escobar și colab., 2017; Tejeda și colab., 2017), indicând astfel că potențarea comportamentului compulsiv indus de D2R poate apărea și prin acțiuni directe asupra celulelor țintă ale neuronilor dopaminici. În acest sens, este de menționat faptul că administrarea repetată de U69593 crește cantitatea de D2Rs în stare de afinitate ridicată (Perreault și colab., 2007). Datele neurochimice indică faptul că nivelul scăzut al dopaminei extracelulare este asociat cu sensibilizarea D2Rs. Co-activarea KOR-urilor nu scade în continuare nivelurile extracelulare de dopamină din NAc deja a scăzut prin activarea repetată a D2Rs (Escobar și colab., 2017), care exclude un rol pentru KOR-uri presinaptice care accelerează sau potențează sensibilizarea D2Rs în NAc prin acest mecanism. Prin urmare, KOR-urile declanșează mecanisme moleculare lente care sensibilizează în continuare efectele neurochimice și comportamentale ale D2Rs, ceea ce sugerează că îmbunătățirea sensibilizării locomoției s-ar putea datora mai degrabă unui efect postsinaptic adaptiv și nu unui efect presinaptic. În acest sens, activarea repetată a KOR-urilor poate declanșa inhibarea D2R a căii striatice indirecte de comutare a echilibrului D1R / D2R la D1R care induce compulsivitatea (Figura 1).
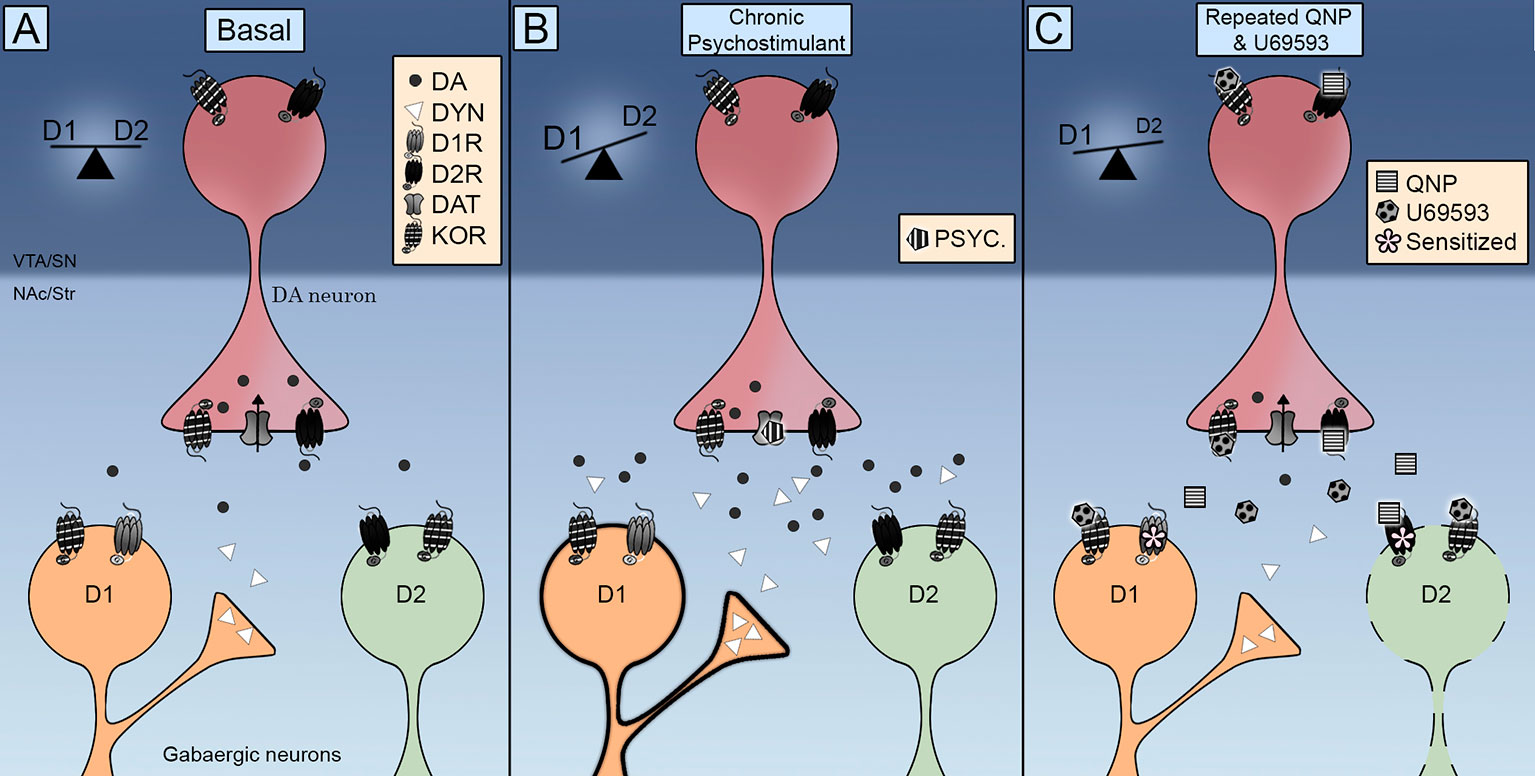 Figura 1 Schema integrativă a controlului receptorilor opioizi Kappa (KOR) pe căile striatale directe (D1R) și indirecte (D2R). (A) KOR sunt localizate pre-sinaptic pe terminalele dopaminei și post sinaptic în neuronii de talie medie (MSN). Activarea sa controlează nivelurile extracelulare de dopamină și localizarea acesteia promovează interacțiunea cu transportorii de dopamină (DAT) și receptorii D2 de dopamină. (B) Expunerea repetată la un psiostimulant este însoțită de o creștere a nivelurilor extracelulare de dopamină și dinorfină. Activarea receptorilor D1 și D2 trece echilibrul pe calea directă D1R care promovează sensibilizarea locomotorie. (C) Administrarea concomitentă de quinpirol și U69593 este însoțită de o scădere a nivelului extracelular de dopamină. Activarea concomitentă a receptorilor KOR și D2 debilitează calea indirectă D2 care induce un comportament compulsiv.
Figura 1 Schema integrativă a controlului receptorilor opioizi Kappa (KOR) pe căile striatale directe (D1R) și indirecte (D2R). (A) KOR sunt localizate pre-sinaptic pe terminalele dopaminei și post sinaptic în neuronii de talie medie (MSN). Activarea sa controlează nivelurile extracelulare de dopamină și localizarea acesteia promovează interacțiunea cu transportorii de dopamină (DAT) și receptorii D2 de dopamină. (B) Expunerea repetată la un psiostimulant este însoțită de o creștere a nivelurilor extracelulare de dopamină și dinorfină. Activarea receptorilor D1 și D2 trece echilibrul pe calea directă D1R care promovează sensibilizarea locomotorie. (C) Administrarea concomitentă de quinpirol și U69593 este însoțită de o scădere a nivelului extracelular de dopamină. Activarea concomitentă a receptorilor KOR și D2 debilitează calea indirectă D2 care induce un comportament compulsiv.
Diferențele de sex ale interacțiunilor KOR-Dopamina în comportamentele compulsive
Studiile clinice au arătat diferențe de sex în comportamentul compulsiv, inclusiv căutarea compulsivă a medicamentelor. Un debut mai precoce al simptomelor TOC este observat la bărbați în comparație cu femeile (Mathis și colab., 2011), femeile manifestând mai multă prevalență a contaminării și simptomelor de curățare (Labad și colab., 2008). În ceea ce privește diferențele de sex în dependența de droguri, dovezi clinice indică faptul că, în timp ce consumul de droguri este mai răspândit la bărbați, femeile prezintă o evoluție mai rapidă decât bărbații în căutarea compulsivă a drogurilor (Hernandez-Avila și colab., 2004; Fattore și Melis, 2016).
În ultima perioadă, dovezile pre-clinice au evidențiat puternic bazele neurobiologice care stau la baza diferențelor de sex în consumul de droguri observate în studiile clinice (Becker și Chartoff, 2019). Primele observații folosind microdialysis cu flux net-net au arătat că concentrația extracelulară de dopamină în striatul dorsal variază în timpul ciclului estru cu niveluri mai mari în proestrus și estru în comparație cu diestrus. Mai mult, în timp ce ovariectomia scade concentrația extracelulară de dopamină striată la șobolani de sex feminin, castrarea șobolanilor masculi nu modifică concentrația extracelulară a dopaminei (Xiao și Becker, 1994), sugerând un rol important al hormonilor ovarului asupra activității dopaminei. În plus, hormonii feminini reglează răspunsul la psiostimulatori. Din timp in vitro experimentele au arătat că estradiolul plus progesteronul reface eliberarea de dopamină indusă de amfetamină din țesutul striatal obținut de șobolani ovariectomizați (Becker și Ramirez, 1981). Mai recent, studii de voltammetrie ciclică cu scanare rapidă au arătat că femeile prezintă o eliberare și o absorbție mai mare de dopamină stimulate electric în comparație cu bărbații (Walker și colab., 2000). Aceste diferențe de sex în neurotransmisia dopaminei pot reprezenta căutarea mai mare a cocainei și a amfetaminei observate la femei. (Roberts și colab., 1989; Cox și colab., 2013).
Reglarea KOR asupra nivelurilor extracelulare de dopamină arată, de asemenea, diferențe de sex (Chartoff și Mavrikaki, 2015). Folosind auto-stimularea intracraniană și voltammetrie ciclică, Conway și colab. (2019) a arătat că sensibilitatea mai mică la efectul anhedonic acut al unui agonist KOR observat la șobolani de sex feminin, comparativ cu șobolanii masculi, este însoțită de o inhibare atenuată a eliberarii de dopamină stimulată în NAConway și colab., 2019). S-a sugerat că estradiolul contribuie la inhibarea descurajată a eliberarii de dopamină observată la șobolanii de sex feminin după activarea KOR (Abraham și colab., 2018). În timp ce intersecția dintre KOR și semnalizarea dopaminei a fost studiată la bărbați (Tejeda și Bonci, 2019), lipsesc cercetările privind această interacțiune și impactul acesteia în procesul de dependență la femei (Chartoff și Mavrikaki, 2015). La șobolanii de sex feminin, administrarea acută a agonistului KOR U69593 a atenuat hiperlocomotia indusă de cocaină atât la șobolani de control, cât și la șobolani ovariectomizați. Interesant, U69593 administrarea repetată a atenuat hiperlocomotia indusă de cocaină într-o manieră dependentă de estradiol (Puig-Ramos și colab., 2008). Aceste date sugerează că estradiolul primează acțiuni de KOR la șobolani de sex feminin, efect care ar putea fi legat de diferențele de sex în răspunsul la stres (Puig-Ramos și colab., 2008). Dacă la șobolani de sex feminin, activarea repetată a KORs facilitează eliberarea de dopamină striată, așa cum s-a observat la bărbați, este o întrebare fără răspuns.
Deși o facilitare a eliberării dopaminei induse de psiostimulant este observată la femei, comparativ cu șobolanii, diferențele de sex în mecanismele dopaminei care stau la baza sensibilizării locomotorii a amfetaminei nu au fost complet elucidate (Becker, 1999). Expunerea repetată la amfetamină induce o activitate locomotorie mai mare atât la adolescenți (Mathews și McCormick, 2007) și șobolani de sex adult (Milesi-Hallé și colab., 2007), cu șobolani de sex feminin care prezintă o sensibilizare locomotorie mai robustă după expunerea repetată la amfetamină. Activarea neonatală a receptorului D2 a potențat sensibilizarea comportamentală indusă de amfetamină numai la șobolani (Brown și colab., 2011). Așa cum am menționat anterior, s-a observat la șobolani masculi expunerea repetată la agonistul D2 induce sensibilizare locomotorie și comportament asemănător compulsivDvorkin și colab., 2006). Mai mult, coactivarea KOR potențează sensibilizarea locomotorie indusă de expunerea repetată la chinpirol, facilitând controlul inhibitor al receptorilor D2 la eliberarea DA în NAc (Escobar și colab., 2017). Diferențe de sex, cum ar fi sensibilitatea scăzută observată la efectul inhibitor al KOR asupra eliberării dopaminei la femei (Conway și colab., 2019) poate reprezenta o contribuție diferențiată a KOR la căutarea compulsivă a medicamentelor.
Concluzii
Cum modulează KORs semnalizarea dopaminei pentru a elabora comportamente motivate și când are ca rezultat un comportament compulsiv sensibilizat? Datele anatomice arată că KOR-urile sunt poziționate în mod deosebit pentru a controla activitatea sinaptică a neuronilor dopaminei cerebrale. Datele funcționale indică faptul că KOR controlează funcționarea DAT și D2R, precum și rata de tragere a neuronilor dopaminici. Primele dovezi care arată că activarea acută a KOR scade eliberarea de dopamină indusă de drogurile de abuz a fost completată cu date care indică faptul că activarea repetată a KOR facilitează eliberarea de dopamină și căutarea compulsivă a medicamentelor. Căile de semnalizare Dopamine echilibrează căile de ieșire directe și indirecte din zonele striatale (Figura 1A). Fie stimulare cronică cu psiostimulatori care crește eliberarea de dopamină activând atât D1R cât și D2R (Figura 1B) sau chinpirol care activează numai D2R (Figura 1C) are ca rezultat sensibilizarea locomotorie și comportamente compulsive printr-o cale indirectă D2R debilitată, schimbând astfel echilibrul pe calea directă D1R. Transmiterea KOR este îmbunătățită în timpul aportului psihostimulant cronic prin creșterea dinorfină în neuronii D1 striatali (Figura 1B). O transmisie îmbunătățită de KOR este emulată în modelul farmacologic al TOC prin administrarea U69593. Această activare KOR concomitentă debilitează în continuare calea indirectă D2 (Figura 1C). Ar trebui efectuate cercetări viitoare pentru a elucida pe deplin consecințele activării KOR asupra activității DAT, pentru a înțelege rolul sistemului KOR endogeu în compulsivitatea indusă de chinpirol și pentru a determina contribuția sistemului KOR la diferențele de sex observate în comportamentele compulsive.
Contribuțiile autorului
AE, MA și JF au contribuit la concepția manuscrisului. AE și JF au scris primul proiect al manuscrisului cu aport de la MA. MA și JC au contribuit la revizuirea critică și la editarea manuscrisului. Toți autorii au aprobat-o spre publicare.
Finanțare
Lucrările autorilor citați în această recenzie au fost susținute de numerele de finanțare FONDECYT: 1110352 și 1150200 către MA; 1141088 la JF; DIPOG acordă 391340281 JF; FONDECYT coleg postdoctoral 3170497 la JC și 3190843 AE.
Conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Editorul de manipulare organizează în prezent o temă de cercetare cu unul dintre autorii JF și confirmă absența oricărei alte colaborări.
Referinte
Abraham, AD, Fontaine, HM, Song, AJ, Andrews, MM, Baird, MA, Kieffer, BL, și colab. (2017). Activarea receptorului opioid Kappa în neuronii dopaminei perturbă inhibarea comportamentului. Neuropsychopharmacology 43 (2), 362–372. doi: 10.1038 / npp.2017.133
Abraham, AD, Schattauer, SS, Reichard, KL, Cohen, JH, Fontaine, HM, Song, AJ, și colab. (2018). Reglarea estrogenului GRK2 inactivează semnalizarea receptorului opioid kappa care mediază analgezia, dar nu aversiunea. J. Neurosci. 38 (37), 8031–8043. doi: 10.1523 / JNEUROSCI.0653-18.2018
Asaoka, N., Nishitani, N., Kinoshita, H., Nagai, Y., Hatakama, H., Nagayasu, K., et al. (2019). Un antagonist al receptorului adenozinei A2A îmbunătățește mai multe simptome ale psihozei induse de chinpirole repetate. eNeuro 6 (1), 1–16. ENEURO.0366-18.2019. doi: 10.1523 / ENEURO.0366-18.2019
Atigari, DV, Uprety, R., Pasternak, GW, Majumdar, S., Kivell, BM (2019). MP1104, un agonist mixt al receptorilor opioizi kappa-delta are proprietăți anti-cocaină cu efecte secundare reduse la șobolani. Neuropharmacology 150, 217-228. doi: 10.1016 / j.neuropharm.2019.02.010
Azocar, VH, Sepúlveda, G., Ruiz, C., Aguilera, C., Andrés, ME, Fuentealba, JA (2019). Blocarea receptorului kappa-opioid inversează schimbările dinamicii dopaminei dorsolaterale în timpul sensibilizării amfetaminei. J. Neurochem. 148, 348–358. doi: 10.1111 / jnc.14612
Béguin, C., Potuzak, J., Xu, W., Liu-Chen, LY, Streicher, JM, Groer, CE, și colab. (2012). Proprietăți de semnalizare diferențială la receptorul opioid kappa al 12-epi-salvinorinei A și analogii săi. Bioorg. Med. Chim. Lett. 15; 22 (2), 1023-1026. doi: 10.1016 / j.bmcl.2011.11.128
Beardsley, PM, Howard, JL, Shelton, KL, Carroll, FI (2005). Efectele diferențiale ale noului antagonist al receptorilor opioizi kappa, JDTic, asupra reintegrării căutării cu cocaină indusă de stresorii de la picior față de primele de cocaină și efectele sale antidepresive la șobolani. Psychopharmacol. (Beri) 183, 118–126. doi: 10.1007/s00213-005-0167-4
Becker, JB, Chartoff, E. (2019). Diferențele de sex în mecanismele neuronale mediază recompensa și dependență. Neuropsychopharmacology 44, 166–183. doi: 10.1038/s41386-018-0125-6
Becker, JB, Ramirez, VD (1981). Diferențele de sex în amfetamina au stimulat eliberarea de catecolamine din țesutul striatal de șobolan in vitro. Brain Res. 204, 361–372. doi: 10.1016/0006-8993(81)90595-3
Becker, JB (1999). Diferențele de gen în funcția dopaminergică în striatum și nucleus accumbens. Pharmacol. Biochem. Behav. 64, 803–812. doi: 10.1016/S0091-3057(99)00168-9
Berridge, KC, Robinson, TE (2016). Teoria gustării, dorinței și stimularea sensibilizării dependenței. A.m. Psychol. 71, 670–679. doi: 10.1037 / amp0000059
Berridge, KC, Venier, IL, Robinson, TE (1989). Analiza reactivității la gust a afagiei induse de 6-hidroxidopamina: implicații pentru ipoteza excitației și anhedoniei funcției dopaminei. Behav. Neurosci. 103, 36-45. doi: 10.1037 / 0735-7044.103.1.36
Broadbear, JH, Negus, SS, Butelman, ER, de Costa, BR, Woods, JH (1994). Efecte diferențiale ale nor-binaltorfina administrată sistemic (nor-BNI) asupra agoniștilor kappa-opioizi în testul de încetinire a șoarecilor. Psychopharmacology 115, 311-319. doi: 10.1007 / BF02245071
Brown, RW, Perna, MK, Noel, DM, Whittemore, JD, Lehmann, J., Smith, ML (2011). Sensibilizarea locomotorie a amfetaminei și preferința condiționată a locului la șobolanii adolescenți de sex masculin și feminin tratate neonat cu chinpirol. Behav. Pharmacol. 22, 374–378. doi: 10.1097/FBP.0b013e328348737b
Bruchas, MR, Chavkin, C. (2010). Cascade kinazice și semnalizare direcționată ligand la receptorul opioid kappa. Psychopharmacol. (Beri) 210, 137–147. doi: 10.1007/s00213-010-1806-y
Callaghan, CK, Rouine, J., O'Mara, SM (2018). Roluri potențiale pentru receptorii opioizi în motivație și tulburare depresivă majoră. Prog. Brain Res. 239, 89–119. doi: 10.1016 / bs.pbr.2018.07.009
Carr, DB, Sesack, SR (2000). Neuronii care conțin GABA în zona tegmentală ventrală a șobolanului se proiectează la cortexul prefrontal. Synapse 38 (2), 114–123. doi: 10.1002/1098-2396(200011)38:2<114::AID-SYN2>3.0.CO;2-R
Casanova, JP, Velis, GP, Fuentealba, JA (2013). Sensibilizarea locomotorie a amfetaminei este însoțită de o eliberare ridicată de dopamină stimulată de K + ridicată în cortexul prefrontal medial al șobolanului. Behav. Brain Res. 237, 313-317. doi: 10.1016 / j.bbr.2012.09.052
Chartoff, EH, Mavrikaki, M. (2015). Diferențele de sex în funcția receptorilor opioizi kappa și impactul lor potențial asupra dependenței. Față. Neurosci. 9, 466. doi: 10.3389 / fnins.2015.00466
Chartoff, EH, Ebner, SR, Sparrow, A., Potter, D., Baker, PM, Ragozzino, ME, și colab. (2016). Momentul relativ între activarea receptorului opioid kappa și cocaină determină impactul asupra recompensei și eliberării dopaminei. Neuropsychopharmacology 41, 989-1002. doi: 10.1038 / npp.2015.226
Chavkin, C., Koob, GF (2016). Dinorfină, disforie și dependență: stresul dependenței. Neuropsychopharmacology 41, 373-374. doi: 10.1038 / npp.2015.258
Chefer, VI, Czyzyk, T., Bolan, EA, Moron, J., Pintar, JE, Shippenberg, T. S. (2005). Sistemele endogene de receptori kappa-opioizi reglează dinamica dopaminei mezoacumbale și vulnerabilitatea la cocaină. J. Neurosci. 25, 5029-5037. doi: 10.1523 / JNEUROSCI.0854-05.2005
Chefer, VI, Zapata, A., Shippenberg, TS, Bungay, PM (2006). Microdialysia cantitativă fără flux net permite detectarea creșterilor și scăderilor absorbției de dopamină în nucleul accumbens de șoarece. J. Neurosci. metode 155, 187-193. doi: 10.1016 / j.jneumeth.2005.12.018
Cole, RL, Konradi, C., Douglass, J., Hyman, SE (1995). Adaptare neuronală la amfetamină și dopamină: mecanisme moleculare de reglare a genei prodynorfin în striatum de șobolan. Neuron 14, 813–823. doi: 10.1016/0896-6273(95)90225-2
Conway, SM, Puttick, D., Russell, S., Potter, D., Roitman, MF, Chartoff, E. H. (2019). Femeile sunt mai puțin sensibile decât bărbații la efectele motivaționale și de suprimare a dopaminei ale activării receptorului opioid kappa. Neuropharmacology 146, 231-241. doi: 10.1016 / j.neuropharm.2018.12.002
Cope, ZA, Huggins, KN, Sheppard, AB, Noel, DM, Roane, DS, Brown, RW (2010). Tratamentul cu quinpirole neonatale îmbunătățește activarea locomotorie și eliberarea de dopamină în nucleul accumbens, ca răspuns la tratamentul cu amfetamină la vârsta adultă. Synapse 64, 289-300. doi: 10.1002 / syn.20729
Cox, BM, Young, AB, See, RE, Reichel, CM (2013). Diferențele de sex în căutarea metamfetaminei la șobolan: impactul oxitocinei. Psychoneuroendocrinology 38, 2343-2353. doi: 10.1016 / j.psyneuen.2013.05.005
Culver, KE, Szechtman, H., Levant, B. (2008). Alterarea receptorilor dopaminei asemănătoare cu D2 la șobolani cu sensibilizare comportamentală la chinpirol: efectele tratamentului pre-tratat cu Ro 41-1049. EURO. J. Pharmacol. 592, 67-72. doi: 10.1016 / j.ejphar.2008.06.101
Dalman, FC, O'Malley, KL (1999). kappa-Toleranță și dependență la opioide în culturile de neuroni dopaminergici ai creierului mediu. J. Neurosci. 19, 5750–5757. doi: 10.1523/JNEUROSCI.19-14-05750.1999
De Vries, TJ, Schoffelmeer, AN, Binnekade, R., Raasø, H., Vanderschuren, LJ (2002). Recidiva la comportamentul de căutare a cocainei și a heroinei mediat de receptorii D2 dopaminei depinde de timp și este asociat cu sensibilizarea comportamentală. Neuropsychopharmacology 26, 18–26. doi: 10.1016/S0893-133X(01)00293-7
Devine, DP, Leone, P., Pocock, D., Wise, RA (1993). Implicarea diferențială a receptorilor tegmentali murali ventrali, delta și kappa opioizi în modularea eliberarii bazolice a dopaminei mesolimbice: studii de vivo de microdializă. J. Pharmacol. Exp. Ther. 266, 1236-1246.
Di Chiara, G., Imperato, A. (1988). Efectele opuse ale agoniștilor opiacei mu și kappa asupra eliberării dopaminei în nucleul accumbens și în caudatul dorsal al șobolanilor în mișcare liberă. J. Pharmacol. Exp. Ther. 244, 1067-1080. doi: 10.1073 / pnas.85.14.5274
Dvorkin, A., Perreault, ML, Szechtman, H. (2006). Dezvoltarea și organizarea temporală a verificării compulsive induse de injecțiile repetate de quinamină agonistă dopamină într-un model animal de tulburare obsesiv-compulsivă. Behav. Brain Res. 169, 303-311. doi: 10.1016 / j.bbr.2006.01.024
Ebner, SR, Roitman, MF, Potter, DN, Rachlin, AB, Chartoff, EH (2010). Efectele de tip depresiv ale agonistei A agonistului receptorului kappa opioid sunt asociate cu scăderea eliberării fazice de dopamină în nucleul accumbens. Psychopharmacol. (Beri) 210, 241–252. doi: 10.1007/s00213-010-1836-5
Edwards, NJ, Tejeda, HA, Pignatelli, M., Zhang, S., McDevitt, RA, Wu, J., și colab. (2017). Specificitatea circuitului în arhitectura inhibitoare a VTA reglementează comportamentul indus de cocaină. Nat. Neurosci. 20 (3), 438–448. doi: 10.1038 / n.4482
Ehrich, JM, Phillips, PEM, Chavkin, C. (2014). Activarea receptorului opioid Kappa potențează creșterea indusă de cocaină în eliberarea de dopamină evocată înregistrată in vivo în nucleul accumbens de șoarece. Neuropsychopharmacology 39, 3036-3048. doi: 10.1038 / npp.2014.157
Ehrich, JM, Messinger, DI, Knakal, CR, Kuhar, JR, Schattauer, SS, Bruchas, MR, și colab. (2015). Aversiunea indusă de receptor opioid Kappa necesită activarea MAPK p38 în neuronele dopaminei VTA. J. Neurosci. 35, 12917-12931. doi: 10.1523 / JNEUROSCI.2444-15.2015
Eilam, D., Szechtman, H. (1989). Efect bifazic al chinpirolului agonist D-2 asupra locomoției și mișcărilor. EURO. J. Pharmacol. 161, 151–157. doi: 10.1016/0014-2999(89)90837-6
Eilam, D., Szechtman, H. (2005). Comportamentul indus de psihostimulant ca model animal al tulburării obsesiv-compulsive: o abordare etologică a formei de ritualuri compulsive. CNS Spectr. 10, 191–202. doi: 10.1017 / S109285290001004X
Escobar, AP, Cornejo, FA, Andrés, ME, Fuentealba, JA (2012). Tratamentul repetat cu agonistul U69593 al receptorului kappa opioid inversează eliberarea crescută de dopamină indusă de K + în nucleul accumbens, dar nu și expresia sensibilizării locomotorii la șobolani sensibilizați la amfetamină. Neurochem. Int. 60 (4), 344–349. doi: 10.1016 / j.neuint.2012.01.014
Escobar, AP, Cornejo, FA, Olivares-Costa, M., González, M., Fuentealba, JA, Gysling, K., et al. (2015). Diminuarea neurotransmisiei de dopamină și glutamat în nucleul obișnuit al șobolanilor sensibilizați la chinpirole indică funcția de autoreceptor D2 inhibitor. J. Neurochem. 134, 1081–1090. doi: 10.1111 / jnc.13209
Escobar, AP, González, MP, Meza, RC, Noches, V., Henny, P., Gysling, K., și colab. (2017). Mecanisme de potențare a receptorilor opioizi kappa a funcției receptorului dopaminei D2 în sensibilizarea locomotorie indusă de chinpirol la șobolani. Int. J. Neuropsychopharmacol. 20, 660–669. doi: 10.1093 / ijnp / pyx042
Everitt, BJ, Robbins, TW (2005). Sisteme neuronale de armare pentru dependența de droguri: de la acțiuni la obiceiuri până la compulsie. Nat. Neurosci. 8, 1481-1489. doi: 10.1038 / nn1579
Fattore, L., Melis, M. (2016). Diferențele de sex în comportamentele impulsive și compulsive: un accent pe dependența de droguri. Addict. Biol. 21 (5), 1043-1051. doi: 10.1111 / adb.12381
Ferrario, CR, Gorny, G., Crombag, HS, Li, Y., Kolb, B., Robinson, TE (2005). Plasticitatea neuronală și comportamentală asociată cu trecerea de la consumul de cocaină controlat la escaladat. Biol. Psi. 58 (9), 751-9.
Figee, M., Pattij, T., Willuhn, I., Luigjes, J., van den Brink, W., Goudriaan, A., și colab. (2016). Compulsivitatea în tulburări obsesive - compulsive și dependențe. EURO. Neuropsychopharmacol. 26, 856-868. doi: 10.1016 / j.euroneuro.2015.12.003
Ford, CP, Mark, GP, Williams, JT (2006). Proprietățile și inhibarea opioidă a neuronilor dopaminei mezolimbice variază în funcție de locația vizată. J. Neurosci. 26, 2788-2797. doi: 10.1523 / JNEUROSCI.4331-05.2006
Ford, CP, Beckstead, MJ, Williams, JT (2007). Inhibarea kappa opioidă a curenților postinaptici inhibitori ai dopaminei somatodendritice. J. Neurophysiol. 97, 883-891. doi: 10.1152 / jn.00963.2006
Fuentealba, JA, Gysling, K., Magendzo, K., Andrés, ME (2006). Administrarea repetată a agonistului selectiv al receptorului kappa-opioid U-69593 crește nivelul stimulat al dopaminei extracelulare stimulat în nucleul accumbens. J. Neurosci. Res. 84, 450-459. doi: 10.1002 / jnr.20890
Fuentealba, JA, Gysling, K., Andrés, ME (2007). Răspuns locomotor crescut la amfetamină indus de administrarea repetată a agonistului selectiv al receptorului kappa-opioid U-69593. Synapse 61, 771-777. doi: 10.1002 / syn.20424
Gehrke, BJ, Chefer, VI, Shippenberg, TS (2008). Efectele administrării acute și repetate de salvinorină A asupra funcției dopaminei în striatul dorsal de șobolan. Psychopharmacol. (Beri) 197, 509–517. doi: 10.1007/s00213-007-1067-6
Giuliano, C., Belin, D., Everitt, BJ (2019). Apariția compulsivă a alcoolului rezultă din eșecul de dezactivării controlului striatal dorsolateral asupra comportamentului. J. Neurosci. 39 (9), 1744–1754. doi: 10.1523 / JNEUROSCI.2615-18.2018
Goto, Y., Grace, AA (2008). Prelucrarea informațiilor limbice și corticale din nucleul accumbens. Tendințe Neurosci. 31, 552-558. doi: 10.1016 / j.tins.2008.08.002
Grace, AA, Bunney, BS (1980). Neuronii dopaminei nigrale: înregistrare și identificare intracelulară cu injecție de L-dopa și histofluorescență. Ştiinţă 210, 654-656. doi: 10.1126 / science.7433992
Grey, AM, Rawls, SM, Shippenberg, TS, McGinty, JF (1999). Agonistul op-opioid, U-69593, scade comportamentele acute evocate de amfetamină și nivelurile de dialamină dependente de calciu de dopamină și glutamat în striatul ventral. J. Neurochem. 73, 1066-1074. doi: 10.1046 / j.1471-4159.1999.0731066.x
Graziane, NM, Polter, AM, Briand, LA, Pierce, RC, Kauer, JA (2013). Receptorii Kappa opioizi reglează căutarea cocainei indusă de stres și plasticitatea sinaptică. Neuron 77, 942-954. doi: 10.1016 / j.neuron.2012.12.034
Heidbreder, CA, Shippenberg, TS (1994). U-69593 previne sensibilizarea la cocaină normalizând dopamina accumbens bazală. Neuroreport 5, 1797–1800. doi: 10.1097/00001756-199409080-00028
Hernandez-Avila, CA 1., Rounsaville, BJ, Kranzler, HR (2004). Femeile dependente de opioide, canabis și alcool arată o evoluție mai rapidă către tratamentul abuzului de substanțe. Alcoolul de droguri depinde. 74 (3), 265–272. doi: 10.1016 / j.drugalcdep.2004.02.001
Hoffman, AF, Spivak, CE, Lupica, CR (2016). Eliberarea îmbunătățită a dopaminei de către inhibitorii transportului de dopamină descrisă de un model de difuzie restrânsă și de voltammetrie ciclică cu scanare rapidă. ACS Chem. Neurosci. 7, 700–709. doi: 10.1021 / acschemneuro.5b00277
Holden, C. (2001). Dependențele „comportamentale”: există? Ştiinţă 294, 980-982. doi: 10.1126 / science.294.5544.980
Imperato, A., Di Chiara, G. (1988). Efectele agoniștilor și antagoniștilor receptorilor D-1 și D-2 aplicate local studiate cu dializă cerebrală. EURO. J. Pharmacol. 156, 385–393. doi: 10.1016/0014-2999(88)90284-1
Jackson, KJ, McLaughlin, JP, Carroll, FI, Damaj, MI (2013). Efectele antagonistului receptorului opioid kappa, norbinaltorfina, asupra stresului și reintegrării induse de medicamente a preferinței locului condiționat de nicotină la șoareci. Psychopharmacol. (Beri) 226, 763–768. doi: 10.1007/s00213-012-2716-y
Kivell, B., Uzelac, Z., Sundaramurthy, S., Rajamanickam, J., Ewald, A., Chefer, V., și colab. (2014). Salvinorina A reglează funcția de transportare a dopaminei printr-un receptor kappa opioid și un mecanism dependent de ERK1 / 2. Neuropharmacology 86, 228-240. doi: 10.1016 / j.neuropharm.2014.07.016
Koeltzow, TE, Austin, JD, Vezina, P. (2003). Sensibilizarea comportamentală la chinpirol nu este asociată cu creșterea excesului de dopamină din nucleul accumbens. Neuropharmacology 44, 102–110. doi: 10.1016/S0028-3908(02)00328-3
Koob, GF, Volkow, ND (2016). Neurobiologia dependenței: o analiză de neurocircuite. Lancet Psihiatrie 3, 760–773. doi: 10.1016/S2215-0366(16)00104-8
Koob, GF (2013). Dependența este un deficit de recompensă și tulburarea surplusului de stres. Față. Psihiatrie 4, 72. doi: 10.3389 / fpsyt.2013.00072
Labad, J. 1., Menchon, JM, Alonso, P., Segalas, C., Jimenez, S., Jaurrieta, N., și colab. (2008). Diferențele de gen în dimensiunile obsesiv-compulsive ale simptomelor. Deprimați anxietatea. 25 (10), 832–838. doi: 10.1002 / da.20332
Maisonneuve, IM, Archer, S., Glick, SD (1994). U50,488, un agonist al receptorilor opioizi kappa, atenuează creșterile induse de cocaină ale dopaminei extracelulare din nucleul accumbens al șobolanilor. Neurosci. Lett. 181, 57–60. doi: 10.1016/0304-3940(94)90559-2
Mansour, A., Fox, CA, Akil, H., Watson, SJ (1995). Expresia ARNm-receptor opioid în SNC de șobolan: implicații anatomice și funcționale. Tendințe Neurosci. 18 (1), 22–29. doi: 10.1016/0166-2236(95)93946-U
Mansour, A., Burke, S., Pavlic, RJ, Akil, H., Watson, SJ (1996). Localizarea imunohistochimică a receptorului kappa 1 clonat în SNC de șobolan și hipofiză. Neuroştiinţe 71, 671–690. doi: 10.1016/0306-4522(95)00464-5
Margolis, EB, Karkhanis, AN (2019). Contribuțiile dopaminergice celulare și ale circuitului la aversiunea mediată de receptorul opioid kappa. Neurochem. Int. 129, 104504. doi: 10.1016 / j.neuint.2019.104504
Margolis, EB, Hjelmstad, GO, Bonci, A., Fields, HL (2003). Agoniștii Kappa-opioizi inhibă direct neuronii dopaminergici din creierul mijlociu. J. Neurosci. 23, 9981–9986. doi: 10.1523/JNEUROSCI.23-31-09981.2003
Margolis, EB, Hjelmstad, GO, Bonci, A., Fields, HL (2005). Atât agoniștii Kappa cât și Mu Opioid inhibă intrarea glutamatergică în neuronele zonei Tegmentale Ventrale. J. Neurophysiol. 93, 3086-3093. doi: 10.1152 / jn.00855.2004
Margolis, EB, Lock, H., Chefer, VI, Shippenberg, TS, Hjelmstad, GO, Fields, HL (2006). Opioizii Kappa controlează selectiv neuronii dopaminergici care se proiectează în cortexul prefrontal. Proc. Natl. Acad. Sci. Statele Unite ale Americii 103, 2938-2942. doi: 10.1073 / pnas.0511159103
Margolis, EB, Mitchell, JM, Ishikawa, J., Hjelmstad, GO, Fields, HL (2008). Neuronii dopaminei cerebrale: ținta de proiecție determină durata potențială de acțiune și inhibarea receptorului dopaminei D (2). J. Neurosci. 28, 8908-8913. doi: 10.1523 / JNEUROSCI.1526-08.2008
Mathews, IZ, McCormick, CM (2007). Șobolanii de sex feminin și de sex masculin în adolescența târzie diferă de adulți în activitatea locomotorie indusă de amfetamină, dar nu se preferă în loc condiționat pentru amfetamină. Behav. Pharmacol. 18, 641–650. doi: 10.1097/FBP.0b013e3282effbf5
Mathis, MA 1., Pd, A., Funaro, G., RC, T., Moraes, I., AR, T., și colab. (2011). Diferențele de gen în tulburarea obsesiv-compulsivă: o revizuire a literaturii Braz. J. Psihiatrie 33 (4), 390-399. doi: 10.1590 / S1516-44462011000400014
McLaughlin, JP, Marton-Popovici, M., Chavkin, C. (2003). Antagonismul receptorilor opioizi Kappa și perturbarea genei prodinorfinei blochează răspunsurile comportamentale induse de stres. J. Neurosci. 23 (13), 5674–5683. doi: 10.1523 / JNEUROSCI.23-13-05674.2003
McLaughlin, JP, Land, BB, Li, S., Pintar, JE, Chavkin, C. (2006a). Activarea prealabilă a receptorilor opioizi kappa prin U50,488 imită stresul de înot forțat repetat pentru a potența condiționarea preferinței locului cocainei. Neuropsychopharmacology 31, 787-794. doi: 10.1038 / sj.npp.1300860
McLaughlin, JP, Li, S., Valdez, J., Chavkin, TA, Chavkin, C. (2006b). Răspunsurile comportamentale induse de înfrângerea socială sunt mediate de sistemul opioid cappa endogen. Neuropsychopharmacology 31 (6), 1241–1248. doi: 10.1038 / sj.npp.1300872
Milesi-Hallé, A., McMillan, DE, Laurenzana, EM, Byrnes-Blake, KA, Owens, SM (2007). Diferențele de sex la (+) - amfetamină și (+) - răspuns comportamental indus de metamfetamină la șobolani Sprague-Dawley, bărbați și femei. Pharmacol. Biochem. Behav. 86, 140-149. doi: 10.1016 / j.pbb.2006.12.018
Negus, SS, Mello, NK, Portoghese, PS, Lin, CE (1997). Efectele opioidelor kappa asupra auto-administrării cocainei de către maimuțele rhesus. J. Pharmacol. Exp. Ther. 282, 44-55.
Nestler, EJ (2001). Baza moleculară a plasticității pe termen lung care stă la baza dependenței. Nat. Rev. Neurosci. 2, 119-128. doi: 10.1038 / 35053570
Perreault, ML, Graham, D., Bisnaire, L., Simms, J., Hayton, S., Szechtman, H. (2006). Agonistul Kappa-opioid U69593 potențează sensibilizarea locomotorie la chinpirolul agonist D2 / D3: mecanisme pre și post sinaptice. Neuropsychopharmacology 31, 1967-1981. doi: 10.1038 / sj.npp.1300938
Perreault, ML, Seeman, P., Szechtman, H. (2007). Stimularea receptorului kappa-opioid accelerează patogeneza verificării compulsive în modelul de sensibilizare la chinpirole al tulburării obsesiv-compulsive (TOC). Behav. Neurosci. 121, 976-991. doi: 10.1037 / 0735-7044.121.5.976
Pierce, RC, Kalivas, PW (1997). Un model de circuit al expresiei sensibilizării comportamentale la psiostimulatoare asemănătoare amfetaminei. Brain Res. Brain Res. Rev. 25, 192–216. doi: 10.1016/S0165-0173(97)00021-0
Polter, AM, Bishop, RA, Briand, LA, Graziane, NM, Pierce, RC, Kauer, JA (2014). Blocul post-stres al receptorilor opioizi Kappa salvează potențializarea pe termen lung a sinapselor inhibitoare și previne reîncadrarea căutării de cocaină. Biol. Psihiatrie 76, 785-793. doi: 10.1016 / j.biopsych.2014.04.019
Puig-Ramos, A., Santiago, GS, Segarra, AC (2008). U-69593, un agonist al receptorilor opioizi kappa, scade sensibilizarea comportamentală indusă de cocaină la șobolani. Behav. Neurosci. 122, 151-160. doi: 10.1037 / 0735-7044.122.1.151
Robbins, TW, Gillan, CM, Smith, DG, de Wit, S., Ersche, KD (2012). Endofenotipuri neurocognitive ale impulsivității și compulsivității: spre psihiatrie dimensională. Tendințe Cognit. Sci. 16, 81-91. doi: 10.1016 / j.tics.2011.11.009
Roberts, DCS, Bennett, SAL, Vickers, GJ (1989). Ciclul estros afectează auto-administrarea de cocaină într-o schemă de raport progresivă la șobolani. Psychopharmacol. (Beri) 98, 408-411. doi: 10.1007 / BF00451696
Robinson, TE, Berridge, KC (1993). Baza neurală a poftei de droguri: o teorie de stimulare a sensibilizării dependenței. Brain Res. Brain Res. Rev. 18, 247–291. doi: 10.1016/0165-0173(93)90013-P
Robinson, TE, Berridge, KC (2001). Incentivant-sensibilizare și dependență. Dependenta 96, 103-114. doi: 10.1046 / j.1360-0443.2001.9611038.x
Rose, JH, Karkhanis, AN, Steiniger-Brach, B., Jones, SR (2016). Efecte distincte ale nalmefenului asupra ratelor de absorbție a dopaminei și a activității receptorului kappa opioid în nucleul accumbens după expunerea cronică intermitentă la etanol. Int. J. Mol. Sci. 17, 1216. doi: 10.3390 / ijms17081216
Schlosburg, JE, Whitfield, TW, Jr., Park, PE, Crawford, EF, George, O., Vendruscolo, LF, și colab. (2013). Antagonismul pe termen lung al receptorilor κ opioizi previne escaladarea și creșterea motivației pentru aportul de heroină. J. Neurosci. 33 (49), 19384–19392. doi: 10.1523 / JNEUROSCI.1979-13.2013
Sesack, SR, Pickel, VM (1992). Localizarea dublă ultrastructurală a impereorectivității enkephalinei și tirozinei hidroxilazei în zona tegmentală ventrală a șobolanului: multiple substraturi pentru interacțiuni opiace-dopamină. J. Neurosci. 12, 1335–a1350. doi: 10.1523/JNEUROSCI.12-04-01335.1992
Sesack, SR, Aoki, C., Pickel, VM (1994). Localizarea ultrastructurală a imunoreactivității asemănătoare receptorului D2 în neuronii dopaminei cerebrale mijlocii și țintele lor striatale. J. Neurosci. 14, 88–106. doi: 10.1523/JNEUROSCI.14-01-00088.1994
Sesia, T., Bizup, B., Grace, AA (2013). Evaluarea modelelor animale ale tulburării obsesiv-compulsive: corelarea cu activitatea neuronilor dopaminici fazici. Int. J. Neuropsychopharmacol. 16, 1295–1307. doi: 10.1017 / S146114571200154X
Shippenberg, TS, Zapata, A., Chefer, VI (2007). Dinorfina și fiziopatologia dependenței de droguri. Farmacol. Ther. 116, 306–321. doi: 10.1016 / j.pharmthera.2007.06.011
Smith, JA, Loughlin, SE, Leslie, FM (1992). inhibiția kappa-opioidă a eliberarii dopaminei [3H] din culturile de celule disociate mezenfalice ventrale de șobolan. Mol. Pharmacol. 42.
Smith, JW, Fetsko, LA, Xu, R., Wang, Y. (2002). Șoarecii knockout receptori Dopamina D2L prezintă deficiențe în proprietățile de întărire pozitive și negative ale morfinei și în învățarea evitării. Neuroştiinţe 113 (4), 755–765. doi: 10.1016/S0306-4522(02)00257-9
Smith, JS, Schindler, AG, Martinelli, E., Gustin, RM, Bruchas, MR, Chavkin, C. (2012). Activarea indusă de stres a sistemului de receptori dinorfină / κ-opioid în amigdala potențează preferința condiționată de nicotină. J. Neurosci. 32, 1488-1495. doi: 10.1523 / JNEUROSCI.2980-11.2012
Spanagel, R., Herz, A., Shippenberg, TS (1992). Sistemele opioide endogene active opuse, tonic, modulează calea dopaminergică mezolimbică. Proc. Natl. Acad. Sci. 89, 2046-2050. doi: 10.1073 / pnas.89.6.2046
Speciale, SG, Manaye, KF, Sadeq, M., Germană, DC (1993). Receptori opioizi în regiunile dopaminergice ale creierului mijlociu la șobolan. II. Autoradiografie a receptorului Kappa și delta. J. Transmul neural. Secțiunea gen. 91, 53-66. doi: 10.1007 / BF01244918
Sperling, RE, Gomes, SM, Sypek, EI, Carey, AN, McLaughlin, JP (2010). Medierea kapo-opioidă endogenă a potențării provocate de stres a preferinței locului etanol condiționate de etanol și auto-administrare. Psychopharmacol. (Beri) 210 (2), 199–209. doi: 10.1007/s00213-010-1844-5
Steketee, JD, Kalivas, PW (2011). Dorința de droguri: sensibilizare comportamentală și recidivă la comportamentul căutător de droguri sibley DR, ed. Pharmacol. Rev. 63, 348-365. doi: 10.1124 / pr.109.001933
Stuchlik, A., Radostová, D., Hatalova, H., Vales, K., Nekovarova, T., Koprivova, J., și colab. (2016). Validitatea modelului de șobolan de sensibilizare la quinpirol al TOC: legătura dintre dovezile obținute din studii clinice la animale și animale. Față. Behav. Neurosci. 10, 209. doi: 10.3389 / fnbeh.2016.00209
Sun, T., Song, Z., Tian, Y., Tian, W., Zhu, C., Ji, G., și colab. (2019). Introducerea amigdala bazolaterală a cortexului prefrontal medial controlează un comportament de verificare obsesiv-compulsiv asemănător tulburării. Proc. Natl. Acad. Sci. Statele Unite ale Americii 116, 3799-3804. doi: 10.1073 / pnas.1814292116
Svingos, AL, Chavkin, C., Colago, EEO, Pickel, VM (2001). Coexpresia majoră a receptorilor - opioizi și a transportorului de dopamină în profilele axonale ale nucleului accumbens. Synapse 42, 185-192. doi: 10.1002 / syn.10005
Szechtman, H., Talangbayan, H., Eilam, D. (1993). Componente de mediu și comportamentale ale sensibilizării induse de chinpirolul agonist dopaminic. Behav. Pharmacol. 4, 405–410. doi: 10.1097/00008877-199308000-00014
Szechtman, H., Talangbayan, H., Canaran, G., Dai, H., Eilam, D. (1994). Dinamica sensibilizării comportamentale indusă de agonistul dopaminei quinpirol și a unui mecanism de control central al energiei propus. Psychopharmacol. (Beri) 115, 95-104. doi: 10.1007 / BF02244757
Szechtman, H., Sulis, W., Eilam, D. (1998). Quinpirolul induce un comportament de verificare compulsivă la șobolani: un model animal potențial de tulburare obsesiv-compulsivă (TOC). Behav. Neurosci. 112, 1475-1485. doi: 10.1037 / 0735-7044.112.6.1475
Szechtman, H., Culver, K., Eilam, D. (1999). Rolul sistemelor de dopamină în tulburarea obsesiv-compulsivă (TOC): implicații ale unui model nou de animal indus de psihotimulatoare. Pol. J. Farmacol. 51, 55-61.
Szechtman, H., Eckert, MJ, Tse, WS, Boersma, JT, Bonura, C a, JZ, M., și colab. (2001). Comportamentul verificării obligatorii a șobolanilor sensibilizați cu quinpirol ca model animal al tulburării obsesiv-compulsive (TOC): formă și control. BMC Neurosci. 2, 4. doi: 10.1186/1471-2202-2-4
Szechtman, H., Ahmari, SE, Beninger, RJ, Eilam, D., Harvey, BH, Edemann-Callesen, H., și colab. (2017). Tulburare obsesiv-compulsivă: perspective asupra modelelor animale. Neurosci. Biobehav. Rev. 76, 254-279. doi: 10.1016 / j.neubiorev.2016.04.019
Tejeda, HA, Bonci, A. (2019). Controlul dinorfină / receptor kappa-opioid al dinamicii dopaminei: implicații pentru stări afective negative și tulburări psihiatrice. Brain Res. 1713, 91-101. doi: 10.1016 / j.brainres.2018.09.023
Tejeda, HA, Wu, J., Kornspun, AR, Pignatelli, M., Kashtelyan, V., Krashes, M. J., și colab. (2017). Modularea receptorilor kappa-opioizi specifici căilor și a celulelor a echilibrului de excitație-inhibiție diferențiază diferit porțile d1 și d2 acumbens activitatea neuronului. Neuron 93 (1), 147–163. doi: 10.1016 / j.neuron.2016.12.005
Thompson, AC, Zapata, A., Justice, JB, Vaughan, RA, Sharpe, LG, Shippenberg, TS (2000). Activarea receptorului Kappa-opioid modifică absorbția de dopamină în nucleul accumbens și se opune efectelor cocainei. J. Neurosci. 20, 9333–9340. doi: 10.1523/JNEUROSCI.20-24-09333.2000
Thompson, D., Martini, L., Whistler, JL (2010). Raportul modificat al receptorilor dopaminei D1 și D2 în striatul de șoarece este asociat cu sensibilizarea comportamentală la cocaină. Plus unu 5, e11038. doi: 10.1371 / journal.pone.0011038
Usiello, A., Baik, J.-H., Rougé-Pont, F., Picetti, R., Dierich, A., LeMeur, M., și colab. (2000). Funcții distincte ale celor două izoforme ale receptorilor dopaminei D2. Natură 408, 199-203. doi: 10.1038 / 35041572
Van Bockstaele, EJ, Pickel, VM (1995). Neuronii care conțin GABA în zona tegmentală ventrală proiectează nucleul accumbens în creierul de șobolan. Brain Res. 682 (1-2), 215–221. doi: 10.1016/0006-8993(95)00334-M
Van't Veer, A., Bechtholt, AJ, Onvani, S., Potter, D., Wang, Y., Liu-Chen, LY și colab. (2013). Ablația receptorilor kappa-opioizi din neuronii dopaminei cerebrale are efecte asemănătoare anxiolitice și îmbunătățește plasticitatea indusă de cocaină. Neuropsychopharmacology 38 (8), 1585–1597. doi: 10.1038 / npp.2013.58
Vanderschuren, LJ, Kalivas, PW (2000). Modificări în transmisia dopaminergică și glutamatergică în inducerea și exprimarea sensibilizării comportamentale: o revizuire critică a studiilor preclinice. Psychopharmacol. (Beri) 151 (2-3), 99–120. doi: 10.1007 / s002130000493
Volkow, ND, Wise, RA, Baler, R. (2017). Sistemul de dopamină: implicații pentru dependența de droguri și alimente. Nat. Rev. Neurosci. 18, 741–752. doi: 10.1038 / nr.2017.130
Walker, QD, Rooney, MB, Wightman, RM, Kuhn, CM (2000). Eliberarea și absorbția de dopamină sunt mai mari la femele decât striatul de șobolan masculin, măsurat prin voltammetrie ciclică rapidă. Neuroştiinţe 95, 1061–1070. doi: 10.1016/S0306-4522(99)00500-X
Wee, S., Koob, GF (2010). Rolul sistemului opioid dinorfină κ în efectele de întărire ale drogurilor de abuz. Psychopharmacol. (Beri) 210, 121–135. doi: 10.1007/s00213-010-1825-8
Wee, S., Orio, L., Ghirmai, S., Cashman, JR, Koob, GF (2009). Inhibarea receptorilor opioizi kappa a atenuat creșterea aportului de cocaină la șobolani cu acces extins la cocaină. Psychopharmacol. (Beri) 205, 565–575. doi: 10.1007/s00213-009-1563-y
Whitfield, TW, Jr., Schlosburg, JE, Wee, S., Gould, A., George, O., Grant, Y., și colab. (2015). κ Receptorii opioizi din nucleul accumbens mediază escaladarea aportului de metamfetamină. J. Neurosci. 35 (10), 4296–4305. doi: 10.1523 / JNEUROSCI.1978-13.2015
Williams, MT, Mugno, B., Franklin, M., Faber, S. (2013). Dimensiunile simptomelor în tulburarea obsesiv-compulsivă: fenomenologia și rezultatele tratamentului cu expunerea și prevenirea ritualului. Psihopatologie 46, 365-376. doi: 10.1159 / 000348582
Wilson, CJ, Young, SJ, Groves, PM (1977). Proprietățile statistice ale trenurilor neuronale în substanța nigra: tipurile de celule și interacțiunile lor. Brain Res. 136, 243–260. doi: 10.1016/0006-8993(77)90801-0
Înțelept, RA (2009). Rolul pentru nigrostriatal - nu doar mezocorticolimbic - dopamină în recompensă și dependență. Tendințe Neurosci. 32, 517-524. doi: 10.1016 / j.tins.2009.06.004
Xiao, L., Becker, JB (1994). Determinarea cantitativă a microdialysei a concentrației de dopamină striatală extracelulară la șobolani bărbați și femei: efecte ale ciclului estros și gonadectomie. Neurosci. Lett. 180, 155–158. doi: 10.1016/0304-3940(94)90510-X
Cuvinte cheie: receptor opioid kappa, dopamină, compulsivitate, amfetamină, chinpirol, sensibilizare locomotorie
Referire: Escobar AdP, Casanova JP, Andrés ME și Fuentealba JA (2020) Crosstalk Între sistemele Kappa Opioid și Dopamina în comportamentele compulsive. Față. Pharmacol. 11: 57. doi: 10.3389 / fphar.2020.00057
Primit: 16 octombrie 2019; Acceptat: 22 ianuarie 2020;
Publicat: 18 Februarie 2020.
Editat de:
Gonzalo E. Yevenes, Universitatea din Concepția, Chile
Revizuite de:
Luis Gerardo Aguayo, Universitatea din Concepția, Chile
Hugo Tejeda, Institutul Național pentru Abuzul de Droguri (NIDA), Statele Unite
Cecilia Scorza, Institutul de Investigații Biologice Clemente Estable (IIBCE), Uruguay
Copyright © 2020 Escobar, Casanova, Andrés și Fuentealba. Acesta este un articol cu acces deschis distribuit în condițiile din Creative Commons Attribution License (CC BY). Utilizarea, distribuirea sau reproducerea în alte forumuri este permisă, cu condiția ca autorii și proprietarii drepturilor de autor să fie creditați și să fie citată publicația originală în acest jurnal, în conformitate cu practicile academice acceptate. Nu este permisă utilizarea, distribuirea sau reproducerea care nu respectă acești termeni.
* Corespondență: José Antonio Fuentealba, [e-mail protejat]
†ORCID: José Antonio Fuentealba, orcid.org/0000-0003-0775-0675
