9月30、2013・バイ タリアラーナー
ドーパミンニューロンは、最も研究されている、最も感覚化されているニューロンです。 最近では、しかし、彼らはアイデンティティ危機のビットを経てきました。 ドーパミンニューロンとは何ですか? ドーパミン研究における最近のいくつかの興味深いねじれは、ドーパミンニューロンはすべて種類のものであるという神話を決定的に非難しました - そして、あなたはそれらをそのように扱うどんな研究にでも疑問を呈するべきです。
ドーパミンニューロン(神経伝達物質ドーパミンを放出するニューロンとして定義される)が単なるドーパミンニューロンではない多くの方法があります。 ここでは、3つの本当に素晴らしい方法に焦点を当てます。
- ドーパミンを放出するすべてのニューロンが同じ場所に同時に放出するわけではありません。 ドーパミンニューロンは、神経回路にどのようにつながっているかによって、脳の機能や行動にさまざまな役割を果たします。
- ドーパミンを放出するすべてのニューロンがドーパミンだけを放出するわけではありません。 他の神経伝達物質を放出する人もいますが、それが神経回路機能にどのように影響するかに大きな影響を与える可能性があります。
- ドーパミンを放出するすべてのニューロンが常にドーパミンを放出するわけではありません。 一部のニューロンは、ドーパミン合成機構をオンまたはオフに切り替えることができます。 この能力のために、それらはドーパミンニューロンであると以前の研究でさえ認識されていなかったかもしれません。
私がこれらのエキサイティングな新しい発見を説明する前に、しかし、私はあなたにドーパミンニューロンへの標準的なNeuroscience 101紹介をさせてください。 ドーパミンニューロン機能のこの影響力のある理論は、Wolfram Schultzとその同僚の1997 Science paperから得られたものです。予測と報酬の神経基盤。」 それは、あるバックグラウンドレートで発火するドーパミンニューロンが、予測されていないが予測されていない報酬に応じてより発火することを示した。 さらに、あなたが報酬を期待していて、それを得ないならば、ドーパミンニューロンはより少なく発射します。 この発見はSchultz等を導きました。 つまり、ドーパミンニューロンが「報酬予測エラー」をコード化することを提案するのです。つまり、物事があなたの予想よりも良い、良い、あるいは悪いかどうかを教えてくれます。 シュルツ等。 これらのニューロンの反応は比較的均質であり、異なるニューロンは同じように反応し、異なる食欲刺激は類似のニューロン反応を誘発する。 すべての反応は、大部分のドーパミンニューロン(55〜80%)で発生します。」
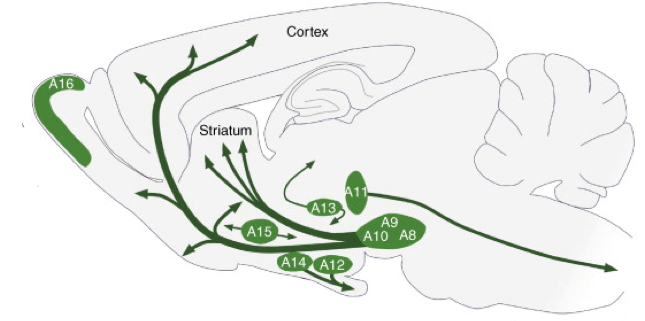
報酬予測誤差の計算機としてのドーパミンニューロンの役割は魅力的で価値のある研究のままであるが、もし報酬予測誤差がドーパミンニューロンがするすべてであれば、それらのために400,000-600,000が何を必要とするのか。脳のドーパミンニューロンは(げっ歯類の脳の断面に)位置しています。
成体げっ歯類脳におけるドーパミンニューロン細胞群A8 ‐ A16の分布 脳内のSBドーパミンニューロンシステムであるBjörklund、A。&Dunnettから改作:最新情報。 Neurosciences 30、194–202(2007)の傾向。
*人間の中で。 サルには160,000-320,000、げっ歯類には20,000-45,000があります。
この図を見ると、ドーパミンニューロンのグループ間にいくつかの大きな解剖学的な違いがすでにあるように見えます。それが、それらがA8-A16とラベル付けされている理由です。 解剖学的な区別も細かいので、機能的な意味合いはそれほど微妙ではありません。 研究の最初の行で、ここで焦点を当てます、Lammel等。 腹側被蓋野(上記の写真ではVTA、またはA10)のドーパミンニューロンを他の脳の領域との結合性によって区別することについて説明します。 ランメル等。 VTA内にドーパミンニューロンの少なくとも2つの分離可能な集団があることを観察した。 1人の母集団は、後外側部と呼ばれる脳領域から入力信号を受け取り、側坐核と呼ばれる脳領域に出力信号を送ります(これらのLDT-ドーパミン-NAcニューロンと呼びます)。 他の集団は外側手綱から入力を得て、前頭前野に出力を送る(これらのLHb-ドーパミン-PFCニューロンと呼ぶ)。 だから何? これらのドーパミンニューロンが異なる脳回路に配線されているという事実は、行動にとって全く重要なのでしょうか。 ランメル等。 それが問題であることを示した。 とき オプトジェネティクス彼らはマウスにおいてLDT-ドーパミン-NAcニューロンへの入力を活性化し、彼らは動物がそれらが刺激された文脈と正の関連を形成することを見出した。 彼らは脳の刺激を受けたボックスの部分でもっと時間を過ごすことを選びました。 対照的に、Lammel等。 LHb−ドーパミン−PFCニューロンへの入力を活性化したところ、正反対が観察された。 動物は刺激を受けた箱の一部を避けた。 別の研究 同じグループによると、マウスが何か良いことや悪いことを自然に経験したとき、これらの異なる回路の強さは異なって変調されました。 コカインを投与されたマウスは、LDT-ドーパミン-NAc経路の強度の増加を示したが、LHb-ドーパミン-PFC経路には変化を示さなかった。 足に刺激を与えたマウスは、LDT-ドーパミン-NAc経路に変化は見られなかったが、LHb-ドーパミン-PFC経路の強度は増加した。
ドーパミンニューロンが同質であるというSchultzらの最初の主張を弱体化させる、Lammelら。 彼らがそうでないことを発見しました。 この改訂は、利用可能なツールの感度が増加したために発生した可能性があり、これは1990から2010にかなり変更されました。 少し優れた創造性と相まって、より新しくより良いツールがLammel et al。を可能にしました。 Schultz等にアクセスできない微妙な点を区別するため。 これらの微妙な点を明らかにすることで、Lammel等。 特に母集団の55-80%の回答があるから、それはあなたがニューロンの全クラスを考え出したと信じることの根拠を示すのに役立ちました。その人口を定義するために使用されています。 (インビボでの神経記録の間にドーパミンニューロンを定義するという問題は、他の全ての問題である)。 Schultz et al。が世界中のすべての称賛を受けています。 ドーパミン研究の火をつけるための、しかしそれは終点よりもむしろ出発点であった。
あなたが脳回路がどのように機能するのか理解しようとしているなら、彼らが参加している脳回路によってニューロンをグループ化することは意味のあるトンになります。 しかし、ドーパミンニューロンのドーパミン部分を見つけ出そうとしているとしたら? ほとんどのドーパミンニューロン研究は、ドーパミンニューロンが発火すると、神経伝達物質ドーパミン、このように見える小さな分子を放出すると仮定しました:
実際、それが「ドーパミンニューロン」を定義した方法です。しかし、科学ではよくあることですが、状況はそれほど単純ではないことがわかりました。 私がここで議論する最近の研究の2行目で、科学者たちはドーパミンニューロンがドーパミンと共にグルタミン酸塩とGABAと呼ばれる他の神経伝達物質分子を共放出できるという証拠を示しました。
実際には、ドーパミンニューロンの異なるサブセットは、グルタメートまたはGABAのどちらかを優先的に共放出する可能性が最も高いです。 による研究 Hnasko等。 (NAIST) および Stuberら。 VTA中のドーパミンニューロンがグルタミン酸を共放出することを実証した。 最初に、彼らは多くのVTAドーパミンニューロンがVGLUTX NUMXと呼ばれるグルタミン酸トランスポーターを発現することに気付いた。 VGLUT2の存在は、ドーパミンニューロンがドーパミンに加えてグルタミン酸をパッケージングしていたことを意味しましたか? この問題を見るために、科学者たちはドーパミンニューロン刺激に対する側坐核(ドーパミンニューロンが出力を送る場所)のニューロンの反応を調べました。 実際、彼らは、ドーパミン作動性反応よりもむしろグルタミン酸作動性反応と一致するであろうタイプのVTAドーパミンニューロンの刺激に対する側坐核ニューロンの迅速で興奮性の反応を観察した。 これらの反応は、グルタミン酸受容体の拮抗薬によって遮断されたが、ドーパミン受容体の拮抗薬によっては遮断されなかった。 さらに、ドーパミンニューロンにおいてVGLUTX NUM Xを欠くように遺伝子操作されたマウスでは、そのような応答は見られなかった。
しかし、グルタミン酸塩の共放出は、すべてのドーパミンニューロンで起こるわけではありません。 Lammelらの研究と同様に、接続性も重要です。 Stuberら。 背側線条体に出力を送る黒質(A9)と呼ばれる隣接領域のドーパミンニューロンは、グルタミン酸放出の証拠を示さなかったことを観察しました。 その否定的な結果はまだ物議をかもしている。 他のグループ Tritsch等。黒質ドーパミンニューロンによるグルタミン酸塩放出のいくつかの証拠を観察した。 さらに、彼らは、これらの黒質ドーパミンニューロンもまた別の神経伝達物質であるGABAを同時放出することを実証した。 しかし、奇妙なことに、黒質ドーパミンニューロンは通常のGABAトランスポーターであるVGATを発現しない。 代わりに、Tritsch等。 ドーパミントランスポーターであるVMATもGABAを共輸送することができ、ドーパミンと共にシナプス放出のためにそれを包装することを発見した。 Tritschらの知見は、黒質ドーパミンニューロンを超えて一般化するかもしれない。 いくつかのGABAがある限り、VMATを表現するものは潜在的にそのGABAも同様にパッケージ化およびリリースすることができます。 Tritschらの研究から生じる一つの重要な問題は、黒質中のGABAが正確にどこでいつ合成されるのかということである。 それにもかかわらず、それはそこにあります。
大部分のドーパミンニューロンからのグルタミン酸塩とGABAの同時放出の影響は、まだ見られていません。 報告されている唯一の行動への影響はHnaskoらによるものです。 紙。 彼らは、ドーパミンニューロンにおいてVGLUTX NUMXを欠くマウスは、正常マウスよりコカインに反応して走り回ることが少ないことを示している。 それは今のところそれです。 他に何もないならば、それは我々が送信機の同時解放の現象についてどれだけ多く学ぶべきであるかを示しています。
これまでのところ、ドーパミンニューロンは、異なる脳回路に接続されている場合は異なることを知らせることができ、少なくとも一部は単にドパミン以外の化学物質を使用して脳回路に割り当てられた役割を果たすことができます。 ここで検討する3行目の研究では、さらに複雑で複雑な絵の層をもう1つ追加します。ドーパミンニューロンは、自分たちが作っているかどうかを変えることによって脳回路への参加方法を変えることができます。全くドーパミンを放出する。 この場合、 Dulcis et al。 視床下部にある、これまで私が今まで話してきたものとは少し違うドーパミンニューロンのグループを見ました。 彼らは、ラットにおけるドーパミンニューロンの数が、ラットが経験する「日光」の長さと共に変動するように思われることに気付いた。 それは本物の昼光ではないので、私は引用符で昼光を入れました - まさしく制御された実験室環境でライトがついているかどうか。 ほとんどの実験動物は1日当たり12時間の光を見ます、しかしDulcis等。 1日に5時間、または19まで試してみました。 長い日数を経験したラットはそれらの視床下部におけるドーパミンニューロンが少なかったが、短い日数を経験したラットはそれよりも多かった。 さらに調べると、彼らは、異なる光条件におけるドーパミンニューロンの数の変化は、ニューロンが死んで生まれたためではないと判断しました。 同じニューロンが実際にすべての条件でそこにいました、しかし、彼らは彼らのドーパミン機能をオンまたはオフに切り替えていました。 なぜ露光量がこれらの変化を引き起こすのか、または正確な行動への影響が何であるのかは依然として不明である。 長い日数を持ち、その結果としてドーパミンニューロンが少なくなったラットは、憂鬱で不安な行動を示しました(ラットは夜行性で暗闇が好きであることに注意してください)。 視床下部ドーパミンニューロンが毒素で殺されたラットもそうでした。 しかしながら、ラットが1日当たりXNUMX時間の光を受けている間にドーパミンニューロンが毒素で殺され、その後ラットが1日当たりXNUMX時間の光しか与えられなかった場合、以前は非ドーパミン作動性ニューロンがドーパミンを放出するために動員された。行動が観察されました。 かなりクール! そして重要なことに、この研究は、ドーパミンニューロンとして以前に同定されたこともなかったニューロンが正しい条件下で変換できることを示しています。 私たちの脳のいくつかの側面は安定しているように構築されていますが、その多くは常に変化しているため、私たちは内在化し、私たちの経験に適応することができます。
これらすべての研究の後、私たちは何を学びましたか? 私にとって、全体像のテイクアウトは、脳を理解することは複雑さを理解することを意味するということです。 もう少し具体的に言うと、それは分子や細胞を回路や行動と結び付けて研究のモダリティにまたがる生物学的実体の定義を提供することを意味します。 彼らが解放することができる1つの神経伝達物質によって単独でこれ以上の神経細胞を分類することはできません。 そのグループ化はまだ関連性がある場合がありますが、上記の調査で見たように、必ずしもそうとは限りません。 正式にドーパミンニューロンとして知られているグループを再定義することを考えて、私達はまた後知恵が提供する見方で過去の文献の何十年も振り返る必要があります。 それは、より古いドーパミンニューロン研究のデータが間違っているということではありませんが、結論は我々が彼らが思っていたものと全く同じではないかもしれません。 これは良いことかもしれません。 ドーパミンニューロンが正確に何をコード化するかについての多くの難解な議論は、実際にはそれらが異なる文脈で多くの異なることをすることを理解することによって解決されるかもしれません。 心配しないでください。混乱しているように思われるかもしれませんが、これは科学が成熟するごく普通のプロセスです。 プロセスが正常であるだけでなく、それは絶対に重要です。 科学者は重要な概念上の進歩を反映するために絶えず私達の定義を疑問視しそして修正しなければならない。
定義は混乱を招く可能性があります。 彼らはまたかなり退屈になる可能性があります、そして私は彼らがあまりにもしばしば科学から人々を追い払うことを心配します。 私が最初の生物学の学生だったとき、私は自分自身が無限の定義のように見えたことを暗記するのを助けるためにフラッシュカードを作るのに何時間も費やしました。 私はそれを退屈だが必要不可欠な生物学者クラブへのイニシエーションと見なしました。 基本的には、それはちょっとしたやり方ではありますが、私は自分自身に、高次の問題を働く科学者と話し合うことができるようになるために語彙を学ぶ必要があると言いました。 私が私のキャリアの中でさらに進歩したときに私が感謝するようになったのは、かつて白黒の、正しいか間違っているかの定義がどれほど微妙であるかです。 ドーパミンニューロンの定義のような科学的定義は、単に共通の言語を提供するのではありません。 それらは私たちの調査の本質を構成しています。 私たちは実験を進めるためにこの構造を必要とします、しかし私たちがそうするように、私たちはまたこれらの定義が私たちを制限することができる方法を知っておく必要があります。 定義されたグループを互いに比較します。 グループ平均について話します。 そのため、私たちのグループにどのようなものが含まれているかは、データの外観や決定する意味に劇的に影響する可能性があります。 したがって、私達は私達の分類に固有の偏りを常に意識しなければなりません。 おそらく定義はそれほど退屈ではありません。 本物の科学者のように考える方法を生徒に教える一方で、これらの警告についての議論はイントロコースの教材をかなり刺激するかもしれません。
神経細胞型を定義するという特定の問題は、実際にはかなり時宜を得たものであることがわかりました。 ほんの数週間前、最初の 中間報告 BRAINイニシアチブワーキンググループのメンバーが出ました(Astra Bryantの記事も参照) 役職 話題になっている)。 その中で、FY 2014のための9つの優先度の高い研究領域が概説されています、それらのうちの最初のものは「細胞型の国勢調査を生成する」です。レポートは私がここで議論している問題を認識します:
経験、結合性および神経調節物質を含む様々な要因が最初に類似した神経細胞の分子的、電気的および構造的性質を多様化させることがあるので、神経細胞型が何であるかについてはまだコンセンサスがない。 場合によっては、サブタイプを互いに区別する明確な境界がないことさえあります。 それにもかかわらず、型は不変で一般的には内在的な特性によって暫定的に定義することができ、そしてこの分類は国勢調査のための良い出発点を提供することができるという一般的な合意があります。 したがって、国勢調査はよく説明された大クラスのニューロン(例えば、皮質の興奮性錐体ニューロン)から始めて、次にこれらの分類内のより細かいカテゴリーに進むべきである。 この国勢調査は、最初は不完全であり、繰り返しを経て改善されるという知識を持って行われます。
「ドーパミンニューロンとは何ですか?」という質問に対する答えは、間もなくやってくるわけではありませんが、その質問に対する注目を集めること、およびそれに続くべき資金調達は、重要な第一歩です。 それを応援します。