 Martin Zack
Martin Zack- 1Laboratorul de psihofarmacologie cognitiv, Departamentul de Neuroștiințe, Centrul de dependență și sănătate mintală, Toronto, ON, Canada
- 2Programul de Neuroștiințe în Translație, Departamentul de Psihiatrie, Scoala de Medicină, Universitatea din Pennsylvania, Philadelphia, PA, SUA
- 3Secția de biopsihologie, Departamentul de Neuroștiințe, Centrul de dependență și sănătate mintală, Toronto, ON, Canada
Dependența este considerată a fi o boală a creierului cauzată de expunerea cronică la medicamente. Sensibilizarea sistemelor dopaminei cerebrale (DA) mediază în parte acest efect. Jocurile de noroc patologice (PG) sunt considerate a fi dependență comportamentală. Prin urmare, PG poate fi cauzată de expunerea cronică la jocurile de noroc. Identificarea unei sensibilizări induse de jocuri de noroc a sistemelor DA ar susține această posibilitate. Recompensele pentru jocuri de noroc evocă eliberarea DA. Un episod al jocului cu sloturi joacă schimbarea răspunsului DA de la livrarea de recompense la debutul indicatorilor (rotitoare) pentru recompensă, în conformitate cu principiile de învățare diferențială temporală. Astfel, stimulii condiționați (CS) joacă un rol-cheie în răspunsurile DA la jocurile de noroc. La primate, răspunsul DA la un CS este mai puternic atunci când probabilitatea recompensării este 50%. În cadrul acestui program, CS provoacă o speranță de recompensă, dar nu furnizează informații despre faptul dacă va avea loc într-un anumit proces. În timpul jocurilor de noroc, un program 50% ar trebui să genereze o eliberare maximă de DA. Acest lucru se potrivește cu frecvența recompensă (46%) într-o mașină de jocuri comerciale. Eliberarea DA poate contribui la sensibilizarea, în special la amfetamină. Expunerea cronică la un CS care prezice recompensa 50% din timp ar putea imita acest efect. Am testat această ipoteză în trei studii cu șobolani. Animalele au primit expuneri 15 × 45-min la un CS care a prezis recompensa cu o probabilitate de 0, 25, 50, 75 sau 100%. CS era o lumină; recompensa a fost o soluție 10% sucroză. După antrenament, șobolanii au primit un regim de sensibilizare a cinci doze separate (1 mg / kg) de d-amfetamină. În cele din urmă, aceștia au primit o provocare de amfetamină 0.5 sau 1 mg / kg înainte de un test de activitate locomotorie 90-min. În toate cele trei studii, grupul 50% a prezentat o activitate mai mare decât celelalte grupuri ca răspuns la ambele doze de provocare. Dimensiunile efectelor au fost modeste, dar consecvente, reflectate de o asociere semnificativă a grupului × rank (φ = 0.986, p = 0.025). Expunerea cronică la un program asemănător cu jocurile de noroc de stimuli predictivi de recompensă poate promova sensibilizarea amfetaminei la fel ca expunerea la amfetamină în sine.
Introducere
Dependența a fost caracterizată ca o boală a creierului cauzată de expunerea cronică la medicamente de abuz (Leshner, 1997). Se crede că neuroplasticitatea mediază efectele unei astfel de expuneri (Nestler, 2001). Sensibilizarea sistemelor de dopamină cerebrală (DA) este o formă de neuroplasticitate implicată în hiper-reactivitate față de stimulii condiționați (CS) pentru medicamente și de căutare a drogurilor compulsive (Robinson și Berridge, 2001). Sensibilizarea a fost definită în mod operațional prin eliberarea crescută de DA ca răspuns la un CS pentru recompensă și prin răspunsul locomotor crescut la provocarea DA farmacologică (Robinson și Berridge, 1993; Pierce și Kalivas, 1997; Vanderschuren și Kalivas, 2000). Deși sensibilizarea este doar una dintre schimbările cerebrale legate de dependență (cf. Robbins și Everitt, 1999; Koob și Le Moal, 2008), s-au sugerat schimbări în eliberarea dopaminergiei presinaptice pentru a reprezenta neuroadaptările comune implicate în depistarea dependenței de droguri (de exemplu, recădere), prin faptul că medicamente care induc sensibilizarea locomotorie la provocarea cu opiacee (de exemplu, morfină) sau stimularea (de exemplu, amfetamina) , determină, de asemenea, reintroducerea răspunsurilor operatorului stingător pentru heroină sau cocaina de autoadministrare - un model animal de recădere (Vanderschuren și colab., 1999). Dovezile că sensibilizarea stimulentelor (valoarea crescută a recompensei de droguri) este cea mai pronunțată după expunerea inițială la medicamentele dependente sugerează în continuare că sensibilizarea poate fi implicată și în stadiile incipiente ale dependenței (Vanderschuren și Pierce, 2010).
Jocurile de noroc patologice (PG) au fost descrise ca dependență comportamentală și reclasificate recent în aceeași categorie ca tulburările de dependență de substanțe în ediția 5th a Manualului de diagnostic și statistic al tulburărilor mintale (Frascella și colab., 2010; APA, 2013). Aceasta implică faptul că PG poate fi cauzată de expunerea cronică la activitatea asemănătoare jocurilor de noroc, că mecanismele comune pot media efectele jocurilor de noroc și ale expunerii la medicament (Zack și Poulos, 2009; Leeman și Potenza, 2012); și că sensibilizarea căilor DA ale creierului poate fi un element important al acestui proces.
Dovezile clinice susțin indirect această posibilitate: Utilizând tomografia cu emisie de pozitroni (PET), Boileau și colegii au descoperit că subiecții PG masculi prezintă o eliberare DA striatală semnificativ mai mare ca răspuns la amfetamină (0.4 mg / kg)Boileau și colab., 2013). Diferențele generale de grup au fost semnificative în striatumul asociativ și somato-sensibil. În striatumul limbic, care include nucleul accumbens, grupurile nu diferă. Cu toate acestea, la subiecții PG, eliberarea DA în striatum limbic corelată direct cu severitatea simptomelor PG. Aceste constatări sunt în concordanță cu sensibilizarea căilor DA ale creierului în PG, dar sugerează, de asemenea, unele diferențe importante cu indivizii dependenți de substanța umană și cu modelul animal clasic de sensibilizare a amfetaminei. Spre deosebire de subiecții PG și de animale expuse la doze mici de amfetamină (cf. Robinson și colab., 1982), oamenii cu dependență de substanță prezintă consistent scăderea eliberării DA la provocarea stimulantă (Volkow și colab., 1997; Martinez și colab., 2007), iar dovezile provenite de la animale sugerează că acest lucru poate reflecta deficitele în funcția DA în timpul stadiilor inițiale de abstinență în urma tiparelor de bruște de abuz de substanțe (Mateo și colab., 2005). În studiile în care se demonstrează sensibilizarea stimulentelor la animale, eliberarea crescută de DA este observată, de obicei, în striatum limbic mai degrabă decât striatul dorsal (asociativ, somatosenzorial) (Vezina, 2004). Cu toate acestea, căutarea de droguri determinată de induse de către animale la animale expuse în mod repetat la cocaină a fost legată de eliberarea îmbunătățită a DA în striatum dorsal, un rezultat considerat a indica o formă mai obișnuită de comportament motivatIto și colab., 2002). Astfel, cresterea globala a eliberarii DA in regiunile dorsale la subiectii PG poate fi legata de cautare de recompense bazata pe obiceiul (inflexibil, rutinizat) care implica o "progresie de la ventral la mai multe domenii dorsale ale striatumului"Everitt și Robbins, 2005, p. 1481), în timp ce eliberarea DA dependentă de severitate în striatum limbic la acești subiecți poate corespunde mai strâns sensibilizării stimulente așa cum a fost modelată în mod obișnuit la animale. Constatările PET nu pot dezvălui dacă hiper-reactivitatea DA a fost o caracteristică preexistentă a acestor subiecți PG, o consecință a expunerii la jocurile de noroc sau rezultatul altui proces în întregime. Pentru a aborda această întrebare, este necesar să se demonstreze inducerea sensibilizării prin expunerea cronică la jocurile de noroc la subiecții care sunt normali înainte de expunere. Acest lucru ridică întrebări cu privire la caracteristicile jocurilor de noroc care determină cel mai probabil sensibilizarea.
Skinner a menționat că programul variabil de întărire a fost fundamental pentru atracția jocurilor de noroc (sau cel puțin pentru persistența acestuia) (Skinner, 1953). Comportamentul de pariere într-un joc de mașină automată este în conformitate cu principiile de bază ale condiționării instrumentale, reflectată de o corelație prospectivă între câștigul monetar și mărimea pariului pe rotiri consecutive (Tremblay și colab., 2011). Astfel, răspunsul operantului cu raport variabil pare să furnizeze un model valabil din exterior pentru jocurile de noroc.
Cercetările recente cu animale oferă un sprijin inițial puternic pentru un efect cauzal al expunerii la jocurile de noroc în ceea ce privește sensibilizarea. Singer și colegii au examinat efectele sesiunilor zilnice 55 1-h ale armării de zaharină fixă (FR20) sau variabilă (VR20) într-o paradigmă operativă a levierului-presă pe răspunsul locomotor la doze mici de amfetamină (0.5 mg / kg) Sprague Dawley) (Singer și colab., 2012). Ei au emis ipoteza că, dacă jocurile de noroc duc la sensibilizare, șobolanii expuși la programul variabil, care imită jocurile de noroc, ar trebui să prezinte un răspuns mai mare la amfetamină decât șobolanii expuși la programul fix. Așa cum a fost prezis, grupul VR20 a prezentat un răspuns locomotor mai mare la amfetamină cu 50% decât grupul FR20. În schimb, grupurile au prezentat o localizare echivalentă după o injecție salină. Aceste constatări confirmă faptul că expunerea cronică la armarea variabilă este suficientă pentru a induce hiper-reactivitatea la o provocare cu DA la animale sănătoase, randomizate pentru programele respective.
Din acest rezultat se ridică o serie de întrebări: În primul rând, în ce măsură contingența percepută - sau lipsa acesteia - între răspunsul operatorului și rezultatul acestuia, mediază aceste efecte? În termeni de învățare, acest efect implică o "așteptare a răspunsului-rezultat", sau un efect similar poate fi văzut în absența unui răspuns operant, adică "o așteptare de stimulare-rezultat" într-o paradigmă Pavloviană (cf. Bolles, 1972)? În al doilea rând, gradul de contingență dintre evenimentul antecedent (răspuns sau stimulare) și rezultatul său influențează gradul de sensibilizare?
A doua întrebare se referă la rolul nesiguranței în sensibilizare. De exemplu, jocurile al căror rezultat este cu adevărat aleatoriu - complet imprevizibil - au un potențial mai mare de a induce sensibilizarea decât jocurile unde șansele de a câștiga sunt clar definite, dar nu aleatoare, chiar dacă rata absolută a recompenselor este scăzută? Cercetarea prezentă a abordat aceste întrebări.
Proiectul experimental a fost informat printr-un studiu semnal privind speranța de recompensă și răspunsul neuronului DA la maimuțe (Fiorillo și colab., 2003). Animalele din acest studiu au beneficiat de o recompensă de suc (în SUA) în cadrul programelor de rapoarte variabile 0, 25, 50, 75 sau 100%. Programările au fost desemnate de 1 de 4 diferite CS (icoane). Programul 0% a oferit o recompensă atât de des ca programul 100%, dar a omis CS. Rata de ardere a neuronilor DA în intervalul dintre debutul CS și livrarea sau omisiunea din SUA a reprezentat măsura cheie dependentă. Studiul a constatat că răspunsul DA a crescut ca o funcție a incertitudinii de livrare a recompensei. Astfel, în cadrul schemei 100% CS a evocat puțină activitate, în cadrul programelor 25 și 75%, CS a evocat niveluri moderate și similare ale activității, iar în cadrul programului 50% CS a evocat activitatea maximă. În fiecare caz, rata de tragere a escaladat pe parcursul intervalului CS-SUA, adică, pe măsură ce speranța se apropia de rezultat.
Aceste constatări indică faptul că activitatea DA nu variază numai dacă este sau nu recompensă (Rata fixă) sau incertă (variabilă), dar variază și proporțional invers proporțional cu cantitatea de informații despre livrarea de recompense transmisă de CS. În condiția 100%, CS evocă speranța de recompensă și, de asemenea, previzionează perfect livrarea. În condițiile 25 și 75%, CS evocă speranța și prezice livrarea de recompense de trei ori din patru. În condiția 50%, CS evocă speranța, dar nu oferă nicio informație despre livrarea de recompense decât din întâmplare. Pe baza constatărilor lor, Fiorillo și colab. a concluzionat: "Această creștere a dopaminei indusă de incertitudine ar putea contribui la proprietățile recompensatoare ale jocurilor de noroc" (p. 1901).
Efectele recompensei variabile de 50% într-o singură sesiune nu ar trebui să se schimbe pe parcursul mai multor sesiuni, deoarece probabilitatea recompensei rămâne cu totul imprevizibilă în fiecare proces. Astfel, atunci când se iau în considerare condițiile care ar maximiza activarea cronică a neuronilor DA în episoadele repetate de jocuri de noroc, programul de 50% ar trebui să genereze efectul cel mai durabil și cel mai robust. Acest lucru este demn de remarcat având în vedere că rata de recompensă pe termen lung (recompensă> 0) observată la mii de rotiri pe un slot machine comercial a fost de 45.8% (Tremblay și colab., 2011). Astfel, recompensa variabilă 50% pare să reflecte cu exactitate programul de plăți administrat de dispozitivele actuale de jocuri de noroc.
Prezentul studiu a folosit aceleași scheme de condiționare ca Fiorillo și colab. într-o expunere cronică, design între grupuri cu șobolani. Animalele au suferit ~ 3 săptămâni de sesiuni de condiționare zilnice, în care un CS (lumină) a fost asociat cu un SU (o cantitate mică de zaharoză). După faza de antrenament, animalele s-au odihnit înainte de evaluarea sensibilizării indexate de răspunsul locomotor la amfetamină. Pe baza literaturii de specialitate, s-a prezis că șobolanii expuși la diferite programe de recompense nu vor diferi în ceea ce privește comportamentul lor locomotor fără medicamente, dar ar prezenta niveluri semnificativ diferite de locomoție după amfetamină, grupul de 50% afișând un răspuns locomotor mai mare la medicament în raport cu celelalte grupuri pe parcursul dozelor, un model care ar fi de așteptat dacă animalele de 50% ar fi fost expuse anterior la doze suplimentare de amfetamină în sine (adică sensibilizare încrucișată).
experimentul 1
Materiale și metode
Subiecții
Patru grupe (n = 8 / grup) de șobolani masculi Sprague-Dawley adulți (300-350 g) (Charles River, St Constant, Quebec, Canada) au fost găzduiți individual în cutii din policarbonat (20 × 43 × 22 cm) 12 ciclu lumină-întuneric. Ei au primit ad libitum accesul la hrană și apă și manipularea zilnică de către un experimentator pentru 2 săptămâni înainte de studiu. Fiecare grup a fost condiționat în unul dintre cele patru scheme de recompensă variabile: 0, 25, 50 sau 100%. Grupul 75% a fost omis în acest studiu inițial, ca Fiorillo și colab. (2003) a găsit o eliberare echivalentă post-CS DA în cadrul programelor de recompensare 25 și 75%, astfel încât ambele condiții au condus la o eliberare mai mare a DA decât la condiția 100% CS-US, dar mai mică decât condiția 50%.
Aparat
Accesul la prezentările de zaharoză și la CS a fost furnizat individual în cutiile de condiționare (33 × 31 × 29 cm). Fiecare cutie a fost echipată cu o revista de întărire, situată pe peretele frontal. O lumină în partea de sus a revistei a servit ca CS. Un dispozitiv de transport cu lichid motorizat, cu comandă solenoidală, ar putea fi ridicat la podeaua magaziei. Evenimentele din cutie au fost controlate de echipamentul și software-ul Med Associates, folosind un program intern scris în MED-PC. Testarea locomotorie a fost efectuată individual în cuști din plexiglas (27 × 48 × 20 cm). Fiecare cușcă era echipată cu un sistem de monitorizare compus din șase celule fotosensibile pentru a detecta mișcarea orizontală.
Procedură
Pregătire. Studiul a fost realizat în conformitate cu orientările etice stabilite de Consiliul canadian privind îngrijirea animalelor. Șobolanii au fost restricționați în funcție de alimente la 90% din greutatea corporală pe durata studiului și adăpostiți individual. Fiecare șobolan a primit 15 zile de pregătire pentru recompensarea sucrozei (10% soluție de apă la 0.06 ml pe recompensă): 5 zile consecutive × 3 săptămâni, cu sfârșit de săptămână. Animalele au fost menținute pe chava standard înainte și după faza de antrenament; expunerea la zaharoză a fost limitată la cele 15 sesiuni de formare 40-min. Fiecare sesiune zilnică a constat în prezentări de stimulare 15 (o lumină, CS), fiecare separată printr-un interval inter-proces al 120 s. Lumina a fost localizată în panoul superior al revistei și a rămas activă pentru 25 s, cu sucroză pusă la dispoziție în ultimele 5 s. În cazul grupului 0, dipperul de zaharoză a fost ridicat la fiecare 140 s (pentru 5 s), dar lumina stimulului nu a fost iluminată. Aceasta a echivalat intervalul dintre prezentările dipperului din grupul 0 și celelalte grupuri (120 + 25 s). Fiecare sesiune de tratament a durat ~ 40 min. În medie, grupul 25 a primit sucroză o dată pentru fiecare patru prezentări CS; grupul 50 a primit sucroză o dată pentru fiecare două prezentări CS, iar grupul 100 a primit sucroză după fiecare prezentare CS.
Testarea. La două săptămâni de la ultima sesiune de acces la zaharoză (sau "condiționat"), a fost evaluat răspunsul locomotor la d-amfetamină (AMPH; ip). Șobolanilor li s-au dat trei sesiuni 2-h pentru a se adapta la cutiile locomotorii, urmate de șase sesiuni de testare AMPH. Durata testelor AMPH a avut loc la intervale 1-wk. În zilele de testare, șobolanilor li sa administrat 30 min pentru a se obișnui cu cutiile, apoi au primit o singură doză 0.5 mg / kg de AMPH urmată, în sesiuni săptămânale separate, cu cinci doze 1.0 mg / kg (o doză pe zi) pe zile de testare 1 prin 5 . Locomoția post-AMPH a fost evaluată pentru 90 min în fiecare sesiune.
Metoda de analiză a datelor
Analizele statistice au fost realizate cu SPSS (v. 16 și v. 21, SPSS Inc., Chicago IL). Răspunsul imediat comportamental la CS a fost evaluat prin prisma nasului în deschiderea în care sa administrat sucroza. Numărul mediu de inflamații ale nasului în acest interval (5 s per studiu) a fost apoi comparat cu numărul mediu de inflamații ale nasului pentru aceeași durată (5 s) medii pentru perioada în care a fost absent CS. Grupul x Sesiunea ANOVA-urilor cu CS-uri prezente și absente au urmărit obținerea de răspunsuri discriminatorii la răspunsul tactic și la răspunsurile nediscriminatorii în cadrul programelor diferite pe parcursul sesiunilor de instruire a sucrozei 15.
Efectele tratamentului asupra răspunsurilor locomotorii au fost evaluate cu ANOVA pentru grupul x sesiune pentru faza de habituire fără droguri (trei sesiuni), pre-sensibilizare 0.5 mg / kg AMPH provocare (o sesiune) și în timpul sesiunii de cinci sesiuni 1 mg / kg AMPH în cazul în care grupurile au fost de așteptat să difere ca răspuns la doze repetate de AMPH. Grupul x Sesiunea ANOVA a evaluat, de asemenea, răspunsurile locomotorii fără medicament în timpul fazei de habituire pre-injectare 30-min din fiecare sesiune de testare AMPH. Comparațiile planificate au evaluat diferența dintre performanța medie pentru grupul 50 față de grupul 0 (fără controlul speranței) și grupul 100 (fără control de incertitudine), prin intermediul t-testuri (Howell, 1992), folosind erorile SM și termenii de eroare df pentru efectul relevant (adică interacțiunea sesiunii de grup sau de grup x) de la ANOVA (Winer, 1971). Analizele trendului polinomial au testat profilul schimbărilor pe parcursul sesiunilor.
Pentru a determina dacă răspunsurile de abordare în prezența și absența CS în timpul sesiunilor de instruire a sucrozei 15 au contribuit la variația răspunsului locomotor la AMPH sau la diferențele mediate de grup în răspunsul AMPH, au fost efectuate analize de urmărire a covarianței (ANCOVA) pe AMPH date locomotorii, inclusiv sumele totale ale nasului (suma pentru sesiunile 15) atunci când CS a fost absent ca covariate. Un efect semnificativ al covariatului ar indica faptul că răspunsurile la abordarea fără droguri au moderat (influențat puterea) efectele grupului sau ale sesiunii. Un declin al semnificației efectelor grupului sau al sesiunii în prezența unei covariate semnificative ar indica faptul că răspunsurile de abordare au mediat (reflectat) efectele grupului sau ale sesiunii. O scădere a importanței efectelor de grup sau de sesiune în absența unui efect covariat semnificativ ar reflecta pur și simplu o pierdere a puterii statistice din cauza realocării df de la termenul de eroare la covarianți și nu ar avea legătură cu interpretarea efectele grupului sau ale sesiunii.
REZULTATE
Nas în timpul sesiunilor de condiționare sucroză
CS prezent. Figura 1A arată frecvența nasului mediu pentru grupurile 25, 50 și 100, în timp ce CS era prezent pe sesiunile de condiționare a sucrozei 15 (nasele nu au fost codificate pentru grupul 0, care nu a primit CS). Un grup 3 × 15 Sesiunea ANOVA a generat efecte principale semnificative ale Grupului, F(2, 21) = 5.63, p = 0.011 și sesiune, F(14, 294) = 14.00, p <0.001, împreună cu o interacțiune semnificativă de grup × sesiune, F(28, 294) = 2.93, p <0.001. Figura 1A indică faptul că efectul principal al sesiunii a reflectat o creștere a numărului de sesiuni din toate cele trei grupuri și efectul principal al grupului a reflectat scoruri generale în general mai mari în grupul 100 față de grupul 25 cu scoruri intermediare în grupul 50. O interacțiune semnificativă a Grupului × Sesiune pentru tendința cubică, F(2, 21) = 4.42, p = 0.030, a indicat o creștere rapidă, scufundare și nivelare în nas în timpul sesiunilor din grupa 100, față de o creștere liniară față de sesiunile din grupul 50 și o creștere liniară mai mică față de sesiunile din grupul 25.
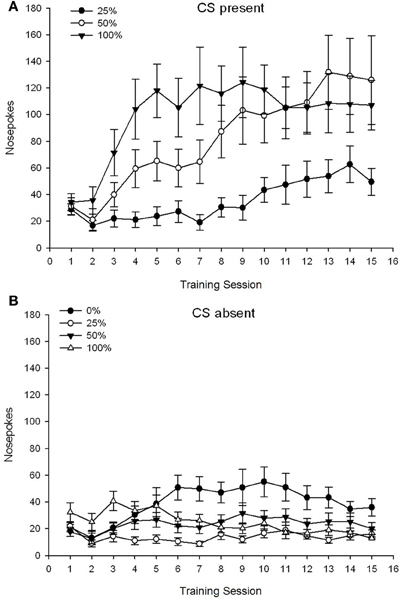
Figura 1. Răspunsurile de abordare medie (SE) (șobolanii) pe sesiunile de antrenament 15 pentru zaharoză în grupuri de șobolani Sprague Dawley (n = 8 / grup) expuse la recompensa de zaharoză (soluție 10%) livrată în cadrul programelor variabile 0, 25, 50 sau 100%. Stimul condiționat a fost o lumină (120 s). Grupul 0 a primit același număr de recompense ca grupul 100 în absența stimulilor condiționați. (A) S-au înregistrat scoruri CS (studii 5 s × 15). (B) S-au scorat când CS a fost absent (media pentru 5 × 15 s în timp ce lumina a fost oprită).
CS absent. Figura 1B arată vârstele medii ale nasului pentru toate cele patru grupe pentru o durată echivalentă (studiile 5 s × 15) medii pentru perioada în care a fost absent CS. Un grup 4 × 15 Sesiunea ANOVA a generat efecte principale semnificative ale Grupului, F(3, 28) = 7.06, p = 0.001 și sesiune F(14, 392) = 2.84, p <0.001, împreună cu o interacțiune semnificativă de grup × sesiune, F(42, 392) = 3.93, p <0.001. O interacțiune semnificativă de grup × sesiune pentru trendul pătratic, F(3, 28) = 3.91, p = 0.019, împreună cu nici o interacțiune pentru tendința cubică, F(3, 28) <0.93, p > 0.44, a reflectat un profil „U inversat” al nasului în timpul sesiunilor din grupa 0, comparativ cu un profil în general stabil în timpul sesiunilor din celelalte grupuri.
Habitatarea camerelor locomotorii
O sesiune 4 × 3 ANOVA a generat un efect principal al sesiunii, F(2, 56) = 5.67, p = 0.006, și nici alte efecte semnificative, F(3, 28) <1.60, p > 0.21. Pauzele medii (SE) ale fasciculului la 2 ore în cutiile locomotorii au fost 1681 (123) în sesiunea 1, 1525 (140) în sesiunea 2 și 1269 (96) în sesiunea 3. Comparațiile planificate nu au găsit diferențe semnificative între grupul 50 și grup 0 sau grupul 100 la prima sau ultima sesiune de obișnuință, t(84) <1.69, p > 0.05. Astfel, în absența AMPH, expunerea repetată la casetele de testare a fost asociată cu o scădere consistentă a activității locomotorii spontane în cele patru grupuri (adică efectul sesiunii) și fără răspuns diferențial în funcție de programul de antrenament al zaharozei (fără interacțiune) .
Sesiuni de testare
Efectele pre-sensibilizării 0.5 mg / kg provocare AMPH.
Locomoție pre-injectabilă. Un ANOVA cu o singură cale 4 a răspunsului locomotor în timpul fazei de habituire pre-injectare 30-min nu a avut efecte semnificative, F(3, 28) <1.05, p > 0.38. Comparațiile planificate nu au găsit nicio diferență semnificativă între grupul 50 și grupul 0 sau grupul 100, t(32) <0.87, p > 0.40. Prin urmare, diferențele inițiale în locomoția pre-injecție nu au luat în considerare diferențele de grup în răspunsul locomotor la AMPH. Pauzele medii (SE) ale fasciculului pentru eșantion au fost de 559 (77).
Locomoția post-injectare față de sesiunea de obișnuință a consumului de droguri. O sesiune de 4 grupuri × 2 ANOVA a comparat răspunsurile locomotorii ale grupurilor la ultima sesiune de obișnuință și imediat după provocarea pre-sensibilizare de 0.5 mg / kg AMPH. Scorurile pentru sesiunea de obișnuință (120 min) au fost reduse pentru a corespunde cu durata sesiunii de testare AMPH (90 min) (scor brut de obișnuință × 90/120). Analiza a produs un efect principal semnificativ al sesiunii, F(1, 28) = 34.16, p <0.001 și fără alte efecte semnificative, F(3, 28) <2.26, p > 0.10. Efectul sesiunii a reflectat o creștere a pauzelor medii (SE) ale fasciculului ca răspuns la doză, de la 952 (72) la 1859 (151). Comparațiile planificate nu au găsit diferențe semnificative între grupul 50 și grupul 0 sau grupul 100 ca răspuns la doză, t(56) <1.72, p > 0.10. Cu toate acestea, ordinea rangului scorurilor de rupere a fasciculului (M; SE) aliniată cu ipoteza: grupul 50 (2205; 264)> grupul 0 (2025; 203)> grupul 100 (1909; 407)> grupul 25 (1296; 299) .
Efectele 1 mg / kg AMPH.
Locomoție pre-injectabilă. O sesiune 4 × 5 ANOVA a răspunsului locomotor în timpul fazei de habituare pre-injectare 30-min pe 1 mg / kg sesiuni de testare AMPH a generat un efect principal al sesiunii, F(4, 112) = 43.64, p <0.0001 și fără alte efecte semnificative, F(3, 28) <0.97, p > 0.42. Comparațiile planificate nu au găsit nicio diferență semnificativă între grupul 50 și grupul 0 sau grupul 100 la prima sau ultima sesiune de test, t(140) <0.84, p > 0.30. Prin urmare, diferențele de bază în locomoție nu au luat în considerare diferențele de grup în răspunsul locomotor la AMPH. Scorurile medii (SE) de rupere a fasciculului pentru faza de obișnuință pre-doză la sesiunile 1-5 au fost: 454 (30), 809 (53), 760 (36), 505 (35), 756 (39).
Locomoția post-injectabilă. Figura 2 arată efectele a cinci injecții de 1 mg / kg AMPH (câte unul pe săptămână) asupra scorurilor de activitate locomotorie în cele patru grupuri. O sesiune 4 × 5 ANOVA a generat un efect principal al sesiunii, F(4, 112) = 8.21, p <0.001, un efect principal marginal al grupului, F(2, 45) = 3.28, p = 0.085, și nici o interacțiune semnificativă, F(12, 122) <0.77, p > 0.68.
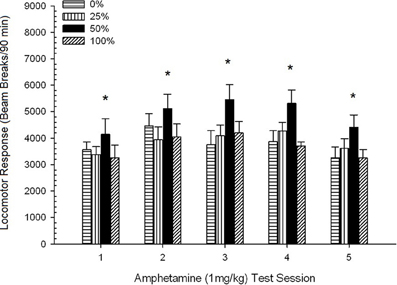
Figura 2. Răspunsul locomotor mediu (SE) (numărul de rupturi ale fasciculului într-o gamă electronică pe 90 min) la 1 mg / kg d-amfetamină (ip) pe sesiuni săptămânale 5 în grupuri de șobolani Sprague Dawleyn = 8 / grup) expuse anterior la sesiunile de condiționare zilnică 15 cu recompensă pentru sucroză (soluție 10%) livrate în cadrul programelor variabile 0, 25, 50 sau 100%. Stimul condiționat a fost o lumină (120 s). Grupul 0 a primit același număr de recompense ca grupul 100 în absența stimulilor condiționați. *p <0.05 pentru diferența medie între grupul 50 și grupul 0, precum și grupul 100, pe baza comparațiilor planificate.
Comparațiile planificate au arătat că scorurile grupului 50 diferă semnificativ de grupul 0, t(14) = 2.19, p = 0.037 și grupul 100, t(14) = 2.36, p = 0.025 [și diferă marginal de grupul 25, t(14) = 2.03, p = 0.051]. Astfel, în grupul 50, răspunsul locomotor la 1 mg / kg AMPH a depășit în mod sigur numărul celorlalte trei grupuri în toate cele cinci sesiuni de testare. Analiza tendințelor polinomiale a detectat o tendință semnificativă în cadrul sesiunilor, F(1, 28) = 32.47, p <0.0001 și fără alte tendințe semnificative, F(1, 28) <1.78, p > 0.19. Figura 2 arată că acest rezultat a reflectat un model "inversat U" în timpul sesiunilor.
Controlați variația nasului care răspunde în timpul antrenamentului cu zahăr
Următoarea ANCOVA a răspunsurilor locomotorii la 1 mg / kg AMPH, cu nasuri (CS prezente) ca covariate, în cele trei grupuri care au primit CS, a dat un efect principal marginal al grupului, F(2, 20) = 3.07, p = 0.069, și nu au efecte legate covariante semnificative, F(4, 80) <0.05, p > 0.85. Astfel, abordarea cued răspuns în timpul antrenamentului nu a explicat variații semnificative în răspunsul locomotor la 1 mg / kg AMPH în grupurile 25, 50 sau 100.
Următoarea analiză ANCOVA a răspunsurilor locomotorii la 1 mg / kg AMPH, cu gonflarea nasului (CS absent) ca o covariată, a produs un efect semnificativ al covariatului, F(1, 27) = 6.17, p = 0.020, un efect principal semnificativ al Grupului, F(3, 27) = 4.13, p = 0.016, o interacțiune marginală Session × Covariate, p = 0.080, și nici alte efecte semnificative, F(4, 108) <1.48, p > 0.21. Astfel, abordarea fără indicii (fără discriminare) care răspunde în timpul antrenamentului a explicat variații semnificative ale răspunsului locomotor la 1 mg / kg AMPH. Cu toate acestea, această variație nu se suprapune cu varianța legată de grup, deoarece includerea covariatei în analiză a crescut mai degrabă decât a scăzut semnificația efectului de grup.
Discuție
Datele nasului în timp ce CS-ul a fost prezent arată că grupurile au dobândit asocierea dintre CS și furnizarea sucrozei, așa cum se reflectă printr-o creștere a răspunsurilor cued peste sesiunile de antrenament. Profilul răspunsurilor la sesiuni în timp ce CS era prezent a sugerat că programele 100 și 50% CS-US au fost la fel de eficiente în abordarea abordării, în timp ce programul 25% a determinat o creștere mai modestă a abordării induse de tac. Datele privind nasul în timp ce CS a fost absent sugerează că grupurile care au primit oricare dintre cele trei programe de pregătire pentru CS-zaharoză (grupul 25, 50, 100) au învățat rapid să reducă nasul în absența CS, în timp ce animalele din grupul 0 , care nu au primit CS, au învățat să-și micșoreze comportamentul de abordare într-o măsură limitată după o pregătire extensivă.
Datele privind obișnuirea arată că grupurile nu au fost diferite înaintea AMPH și că expunerea repetată la cutiile de testare a fost asociată cu un răspuns locomotor redus fără medicament. Prin urmare, diferențele dintre grupuri și răspunsul crescut față de dozele repetate de AMPH nu pot fi atribuite diferențelor preexistente în comportamentul locomotor.
Rezultatele provocării pre-sensibilizării cu 0.5 mg / kg AMPH au confirmat că medicamentul a crescut activitatea locomotorie în raport cu ziua de obișnuință fără uz de droguri. În concordanță cu ipoteza, grupul 50 se situează mai înalt decât grupurile 0 sau 100 (precum și grupul 25) în ceea ce privește răspunsul mediu la doză, deși diferențele medii între grupuri nu au fost semnificative.
Pentru sesiunile de sensibilizare, comparațiile planificate între grupuri au arătat că expunerea anterioară la recompensa de zaharoză condiționată de 50% a dus la o creștere semnificativă a răspunsului locomotor la o doză de 1.0 mg / kg de amfetamină față de celelalte trei scheme. Acest efect a fost evident de la prima doză și nu s-a modificat semnificativ în cazul dozelor repetate. Analiza tendinței a indicat un răspuns bifazic (pentru proba completă) la doze repetate de AMPH, crescând până la a treia doză și scăzând după aceea. Rezultatele ANCOVA de urmărire cu lovituri de nas (CS absente) ca covariate au confirmat că diferențele în răspunsurile locomotorii ale celor patru grupuri la 1 mg / kg AMPH nu au fost mediate de abordarea fără răspuns în timpul sesiunilor de antrenament zaharoză.
Efectul de grup în timpul sesiunilor de sensibilizare este în concordanță cu ipoteza noastră. Efectul sesiunii bi-fazice nu este în concordanță cu escaladarea preconizată a răspunsurilor locomotorii cu doze repetate de AMPH. Aceasta poate fi legată de intervalul de dozare. Pentru a aborda această problemă, ar trebui să se utilizeze o procedură (doze zilnice alternative) care să indice o creștere esențială a răspunsului locomotor la doze 1.0 mg / kg de AMPH (adică sensibilizarea comportamentală). Impactul unui regim sensibilizat al AMPH asupra răspunsului ulterior la o a doua provocare 0.5 mg / kg ar susține în continuare generalizarea acestui efect. Includerea unei provocări saline înaintea AMPH ar determina rolul speranței sau efectelor legate de injectare (de exemplu, stresul) asupra răspunsului locomotor la AMPH. Includerea unui grup de zaharoză condiționată cu 75% ar ajuta la clarificarea rolului incertitudinii de răsplată față de rata de recompensă asupra tiparului de răspunsuri pentru grupurile 50 și 25. În plus, pentru a permite evaluarea (prin ANCOVA) a aportului de răspunsuri de abordare fără răspuns medicamentos la locomoție sub AMPH (folosind nasul cu CS prezent ca covariate), nasurile au fost de asemenea codate pentru grupa 0 în intervalul în care CS a fost prezentă în celelalte patru grupuri (adică, astfel încât nasul din toate cele cinci grupuri - inclusiv grupul 0 care nu a primit CS - ar putea fi inclus în analiza covarianței cu CS prezentată ca covariat). Aceste rafinări au fost încorporate în experimentul 2.
experimentul 2
Materiale și metode
Metodologia experimentului 2 a fost similară cu cea a experimentului 1, dar a fost revizuită pentru a apropia mai bine un regim găsit pentru a determina în mod fiabil sensibilizarea AMPH (Fletcher și colab., 2005). Modificările au fost după cum urmează: (a) grupul 75% CS-zaharoză (n = 8); (b) În timpul antrenamentului cu zaharoză, șobolanii (cu excepția grupului 0) au primit prezentări 20 CS (ușoare) (spre deosebire de 15 în experimentul 1); (c) prezentările CS s-au separat fiecare printr-un interval mediu inter-proces al 90 s; : 30-180 s (față de 120 s în experiment 1), care au compensat creșterea studiilor de antrenament pentru a egala durata fiecărei sesiuni de antrenament cu cea a experimentului 1; (d) durata fiecăreia dintre cele trei sesiuni de obișnuință a fost redusă de la 120 la 90 min pentru a corespunde duratei sesiunilor de testare; (e) A fost adăugată o soluție salină (ip, 1 ml / kg) (90 min) (zi de antrenament post-zaharoză 8), pentru a evalua efectele locomotorii ale injectării în sine (de exemplu, așteptare, stres); (f) Sesiunile de sensibilizare 1 mg / kg s-au desfășurat în zilele săptămânii alternative (zile post-antrenament 12-21), mai degrabă decât la intervale săptămânale, ca în experimentul 1; (g) Pe lângă pre-sensibilizarea 0.5 mg / kg AMPH provocare (ziua post-antrenament 9) a fost adăugată oa doua reacție post-sensibilizare 0.5 mg / kg AMPH (zi de antrenament post-zahăr 28), pentru a testa genericitatea efectul de sensibilizare în doze; (h) bolile nasului în timp ce CS a fost prezent au fost codificate pentru toate grupurile (inclusiv grupa 0); (i) nasurile în timp ce CS a fost absent au fost înregistrate în mod specific din intervalul 5 imediat înainte de debutul CS la indexarea abordării premature care răspunde.
REZULTATE
Nas în timpul sesiunilor de condiționare sucroză
O grupă 5 × Sesiune 15 × Faza 2 (CS prezent, CS absent) ANOVA a nasului a generat efecte principale semnificative ale grupului, F(4, 19) = 2.89, p = 0.050, sesiune F(14, 266) = 2.28, p = 0.006, și Phase, F(1, 19) = 14.72, p = 0.001, precum și o interacțiune semnificativă în trei direcții, F(56, 266) = 1.38, p = 0.050. panouri (A, B) din figura 3 trasați punctajele medii ale grupului cu vârfurile nasului pentru fazele CS prezente și respectiv CS absente. Comparația celor două panouri relevă faptul că efectul principal al fazei a reflectat mai multe răspunsuri generale la vârful nasului atunci când CS a fost prezent vs. Prin urmare, răspunsurile cued au apărut semnificativ mai des decât răspunsurile premature fără răspuns. Efectele principale ale grupului și sesiunii nu au fost interpretate cu ușurință din cauza interacțiunii de ordin superior. Acest ultim rezultat a reflectat o convergență a scorurilor pentru cele cinci grupuri la un nivel relativ stabil scăzut între sesiuni când CS a fost absent (Figura 3B), împreună cu o diferență de scoruri în grupuri mari (grup 75, grup 100), intermediar (grup 50) și scăzut (grup 0, grup 25) 3A). Din tendințele polinomiale de ordine inferioară (liniară, patratică, cubică), numai interacțiunea cu trei căi pentru tendința liniară a abordat semnificația, F(4, 19) = 2.32, p = 0.094, reflectând creșterea în general monotonă a naselor în timpul grupului 75 și stabilizarea relativ mai rapidă la niveluri ridicate, intermediare și scăzute ale răspunsului la celelalte grupuri atunci când a fost prezent CS.
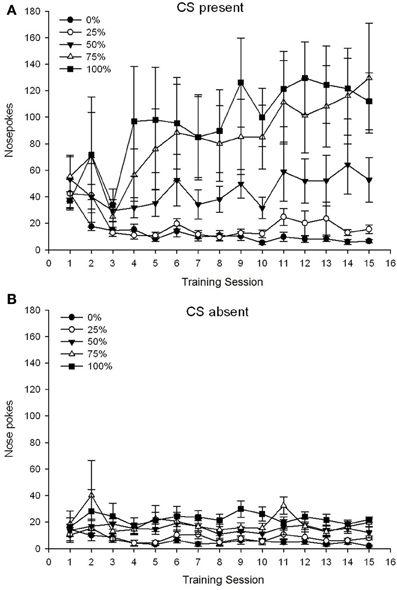
Figura 3. Răspunsurile de abordare medie (SE) (șobolanii) pe sesiunile de antrenament 15 pentru zaharoză în grupuri de șobolani Sprague Dawley (n = 8 / grup) expuse la recompensa de zaharoză (soluție 10%) livrată în cadrul programelor variabile 0, 25, 50, 75 sau 100%. Stimul condiționat a fost o lumină (120 s). Grupul 0 a primit același număr de recompense ca grupul 100 în absența stimulilor condiționați. (A) S-au înregistrat scoruri CS (studii 5 s × 20). (B) S-au scorat când CS a fost absent (media pentru 5 × 20 s în timp ce lumina a fost oprită).
Habitat pentru cutiile locomotorii
O sesiune 5 × 3 ANOVA de răspunsuri locomotorii fără droguri a produs un efect principal semnificativ al sesiunii, F(2, 70) = 60.01, p <0.0001 și fără alte efecte semnificative, F(4, 35) <0.70, p > 0.60. Comparațiile planificate ale grupului 50 cu grupul 0 și cu grupul 100 la prima și ultima sesiune de obișnuință nu au avut efecte semnificative, t<0.84, p > 0.40. Prin urmare, răspunsul locomotor mediu fără droguri în grupurile cheie nu a diferit înainte de testare. Numărul mediu (SE) de pauze ale fasciculului pe 90 de minute a fost de 2162 (118) în sesiunea 1, 1470 (116) în sesiunea 2 și 1250 (98) în sesiunea 3.
Sesiuni de testare
Ser fiziologic. O sesiune 5 Group × 2 ANOVA a comparat răspunsul locomotor la sesiunea de obișnuință de obișnuință și la sesiunea de provocare salină. ANOVA a generat un efect principal al sesiunii, F(1, 35) = 62.46, p <0.0001 și fără alte efecte semnificative, F(4, 35) <0.65, p > 0.64. Figura 4 arată modul în care grupul indică faptul că efectul Sesiunii a reflectat o scădere generală a răspunsului locomotor din sesiunea de obișnuință de hrănire fără droguri până la sesiunea de salină, care nu a variat în funcție de grup. Astfel, declinul răspunsului locomotor observat în cele trei sesiuni de habituare a continuat cu cea de-a patra expunere fără droguri la cutiile de test.
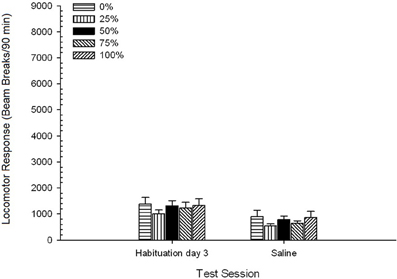
Figura 4. Răspunsul locomotor mediu (SE) (numărul de rupturi ale fasciculului într-o gamă electronică pe 90 min) pe ultima sesiune de habituire fără droguri 3 și pe o sesiune ulterioară după injectarea salinei (ip, 1 ml / kg) în grupuri de Sprague Dawley șobolani (n = 8 / grup) expuse anterior la sesiunile de condiționare zilnică 15 cu recompensă pentru sucroză (soluție 10%) livrate în cadrul programelor variabile 0, 25, 50, 75 sau 100%. Stimul condiționat a fost o lumină (120 s). Grupul 0 a primit același număr de recompense ca grupul 100 în absența stimulilor condiționați.
Efectele 0.5 mg / kg AMPH.
Locomoție pre-injectabilă. O sesiune 5 × 2 ANOVA de locomoție pre-injectabilă (30-min) la zilele de testare pre-și post-sensibilizare 0.5 mg / kg AMPH a generat un efect principal semnificativ al sesiunii, F(1, 35) = 13.39, p = 0.001, și nici alte efecte semnificative, F(4, 35) <1.79, p > 0.15. Comparațiile planificate nu au găsit diferențe semnificative între grupul 50 și grupul 0 sau grupul 100 în prima sesiune, t(70) <1.00, p > 0.30. Cu toate acestea, în cea de-a doua sesiune (post-sensibilizare), grupul 50 (1203; 121) a afișat semnificativ mai multe rupturi ale fasciculului pre-injecție (M; SE) decât grupul 100 (756; 103), t(70) = 5.11, p <0.001, dar nu diferă de grupul 0 (1126; 211), t(7) <0.88, p > 0.40. Prin urmare, diferențele inițiale de locomoție nu au luat în considerare diferențele de grup în răspunsul locomotor la prima doză de 0.5 mg / kg de AMPH, dar ar fi putut contribui la diferențe între grupul 50 și grupul 100 în răspunsul locomotor la a doua doză de 0.5 mg / kg de AMPH . Pauzele medii (SE) ale fasciculului pentru faza de pre-injecție la prima și a doua sesiune de test de 0.5 mg / kg AMPH au fost 757 (41) și 974 (59).
Locomoția post-injectabilă. O sesiune 5 × 2 ANOVA a răspunsului locomotor la 0.5 mg / kg AMPH înainte și după schema de sensibilizare a dozei de 5 a produs un efect principal al sesiunii, F(1, 35) = 76.05, p <0.0001 și fără alte efecte semnificative, F(4, 35) <1.10, p > 0.37. Figura 5 arată scorurile medii pentru fiecare grup și sesiune.
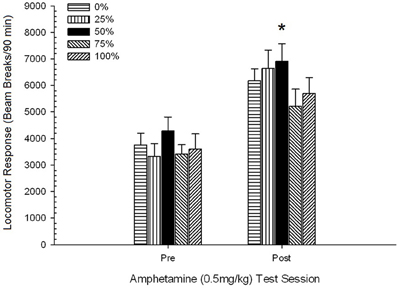
Figura 5. Raspunsul locomotor mediu (SE) (numarul de spargeri ale fasciculului intr-o matrice electronica pe 90 min) la 0.5 mg / kg d-amfetamina pe sesiuni separate inainte si dupa o sesiune de sesizare 5 a d-amfetaminei (1.0 mg / kg; per ședință) în grupuri de șobolani Sprague Dawley (n = 8 / grup) expuse anterior la sesiunile de condiționare zilnică 15 cu recompensă pentru sucroză (soluție 10%) livrate în cadrul programelor variabile 0, 25, 50, 75 sau 100%. Stimul condiționat a fost o lumină (120 s). Grupul 0 a primit același număr de recompense ca grupul 100 în absența stimulilor condiționați. *p <0.05 pentru diferența medie între grupul 50 și grupul 0, precum și grupul 100, pe baza comparațiilor planificate.
Figura arată că efectul sesiunii a implicat o creștere semnificativă a întreruperii fasciculului mediei (SE) pe 90 min de la 0.5 mg / kg doză 1, 3674 (216) la 0.5 mg / kg doză 2, 6123 (275). Lipsa interacțiunii sau a efectului de grup a sugerat că sensibilizarea la AMPH nu a variat fiabil în rândul grupurilor. În ciuda lipsei de efecte semnificative legate de grup în ANOVA, inspectarea figurii arată că grupul 50 a prezentat cel mai mare răspuns atât la prima, cât și la cea de-a doua doză de 0.5 mg / kg. Comparațiile planificate ale răspunsului la prima doză de 0.5 mg / kg nu au evidențiat diferențe semnificative între grupul 50 și grupul 0 sau grupul 100, t„s(35) <0.48, p > 0.50. Cu toate acestea, ca răspuns la a doua doză (post-sensibilizare) de 0.5 mg / kg, grupul 50 a prezentat locomoție semnificativ mai mare decât grupul 0, t(35) = 2.00, p <0.05, precum și grupul 100, t(35) = 3.29, p <0.01.
Având în vedere diferența semnificativă a grupului de locomoție pre-injectabilă la cea de-a doua sesiune 0.5 mg / kg AMPH raportată mai sus, s-a efectuat o urmărire 5 Group × 2 Session ANCOVA a răspunsului locomotor la 0.5 mg / kg AMPH, locomoție de injectare în a doua sesiune. Această analiză a determinat un efect semnificativ al covariatului, F(1, 34) = 8.65, p = 0.006, efectul principal al sesiunii F(1, 34) = 10.83, p = 0.002, și nici alte efecte semnificative, F(4, 34) <0.85, p > 0.50. Foarte important, comparațiile planificate bazate pe eroarea MS și eroarea df de la ANCOVA au confirmat că răspunsul locomotor mediu la a doua doză de 0.5 mg / kg de AMPH a rămas semnificativ mai mare în grupul 50 decât în grupul 100, t(34) = 3.09, p <0.01 și grupul 0, t(34) = 1.88, p <0.05 (cu o singură coadă), când variația pre-injecției de la sesiunea 2 a fost controlată. Astfel, grupul 50 a prezentat un răspuns locomotor post-sensibilizare semnificativ mai mare la 0.5 mg / kg AMPH decât grupul 100 sau grupul 0, iar aceste diferențe de grup nu au fost mediate de locomoția pre-injecție în zilele de testare.
Efectele 1.0 mg / kg AMPH.
Locomoție pre-injectabilă. O sesiune 5 Group × 5 ANOVA de scoruri pre-injectare 30-min pentru sesiunile de sensibilizare 1 mg / kg AMPH a generat un efect principal al sesiunii, F(4, 140) = 16.70, p <0.0001 și fără alte efecte semnificative, F(4, 35) <0.94, p > 0.45. Comparațiile planificate nu au găsit nicio diferență semnificativă în locomoția pre-injecție între grupul 50 și grupul 0 sau grupul 100 în prima sesiune, t(175) <1.66, p > 0.10. Cu toate acestea, în ultima sesiune, grupul 50 (1167; 140) a afișat semnificativ mai multe pauze ale fasciculului (M; SE) decât grupul 100 (1000; 99), t(175) = 2.35, p <0.05, dar nu diferă de grupul 0 (1085, 120), t(175) <1.16, p > 0.20. Prin urmare, diferențele în locomoția pre-injecție au contribuit la diferențe între grupurile 50 și 100 în răspunsul locomotor la doza finală de 1 mg / kg AMPH. Pauzele medii (SE) ale fasciculului global pentru eșantion în timpul fazei de pre-injecție pentru sesiunile 1 până la 5 au fost: 810 (46), 784 (52), 760 (53), 726 (46), 1009 (51).
Locomoția post-injectabilă. O sesiune 5 × 5 ANOVA a răspunsurilor la 1 mg / kg AMPH a generat un efect principal semnificativ al sesiunii, F(4, 140) = 6.72, p <0.001, o interacțiune marginală de grup × sesiune, F(16, 140) = 1.57, p = 0.085, și nici un efect principal al Grupului, F(4, 35) <0.44, p > 0.77. Analizele tendințelor polinomiale au relevat o tendință liniară semnificativă, F(1, 35) = 9.19, p = 0.005, și trendul cubic, F(1, 35) = 21.63, p <0.001, peste sesiunile de la 1 la 5. Figura 6 arată scorurile locomotorii medii pentru fiecare grup și sesiune.
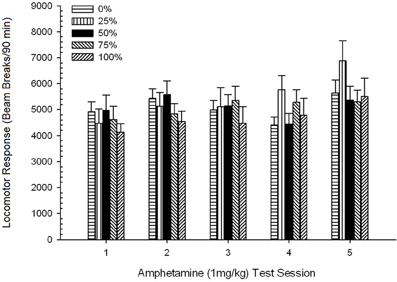
Figura 6. Răspunsul locomotor mediu (SE) (numărul de rupturi ale fasciculului într-o gamă electronică pe 90 min) la 1 mg / kg d-amfetamină (ip) pe sesiuni săptămânale 5 în grupuri de șobolani Sprague Dawleyn = 8 / grup) expuse anterior la sesiunile de condiționare zilnică 15 cu recompensă pentru sucroză (soluție 10%) livrate în cadrul programelor variabile 0, 25, 50, 75 sau 100%. Stimul condiționat a fost o lumină (120 s). Grupul 0 a primit același număr de recompense ca grupul 100 în absența stimulilor condiționați.
Figura arată că efectul Sesiunii a reflectat o creștere semnificativă a întreruperii fasciculului mediu (SE) pentru întregul eșantion de la sesiunea 1, 4624 (213) la sesiunea 5, 5736 (272), confirmând apariția sensibilizării la AMPH. Tendința cubică a indicat maximele relative la sesiunile 1, 3 și 5, cu scufundări la sesiunile 2 și 4, în special pentru grupurile 0 și 50. Cifra, de asemenea, arată că, în ciuda lipsei unei interacțiuni semnificative, grupul 25 a afișat un răspuns locomotor progresiv mai mare în timpul sesiunilor și sa deosebit considerabil de celelalte grupuri la sesiunile 4 și 5 (9 și 22% mai mare decât cel mai mare grup de vârf). Comparațiile planificate au constatat că grupul 50 nu diferă semnificativ de grupurile 0 sau 100, t(175) <0.89, p > 0.40 la prima sau ultima sesiune de test de 1 mg / kg AMPH.
Controlați variația nasului care răspunde în timpul antrenamentului cu zahăr
Două grupuri 5 x 2 ANCOVA ale răspunsului locomotor la 0.5 mg / kg AMPH înainte și după schema de sensibilizare, incluzând nivelele totale ale nasului în timpul antrenamentului cu zaharoză cu CS prezent și CS absent ca covariate separate, nu au găsit efecte semnificative nici pentru covariate, F(1, 18) <1.03, p > 0.31. Prin urmare, abordarea care a răspuns în timpul antrenamentului nu a mediat diferențele de grup ca răspuns la 0.5 mg / kg AMPH.
Două grupuri 5 x 5 Sesiunea ANCOVA a răspunsului locomotor la 1 mg / kg în timpul sesiunilor de sensibilizare cu nasul total (CS prezent, CS absent) ca covariate separate nu a produs efecte semnificative ale covariatului în timp ce CS a fost prezent, F(4, 104) <1.04, p > 0.38 și un efect principal marginal al covariatei în timp ce CS a fost absent, F(1, 18) = 3.32, p = 0.085.
Discuție
Rezultatele acestui studiu nu au susținut în mod constant ipoteza că grupul 50 ar demonstra un răspuns locomotor mai mare în timpul sesiunilor comparativ cu celelalte grupuri. Datele 1 mg / kg AMPH confirmă apariția sensibilizării cu regimul de dozare alternativă. Modelul din grupuri a indicat o tendință de sensibilizare mai mare în timpul ultimelor sesiuni din grupul 25, fără dovezi pentru grupul 50. În contrast, rezultatele dozei 0.5 mg / kg au indicat o tendință de sensibilizare mai mare la grupul 50, confirmând în același timp o creștere globală semnificativă a răspunsului locomotor pe grupuri față de cea de-a doua versus prima doză 0.5 mg / kg AMPH. Efectul nul al injectării saline a confirmat că speranța sau stresul asociat injectării nu au contribuit la efectele AMPH.
Datele privind nasul din nou au evidențiat o creștere generală a abordării care a răspuns în cursul sesiunilor de formare atunci când CS a fost prezent, fără o creștere corespunzătoare atunci când CS a fost absent. Prin urmare, animalele par să dobândească asocierea dintre CS și perspectiva recompensei de zaharoză. Diferențele de grup în ceea ce privește frecvența nasului în momentul prezenței CS corespundeau în mod frecvent cu frecvența de livrare a recompenselor în cadrul programelor respective, grupurile 75 și 100 afișând cele mai multe vârfuri ale nasului, grupul 50 afișează numerele intermediare ale nasului și grupurile 0 și 25 care afișează cele mai puține bătăi ale nasului. Aceste rezultate sugerează că CS a venit să controleze abordarea care răspunde într-un mod compatibil cu probabilitatea generală de recompensă. Deși speculativă, o posibilă explicație pentru ratele de nas mai mici cu CS prezentă în grupul 50 în experimentul 2 față de experimentul 1 poate fi scurtarea intervalului inter-proces, deoarece intervale inter medii mai mari (experiment 1) par să încurajeze impulsiv tendințe și acest lucru este asociat cu creșterea cifrei de afaceri a DA în cortexul anterior cingulate, prelimbic și infralimbic (Dalley și colab., 2002). Prin urmare, reducerea cu 30% a intervalului inter-experimental în experimentul 2 (și 3) poate avea modificări ale nivelelor DA corticale și promovată mai selectivă (adică, ghidată de frecvența relativă a recompenselor) față de abordarea impulsivă care răspunde în grupul 50 în timpul studiilor de formare în experimentul 2 comparativ cu experimentul 1.
Lipsa efectelor semnificative legate de covariate pentru nasul în starea CS prezentă în ANCOVA indică faptul că abordarea care răspunde în timpul antrenamentului cu zaharoză nu a mediază efectele diferitelor scheme de CS-zaharoză asupra răspunsurilor la AMPH. Efectul semnificativ marginal al covariatului pentru starea absentă CS în ANCOVA al răspunsurilor locomotorii la 1 mg / kg AMPH sugerează că tendința de a răspunde fără medicament prematură a explicat o parte din variabilitatea efectelor locomotorii ale AMPH în timpul sesiunilor de sensibilizare.
Împreună, dovezile sugerează că efectele istoricului condiționării pot fi mai discerneabile cu 0.5 AMPH decât cu 1 mg / kg AMPH și că un protocol care generează sensibilizare în absența oricărei alte manipulări poate observa sau redundanța efectelor unui presupus sensibilizarea care promovează manipularea comportamentală (adică, recompensa cronică variabilă).
Sensibilizarea comportamentală la AMPH este un efect robust în laborator. Cu toate acestea, în afara laboratorului, doar o minoritate de indivizi care joacă un rol escaladat cronic la niveluri patologice. Deși riscul de sensibilizare este legat de riscul de dependență (sau solicitarea de droguri), în special pentru psihostimulatori (Vezina, 2004; Flagel și colab., 2008), mulți factori în afara riscului de sensibilizare pot predispune la dependență (de exemplu, Verdejo-Garcia și colab., 2008; Conversano și colab., 2012; Volkow și colab., 2012). Cu toate acestea, factorii de trăsătură care conferă vulnerabilitate sensibilizării pot interacționa cu istoria condiționării pentru a accentua efectele unei recompense imprevizibile (de exemplu, programul 50% CS-US) asupra reactivității sistemului DA. Pentru a investiga această posibilitate, experimentul 3 a folosit aceeași procedură ca experimentul 2, dar a folosit tulpina Lewis în locul șobolanilor de tulpină Sprague Dawley.
Șobolanii Sprague Dawley prezintă nivele intermediare de transportatori DA, cu niveluri mai scăzute decât șobolanii tulpini Wistar (Zamudio și colab., 2005), dar niveluri mai ridicate decât șobolanii Wistar-Kyoto (o tulpină "depresivă") în nucleul accumbens, amigdala, zona tegmentală ventrală și substantia nigra (Jiao și colab., 2003). Acest profil poate face șobolanii Sprague Dawley să fie sensibili doar la mediul sau manipularea farmacologică a funcției DA. În contrast, șobolanii Lewis prezintă niveluri scăzute de transportatori DA, precum și receptorii D2 și D3 DA în nucleul accumbens și striatum dorsal comparativ cu alte tulpini (de exemplu, F344) (Flores și colab., 1998). Aceste diferențe morfologice pot contribui la răspunsul diferențial al șobolanilor Lewis la manipulările DA. Șobolanii Lewis prezintă, de asemenea, o serie de răspunsuri accentuate la manipulările experimentale ale medicamentelor comparativ cu alte tulpini (de exemplu, F344). Cel mai important, șobolanii Lewis prezintă o sensibilizare mai mare la metamfetamină, caracterizată printr-un răspuns scăzut la dozele inițiale, dar un răspuns mai mare la dozele ulterioare (Camp și colab., 1994). Lewis, de asemenea, prezintă o sensibilizare locomotorie mai mare la o serie de doze de cocaină (Kosten și colab., 1994; Haile și colab., 2001). Pe baza acestui model de efecte, am presupus că șobolanii Lewis ne-ar permite să investigăm dacă susceptibilitatea la sensibilizare amplifică efectele programării condiționării asupra răspunsului ulterior la AMPH.
experimentul 3
Materiale și metode
Metodologia a fost aceeași ca și în experimentul 2, în afară de utilizarea șobolanilor Lewis (200-225 g la sosire, Charles River, Quebec, Canada).
REZULTATE
Nas în timpul sesiunilor de condiționare sucroză
O grupă 5 × Sesiune 15 × Faza 2 (CS prezent, CS absent) ANOVA a nasului a generat efecte principale semnificative ale grupului, F(4, 34) = 6.12, p = 0.001, sesiune, F(14, 476) = 3.42, p <0.001 și fază, F(1, 34) = 20.83, p <0.001, precum și o interacțiune semnificativă în trei direcții, F(56, 476) = 1.56, p = 0.008. panouri (A, B) din figura 7 trasați punctajele medii ale grupului cu vârfurile nasului pentru fazele CS prezente și respectiv CS absente. Comparația celor două panouri relevă faptul că efectul principal al fazei a reflectat mai multe răspunsuri generale la vârful nasului atunci când CS a fost prezent vs. absent. Prin urmare, răspunsurile indicate au apărut semnificativ mai des decât răspunsurile pre-mature. Efectele principale ale grupului și sesiunii nu au fost interpretate cu ușurință din cauza interacțiunii de ordin superior. Interacțiunea în trei direcții a reflectat o convergență a scorurilor pentru cele cinci grupuri la un nivel relativ stabil scăzut între sesiuni când CS a fost absent [Panoul (B)], împreună cu o divergență a scorurilor atunci când CS a fost prezentă în profiluri relativ discrete pentru fiecare grup care a paralel cu ordinea rangului lor de frecvență de recompensă: de la cea mai înaltă (grupa 100) la cea mai mică (grupul 25) [Panel (A)]. Numai tendința liniară a interacțiunii a fost semnificativă, F(4, 34) = 4.03, p = 0.009, reflectand cresterea general consistenta a naselor asupra sesiunilor in grupul 100 atunci cand CS a fost prezent in raport cu profilul relativ inconsistent de crestere a naselor in timpul sesiunilor in celelalte grupuri in aceasta faza.
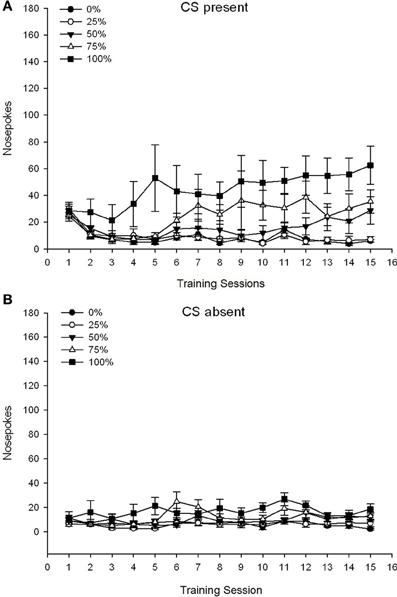
Figura 7. Reacțiile de abordare medie (SE) (șobolanii) asupra sesiunilor de antrenament pentru zahăr 15 în grupuri de șobolani Lewis (n = 8 / grup) expuse la recompensa de zaharoză (soluție 10%) livrată în cadrul programelor variabile 0, 25, 50, 75 sau 100%. Stimul condiționat a fost o lumină (120 s). Grupul 0 a primit același număr de recompense ca grupul 100 în absența stimulilor condiționați. (A) S-au înregistrat scoruri CS (studii 5 s × 20). (B) S-au scorat când CS a fost absent (media pentru 5 × 20 s în timp ce lumina a fost oprită).
Habitat pentru cutiile locomotorii
O sesiune 5 × 3 ANOVA a generat un efect principal al sesiunii, F(2, 70) = 23.07, p <0.0001 și fără alte efecte semnificative, F(8, 70) <1.47, p > 0.18. Un model curviliniu al scorurilor locomotorii medii (SE) a apărut din sesiunea 1, 1076 (74), până la sesiunea 2, 644 (48), până la sesiunea 3, 762 (59). Comparațiile planificate ale grupului 50 cu grupul 0 și cu grupul 100 la prima și ultima sesiune de obișnuință au relevat semnificativ mai puține pauze ale fasciculului în grupul 50 (M = 911; SE = 109) vs. grupul 0 (M = 1103; SE = 176) în sesiunea de obișnuință 1, t(105) = 2.02, p <0.05, dar nu există nicio diferență între grupul 50 și grupul 100 (M = 1066; SE = 150), t(105) <1.20, p > 0.20, la această sesiune. Grupa 50 nu a diferit semnificativ nici de grupul 0, nici de grupul 100 din ultima sesiune de obișnuință, t(105) <0.93, p > 0.30. Prin urmare, răspunsul locomotor mediu fără droguri în grupurile cheie nu a diferit în mod constant înainte de testare.
Sesiuni de testare
Ser fiziologic. O sesiune 5 × 2 ANOVA a răspunsurilor locomotorii la sesiunea de obișnuință de obișnuință și la sesiunea de testare salină a generat un efect principal semnificativ al sesiunii, F(1, 35) = 50.12, p <0.0001 și fără alte efecte semnificative, F(4, 35) <0.57, p > 0.68. Figura 8 arată scorurile medii ale grupului pentru cele două sesiuni și indică faptul că efectul Sesiunii a reflectat o scădere semnificativă de la obișnuirea la testul salin. Astfel, primirea injecției în sine (de exemplu, speranța, stresul) nu au îmbunătățit răspunsul locomotor.
Figura 8. Răspunsul locomotor mediu (SE) (numărul de rupturi ale fasciculului într-o gamă electronică pe 90 min) la ultima sesiune de habituire fără droguri 3 și la o sesiune ulterioară după injectarea salinei (ip, 1 ml / kg) în grupuri de șobolani Lewis (n = 8 / grup) expuse anterior la sesiunile de condiționare zilnică 15 cu recompensă pentru sucroză (soluție 10%) livrate în cadrul programelor variabile 0, 25, 50, 75 sau 100%. Stimul condiționat a fost o lumină (120 s). Grupul 0 a primit același număr de recompense ca grupul 100 în absența stimulilor condiționați.
Efectele 0.5 mg / kg AMPH.
Locomoție pre-injectabilă. O sesiune 5 × 2 ANOVA a locomoției pre-injectare a produs un efect principal semnificativ al sesiunii, F(1, 35) = 15.04, p <0.001 și fără alte efecte semnificative, F(4, 35) <1.19, p > 0.33. Comparațiile planificate nu au găsit nicio diferență semnificativă între grupul 50 și grupul 0 sau grupul 100 în ambele sesiuni de testare, t(70) <0.99, p > 0.30. Prin urmare, diferențele inițiale în locomoția pre-injecție nu au luat în considerare diferențele de grup în răspunsul locomotor la 0.5 mg / kg AMPH. Pauzele medii (SE) ale fasciculului pentru faza de pre-injecție pentru prima și a doua (post-sensibilizare) sesiuni de 0.5 mg / kg au fost 325 (25) și 473 (36).
Locomoția post-injectabilă. O sesiune 5 × 2 ANOVA a răspunsului locomotor la dozele 0.5 mg / kg livrate înainte și după administrarea 1 cronică mg / kg AMPH a generat un efect principal al sesiunii, F(1, 34) = 87.44, p <0.0001 și fără alte efecte semnificative, F(4, 34) <0.94, p > 0.45. Figura 9 descrie scorurile medii locomotorii pentru fiecare grup și sesiune și arată că efectul sesiunii a reflectat un răspuns global crescut la cea de-a doua doză 0.5 mg / kg, în concordanță cu sensibilizarea. Cifra arată, de asemenea, că grupurile s-au desfășurat foarte similar la sesiunea 1, dar grupul 50 a afișat activitate mai locomotorie decât celelalte grupuri din sesiunea 2. Comparațiile planificate ca răspuns la prima doză 0.5 mg / kg nu au evidențiat diferențe semnificative între grupul 50 și grupul 0 sau grupul 100, t(35) <1.28, p > 0.20. Cu toate acestea, grupul 50 a prezentat un răspuns locomotor semnificativ mai mare la a doua doză de 0.5 mg / kg decât grupul 0, t(35) = 4.32, p <0.001 sau grupul 100, t(35) = 2.24, p <0.05.
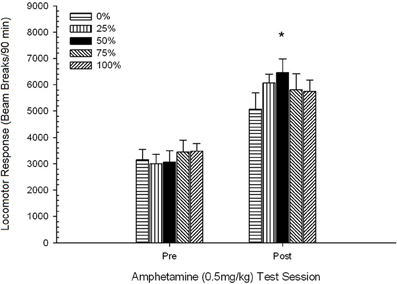
Figura 9. Raspunsul locomotor mediu (SE) (numarul de spargeri ale fasciculului intr-o matrice electronica pe 90 min) la 0.5 mg / kg d-amfetamina pe sesiuni separate inainte si dupa o sesiune de sesizare 5 a d-amfetaminei (1.0 mg / kg; per ședință) în grupuri de șobolani Lewis (n = 8 / grup) expuse anterior la sesiunile de condiționare zilnică 15 cu recompensă pentru sucroză (soluție 10%) livrate în cadrul programelor variabile 0, 25, 50, 75 sau 100%. Stimul condiționat a fost o lumină (120 s). Grupul 0 a primit același număr de recompense ca grupul 100 în absența stimulilor condiționați. *p <0.05 pentru diferența medie între grupul 50 și grupul 0, precum și grupul 100, pe baza comparațiilor planificate.
Efectele 1 mg / kg AMPH.
Locomoție pre-injectabilă. O sesiune 5 × 5 ANOVA a scorurilor pre-injectare 30-min pentru sesiunile de sensibilizare a generat un efect principal al sesiunii, F(4, 140) = 4.10, p = 0.004, și nici alte efecte semnificative, F(4, 35) = 1.25, p > 0.31. Comparațiile planificate au constatat că rupturile fasciculului în timpul fazei de pre-injecție (M; SE) au fost semnificativ mai mici în grupul 50 (395; 62) decât în grupul 100 (508; 62), t(175) = 2.58, p <0.01, dar nu grupul 0, t(175) <1.83, p > 0.10, la sesiunea de 1 mg / kg AMPH 1. La ultima sesiune de 1 mg / kg AMPH, comparațiile planificate au constatat, de asemenea, că locomoția pre-injecție în grupul 50 (378; 60) a fost semnificativ mai mică decât în grupul 100 (650; 75 ), t(175) = 6.17, p <0.001, dar nu în grupul 0, t(175) <1.84, p > 0.10. Deoarece direcția acestor diferențe de grup (grupul de control = grupul 50) a fost opusă tiparului ipotezat, diferențele de grup în locomoția post-injecție care se aliniază cu ipoteza nu pot fi atribuite diferențelor de bază înainte de injectare. Pauzele medii (SE) ale fasciculului în timpul fazei de pre-injecție pentru sesiunile 1 până la 5 au fost: 442 (34), 452 (32), 542 (40), 411 (26), 504 (37).
Locomoția post-injectabilă. O sesiune 5 × 5 ANOVA a răspunsurilor la dozele de 1 mg / kg a dat naștere unui efect principal semnificativ al sesiunii, F(4, 140) = 6.15, p <0.001 și fără alte efecte semnificative, F(4, 35) <0.57, p > 0.68. Analizele tendințelor polinomiale au relevat o tendință liniară semnificativă, F(1, 35) = 9.34, p = 0.004, și trendul cubic, F(1, 35) = 5.08, p = 0.031, ultimul rezultat denotând maximele relative la sesiunile 3 și 5. Figura 10 descrie aceste scoruri și arată că, în ciuda lipsei unei interacțiuni semnificative în ANOVA, grupul 50 a prezentat o locomoție substanțial mai mare decât celelalte patru grupuri ca răspuns la doza finală de 1 mg / kg. În consecință, comparațiile planificate au relevat scoruri medii semnificativ mai mari la sesiunea 5 în grupa 50 decât la toate celelalte grupuri, t(35) > 3.68, p <0.001.
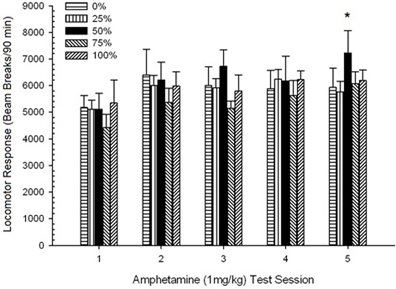
Figura 10. Răspunsul locomotor mediu (SE) (numărul de rupturi ale fasciculului într-o matrice electronică pe 90 min) la 1 mg / kg d-amfetamină (ip) pe sesiunile săptămânale 5 în grupuri de șobolani Lewisn = 8 / grup) expuse anterior la sesiunile de condiționare zilnică 15 cu recompensă pentru sucroză (soluție 10%) livrate în cadrul programelor variabile 0, 25, 50, 75 sau 100%. Stimul condiționat a fost o lumină (120 s). Grupul 0 a primit același număr de recompense ca grupul 100 în absența stimulilor condiționați. *p <0.05 pentru diferența medie între grupul 50 și grupul 0, precum și grupul 100, pe baza comparațiilor planificate.
Controlați variația nasului care răspunde în timpul antrenamentului cu zahăr
Două grupuri 5 x 2 ANCOVA ale răspunsului locomotor la 0.5 mg / kg AMPH înainte și după schema de sensibilizare, incluzând nivelele totale ale nasului în timpul antrenamentului cu zaharoză cu CS prezent și CS absent ca covariate separate, nu au găsit efecte semnificative nici pentru covariate, F(1, 32) <0.44 p > 0.51. Două ANCOVA-uri din sesiunea de 5 grupe × 5 de răspuns locomotor la 1 mg / kg AMPH în timpul sesiunilor de sensibilizare cu lovituri totale de nas (CS prezent, CS absent), deoarece covariate separate nu au produs efecte semnificative ale covariatei în timp ce CS era prezent sau absent, F(1, 33) <0.14, p > 0.71. Prin urmare, răspunsul la abordarea fără medicamente nu a luat în considerare diferențele de grup în răspunsurile locomotorii la nici o doză de AMPH.
Discuție
Sensibilizarea s-a dezvoltat la efectele repetate de 1.0 mg / kg de amfetamină. Datele privind obișnuința și soluția salină confirmă faptul că acest efect nu s-a datorat diferențelor preexistente, speranței sau răspunsurilor legate de stres la injecție. ANCOVA-urile cu nasuri confirmă faptul că aceste efecte nu s-au datorat comportamentului de abordare fără droguri. Datele despre nasul însuși au indicat că grupurile au dobândit asocierea dintre CS și perspectiva recompensei de zaharoză. Nivelul de răspuns al grupurilor de răspuns la sfârșitul antrenamentului s-a potrivit cu frecvența generală a recompensei în cadrul diferitelor programe de la cea mai mare (grupa 100) la cea mai mică (grupa 0), așa cum a făcut în experimentul 2. Media generală relativ mai mică nivelurile de nas în acest experiment, comparativ cu experimentele 1 și 2, pot reflecta o abordare mai selectivă, care răspunde la indicii pentru recompensă la șobolanii Lewis (Kosten și colab., 2007).
Datele privind doza 0.5 mg / kg au arătat că răspunsul locomotor inițial la AMPH la șobolanii Lewis (Figura 9) a fost oarecum suprimată comparativ cu șobolanii Sprague Dawley (experimentul 2; 5), dar creșterea în interiorul grupului ca răspuns la a doua doză la șobolanii Lewis a fost considerabilă (aproape dublul răspunsului la prima doză de 0.5 mg / kg) în urma tratamentului AMPH cu 5. Mai ales, grupul 50 a prezentat un răspuns locomotor mai mare decât toate grupurile, cu excepția grupului 25, pentru a doua doză (de exemplu, post-sensibilizare) 0.5 mg / kg AMPH și un răspuns locomotor mai mare decât toate celelalte grupuri, inclusiv grupul 25, până la doza finală de 1 mg / kg AMPH (sesiune de sensibilizare finală) .
Rezumat analiză a clasamentului de grup în cadrul experimentelor
Pentru a determina fiabilitatea diferențelor de grup în sensibilizare, o analiză neparametrică a evaluat contingența dintre grup și rangul răspunsului locomotor mediu la cea de-a doua (post AMPHPH) 0.5 mg / kg și doza finală 1.0 mg / kg de AMPH din experimentele 3. Analiza a dat un efect semnificativ, φ = 0.986, p = 0.025, reflectând faptul că grupul 50 sa clasat pe primul loc în toate, cu excepția uneia dintre comparații. Clasa superioară a grupului 50 comparativ cu celelalte grupuri ca răspuns la cea de-a doua (doză post-cronică AMPH) 0.5 mg / kg este prezentată în figura 5 (experiment 2) și Figura 9 (experiment 3). Clasamentul superior al grupului 50 față de alte grupuri ca răspuns la doza finală de 1.0 mg / kg este prezentat în figura 2 (experiment 1) și Figura 10 (experiment 3). Singura excepție de la acest model a fost răspunsul la doza finală de 1.0 mg / kg la șobolanii Sprague-Dawley în experimentul 2.
discutie generala
Actuala serie de experimente a testat ipoteza că expunerea cronică la un program de recompensă asemănător cu jocurile de noroc poate sensibiliza căile DA ale creierului, la fel ca expunerea cronică la medicamentele de abuz. Dovezile pentru un astfel de efect ar sugera că neuroplasticitatea, de același tip considerată a contribui la dependența de droguri, poate fi indusă de expunerea cronică la programele de recompensare imprevizibile. În conformitate cu literatura de specialitate privind dependența de droguri, răspunsul locomotor la dozele 0.5 și 1.0 mg / kg de reactivitate a sistemului DA DA indexat AMPH, cu locomoție mai mare ca răspuns la dozele ulterioare care definesc în mod operativ sensibilizarea (cf. Robinson și Berridge, 1993; Pierce și Kalivas, 1997; Vanderschuren și Kalivas, 2000).
În ansamblu, rezultatele sunt în concordanță cu ipoteza noastră. Cu toate acestea, ele indică, de asemenea, o variabilitate considerabilă a efectelor experimentale datorită factorilor de procedură. Efectele programării condiționării au fost modeste, dar consecvente, grupul 50 demonstrând un răspuns mai mare decât celelalte patru grupuri în ambele doze după regimul cu doze de cinci. Deși în general F- valorile pentru efectele legate de grup în analizele varianței au fost adesea nesemnificative, diferențele de grup cheie fiind confirmate prin comparații planificate pe perechi. În această privință, trebuie remarcat faptul că "Gândirea actuală este totuși o semnificație generală F în ANOVA] nu este necesar. În primul rând, ipotezele testate de testul global și de un test de comparație multiplă sunt destul de diferite, cu niveluri diferite de putere. De exemplu, în general F distribuie efectiv diferențele între grupuri în ceea ce privește numărul de grade de libertate pentru grupuri. Acest lucru are ca efect diluarea generală F în situația în care mai multe mijloace de grup sunt egale una cu alta, dar diferite de alte mijloace "(Howell, 1992, p. 338). Aceasta este tocmai situația aplicată în experimentele actuale, unde grupul 50 era de așteptat să difere de grupurile 0 și 100, dar nici o diferență între aceste grupuri de control nu a fost prevăzută pentru grupul 25 sau grupul 75.
Datele nasului au confirmat faptul că, în fiecare experiment, animalele au dobândit asocierea dintre CS și perspectiva recompensei de zaharoză. Corespondența dintre frecvența nasului pentru diferitele grupe și frecvența generală a recompenselor în cadrul programelor de formare respective sugerează că rata medie de răsplată a zahărului recompensează ghidul de abordare fără răspuns. Cu toate acestea, lipsa efectului de mediere a naselor asupra răspunsurilor locomotorii legate de grup la AMPH în ANCOVA a indicat faptul că procesele separate se află la baza celor două comportamente.
În unele cazuri, efectul programului de condiționare a fost evident ca răspuns la prima doză AMPH; în alte cazuri a apărut numai după doze repetate. Diferențele de grup în răspunsul locomotor la prima doză AMPH sugerează că expunerea la programe de recompensare asemănătoare jocurilor de noroc este suficientă pentru a induce sensibilizarea. Diferențele de grup în locomoție după doze multiple de AMPH indică un efect mai subtil care poate fi caracterizat drept "susceptibilitate", care se manifestă numai atunci când este combinată cu expunerea continuă la agentul sensibilizant primar (adică amfetamină).
Diferențele în modelul de răspuns în cadrul experimentelor sugerează că un interval mai lung între antrenament și provocarea inițială cu AMPH poate maximiza posibilitatea de a detecta efectul sensibilizant inerent al tratamentului condiționat. Aceasta, la rândul său, sugerează că efectele expunerii condiționate a expunerii pot fi incubate în timp, un fenomen văzut și cu sensibilizarea stimulentelor (Grimm și colab., 2006). Modelul răspunsului la cele două doze de amfetamină sugerează că doza de 0.5 mg / kg poate fi mai eficientă pentru a descoperi efectele istoricului condiționării. Aceasta, la rândul său, sugerează că efectele de condiționare în cadrul protocolului actual de antrenament sunt oarecum subtile și pot fi camuflate de efectele tavanului în doze de AMPH și de condițiile care generează novo sensibilizare.
În experimentul 3, modelul bifazic al răspunsului la dozele de 0.5 mg / kg și apariția progresivă a superiorității în grupul 50 este în concordanță cu profilul așteptat pentru șobolanii Lewis ca răspuns la metamfetamină (Camp și colab., 1994). Acest lucru susține validitatea constatărilor prezente și sugerează suprapunerea între factorii care atenuează vulnerabilitatea la sensibilizarea psihostimulantului și la programele de recompensare asemănătoare jocurilor de noroc.
În cadrul experimentelor, răspunsul locomotor post-sensibilizare al grupului 50 a depășit, în general, cel al altor grupuri în doze diferite de amfetamină și în diferite tulpini de animale. Cu toate acestea, variabilitatea ridicată în cadrul grupului și dimensiunile modeste ale efectului între grupuri indică un rol pentru alți factori în reactivitatea sistemului DA la amfetamină ca urmare a expunerii la diferitele scheme de recompensă sucroză condiționată. Deși răspunsurile neuronilor DA pentru recompensarea semnalelor pot oferi un model grosier de jocuri de noroc (Fiorillo și colab., 2003), la fel ca toate modelele, există o pierdere de informații din motive de parsimonie - de exemplu, pentru a demonstra un proces cheie. Ca rezultat, modelul de efecte în condițiile CS-SUA în originalul Fiorillo et al. studiul nu generalizează pe deplin răspunsul locomotor la amfetamină. Modificări suplimentare ale modelului sunt solicitate pentru a capta pe deplin aspectele legate de jocurile de noroc care au impact asupra funcției sistemului DA.
Luate împreună, rezultatele acestei serii de experimente oferă un sprijin provizoriu pentru ipoteza că expunerea cronică la programele de recompensă asemănătoare jocurilor de noroc sporește reactivitatea sistemului DA creier la provocarea psihostimulantului. Ca atare, ele extind constatările Singer și colab. (2012) care a demonstrat că, în raport cu un program fix, expunerea prealabilă la un program de armare variabilă într-o paradigmă operantă îmbunătățește răspunsul locomotor ulterior la amfetamină. Mai exact, concluziile prezente indică incertitudinea furnizării de recompense ca factor critic care stă la baza efectelor unei recompense variabile. Mărimea efectelor în paradigma operantă a fost substanțial mai mare decât efectele descoperite în experimentele de față. Acest lucru poate reflecta o expunere mai mare cronica la activitatea de tip joc (55 vs 15 zile); acesta poate reflecta efectele solicitării unui răspuns operator pentru a genera recompensa (adică un rol pentru agenție), mai degrabă decât expunerea pasivă, ca în studiul prezent. Creșterea duratei de formare în paradigma actuală ar ajuta la rezolvarea acestor întrebări.
Valabilitatea programelor variabile de recompensare și de întărire ca modele de jocuri de noroc nu poate fi obținută din aceste experimente. Cercetările viitoare care examinează impactul istoricului condiționării asupra comportamentului de asumare a riscurilor în sarcinile legate de jocuri de noroc pot explica această problemă. În mod similar, corespondența dintre sensibilizarea comportamentală găsită aici și răspunsul DA crescut striatal la amfetamina recent găsită în jocurile de noroc gameri trebuie să aștepte investigații suplimentare (Boileau și colab., 2013). Micro-dializa ar putea aborda această problemă, iar predicția bazată pe datele umane ar fi că eliberarea mai mare a DA în grupul 50 "fenotip de jocuri de noroc" ar fi cel mai clar observată în striatumul dorsal (senzorimotor) mai degrabă decât striatul ventral (limbic) . Validarea expunerii 50% variabile CS + în aceste alte paradigme ar susține utilitatea sa ca model bona fide experimental al PG.
În timp ce unele forme de jocuri de noroc implică în mod clar un răspuns instrumental (de exemplu, slot machine), în alte forme de jocuri de noroc (de exemplu, loterie) legătura dintre acțiune (cumpărarea biletului, adică plasarea pariului), indicii pentru recompensă (adică , numerele de loterie) și recompensa în sine (numărul câștigător și recompensa monetară) este mult mai difuză. Cu toate acestea, activarea DA în timpul intervalului CS-SUA poate să apară. Acest lucru poate explica de ce, atunci când este anunțat „numărul câștigător”, atenția este nituită pe măsură ce fiecare minge de loterie individuală cade succesiv pentru a compune secvența specifică de cifre din numărul câștigător. Deși probabilitatea ca o anumită cifră să apară este definită matematic, rezultatul pentru fiecare minge de loterie individuală este binar - lovit (se potrivește cu numărul jucătorului) sau ratat (nu se potrivește cu numărul jucătorului) - iar rezultatul pentru orice proces dat este necunoscut. Un astfel de scenariu poate caracteriza mai bine experiența grupului 50 din experimentele prezente, în care recompensa a fost furnizată în mod necontenit, dar și imprevizibil, iar CS a indicat doar potențialul recompensei fără a dezvălui dacă ar avea loc într-un proces dat. Sloturile sunt mai puternic legate de PG decât biletele de loterie (Cox și colab., 2000; Bakken și colab., 2009), indicând rolul important al factorilor (și al imediatei) factori în aspectele pline de satisfacție ale jocurilor de noroc pentru această populație (Loba și colab., 2001). Cu toate acestea, procesul Pavlovian modelat în experimentele actuale (recompensa nesigură CS +) pare a fi un element necesar, dacă nu suficient, al experienței jocurilor de noroc.
Odată cu lipsa unei cerințe instrumentale clare, o serie de alte caracteristici de proiectare ar fi putut contribui la modelul relativ modest și variabil al efectelor experimentale. Grupurile au fost diferite în ceea ce privește expunerea totală la zaharoză, precum și contingența dintre CS și recompensa de zaharoză. Deși acest lucru a contribuit la variabilitatea intergrupului, nu poate explica cu ușurință de ce animalele cu cea mai mare expunere la zaharoză (grupul 100) prezintă o sensibilizare mai redusă decât grupul 50. În plus, grupul 0 nu a primit niciun stimul înainte de expunerea la zaharoză la fiecare studiu. Deși acest lucru a împiedicat o așteptare indusă de recompensă, nu a controlat prezența unui stimul înainte de livrarea de recompense, care a existat în toate celelalte grupuri. Pentru a aborda această problemă, cercetarea viitoare ar trebui să includă o condiție în care animalele primesc recompensă în fiecare studiu după expunerea aleatorie la un stimul neutru (adică, a cărui prezență nu indică potențialul de recompensă).
O altă limitare a designului este apariția potențială a comportamentului adjuvant care ar putea influența efectele programului de antrenament. În fața incertitudinii, animalele pot dezvolta comportamente superstițioase menite să sporească controlul perceput și să reducă activarea DA indusă de incertitudine (cf. Harris și colab., 2013). Prin urmare, este posibil ca aspectele necontrolate ale designului experimental să permită animalelor să compenseze efectele programului de condiționare. Un astfel de efect ar putea contribui la răspunsul relativ modest și variabil la amfetamină din grupul 50, urmând formarea CS + sucroză. Cercetările viitoare ar trebui să înregistreze un comportament spontan, în afară de bătaia nasului, în timpul sesiunilor de formare pentru a testa această posibilitate și să controleze statistic dacă ar trebui să apară. Deoarece un astfel de comportament ar fi de așteptat să contracareze sau să atenueze efectele incertitudinii induse de program, răspunsul locomotor la amfetamină din grupul 50 ar trebui îmbunătățit când este controlat (procedural sau statistic). Prin urmare, proiectul prezent (necontrolat) oferă un test conservator al efectelor recompensării 50% CS + asupra sensibilizării amfetaminei.
În ceea ce privește valabilitatea externă, utilizarea șobolanilor masculi limitează și generalizarea rezultatelor. Lipsa unei condiții clare de "pedeapsă" diferă și de jocurile de noroc, unde pierderile monetare mari sunt comune și exercită importante efecte motivaționale (Nieuwenhuis și colab., 2005; Singh și Khan, 2012). Abilitatea de a acumula recompense este, de asemenea, absentă din paradigma actuală, iar câștigurile cumulate într-un joc de mașini automate au fost găsite a interacționa cu manipularea DA la om (Tremblay și colab., 2011; Smart și alții, 2013). În mod similar, oportunitatea pentru un jackpot este o diferență importantă între modelul prezent și jocurile actuale.
În ciuda acestor limitări, rezultatele prezente sugerează că recompensa 50% variabilă CS + poate angaja căi DA implicate în efectele de întărire ale jocurilor de noroc (Fiorillo și colab., 2003; Anselme, 2013). Sensibilizarea încrucișată a răspunsului la AMPH în urma acestui program de jocuri de noroc este în concordanță cu rolul central al DA în jocurile de noroc și în efectele psihostimulante ale medicamentelor (Zack și Poulos, 2009) și extinde studiile anterioare cu privire la amorsarea încrucișată a motivației de a paria de către AMPH la jucătorii patologici (Zack și Poulos, 2004). Rezultatele prezente sugerează, de asemenea indirect, că dozele modeste de AMPH, care nu provoacă eliberarea DA super-fiziologică, pot să modeleze mai bine activitatea creierului ca răspuns la semnalele de recompensă intermitentă (adică în timpul jocurilor de noroc) decât expunerea la un nivel ridicat (de exemplu, doze de medicamente stimulante (cf. Vanderschuren și Pierce, 2010). Sprijinul direct pentru această corespondență ar putea fi obținut prin evaluarea eliberării DA ca răspuns la schema CS-US variabilă 50% și doze diferite de AMPH utilizând microdializa.
Din punct de vedere experimental, modelul actual Pavlovian și modelul precedent de operare a armăturii variabile par să provoace un fenotip asemănător unui jucător patologic uman. Ca atare, ele oferă o completare valoroasă pentru sarcinile legate de jocurile de noroc care modelează comportamentul jocurilor de noroc (ca măsură dependentă), dar până acum au folosit numai animale sănătoase, echivalentul jucătorilor sociali umani. Bazându-se pe literatura de specialitate, animalele expuse în mod cronic la o recompensă variabilă pot diferi foarte bine în aceste sarcini, în special ca răspuns la medicamentele DA-ergice. Combinarea fenotipului de jocuri de noroc cu șobolani cu sarcinile de jocuri de noroc poate permite dezvoltarea sistematică a medicamentelor pentru tratamentul PG, care s-ar putea să nu fie realizată pe deplin numai cu animale sănătoase. Îmbunătățirile suplimentare în schema de experimentare și de formare experimentală, descrise mai sus, ar trebui să îmbunătățească corespondența dintre animalele instruite în această paradigmă și jucătorii patologi reali.
Din punct de vedere clinico-sociologic, constatarea că expunerea la recompensa 50% variabilă CS +, care se potrivește strâns programului de recompensare pe un slot machine comercial (Tremblay și colab., 2011), modifică sistemul DA creier în moduri fiabile și durabile sugerează că, în unele cazuri, activitatea de jocuri de noroc, cum ar fi drogurile de abuz, poate fi un "agent patogen" capabil să cauzeze dependență. Cu toate acestea, dimensiunea modestă a efectului și variabilitatea ridicată ca răspuns la recompensa 50% CS + sugerează că, ca și drogurile de abuz, tendința de programare a recompensărilor de tip joc de noroc pentru a promova dependența va depinde în mare măsură de profilul de risc pre-existent al jucătorului. Cu toate acestea, pentru a evita expunerea persoanelor cu risc sporit la potențialele efecte adverse legate de jocurile de noroc, pare rezonabil ca politicile aplicate pentru a descuraja utilizarea și a reduce la minimum efectele nocive ale drogurilor de abuz ar putea fi extinse și la jocurile de noroc.
Declarația privind conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
recunoasteri
Această cercetare a fost finanțată prin granturi de la Consiliul Științelor Naturale și Inginerie Cercetare din Canada către Paul J. Fletcher. Îi mulțumim sincer domnului Djurdja Djordjevic pentru pregătirea cifrelor.
Referinte
Anselme, P. (2013). Dopamina, motivația și semnificația evolutivă a comportamentului asemănător cu jocurile de noroc. Behav. Brain Res. 256, 1-4. doi: 10.1016 / j.bbr.2013.07.039
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
APA (2013). Manualul Diagnostic și Statistic al Tulburărilor Mentale, 5th Edn. Arlington, VA: American Psychiatric Publishing.
Bakken, IJ, Gotestam, KG, Grawe, RW, Wenzel, HG și Oren, A. (2009). Jocurile de noroc și problemele de jocuri de noroc în Norvegia 2007. Scand. J. Psychol. 50, 333-339. doi: 10.1111 / j.1467-9450.2009.00713.x
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Boileau, I., Payer, D., Chugani, B., Lobo, DS, Houle, S., Wilson, AA și colab. (2013). In vivo dovezi privind o mai mare eliberare a dopaminei induse de amfetamină în jocurile de noroc patologice: un studiu cu tomografie cu emisie de pozitroni cu [C] - (+) - PHNO. Mol. Psihiatrie doi: 10.1038 / mp.2013.163. [Epub înainte de imprimare].
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Bolles, RC (1972). Consolidarea, speranța și învățarea. Psychol. Rev. 79, 394-409. doi: 10.1037 / h0033120
Camp, DM, Browman, KE și Robinson, TE (1994). Efectele metamfetaminei și cocainei asupra comportamentului motor și a dopaminei extracelulare în striatumul ventral al șobolanilor Lewis versus Fischer 344. Brain Res. 668, 180–193. doi: 10.1016/0006-8993(94)90523-1
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Conversano, C., Marazziti, D., Carmassi, C., Baldini, S., Barnabei, G. și Dell'Osso, L. (2012). Jocuri de noroc patologice: o revizuire sistematică a descoperirilor biochimice, neuroimagistice și neuropsihologice. Harv. Rev. Psihiatrie 20, 130-148. doi: 10.3109 / 10673229.2012.694318
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Cox, BJ, Kwong, J., Michaud, V. și Enns, MW (2000). Problema și probabilitatea jocurilor de noroc patologice: considerații dintr-un sondaj comunitar. Poate sa. J. Psychiatry 45, 548-553.
Dalley, JW, Theobald, DE, Eagle, DM, Passetti, F. și Robbins, TW (2002). Deficitele în controlul impulsurilor asociate cu funcția serotoninergică tonic-crescută în cortexul prefrontal al șobolanului. Neuropsychopharmacology 26, 716–728. doi: 10.1016/S0893-133X(01)00412-2
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Everitt, BJ și Robbins, TW (2005). Sisteme neurale de întărire a dependenței de droguri: de la acțiuni la obiceiuri la constrângere. Nat. Neurosci. 8, 1481-1489. doi: 10.1038 / nn1579
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Fiorillo, CD, Tobler, PN și Schultz, W. (2003). Codificarea discretă a probabilității de recompensă și a incertitudinii prin neuroni dopaminergici. Ştiinţă 299, 1898-1902. doi: 10.1126 / science.1077349
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Flagel, SB, Watson, SJ, Akil, H. și Robinson, TE (2008). Diferențe individuale în atribuirea caracterului stimulativ către un tact de recompensă: influența asupra sensibilizării cocainei. Behav. Brain Res. 186, 48-56. doi: 10.1016 / j.bbr.2007.07.022
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Fletcher, PJ, Tenn, CC, Rizos, Z., Lovic, V. și Kapur, S. (2005). Sensibilizarea la amfetamină, dar nu la PCP, afectează schimbarea setată atentă: inversarea de către un agonist al receptorului D1 injectat în cortexul prefrontal medial. Psihofarmacologie (Berl.) 183, 190–200. doi: 10.1007/s00213-005-0157-6
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Flores, G., Wood, GK, Barbeau, D., Quirion, R. și Srivastava, LK (1998). Lewis și Fischer șobolani: o comparație a nivelurilor de transportator de dopamină și de receptori. Brain Res. 814, 34–40. doi: 10.1016/S0006-8993(98)01011-7
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Frascella, J., Potenza, MN, Brown, LL și Childress, AR (2010). Vineabilitățile comune ale creierului deschid calea pentru dependența de substanțe non-dependente: sculptură de dependență la o nouă comună? Ann. NY Acad. Sci. 1187, 294-315. doi: 10.1111 / j.1749-6632.2009.05420.x
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Grimm, JW, Buse, C., Manaois, M., Osincup, D., Fyall, A., și Wells, B. (2006). Disocierea dependentă de timp a efectelor doză-răspuns ale cocainei asupra poftei și locomoției cu sucroză. Behav. Pharmacol. 17, 143-149. doi: 10.1097 / 01.fbp.0000190686.23103.f8
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Haile, CN, Hiroi, N., Nestler, EJ și Kosten, TA (2001). Răspunsurile comportamentale diferențiate la cocaină sunt asociate cu dinamica proteinelor mezolimbice de dopamină la șobolanii Lewis și Fischer 344. Synapse 41, 179-190. doi: 10.1002 / syn.1073
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Harris, JA, Andrew, BJ și Kwok, DW (2013). Abordarea prin reviste în timpul unui semnal pentru alimente depinde de condiționarea Pavlovian, nu instrumentală. J. Exp. Psychol. Anim. Behav. Proces. 39, 107-116. doi: 10.1037 / a0031315
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Howell, DC (1992). Metode statistice pentru psihologie. Boston, MA: Duxbury.
Ito, R., Dalley, JW, Robbins, TW și Everitt, BJ (2002). Eliberarea dopaminei în striatul dorsal în timpul comportamentului căutător de cocaină sub controlul tacului asociat consumului de droguri. J. Neurosci. 22, 6247-6253.
Jiao, X., Pare, WP și Tejani-Butt, S. (2003). Diferențele de diferențiere în distribuția situsurilor transportatoare de dopamină în creierul de șobolan. Prog. Neuropsychopharmacol. Biol. Psihiatrie 27, 913–919. doi: 10.1016/S0278-5846(03)00150-7
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Koob, GF și Le Moal, M. (2008). Revizuire. Mecanisme neurobiologice pentru procese motivaționale opuse în dependență. Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 3113-3123. doi: 10.1098 / rstb.2008.0094
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Kosten, TA, Miserendino, MJ, Chi, S. și Nestler, EJ (1994). Tulpinile de șobolani Fischer și Lewis prezintă efecte diferențiale de cocaină în preferința locului condiționat și sensibilizarea comportamentală, dar nu în activitatea locomotorie sau aversiunea condiționată a gustului. J. Pharmacol. Exp. Ther. 269, 137-144.
Kosten, TA, Zhang, XY și Haile, CN (2007). Diferențele de diferențiere în menținerea autoadministrării de cocaină și relația lor cu răspunsurile la activitatea de noutate. Behav. Neurosci. 121, 380-388. doi: 10.1037 / 0735-7044.121.2.380
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Leeman, RF și Potenza, MN (2012). Asemănări și diferențe între tulburările patologice și jocurile de noroc: o concentrare asupra impulsivității și compulsivității. Psihofarmacologie (Berl.) 219, 469–490. doi: 10.1007/s00213-011-2550-7
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Leshner, AI (1997). Dependența este o boală a creierului și contează. Ştiinţă 278, 45-47. doi: 10.1126 / science.278.5335.45
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Loba, P., Stewart, SH, Klein, RM și Blackburn, JR (2001). Manipularea caracteristicilor jocurilor terminale standard de loterie video (VLT): efecte asupra jucătorilor patologici și non-patologi. J. Gambl. știft. 17, 297-320. doi: 10.1023 / A: 1013639729908
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Martinez, D., Narendran, R., Foltin, RW, Slifstein, M., Hwang, DR, Broft, A., și colab. (2007). Eliberarea de dopamină indusă de amfetamină: puternic afectată de dependența de cocaină și predicția alegerii de autoadministrare a cocainei. A.m. J. Psychiatry 164, 622-629. doi: 10.1176 / appi.ajp.164.4.622
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Mateo, Y., Lipsa, CM, Morgan, D., Roberts, DC, și Jones, SR (2005). Reducerea funcției terminale a dopaminei și insensibilitatea față de cocaină după administrarea de cocaină și deprivarea. Neuropsychopharmacology 30, 1455-1463. doi: 10.1038 / sj.npp.1300687
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Nestler, EJ (2001). Baza moleculară a plasticității pe termen lung care stă la baza dependenței. Nat. Rev. Neurosci. 2, 119-128. doi: 10.1038 / 35053570
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Nieuwenhuis, S., Heslenfeld, DJ, von Geusau, NJ, Mars, RB, Holroyd, CB și Yeung, N. (2005). Activitatea în zonele creierului sensibil la răsplată umană este puternic dependentă de context. Neuroimage 25, 1302-1309. doi: 10.1016 / j.neuroimage.2004.12.043
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Pierce, RC și Kalivas, PW (1997). Un model de expresie a sensibilizării comportamentale la psihostimulatori de tip amfetaminic. Brain Res. Brain Res. Rev. 25, 192. doi: 10.1016 / S0165-0173 (97) 00021-0
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Robbins, TW și Everitt, BJ (1999). Dependența de droguri: obiceiurile proaste se adaugă. Natură 398, 567-570. doi: 10.1038 / 19208
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Robinson, TE, Becker, JB și Presty, SK (1982). Facilitarea pe termen lung a comportamentului rotativ indus de amfetamină și a eliberării dopaminei striate produse de o singură expunere la amfetamină: diferențe de sex. Brain Res. 253, 231–241. doi: 10.1016/0006-8993(82)90690-4
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Robinson, TE și Berridge, KC (1993). Baza neurală a poftei de droguri: o teorie de stimulare-sensibilizare a dependenței. Brain Res. Brain Res. Rev. 18, 247-291. doi: 10.1016 / 0165-0173 (93) 90013-P
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Robinson, TE și Berridge, KC (2001). Stimularea sensibilizării și dependența. Dependenta 96, 103-114. doi: 10.1046 / j.1360-0443.2001.9611038.x
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Singer, BF, Scott-Railton, J. și Vezina, P. (2012). Protecția impresionantă a zaharinei sporește reacția locomotorie la răspunsul la amfetamină. Behav. Brain Res. 226, 340-344. doi: 10.1016 / j.bbr.2011.09.003
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Singh, V. și Khan, A. (2012). Luarea deciziilor în variantele de recompensare și pedeapsă ale sarcinii de jocuri de noroc iowa: dovezi de "previziune" sau "încadrare"? Față. Neurosci. 6: 107. doi: 10.3389 / fnins.2012.00107
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Skinner, BF (1953). Știință și comportament uman. New York, NY: presă liberă.
Smart, K., Desmond, RC, Poulos, CX și Zack, M. (2013). Modafinil mărește rolul de recompensă într-un joc de mașini de slot în jucători patologici cu nivel scăzut și ridicat de impulsivitate. Neuropharmacology 73, 66-74. doi: 10.1016 / j.neuropharm.2013.05.015
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Tremblay, AM, Desmond, RC, Poulos, CX și Zack, M. (2011). Haloperidolul modifică aspectele instrumentale legate de jocurile de noroc la jucători patologici și controale sănătoase. Addict. Biol. 16, 467-484. doi: 10.1111 / j.1369-1600.2010.00208.x
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Vanderschuren, LJ și Kalivas, PW (2000). Alterarea transmiterii dopaminergice și glutamatergice în inducerea și exprimarea sensibilizării comportamentale: o analiză critică a studiilor preclinice. Psihofarmacologie (Berl.) 151, 99-120. doi: 10.1007 / s002130000493
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Vanderschuren, LJ și Pierce, RC (2010). Procesele de sensibilizare în dependența de droguri. Curr. Top. Behav. Neurosci. 3, 179-195. doi: 10.1007 / 7854_2009_21
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Vanderschuren, LJ, Schoffelmeer, AN, Mulder, AH și De Vries, TJ (1999). Mecanisme dopaminergice care mediază exprimarea pe termen lung a sensibilizării locomotorii după pre-expunerea la morfină sau amfetamină. Psihofarmacologie (Berl.) 143, 244-253. doi: 10.1007 / s002130050943
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Verdejo-Garcia, A., Lawrence, AJ și Clark, L. (2008). Impulsivitatea ca marker de vulnerabilitate pentru tulburările de utilizare a substanțelor: revizuirea constatărilor rezultate din cercetările cu grad ridicat de risc, gamblerele problematice și studiile de asociere genetică. Neurosci. Biobehav. Rev. 32, 777-810. doi: 10.1016 / j.neubiorev.2007.11.003
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Vezina, P. (2004). Senzibilizarea reactivității neuronului dopaminei midbrain și autoadministrarea medicamentelor stimulente psihomotorii. Neurosci. Biobehav. Rev. 27, 827-839. doi: 10.1016 / j.neubiorev.2003.11.001
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Volkow, ND, Wang, GJ, Fowler, JS, Logan, J., Gatley, SJ, Hitzemann, R. și colab. (1997). S-a redus receptivitatea dopaminergică striatală la subiecții dependenți de cocaină. Natură 386, 830-833. doi: 10.1038 / 386830a0
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Volkow, ND, Wang, GJ, Fowler, JS și Tomasi, D. (2012). Circuite de dependență în creierul uman. Annu. Rev. Pharmacol. Toxicol. 52, 321-336. doi: 10.1146 / anurev-pharmtox-010611-134625
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Winer, B. (ed.). (1971). Principiile statistice în designul experimental. New York, NY: McGraw-Hill.
Zack, M. și Poulos, CX (2004). Amfetamina primește motivația de a juca jocuri de noroc legate de jocuri semantice legate de jocurile de noroc. Neuropsychopharmacology 29, 195-207. doi: 10.1038 / sj.npp.1300333
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Zack, M. și Poulos, CX (2009). Rolul paralel al dopaminei în jocurile de noroc patologice și al dependenței psihostimulante. Curr. Abuzul de droguri Rev. 2, 11-25. doi: 10.2174 / 1874473710902010011
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Zamudio, S., Fregoso, T., Miranda, A., De La Cruz, F. și Flores, G. (2005). Diferențele de diferențe ale nivelurilor receptorilor de dopamină și comportamentele legate de dopamină la șobolani. Brain Res. Taur. 65, 339-347. doi: 10.1016 / j.brainresbull.2005.01.009
Pubmed Abstract | A publicat un text complet | CrossRef Full Text
Cuvinte cheie: jocuri de noroc patologice, sensibilizare, amfetamină, dopamină, incertitudine
Referirea: Zack M, Featherstone RE, Mathewson S și Fletcher PJ (2014) Expunerea cronică la un program asemănător cu jocurile de noroc de stimuli predictivi poate recompensa sensibilizarea amfetaminei la șobolani. Față. Behav. Neurosci. 8: 36. doi: 10.3389 / fnbeh.2014.00036
Primit: 01 Noiembrie 2013; Acceptat: 23 ianuarie 2014;
Publicat online: 11 Februarie 2014.
Editat de:
Bryan F. Singer, Universitatea din Michigan, Statele Unite ale Americii
Revizuite de:
Louk Vanderschuren, Universitatea din Utrecht, Olanda
Ruud Van Den Bos, Universitatea Radboud Nijmegen, Olanda
Patrick Anselme, Universitatea din Liège, Belgia
Copyright © 2014 Zack, Featherstone, Mathewson și Fletcher. Acesta este un articol cu acces deschis, distribuit în termenii lui Creative Commons Attribution License (CC BY). Utilizarea, distribuirea sau reproducerea în alte forumuri este permisă, cu condiția ca autorii sau licențiatorii originali să fie creditați și să fie citată publicația originală din acest jurnal, în conformitate cu practica academică acceptată. Nu este permisă utilizarea, distribuirea sau reproducerea care nu respectă acești termeni.
* Corespondență: Martin Zack, Laboratorul de psihofarmacologie cognitivă, Departamentul de Neuroștiințe, Centrul de dependență și sănătate mintală, 33 Russell Street, Toronto, ON M5S 2S1, Canada e-mail: [e-mail protejat]
