Front Behav Neurosci。 2015は19、9は128です。 doi:10.3389 / fnbeh.2015.00128 eコレクション2015。
抽象
古典的条件付けでは、最初は中立的な刺激(条件付け刺激、CS)は、生物学的に顕著なイベント(無条件刺激、米国)に関連付けられます。これは、痛み(嫌悪条件付け)または食物(食欲条件付け)である可能性があります。 いくつかの関連付けの後、CSはそれぞれ防御的または完全な応答を開始できます。 嫌悪条件付けとは対照的に、食欲条件付けが人間で調査されることはめったにありませんが、正常および病理学的行動(例えば、肥満、中毒)に対するその重要性は否定できません。 本研究は、食欲調節に関する動物の所見を、米国として食物を使用する人間に翻訳することを目的としています。 8人の参加者は、彼らが空腹を感じていることを確認するために、朝食なしで午前10時からXNUMX時の間に調査されました。 XNUMXつの取得フェーズで、XNUMXつの幾何学的形状(avCS +)は嫌悪的なUS(痛みを伴う電気ショック)を予測し、別の形状(appCS +)は食欲をそそるUS(参加者の好みに応じてチョコレートまたは塩味のプレッツェル)を予測し、XNUMX番目の形状(CS– )どちらの米国も予測しませんでした。 絶滅の段階で、これらのXNUMXつの形状と新しい形状(NEW)が、米国での配信なしで再び提示されました。 驚愕反応と皮膚コンダクタンス反応(SCR)だけでなく、価数と覚醒の評価も学習指標として収集されました。 嫌悪的で食欲をそそる条件付けが成功したことがわかりました。 一方では、avCS +は、CS–よりも否定的で刺激的であると評価され、驚異的な増強と強化されたSCRを誘発しました。 一方、appCS +はCS–よりもポジティブと評価され、驚愕減衰とより大きなSCRを誘発しました。 要約すると、食欲学習と通常の嫌悪学習を実証することにより、(空腹の)人間の動物の発見を確認することに成功しました。
イントロダクション
脅威と食物を予測することは、あらゆる生物の生存にとって非常に重要です。 古典的条件付け(パブロフ、1927)軽度の痛みを伴う電気ショック(嫌悪無条件刺激、米国)などの嫌悪イベント、または食物ペレット(米国嗜好)などの嗜好イベントの前に、合図が数回繰り返される。 その後、この刺激のみ(現在はCS、CS +と表示されている)は、防御的または完全な応答をそれぞれ引き出すことができる。 前者の連想学習は嫌悪条件付けと呼ばれ、後者は食欲の条件付けと呼ばれます。
食欲の事象の予測は、嫌悪の事象の予測と同じくらい生存のために重要ですが、食欲の条件付けは動物ではあまり研究されていません(ブートン&ペック、1989; Koch他、1996; McDannaldら、2011レビューを見るには Martin-Soelchら、2007)人間だけでなくKluckenら、2009, 2013; オースティンとドゥカ、2010; Delgado他、2011; レヴィとグリムチャー、2011) この研究の欠如は恐らく嫌悪的なパラダイムと比較して食欲をそそるパラダイムの複雑さによるものかもしれません。 例えば、主な強化剤としての食べ物は、その生物が飢えているときには、やりがいのあるものになるように届けられなければなりません(最近の総説については、 Dickinson and Balleine、1994; Clarkら、2012) 人間の研究では、この困難はお金を使うことによって克服されました。オースティンとドゥカ、2010; Delgado他、2011; レヴィとグリムチャー、2011)またはエロティックな写真(Kluckenら、2009, 2013) しかしながら、神経画像研究は、一次(すなわち、スナックまたは飲料)および二次(すなわち、金)補強剤がいくつかの一般的な脳領域(例えば、線条体)を活性化するが異なる活性化パターンも伴うことを指摘した。Delgado他、2011; レヴィとグリムチャー、2011).
我々の知る限りでは、ほんの少しの人間のコンディショニング研究が臭いのような一次食欲増強剤の効果を調べた。Gottfried他、2002)、 水 (Kumarら、2008)、または食べ物(Prévostet al。、2012) 最初の研究では、 Gottfried等。 (2002) 不快、愉快、または中立臭のいずれかを伴う中立面(CS)。 興味深いことに、彼らは、食欲不振CS +対嫌悪CS +に応答して、眼窩前頭皮質(OFC)および腹側線条体においてより大きな活性化を見出した。 彼らは、OFCが匂いの価値を処理し、感情的価値を嗅覚(米国)から視覚(CS)システムに移すことに関与していると結論を下しました。 (腹側)線条体活性化は、食欲CS +によって誘発される食欲CRを反映すると解釈されてきた。 2番目の研究では、 Kumar et al。 (2008) 彼らは参加者(健常者と大うつ病患者)を早朝に検査室に招待し、スキャン前にのどが渇いていることを確認するために夜間は飲酒を控えるように求めました。 フラクタル写真は米国のCSと0.1 mlの水でした。 興味深いことに、他の活性化の中でも、健常者(うつ病患者ではない)は、食欲をそそるCS +に対して腹側線条体においてより大きな活性化を示し、この刺激がやりがいのあるものとして処理されたことを示唆する。 3番目の研究では、 Prévost等。 (2012) また、参加者の好みに応じて、フラクタル写真をCSとして、甘いまたは塩辛いスナックを米国として提示しました。 CSは6秒間提示され、最後のXNUMX秒間に食べ物の写真が追加で提示されました。 食べ物の写真が表示されるたびに、実験者は参加者の手に食べ物を置き、参加者はすぐに軽食を食べることができました。 残念ながら、著者らは古典的条件付け段階で脳の活性化を食欲のあるCS +に報告しませんでした。 しかし、彼らは、報酬のないCS–と比較して、報酬のあるCS +に対する心臓の減速が少ないことを観察しました。これは、異なる条件付け効果を示しています。
食欲の条件付けのための指標として驚愕反応を用いた研究の欠如は、特に嫌悪条件付けにおけるその広範な使用を考慮すると、驚くべきことである。 驚愕反応は、突然の予期せぬ強い嫌悪的な出来事に対する先祖的で自動的な防御反応です(コッホ、1999) この防御反応は、蝸牛神経根ニューロン、網状層の尾側核(PnC)および脊髄運動ニューロンを含む比較的単純な神経経路によって仲介される(フェントとファンスロー、1999; コッホ、1999) 動物実験は、驚愕増強が扁桃体からPnCへの予測に依存することを明らかにした。フェントとファンスロー、1999; コッホ、1999一方、驚愕の減衰は無傷の側坐核(NAcc、 コッホ、1999) 驚愕反応のそのような調節は、提示された前景刺激の価数に対する有用な暗黙の尺度である。 したがって、増強は負の原子価を示し、減衰は正の原子価を示し、両方とも認知プロセスによって大きく影響されることはない(ハムアンドウィーク、2005; Andreattaら、2010) 我々の知る限りでは、依存測定として驚愕反応を測定することによって食欲調節を調査した動物研究は1つだけである。 興味深いことに、試験したラットは、スクロース溶液の送達に関連するCS +(すなわち、光)への驚異的な減衰を示した。 特に、そのような弱毒化はNAcc病変を有する動物では損なわれたが、扁桃体病変を持つ動物では損なわれず、NAccが食欲増進性CRの誘発および驚愕反応の減弱において特定の役割を果たすことを示唆する。Koch他、1996).
現在の研究は我々の知る限りでは、アメリカのように一次強化剤を使用することによってこの食欲をそそるコンディショニングパラダイムを人間に翻訳する、すなわち甘い(チョコレートSmarties)®)または塩味(小塩味プレッツェル)食品、およびCRの尺度として驚きの変調。 我々は、他の刺激剤、すなわちavCS +およびCS-と比較して、驚くべき減衰、向上したSCR、および正の原子価評価に反映されるように、appCS +が強い食欲CRを誘発すると予想した。
材料と方法
一般
5人のボランティアが研究への参加を受け入れ、コースクレジットを受け取りました。 2人の参加者は分析から除外されました。33人は技術的な問題のため、16人は非応答者としてコード化されたため(平均驚愕振幅<22.09μV)、XNUMX人は条件ごとに十分な驚愕反応を示さなかったため(最小= XNUMX;詳細については、材料と方法を参照してください)。 最終的に、XNUMX人の参加者が分析で考慮されました(XNUMX人の男性;平均年齢:XNUMX歳、 SD:2.84; 範囲:18 - 29年) 4人の参加者はネイティブのドイツ人ではなく、6人は左利きでした。 1人の参加者は実験を通してCS-US協会に気づかないままでした(手順を見てください)、私達は彼の応答が正常で結果に影響を及ぼさなかったのでこの参加者を除外しないことにしました。
材料
無条件刺激(アメリカ)
2種類のアメリカが使われました。 嫌悪的な米国として、我々は参加者の非優勢な前腕に軽度の痛みを伴う電気ショックを与えた。 感電は、直径9 mm、間隔30 cmの2つの電極を用いて行いました。 電気刺激は、電流刺激装置(Digitimer DS50A、Welwyn Garden City、英国、200 V、最大7 mA)によって発生させた、周波数400 Hzおよび持続時間9.99 msのパルス刺激からなった。 電気ショックの強度は、前述の閾値手順を介して個々に決定された。Andreattaら、2010) 手短に言えば、参加者は、0.5 mAの間隔で2つの一連の上昇および下降強度を受けた。 彼らは0(全く痛みを感じない)から10(本当に激しい痛み)までしきい値のアンカーとしての4(ちょうど目立った痛み)の範囲の視覚スケールで各刺激を評価しなければなりませんでした。 電気刺激の平均強度は2.12 mA(SD それは痛みを伴うと評価されました(= 0.56)。M = 6.45、 SD = 1.73) 食欲をそそるアメリカはチョコレート(Smarties)から成りました®)または小さな塩味のプレッツェル。 食欲をそそる米国の選択は、予備面接中に報告された参加者の個人的な好みに依存していました。 つまり、参加者は、朝食時に通常塩辛い食べ物を食べるのか甘い食べ物を食べるのかを報告する必要がありました。 また、実験中にチョコレートと塩味のプレッツェルのどちらを好むかを自由に選択することもできました。 結局、22人の参加者がチョコレートを選び、11人が小さな塩味のプレッツェルを選びました。
条件付き刺激(CS)
対角線が8 cmの幾何学的形状(青い正方形、黄色い円、緑色の三角形、赤い六角形)をCSとして提示した。 8の場合、図形は黒いコンピューター画面の中央に表示されていました。 1つの形状(avCS +)は常に嫌悪的な米国(痛みを伴う感電)と関連し、1つの形状(appCS +)は常に食欲をそそるUS(チョコレートまたは塩味のプレッツェル)と関連しました。第4の形(NEW)は絶滅の段階で提示されたが、その中立性を保証するための獲得段階の間ではなかった。
驚きのプローブ
驚愕プローブとして、持続時間が103ミリ秒の50dBのホワイトノイズを使用しました。 音響刺激は、ヘッドホンを介してバイノーラルで提示され、形状の開始から4〜6秒後にランダムに発生しました。
アンケート
実験の前後に、参加者はドイツ語版の国家特性不安インベントリー(STAI、 Lauxら、1981)および正の負の影響スケジュール(PANAS、 Krohneら、1996)。 STAIは、参加者の特性および/または状態不安を評価するためのインベントリであり、特性バージョンと状態バージョンの両方の20項目で構成されています。 以前の参加者の不安レベル(M = 37.06、 SD = 7.80)以降M = 39.33、 SD = 9.16)実験は有意に変化しませんでした[t(32) = 1.61、 p = 0.117]。 現在のサンプルの特性不安スコアは、20と58の間の範囲でした(M = 36.6、 SD = 8.98)、これは成人の公表されている正常範囲と同等です(Lauxら、1981) パナス(Krohneら、1996)は、ポジティブな気分とネガティブな気分の指標です。 ポジティブな感情の尺度(PAS)で高いスコアを持つ個人は、熱意などの感情になりがちですが、ネガティブな感情の尺度(NAS)で高いスコアを持つ個人は、苦痛などの感情になりやすいです。 各項目は形容詞で構成されており、参加者は形容詞がその特定の瞬間の感情をどの程度反映しているかを1(ごくわずか)から5(非常に)の範囲で示します。 実験の結果、参加者の悪影響に有意差は見られませんでした[開始: M = 11.67、 SD = 2.29; 終わり: M = 12.88、 SD = 4.69; t(32) = 1.55、 p = 0.130]。 どういうわけか、参加者は最初から積極的な気分を大きく変えました(M = 26.72、 SD 最後まで(= 4.70)M = 23.66、 SD 実験の= 7.02)t(31) = 3.11、 p = 0.004]。 参加者の前向きな気分のこの低下は、パラダイムの不快感に関連している可能性があります(痛みを伴う電気ショックと嫌悪的なホワイトノイズが提示されました)。
手順
研究室に到着すると、参加者はヴュルツブルク大学心理学科の倫理委員会によって承認されたインフォームド・コンセント用紙を読み、署名した。 彼らはCSとUSの間の偶発事故について知らされていなかった。 質問票に記入した後、電極を取り付け、痛み閾値処置を上記のように実施した。
間に 慣れフェーズ4つの幾何学的形状は、XNUMXとXNUMXとの間で変動する試行間間隔(ITI)で2回提示された(平均:XNUMX)。 この段階では、米国や驚愕のプローブは届けられませんでした。
取得段階の前は、初期の驚愕反応性を低下させるために、7〜15ごとに7回のホワイトノイズが送出されました。
次の二つ 取得フェーズ 同一でした(図 1) 各取得段階は、24 CS–試行、8 avCS +試行、および8 appCS +試行の8試行で構成されています。 CSシーケンスは擬似ランダムであり、唯一の制限は、同じ刺激が連続して2回を超えて提示され得ないということである。 特に、avCS +は稲妻と一緒に感電の象徴として提示され、痛みを伴う米国はそのオフセットで配達されました。 appCS +はSmartiesまたは塩味のプレッツェルのイメージと組み合わせて表示され、参加者はjarからSmartiesまたはプレッツェルを選ぶことができます。 CS - は禁止記号と一緒に表示され、米国は配達されませんでした。 各タイプの3回のXNUMX CS提示の間、刺激開始後、驚愕プローブがXNUMXとXNUMXとの間に送達された。 それらの予測不可能性を保証し、驚きの慣れを減らすために、3つの追加の驚きプローブがITIの間に発表されました。 黒い画面で構成されるITIは、8と4の間で変化し、平均は6です。
図1
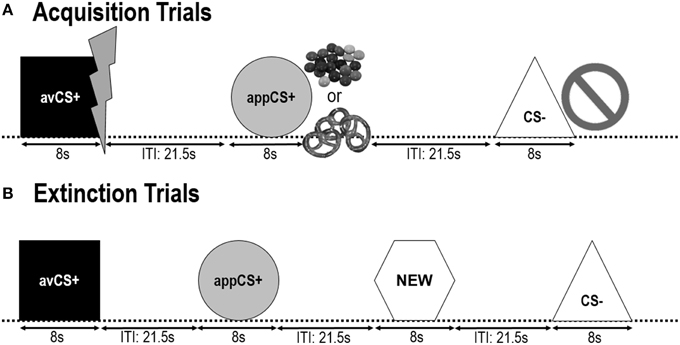
図1 2つの取得段階(A)と消滅段階(B)の間の試行。 参加者は、1つの形状(avCS +)は軽い痛みを伴う感電を予測し、1つの形状(appCS +)はチョコレートまたは小さな塩味のプレッツェル(自分の好みによる)を予測し、3つ目の形状(CS–)は予測できないことを知りました。生物学的に顕著なイベント。 それぞれの形は、感電、賢い/塩味のプレッツェル、または米国との関係に応じて何も描かれていない絵を描いて提示されました。 消滅段階の間に、3つの幾何学的な形は再び提示されました、しかし、米国は届けられませんでした。 加えて、第4の幾何学的形状(NEW)が中立対照として提示された。
間に 消滅フェーズ参加者は、3つの幾何学的形状(avCS +、appCS +、CS - )と、新たな中立形状(NEW)を再び見ました。 米国は配達されず、形はフラッシュ、チョコレート/塩味のプレッツェル、または禁止の絵と一緒に提示されませんでした。 各刺激は擬似乱数順に8回提示され(すなわち、同じ刺激は2回以上続けて提示されなかった)、32試行を作成した。 驚愕プローブ刺激は、各CSタイプの4刺激提示のうち8中に提示された。 取得および慣れ段階と同様に、ITIはXNUMXとXNUMXとの間で変化し、そしてXNUMX追加驚愕プローブはITIの間に予測不可能に送達された。
各段階の後、参加者は1から9までの範囲の視覚的アナログ尺度(VAS)でCSの価数(快感)と覚醒(興奮)を評価した。 バランススケールは、「非常に不快」を示す「XNUMX」から「非常に快適」を示す「XNUMX」までの範囲であり、覚醒スケールは、「穏やか」を示す「XNUMX」から「興奮」を示す「XNUMX」までの範囲であった。 2つの取得段階と絶滅段階の後に評価されます。 参加者は1の幾何学的形状を見た後、この形状が電気ショックに関連しているのか、チョコレート/塩味のプレッツェルに関連しているのか、何も関連がないのかを示す必要がありました。 特に、すべての参加者(1人を除く)は、アクイジション9後の偶発事故を認識していました。
データ削減
生理学的応答は、V-Amp 16増幅器およびVision Recorder V-Amp Editionソフトウェア(バージョン1.03.0004、BrainProducts Inc.、ドイツ、ミュンヘン)を用いて記録した。 1000 Hzのサンプリングレートと50 Hzノッチフィルターを適用した。 オフライン分析はブレインビジョンアナライザー(バージョンXNUMX;ブレインプロダクツ社、ミュンヘン、ドイツ)を用いて行った。
驚きの反応
驚愕反応は左側の筋電図検査(EMG)によって測定された 眼窩 2本の5 mm Ag / AgCl電極を備えた筋肉。 ガイドラインに従って(Blumenthalら、20051つの電極は瞳孔の下に配置され、2つ目の電極は横方向にXNUMXcmに配置された。 接地電極および参照電極をそれぞれ左右のマストイドに配置した。 電極を取り付ける前に、インピーダンスを1kΩ以下に保つために、皮膚を軽く研磨し、アルコールで洗浄しました。 筋電図信号を、10 Hz低カットオフフィルターおよび28 Hz高カットオフフィルターでオフラインフィルタリングした。 次に、EMG信号を修正し、移動平均500 msを適用しました。 ベースラインとして驚愕プローブの発症前の50 msを使用した(Grillonら、2006) 驚愕プローブに対する応答を手動でスコア化し、過度のベースラインシフト(±5μV)または運動アーチファクトを伴う試験はさらなる分析から除外した。 5μVより低い驚愕反応はゼロとしてコード化され、驚愕の大きさの計算のために考慮された。Blumenthalら、2005) 全体として、10.4%の試行が棄却され、参加者を分析プールに保持するには、各条件について、獲得段階における2驚愕反応のうち最低3、および消光相における4驚愕反応のうち8が必要であった。 このため、4名の参加者は除外されました。 ピーク振幅は、驚愕プローブの開始後の20〜120 msの時間枠の間のベースラインに対する最大ピークとして定義されました。 生データは、被験者内で正規化されました。 z個人差の影響を減らし、心理的プロセスをよりよく検出するために、スコアとTスコアを選択します。 各条件(avCS +、appCS +、CS - 、NEW、およびITI)についてTスコアを平均しました。 驚愕増強または驚愕減衰を調べるために、ITI驚愕反応についてのスコアを、各状態についての驚愕反応から差し引いた。
皮膚コンダクタンス応答(SCR)
非利き手の手のひらに置いた2つの5 mm Ag / AgCl電極を使用して、皮膚コンダクタンス応答(SCR)を記録した。 ガルバニック応答は、1 Hz高カットオフフィルターを用いてオフライン濾過した。 SCRは、応答開始(刺激開始後の1〜3)と応答ピーク(µS)との差(μS)として定義されました。トラネルとダマシオ、1994; Delgado他、2011) 驚愕プローブを含む試験はSCRの分析において考慮されなかった。 0.02μS未満の応答はゼロとしてコード化されました。 5人のさらなる参加者は、平均SCRがXNUMXμSより低いため、SCR分析から除外した。 生の皮膚コンダクタンスデータは、分布を正規化するために平方根変換され、スコアは2つの取得フェーズ(avCS +、appCS +、CS–)と消光フェーズ(avCS +、appCS +、CS–、およびNEW)で別々に平均化されました。 )
統計分析
全てのデータは、ウィンドウズ用SPSS(バージョンXNUMX、SPSS Inc.)で分析した。 生理学的応答については、2つの取得段階および消光段階について分離多変量分散分析(ANOVA)を計算した。 取得フェーズのANOVAは、被験者内要因として刺激(avCS +、appCS +、CS–)とフェーズ(取得20.0、取得1)を持っていました。 絶滅期のANOVAは、被験者内因子として刺激(avCS +、appCS +、CS–、NEW)のみでした。 価数、覚醒度、および偶発性の評価は、被験者内要因の刺激(avCS +、appCS +、CS–、およびNEW)と位相を含む別々のANOVAで分析しました。 この因子は、価数および覚醒評価について4つのレベル(T2:習慣化フェーズ後、T1:2回目の取得フェーズ後、T2:消滅フェーズ後)を、3つのレベルで分割された(T3:後)最初の取得フェーズ、T4:2番目の取得フェーズの後、T1:消光フェーズの後)
すべての分析についてアルファ(α)レベルをXNUMXに設定した。 効果の大きさは部分ηとして報告されます。2.
結果
各相の価数と覚醒度の評価を図に示します。 2; 驚愕反応とSCRを図に示す。 3.
図2
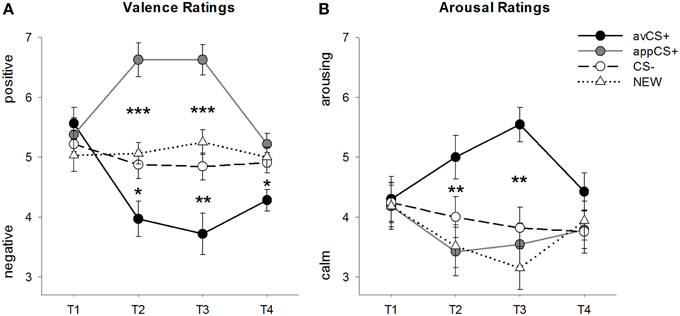
図2 原子価(A)と覚醒(B)の評価。 行(標準誤差付き)は、慣れフェーズ(T1)、取得1(T2)、取得2(T3)、および消光フェーズ(T4)の後の格付けを表します。 嫌悪的なCS +(黒い実線)は、CS-(黒い破線)とNEW(黒い点線)と比較して、2つの獲得フェーズの後に負の価数と高い覚醒を獲得しました。 重要なことに、食欲をそそるCS +(灰色の実線)は、CS - およびNEWと比較して正の原子価を獲得しました。 *p <0.05、**p > 0.01、***p <0.001。
図3
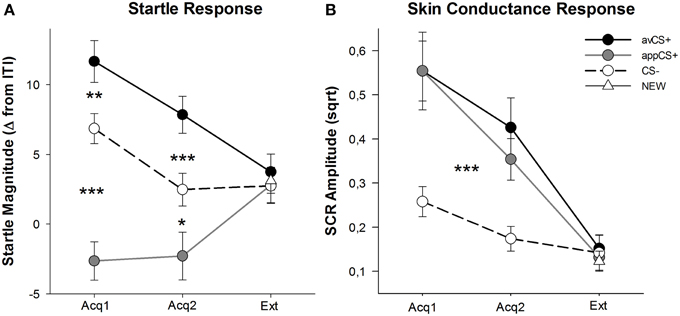
図3 第一取得段階(Acq1)、第二取得段階(Acq2)、および消光段階(Ext)中の驚愕反応(A)および皮膚コンダクタンス(B)(標準誤差あり)。 驚愕反応は、獲得期のCS-(黒い破線)と比較して、嫌悪的なCS +(黒い実線)に対して有意に増強され、食欲のあるCS +(灰色の実線)に対して有意に減衰した。 SCRはCS–と比較してavCS +とappCS +の方が有意に大きかった。 消滅段階の間に違いは明らかにされなかった。 *p <0.05、**p > 0.01、***p <0.001。
評価
の分散分析 原子価評価 獲得の間に刺激の重要な主な効果を明らかにしました[F(3、93) = 17.26、GG-ε= 0.801、 p <0.001、部分η2 = 0.358]とフェーズ[F(3、93) = 3.30、GG-ε= 0.731、 p = 0.039、部分η2 刺激と位相の間の重要な相互作用と同様に= 0.096]F(9、279) = 9.54、GG-ε= 0.463、 p <0.001、部分η2 = 0.235]。 ファローアップ t実験は、実験開始時の4つの幾何学的形状の原子価が同一であることを示している(ps> 0.19)、一方、Acquisition1とAcquisition2の両方の後、avCS +は特にネガティブ、appCS +は特にポジティブと評価されました。 具体的には、avCS +は、CS– [Acq1:]と比較してより負の価数であると評価されました。 t(31) = 2.34、 p = 0.026; Acq2: t(31) = 3.07、 p = 0.004]、新しい[Acq1: t(31) = 2.70、 p = 0.011; Acq2: t(31) = 3.89、 p <0.001]、およびappCS + [Acq1: t(31) = 5.41、 p <0.001; Acq2: t(31) = 6.11、 p <0.001]。 appCS +は、CS– [Acq1:よりも大幅にポジティブであると評価されました。 t(31) = 4.99、 p <0.001; Acq2: t(31) = 5.31、 p <0.001]および新しい[Acq1: t(31) = 4.92、 p <0.001; Acq2: t(31) = 4.14、 p <0.001]。 CS–とNEWの違いは決して重要ではありませんでした(ps> 0.18)。
重要なことに、チョコレートと塩味のプレッツェルに関連するappCS +の価数評価を比較しても、大きな違いは見つかりませんでした[Acq1: t(30) = 0.03、 p = 0.477; Acq2: t(30) = 0.29、 p = 0.775]。
消滅段階の後でも、avCS +はCS–と比較してさらにマイナスと評価されました。t(31) = 2.40、 p = 0.023]、新しい[t(31) = 3.40、 p = 0.002]、およびappCS + [t(31) = 3.35、 p = 0.002]。 それどころか、appCS +の価数はCS-とそれ以上変わらなかった。t(31) = 1.77、 p = 0.086]またはNEW [t(31) = 1.07、 p = 0.293]。
の分散分析 覚醒度の評価 獲得の間に刺激の重要な主な効果を明らかにしました[F(3、96) = 7.07、GG-ε= 0.737、 p = 0.001、部分η2 = 0.181]、ただし相ではない[=F(3、96) = 1.27、GG-ε= 0.805、 p = 0.289、部分η2 = 0.038]、および刺激と位相の間の重要な相互作用[F(9、288) = 4.53、GG-ε= 0.582、 p = 0.001、部分η2 = 0.124]。 ファローアップ t- テストは彼らの初期の覚醒に関して刺激の間に有意差を示さなかった(ps> 0.74)。 ただし、1つの取得フェーズの後、avCS +はNEWよりも刺激的であると評価されました[AcqXNUMX: t(32) = 2.99、 p = 0.005; Acq2: t(32) = 5.97、 p <0.001]およびappCS + [Acq1: t(32) = 2.62、 p = 0.013; Acq2: t(32) = 4.42、 p <0.001]、最初の取得フェーズの後、CSよりもわずかに興奮します[t(32) = 1.96、 p = 0.058]、しかし2回目の取得段階の後にはもっともっと興奮t(32) = 3.65、 p = 0.001]。 原子価評価とは対照的に、appCS +の覚醒はCS–およびNEWの覚醒と変わらなかった(ps > 0.13)取得フェーズ後。
原子価評価と同様に、CS–とNEWは覚醒度に違いはありませんでした(ps> 0.07)。
価数の場合と同様に、チョコレートに関連付けられたappCS +と塩味のプレッツェルに関連付けられたappCS +に違いは見られませんでした[Acq1: t(31) = 0.26、 p = 0.797; Acq2: t(31) = 0.33、 p = 0.724]。
消滅段階の後、覚醒度の格差に有意な差は見られなかった(ps> 0.08)。
追加の 事後t慣れ段階、1回目と2回目の獲得段階、および絶滅段階後の評価を比較するテスト、補足資料を参照してください。
驚きの反応
取得段階のANOVAは刺激の主な効果を返しました[F(2、64) = 49.92、GG-ε= 0.964、 p <0.001、部分η2 = 0.609]、しかし相[]ではないF(1、32) = 3.16、 p = 0.085、部分η2 = 0.090]、および有意な相互作用F(2、64) = 3.37、GG-ε= 0.875、 p = 0.048、部分η2 = 0.095]。 ファローアップ t最初の両方の試験において、CS試験と比較してavCS +に対する有意な驚愕増強作用が明らかにされた。t(32) = 3.27、 p = 0.003]と2番目の[t(32) = 4.00、 p <0.001]取得フェーズ。 avCS +に対する驚愕反応も、appCS +に対する驚愕反応と比較して大幅に増強されました。これも、両方の取得1 [t(32) = 8.20、 p <0.001]および取得2 [t(32) = 5.74、 p <0.001]。 重要なことに、appCS +に対する驚愕の大きさは、両方の取得1の間にCS–と比較して大幅に減衰しました[t(32) = 6.34、 p <0.001]および取得2 [t(32) = 2.91、 p = 0.007]。 やはり評価と一致して、チョコレートまたは塩味のプレッツェルと関連した場合、appCS +に関して驚異的な反応は見られなかった[Acq1: t(31) = 1.04、 p = 0.309; Acq2: t(31) = 0.07、 p = 0.947]。 消滅期の間、有意な影響は見られませんでした。F(3、96) = 0.26、GG-ε= 0.906、 p = 0.833、部分η2 = 0.008]。
皮膚コンダクタンス応答(SCR)
2つの取得段階におけるSCRのANOVAから、主な効果は刺激されます[F(2、54) = 18.04、GG-ε= 0.908、 p <0.001、部分η2 = 0.401]とフェーズ[F(1、27) = 20.91、 p <0.001、部分η2 = 0.436]は重要であることが判明しましたが、それらの相互作用はそうではありません[F(2、54) = 0.68、GG-ε= 0.637、 p = 0.451、部分η2 = 0.024]。 事後処理テストはavCS +に対して有意に大きいSCRを示しました[t(27) = 6.46、 p <0.001]およびappCS + [t(27) = 4.84、 p CS–と比較して<0.001]、参加者はavCS +およびappCS +と同等のSCRを示しました[t(27) = 0.64、 p = 0.527]。 特に、SCRにチョコレートappCS +と塩味プレッツェルappCS +との違いは見られませんでした[Acq1: t(26) = 2.55、 p = 0.120; Acq2: t(26) = 1.29、 p = 0.210]。 格付けと驚愕反応に当てはまるように、絶滅相に有意な影響は見られませんでした。F(3、81) = 0.28、GG-ε= 0.634、 p = 0.743、部分η2 = 0.010]。
議論
この研究の目的は、無条件刺激としての一次強化剤を用いた古典的な食欲調節条件付けパラダイムを使用することによって動物の知見をヒトに翻訳することであった。 そうするために、参加者は空腹であることを確認するために朝食なしで早朝に研究室に来ました、そして、彼らの好みに応じて、チョコレートや塩味のプレッツェルは食欲をそそるUSとして使われました。 取得段階では、1つの幾何学的形状(avCS +)が軽い痛みを伴う感電(嫌悪感のあるUS)、別の形状(appCS +)が嗜好的なUS、および3番目の形状(CS–)と関連しました。嫌悪する米国。 結果は、明示的な言語レベル(すなわち評価)、潜在的な行動レベル(すなわち驚愕反応)、および生理学的レベル(すなわちSCR)での嫌悪的かつ食欲的な条件付けの成功を示しています。 具体的には、CSと比較したavCS +は、より負の原子価評価、より高い覚醒評価、驚愕増強、およびより高いSCRを引き出した。 最も重要なことには、appCS +がCS - と比較して、より正の価数評価、驚きの減衰、およびより大きなSCRを引き起こしました。 以前の研究では、脅威を予測する刺激(avCS +)が嫌悪として評価され、より大きな恐怖反応を引き出し、そして生理的覚醒を増加させることがわかったので、嫌悪条件付けに関する我々の発見は予想と一致した。フェントとファンスロー、1999; ハムアンドウィーク、2005; Andreattaら、2010, 2013) 食欲のコンディショニングに関する我々の結果はまた、報酬予測予測(appCS +)がポジティブであると評価され、恐怖反応を抑制し、そして生理学的覚醒を増加させることを示す以前のヒトおよび動物の研究と一致した。Koch他、1996; Gottfried他、2002; Kumarら、2008; Kluckenら、2009, 2013; オースティンとドゥカ、2010; Prévostet al。、2012) 我々の知る限りでは、これは一次報酬を予測する刺激に対するヒトの条件付き驚愕の減弱を示す最初の研究である。 重要なことに、動物実験の結果を転送し確認することができました。Koch他、1996) この動物実験は、ラットにおける驚愕の減弱が、NAcc(腹側線条体の一部)からPnCへの投射に依存することを実証した。 したがって、我々の研究における驚愕の減弱はNAcc活性を意味する可能性があり、これもfMRI所見と一致している(Gottfried他、2002; Kumarら、2008; Kluckenら、2009, 2013; Delgado他、2011; レヴィとグリムチャー、2011) したがって、明示的(格付け)と暗黙的(驚愕の減衰)の両方の正の原子価が示すように、我々は我々の食欲をそそる条件付けパラダイムは成功したと結論する。
この新しいだがかなり予測可能な発見に加えて、さらに2つの興味深い結果が言及に値する。 第一に、appCS +に対する口頭および生理的覚醒反応が解離した。 第二に、我々は嫌悪的なCRと比較して食欲をそそるCRのより速い消滅を見いだした。
SCRは交感神経活性化を示し、これは報酬関連刺激(appCS +)および脅威関連刺激(avCS +)の両方に増加する。 この結果は、エロティックな写真が食欲をそそる米国として使用されたという以前のコンディショニング研究と一致しています(Kluckenら、2013そして、お金を予測する手がかりと嫌悪音を予測する手がかりに匹敵するSCRを明らかにした研究(オースティンとドゥカ、2010) 特に、SCRは交感神経系の活性化に関連する指向性反応です。 この反応は、やる気を起こさせるような顕著な出来事に対する行動反応の準備を反映していると示唆されている(ブラッドリー、2009) これに基づいて、我々は、脅威関連刺激と報酬関連刺激の両方が、連続的な行動反応に対する準備反応を誘発したと考える。 言い換えれば、食物と痛みの信号は、それぞれ接近行動と回避行動の準備を始めました。 高い生理学的覚醒とは対照的に、言葉による反応は報酬に関連した刺激に対する低い覚醒を示した。 言葉による覚醒は、生理的活性化よりもむしろ米国の興奮性により影響される可能性があります。 それ自体が。 実際、appCS +とavCS +はそれぞれ低と高興奮のUSにリンクされているため、低と高興奮の両方と評価されました。 残念ながら、我々は米国の覚醒評価を収集していないので、この仮説を明示的に検証することは将来の研究次第です。
消滅段階の間、米国は届けられませんでした。 これは絶滅と呼ばれる新しい抑制的学習を誘発したかもしれません(総説については Milad and Quirk、2012その結果、嫌悪CRと食欲CRの両方が減少する。 明らかに、行動的(すなわち驚愕反応)および生理学的(すなわちSCR)レベルでは、avCS +、appCS +、およびCS-に対する異なる反応はもはや検出されなかった。 同様に、絶滅相の後に評価された条件付き刺激の明白な覚醒は低レベルで等しく、絶滅学習の成功を示唆している。 ただし、avCS +は依然としてCS - よりも大幅にマイナスに評価されていましたが、appCS +の価数はCS - の価数と変わらなくなりました。 嫌悪的な明白な反応のより遅い消滅は進化論的保守主義によるかもしれません、それは脅威の信号への無反応が生命を脅かすかもしれないので脅威の信号が特に忘れがたいことを意味します。
それでもなお一つの問題が残っています:それでは、なぜ驚愕反応(すなわち暗黙の原子価)が消滅段階の間に完全に消えたのでしょうか? まず、これらの回答はフェーズの全過程にわたって計算されました。 したがって、絶滅相の最初の試験中に差別的CRが依然として検出可能であると考えられる。 探索的な方法で、我々はこの仮説をたどり、絶滅期を通して驚愕反応とSCRの両方を調べました(補足資料参照)。 有意な差は見られませんでしたが、CS - および新規対照刺激と比較して、avCS +の方がわずかに高い驚愕強度を観察しました。 また、CS - や消滅期の最初の段階での新規対照刺激と比較して、appCS +に対する驚異的な減衰がわずかに多くなっていますが、数回の試行で消失しました。 appCS +へのSCRは、2ndの絶滅試験の後にすでに低下していますが、avCS +へのSCRは、NEWへのSCRと比較して、ほとんどすべての絶滅に対してより高いままでした。 我々はこれらの結果を細心の注意を払って解釈すべきであるが、驚愕反応は原子価評価と平行しているように思われた。 さらに、驚愕反応とSCRの両方は、脅威の刺激に対する進化的保守主義の考えをさらに支持する。
最後に、我々はこの研究のいくつかの制限を認識しなければなりません。 まず、技術的な問題により、チョコレートと塩味のプレッツェルについての快適さ(および覚醒)の評価を報告できませんでした。 しかし、食欲をそそるCRは、参加者が実際に2人の米国を食欲をそそるものとして経験したことを示唆している。 第二に、嫌悪感と食欲のアメリカの期間は非常に異なっていました。 したがって、痛みを伴う感電はavCS +のオフセットで行われ、正確に200 ms持続しましたが、appCS +の開始後、チョコレートと塩味のプレッツェルは2 s前後で参加者に提示されました。個人はそれらを食べました。 このようにして食欲をそそる米国を提供するという選択は、以前のヒトでの研究に基づいていました(Prévostet al。、2012) ただし、appCS + offsetでジュースやジンジャーエールを一口食べるなど、嫌悪的なUSに匹敵する食欲をそそるUSを提供することは方法論的によりエレガントです。 この大きな知覚の違いを補うために、米国を象徴する視覚的刺激と組み合わせて幾何学的形状を提示しました。 第三に、我々は絶滅期に急速に消滅した生理学的反応が方法論的側面に起因するという可能性を確実に排除することはできない。 実際、この段階での視覚的刺激は、獲得段階の場合のように米国のシンボルと関連して提示されていません。 CSの提示のみがCRに影響を及ぼした可能性があり、そのために最初の絶滅試験中に有意差は見られなかった。 しかしながら、我々の素早い消滅反応は、CS +(顔面)が獲得中に米国(悲鳴)と複合的に提示されたが消滅中には消滅しなかった以前の研究における消滅反応と一致している。Lissek他、2008).
結論として、我々は脅威に関連した刺激と報酬に関連した刺激に対してそれぞれ巧妙な嫌悪感と食欲をそそる条件付き反応を見出した。 興味深いことに、明示的(格付け)および暗黙的(驚愕反射)レベルの反応は、avCS +が陰性および誘発性驚愕増強として報告され、appCS +が陽性および誘発性驚愕減衰として報告されたという点で相乗的に機能した。 さらに、appCS +の明示的(評価)および生理学的(SCR)覚醒は、2つの異なるプロセスを反映して解離しました。
利益相反に関する声明
著者らは、潜在的な利益相反として解釈される可能性がある商業的または金銭的関係がない状態で研究が行われたと宣言しています。
謝辞
この研究は、共同研究センター「恐怖、不安、および不安障害」、SFB-TRR 58、プロジェクトB1によって支援された。
補足資料
この記事の補足資料は、次のWebサイトで入手できます。 http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
参考情報
Andreatta、M。、Mühlberger、A。、Glotzbach-Schoon、E。およびPauli、P。(2013)。 疼痛の予測可能性は、救済関連刺激の原子価評価を逆転させる。 前面。 シスト 神経症。 7:53 土井:10.3389 / fnsys.2013.00053
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Andreatta、M。、Mühlberger、A。、Yarali、A。、Gerber、B。、およびPauli、P。(2010)。 ヒトにおける疼痛緩和学習後の暗黙的および明示的条件付き原子価間の裂け目 手順 Biol。 サイ。 277、2411〜2416。 土井:10.1098 / rspb.2010.0103
PubMed要約 | CrossRefフルテキスト | Google Scholarの
オースティン、AJ、およびDuka、T.(2010)。 パブロフの条件付けにおける食欲と嫌悪の結果に対する注意のメカニズム ふるまい。 脳の解像度。 213、19〜26。 土井:10.1016 / j.bbr.2010.04.019
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Blumenthal、TD、Cuthbert、BN、Filion、DL、Hackley、S、Lipp、OV、およびvan Boxtel、A(2005)。 委員会報告ヒトの驚きの瞬目筋電図検査のための指針 精神生理学 42、1〜15。 土井:10.1111 / j.1469-8986.2005.00271.x
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Bouton、ME、およびPeck、CA(1989)。 食欲をそそるコンディショニング製剤におけるコンディショニング、絶滅、および回復に対する文脈効果。 アニメ。 学びなさい。 ふるまい。 17、188〜198。 土井:10.3758 / BF03207634
CrossRefフルテキスト | Google Scholarの
ブラッドリー、MM(2009)。 自然な選択的注意:方向づけと感情 精神生理学 46、1〜11。 土井:10.1111 / j.1469-8986.2008.00702.x
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Clark、JJ、Hollon、NG、およびPhillips、PEM(2012)。 学習と意思決定におけるパブロフ評価システム Curr。 意見 ニューロビオール。 22、1054〜1061。 土井:10.1016 / j.conb.2012.06.004
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Delgado、MR、Jou、RL、およびPhelps、EA(2011)。 一次および二次強化剤によるヒトの嫌悪条件付けの基礎となる神経系 前面。 神経症。 5:71 土井:10.3389 / fnins.2011.00071
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Dickinson、A。およびBalleine、B(1994)。 目標指向行動の動機づけ制御 アニメ。 学びなさい。 ふるまい。 22、1〜18。 土井:10.3758 / BF03199951
CrossRefフルテキスト | Google Scholarの
Fendt、M。、およびFanselow、MS(1999)。 条件付き恐怖の神経解剖学的および神経化学的基礎 神経症。 バイオ行動。 改訂。 23、743〜760。 土井:10.1016 / j.ijpsycho.2012.09.006
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Gottfried、JA、O'Doherty、J。、およびDolan、RJ(2002)。 イベント関連の機能的磁気共鳴画像法を使用して研究された人間の食欲と嫌悪の嗅覚学習。 J.ニューロシ。 22、10829〜10837。 オンラインで入手可能: http://www.jneurosci.org/content/22/24/10829.full
Grillon、C.、Baas、J.M.、Connwell、B.およびJohnson、L.(XNUMX)。 仮想現実環境における文脈条件付けと行動回避予測可能性の効果 Biol。 精神医学 60、752〜759。 土井:10.1016 / j.biopsych.2006.03.072
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Ham、AO、そしてWeike、AI(2005)。 恐怖学習と恐怖規制の神経心理学 Int。 J.サイコフィジオル。 57、5〜14。 土井:10.1016 / j.ijpsycho.2005.01.006
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Klucken、T.、Schweckendiek、J.、Merz、C.J.、Tabbert、K.、Walter、B. (2009) 条件付けされた性的覚醒の獲得の神経活性化:偶発性意識と性の影響 J.セックス。 中。 6、3071〜3085。 土井:10.1111 / j.1743-6109.2009.01405.x
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Klucken、T.、Wehrum、S.、Schweckendiek、J.、Merz、C.J。、Hennig、J.、Vaitl、D。 (2013) 5-HTTLPR多型は食欲調節中の血行動態反応の変化と関連している。 ハム。 ブレインマップ。 34、2549〜2560。 土井:10.1002 / hbm.22085
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Koch、M.(1999)。 驚愕の神経生物学 プログレ ニューロビオール. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Koch、M.、Schmid、A.、およびSchnitzler、H。 (1996) 驚愕の快楽減弱は側坐核の損傷によって破壊される。 神経報告 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Krohne、HW、Egloff、B.、Kohmann、C.-W.およびTausch、A.(1996)。 「肯定的および否定的影響スケジュール」(PANAS)についての考察をより深く理解することができます。 Diagnostica 42、139-156。
Kumar、P。、Waiter、G。、Ahearn、T。、Milders、M。、Reid、I。、およびSteele、JD(2008)。 大うつ病における異常な時間差報酬学習信号 脳 131、2084〜2093。 土井:10.1093 / brain / awn136
PubMed要約 | CrossRefフルテキスト | Google Scholarの
、Lax、L.、Glanzmann、P。、Schaffner、P。、およびSpielberger、CD(XNUMX)。 Das State-Trait Angstinventar。 ワインハイム:ベルトテスト。
Levy、DJ、そしてGlimcher、PW(2011)。 りんごとオレンジの比較:脳内の報酬特異的および報酬全般的な主観的価値表現の使用 J.ニューロシ。 31、14693〜14707。 土井:10.1523 / JNEUROSCI.2218-11.2011
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Lissek、S.、Levenson、J.、Biggs、A.L.、Johnson、L.L.、Ameli、R. (2008) 社会不安障害における社会的に関連のある無条件刺激に対する恐怖条件付けの高揚 Am。 J.精神医学 165、124〜132。 土井:10.1176 / appi.ajp.2007.06091513
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Martin-Soelch、C.、Linthicum、J.、およびErnst、M.(2007)。 食欲調節:神経基盤と精神病理学に対する意義 神経症。 バイオ行動。 改訂。 31、426〜440。 土井:10.1016 / j.neubiorev.2006.11.002
PubMed要約 | CrossRefフルテキスト | Google Scholarの
McDannald、MA、Lucantonio、F、Burke、KA、Niv、Y、およびSchoenbaum、G(2011)。 腹側線条体および眼窩前頭皮質はどちらもモデルベースであるがモデルフリーではない強化学習に必要である。 J.ニューロシ。 31、2700〜2705。 土井:10.1523 / JNEUROSCI.5499-10.2011
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Milad、MR、およびQuirk、G。(2012)。 並進神経科学のモデルとしての恐怖の絶滅10年の進歩 アンヌ。 サイコル牧師。 63、129〜111。 土井:10.1146 / annurev.psych.121208.131631
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Pavlov、IP(1927)。 条件付き反射:大脳皮質の生理活性の調査。 ロンドン:オックスフォード大学出版局。
Prévost、C.、Liljeholm、M.、Tyszka、JM、およびO'Doherty、JP(2012)。 ヒト扁桃体サブ領域内の特定および一般的なパブロフから器具への移動の神経相関:高解像度fMRI研究。 J.ニューロシ。 32、8383〜8390。 土井:10.1523 / jneurosci.6237-11.2012
PubMed要約 | CrossRefフルテキスト | Google Scholarの
Tranel、D。、およびDamasio、H(1994)。 皮膚皮膚コンダクタンス反応の神経解剖学的相関 精神生理学 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
PubMed要約 | CrossRefフルテキスト | Google Scholarの
キーワード:古典的条件づけ、報酬、罰、驚愕反射、皮膚コンダクタンス反応
引用:Andreatta MとPauli P(2015)人の食欲対嫌悪条件付け。 前面。 ふるまい。 神経症. 9:128 土井:10.3389 / fnbeh.2015.00128
受け取ったもの:11 2月2015。 受け入れられる:05 5月2015。
公開日:19 5月2015。
によって編集:
ニルス・ビルバウマードイツ、テュービンゲン大学
したレビュー:
ティム・クルッケンドイツ、ギーセン、Justus Liebig大学
アレッサンドロ・アングリーリ、パドバ大学、イタリア
著作権©2015 AndreattaとPauli。 これは、の条件の下で配布されているオープンアクセスの記事です。 クリエイティブコモンズ表示ライセンス(CC BY)。 他のフォーラムでの使用、配布または複製は、原著者またはライセンサーに認められ、本ジャーナルの原著論文が認められた学術慣習に従って引用されていることを条件として許可されます。 これらの条件に従わない使用、配布、複製は許可されていません。
*対応:マルタ・アンドレッタ、心理学科(生物心理学、臨床心理学、心理療法)、ヴュルツブルク大学、マルクス通り9-11、D-97070ヴュルツブルク、ドイツ、 [メール保護]