 マルクス・J・ランタラ
マルクス・J・ランタラ セヴェリ・ルオト
セヴェリ・ルオト タッジャナ・クラマ
タッジャナ・クラマ インドリス・クラムス
インドリス・クラムス- 1フィンランド、トゥルクのトゥルク大学生物学部
- 2英語、演劇、ライティング研究、オークランド大学、オークランド、ニュージーランド
- 3ニュージーランド、オークランド、オークランド大学心理学部
- 4ラトビア、ダウガフピルス大学、バイオテクノロジー学部
- 5タルトゥ大学、タルトゥ、エストニアの生態地球科学研究所
摂食障害は進化的に新しい病気です。 それらは、すべての精神障害の中で最も高い死亡率のいくつかにつながります。 摂食障害についてはいくつかの進化仮説が提案されていますが、 性内競争仮説 証拠によって広く支持されています。 私たちは提示します ミスマッチ仮説 摂食障害の現在の理論的枠組みへの必要な拡張として。 この仮説は進化的に小説を説明します 適応メタ問題 交配の動機が、大規模で容易に入手できるが、報酬が高いが肥満を引き起こす食物と対立するときに生じた。 この状況は、特に座りがちなライフスタイル、常に存在するジャンクフード、カロリーの余剰、およびソーシャルメディアを介して行われる社会比較の遍在によって特徴付けられる現代の環境で悪化しています。 私たちの精神神経免疫モデルは、交尾動機と食物報酬の間の適応的メタ問題がどのように慢性的なストレス、さらには摂食障害につながるかを示すことにより、究極レベルの因果関係を近接メカニズムに結び付けます。 慢性的なストレスは神経炎症を引き起こし、これは通常摂食障害と共起するOCDのような行動に対する感受性を高めます。 慢性ストレスはセロトニン系をアップレギュレートし、神経性食欲不振症患者に不快気分を引き起こします。 しかし、ダイエットはセロトニンレベルと不快気分を低下させ、厳しいダイエットを通じてコルチゾールと神経炎症を増加させる悪質なセロトニン作動性恒常性ストレス/飢vサイクルにつながります。 私たちの精神神経免疫モデルは、摂食障害の個人間および個人内変動が、腸内微生物叢の(共)変動とストレス応答性から部分的に生じ、神経炎症とセロトニン系に影響することを示しています。 摂食障害の最善の治療方法を理解するために近年行われた進歩をレビューし、将来の臨床研究の方向性を概説します。 現在のエビデンスは、摂食障害の治療は、慢性的なストレス、神経炎症、ストレス応答性、および障害の原因となる腸の共生を減らすことを目的とすべきであることを示しています。 究極の原因を近接するメカニズムと結び付け、生物心理社会的治療を行う 原因 明らかな症状ではなく、摂食障害に苦しむ何百万人もの人々に対して、より効果的で洗練された長期的な介入をもたらすことが期待されています。
イントロダクション
摂食障害は重度の精神障害であり、生物心理社会的病因とEUだけで年間約1兆ユーロの費用がかかります(シュミット他、2016)。 彼らは慢性的で衰弱する可能性があり、死亡率の大幅な増加に関連付けられています(シュミット他、2016)。 たとえば、神経性食欲不振は、すべての精神障害の中で最も高い死亡率を示しています(罹患した5.10人あたり1,000人の死亡者: Arcelus et al。、2011)。 摂食障害の治療は、他の精神障害の治療と比較して効果がないことはよく知られています(Arcelus et al。、2011; マレーら、2019)。 この非効率性は、現在の治療が摂食障害の根本的な原因を治療するのではなく、症状を軽減することに焦点を合わせているという事実によって強調されています。 既存の治療法が無効である主な理由は、率直に言って、摂食障害の病因が適切に理解されていないことです(van Furth et al。、2016; Frankら、2019; マレーら、2019)。 摂食障害の科学的知識の進歩が緊急に必要です。
特性または行動の完全な分析は、理想的には1つの異なるが補完的なレベルで提供されます。(XNUMX) 近接メカニズム 特性の基礎:どのように機能しますか? –(2)何が 究極の理由 それは進化しました:もしあれば、それは生物にどんなフィットネスの利益を提供しますか? (Bateson and Laland、2013; ランタラら、2018; ルオトら、2019a)。 これらXNUMXつのレベルの分析を統合し、近接メカニズムと究極の原因の両方を理解しないと、摂食障害を予防し、それらの効果的な治療法を見つけることは難しいと主張します。
精神障害の診断および統計マニュアル(DSM-5)の第5版では、神経性食欲不振(AN)、神経性過食症(BN)、および過食症(BED)のXNUMXつの摂食障害が規定されています。 これらXNUMXつの障害に加えて、DSM-XNUMXは、XNUMXつの特定の摂食障害(OSFED)サブタイプを指定することにより、サブスレッショルドおよび非定型状態の重要性を認識しています。
1.非定型神経性食欲不振(すなわち、体重の少ない食欲不振の特徴);
2.神経性過食症(低頻度および/または限られた期間);
3.過食症(低頻度および/または限られた期間);
4.障害の除去。
5.夜食症候群。
DSM-5には、これらXNUMXつのカテゴリのいずれにも当てはまらない人、または特定のOSFED診断を下すのに十分な情報がない人を含む不特定摂食障害(UFED)と呼ばれるカテゴリも含まれます(アメリカ精神医学会、2013).
摂食障害を説明するために、いくつかの進化仮説が提案されています。 これらの究極レベルの仮説(セクション「摂食障害の既存の進化心理学的仮説」)を批判的にレビューし、摂食障害の根底にある生理学的メカニズムの新しい近似説明(セクション「摂食障害の精神神経免疫モデル」)でそれらを統合します。 私たちの精神神経免疫学的モデルは、摂食障害は別個の病気ではなく、連続体を形成することを示唆しています。 患者の生体行動状態の変化に基づいて、連続体モデルは、「不特定の摂食障害」が一般的な診断である理由を説明します(ケースの11〜50.8%: マチャドら、2013; Caudle et al。、2015; Mancuso et al。、2015)および患者の診断が時間の経過とともに摂食障害間で移行する理由(セクション「性内競争に対する反応の個人差の原因」)。 このモデルは、摂食障害が他の精神障害と共存することが多いという発見の説明を提供します(セクション「摂食障害の併存」)。このモデルによると、この共有伝達は、神経炎症およびストレス応答性に対する脆弱性によって媒介されます(セクション「リスク要因」摂食障害」)。 私たちの精神神経免疫学的モデルは、近接メカニズムと究極の原因の両方の総合的な理解に基づいた治療法(「精神神経免疫学に基づく摂食障害の治療」および「摂食障害の究極レベルの予防」のセクション)を提案します。 これらの治療法は、現在の摂食障害の治療に大きな進歩をもたらす可能性があります。これについては、「現在の摂食障害の治療」セクションで説明します。
摂食障害に対する既存の進化心理学的仮説
摂食障害の究極の原因には、1つの既存の進化仮説があります。 BNおよびBEDの主要な進化仮説は(XNUMX) rif約遺伝子型仮説。 過食は心理的適応であることを示唆している(例えば、 ルイスら、2017 余分なエネルギー貯蔵が私たちの種の進化の歴史において保護的だったために生じた心理的適応の議論のために:それらは栄養失調の回避を助け、飢amine中の生存を助け、生殖を制御しました(チャクラバルシーアンドブース、2004; ウェルズ、2006)。 この仮説を拡張して、 二重介入ポイント モデルは、体が体脂肪症のレベルの上限と下限の設定点を持っていると仮定します。 これらを超えると、生理学的フィードバックメカニズムがトリガーされます(スピークマンら、2011; スピークマン、2018)。 飢positを避けるために肥満の最小設定値が必要である一方、最大設定値は捕食のリスクによって決定されます。 捕食のリスクが低下するにつれて、より高い最大設定値をコードする遺伝子がより一般的になり、体重増加を防ぐためにカロリー摂取量を減らす人は少なくなります(スピークマンら、2011).
(2) 性内競争仮説 (Abed、1998)摂食障害の究極の原因は、配偶者に対する激しい性内競争であることを示唆しています。 この仮説は、女性の体型は、女性の生殖歴、生殖能力、および配偶者価値の指標であり、ウエストヒップ比およびボディマス指数(BMI)(アンドリュース他、2017; デル・ゾットとペグナ、2017)。 女性が老化および/または生殖すると、体重が増え、砂時計の体型が失われる傾向があります(ブトフスカヤ他、2017)男性にとって性的に望ましい特性です(例: ボベット、2019)。 女性の生殖窓は有限であるため、男性は出生率と若さの手がかりを好むようになりました(ソン、2016; ラセクとゴーリン、2019)。 これは、若々しくスリムに見えることにより、男性の注意をめぐって競合する女性につながる可能性があります:結局のところ、女性の生殖価値は若々しさに関連し、若々しさはスリムに関連しています(Abed、1998; ラセクとゴーリン、2019).
性内競争の仮説は、女性の身体に対する男性の選好が環境や社会によって異なる可能性があることを示す研究と一致しています(cf. ファーナムとバグマ、1994; Tovee et al。、2006)。 ふっくら感は、栄養失調が一般的な国での出生率が高いことの指標になる可能性があります。 対照的に、十分に栄養を与えられた集団では、相対的なふっくら感が加齢と出生率の低下に関連しています(例、 Tovee et al。、2006)。 性内競争の仮説は、西洋社会における摂食障害の有病率の増加は、女性間の性内競争の激化および/または相対的な豊富な食物(Abed et al。、2012; Baumeister et al。、2017; Nettersheim et al。、2018).
性内競争を激化させる多くの要因があります(Abed et al。、2012):(a)妊t性の低下は、年配の女性の魅力的な外観の保存の増加につながります。 (b)現代の西部社会では、女性は生殖行動を規制する能力がより高い(親族からの干渉が最小限である)。 (c)人類の祖先の状態と比較して、近代的な都市では、若くて若々しい女性、すなわち潜在的な競争相手が異常に多い。 (d)メディアは魅力的な競合他社の画像を提供します。 (e)食物は豊富で、人口は十分に栄養が与えられているので、体重増加と乳形の悪化は、加齢に伴う典型的な特徴です。 (f)結婚生活の不安定化と離婚のthe延により、男性と女性の両方が交配市場に繰り返し戻ってきた。 (g)現代医学の出現と平均余命の増加(すなわち、閉経後女性の増加)により、若者は女性の配偶者価値の主要な決定要因のXNUMXつになりました( Abed et al。、2012も参照してください Baumeister et al。、2017; サンダースとイートン、2018; ラセクとゴーリン、2019; ルオト、2019a).
26か国で7,000人以上の参加者が行った研究では、社会経済的地位の高い地域では薄い体型が好まれ、メディアへの露出は理想体重と有意な関係があることがわかりました(スワミ他、2010) さらに、 スワミ他 (2010) 女性は、男性の考えよりも薄い女性の方が魅力的だと一貫して考えていることがわかりました。 Baumeister et al。 (2017) 地元の交配市場で男性が不足していると認識する女性が増えれば増えるほど、thinせたいと思うようになり、身体の不満の兆候が多くなると報告しています。 これらの発見は、女性間の性内競争が女性のthin身追求を促進するという考えを裏付けています。
摂食障害の背後にある性内競争の仮定された役割に対するより多くのサポートは、男性のボディービルダーに影響を与える「逆食欲不振」から来ています(ポープら、1993)そして、より一般的に男の子と男性の身体の不満を駆動することができます(Karazsia et al。、2017)。 影響を受けた個人は、筋肉質であるにも関わらず小さすぎるという信念を表明しているため、身体イメージが歪んでいます。 筋肉質と大きな体の大きさは、他の哺乳類と同様に、人間のオスとオスの競争において明確な利点を提供します。 筋肉質は、私たちの進化史において女性にとって性的に魅力的な特性であった可能性があり、現在の男性でさえ性的選択が特性に作用している(フレデリックとハセルトン、2007; Sellら、2017).
上記の激化する競争と他の進化的に新しい要因により、西洋世界のほとんどの女性は体の大きさと形に不満を抱いており、XNUMX代の少女の半分が体重をコントロールしようとしています(ノイマルク=シュテナー、2005)。 実験的研究は、競合他社の魅力や薄さに関連する手がかりがなくても、性内の状態の動機が若い女性の摂食障害に似た摂食態度を引き起こすことができることを示しました(Liら、2010; カステリーニら、2017)。 同様の効果は異性愛者の男性では見られません(Liら、2010)。 さらに、摂食障害は異性愛者の男性よりも同性愛者の男性の間ではるかに一般的です(Liら、2010; カルゾら、2018)。 これらの調査結果の潜在的な説明は、同性愛者の男性内での性的競争は身体的魅力に焦点を当てていることです。Liら、2010)。 同性愛者の男性は、性的欲求の競合に対しても、否定的な食事態度と自分の身体イメージについてのより悪い認識で反応します(Liら、2010).
性内競争が摂食障害の発症の重要な要因である場合、特に交配に関連する社会的地位の達成を志向している人は、摂食障害のリスクが高くなります。 実際、摂食障害は、性内競争が最も強い年齢の頃に最も頻繁に引き起こされます(Liら、2010)。 さらに、女子学生の割合が高い学校の女子は、摂食障害を発症する可能性が高いことが示されています(ボールドら、2016)、これは、より高い性内競争環境が摂食障害の有病率を高めることを示唆しています(cf. Baumeister et al。、2017; サンダースとイートン、2018).
摂食障害は、多くの場合、友情グループで社会的に伝染しやすく、学校環境に広がる可能性があります(ボールドら、2016)。 たとえば、摂食障害のために友人のBMIが低い場合、自分の体が比較的大きいと感じ、身体の不満が高くなり、摂食障害を発症する可能性が高くなります。 摂食障害のこの社会発達の病因は、低体重の少女の割合が高い学校では、他の少女が体重を減らそうとする可能性が高いという発見によって裏付けられています(Muellerら、2010).
(3)など、経験的サポートが少ない他の仮説もあります。 生殖抑制仮説、これはANが罹患女性による生殖抑制の適応的試みであることを示唆しています(ワッサーとバラッシュ、1983; Surbey、1987; Voland and Voland、1989)。 (4) 親の操作仮説 ANは血縁選択によって維持されることを示唆している:親は女性の子孫を操作して、兄弟間の生殖投資における戦略的シフトを促進する(Voland and Voland、1989)。 (5) 支配的な女性仮説による生殖抑制 ANは、女性と女性の生殖競争の過程における支配的な女性による下位の女性の生殖抑制の発現であると仮定します(ミーリー、2000)。 (6) 飢amine仮説から逃げる ANの症状(多動や摂食制限など)により、飢amineがより豊富な食物のある地域に到達するための移動を可能にしたことが示唆されていますGuisinger、2003).
これらの仮説はANに焦点を当てており、他の摂食障害、特にBEDについては説明がありません。 仮説3–5では、男性でも同様に摂食障害が発生する理由を完全に説明できていません。 また、性的指向が摂食障害の可能性に影響する理由を説明できません(Liら、2010; カルゾら、2018)。 既存の研究では、神経性食欲不振症の個人は「敗北戦略」を採用する社会的従属者であるという考えを支持していません(Faer et al。、2005)。 上記の仮説のほとんどは、摂食障害は進化的適応であるという考えに基づいています。
他の仮説とは対照的に、性内競争仮説は、摂食障害が適応であるとは想定していません。 さらに、ANのみに適用されるわけではありませんが、摂食障害の全範囲を、女性の性内競争への適応と、それらの適応が失敗する現代の環境との不一致の病理学的結果とみなしています。 性内競争仮説の説明力にもかかわらず、仮説に関する先行研究は、摂食障害の進化的起源を完全に説明するには不十分に定式化されています。 したがって、 摂食障害のミスマッチ仮説.
摂食障害の不一致仮説
大規模肥満は進化の目新しさです。 人類の文化的進化は、大量の エネルギー的に高密度 (NAIST) および 不快なほどの報酬 先進国ではほとんどの人が食物を容易に入手できます(リンデバーグ、2010; パワー、2012; ロジンとトッド、2015; Corbett et al。、2018)。 環境からエネルギーを抽出することは、先進社会に住むほとんどの現代人にとって実質的なエネルギーコストを必要としません。 現代の先進国が享受している現在のエネルギーの豊富さは、進化の目新しさです。先祖の人間は、現代の人間よりも食料資源を獲得するために(平均して)より多くのエネルギーを費やすことを余儀なくされました。 この単純なエネルギーの不均衡(カロリーイン>カロリーアウト)は、肥満の蔓延と現代の健康問題の広がりをもたらしました(リンデバーグ、2010; パワー、2012; Corbett et al。、2018)、メンタルヘルス(Milaneschi et al。、2018; ランタラら、2018).
人間には、食物摂取の原因となる進化した心理的メカニズム(モジュール)の洗練されたスイートがあります(キング、2013; アルシャワフ、2016; ロール、2017; 愛とスリコウスキー、2018)および交配を担当する別の一連のメカニズム(モジュール)(Weekes-Shackelford and Shackelford、2014; ルオト、2019a, b)。 相対的なエネルギー量の現在の環境(リンデバーグ、2010; パワー、2012)食物摂取と交配に関与する心理学的モジュール間に進化的に新しい対立を生み出した。 一方で、人間は食料の存在を最大限に活用するように進化しています(チャクラバルシーアンドブース、2004; キング、2013; アルシャワフ、2016); 一方、人間は表現型の性的装飾を介して生殖能力を示すように進化しています(杉山、2015; ラセクとゴーリン、2019)。 文化的進化は、人間の進化史上初めて、これらの心理的適応が互いに大規模な矛盾を起こす状況を作り出しました。 したがって、 摂食障害のミスマッチ仮説 食物摂取と交配の以前に共適応した心理的メカニズムが拮抗するという新しい状況を認識します。 この拮抗は、対立するインセンティブ、つまり食物の報酬と交配の報酬の間で個人が引き裂かれる状況を作り出します。 矛盾する適応問題の同時提示は、 適応メタ問題 (アルシャワフ、2016)。 カロリーが高く、感覚的に報酬の多い食物が豊富であるという基本的な拮抗作用(リンデバーグ、2010; ロジンとトッド、2015)交尾の動機と食物の報酬の間に引き起こされたものが、現代の人間におけるそのような適応的メタ問題を引き起こし、最終的にはさまざまな摂食障害に現れます。
ミスマッチ仮説は、摂食障害が現代の先進社会と同じように伝統的な狩猟採集社会でも同様にvalent延していることを示すことによって偽造される可能性があります。 狩猟採集者の自給スタイルは、進化的順応性の人間環境の条件にある程度匹敵します(たとえば、 アルシャワフ、2016; ルイスら、2017)。 私たちは、狩猟採集社会におけるAN、BN、BEDの存在に関する証拠を認識していません。逆に、飢erは現代の狩猟採集社会の広範な側面のようです( アルシャワフ、2016).
ミスマッチ仮説は、人間以外の動物の研究によって間接的に支持されており、人間が動物を飼育下に置いている場合にのみ肥満が重大な問題になることが示されています(パワー、2012)。 捕われの身は、現代の座りがちな生活様式が人間に対して行うように、人間以外の動物にとって進化的に類似した条件を表します(ウィリアムズ、2019)、両方の状況下での肥満表現型の有病率の大幅な増加につながる(パワー、2012)。 これらの発見は、性的競合仮説への必要な拡張として提案する進化的ミスマッチ仮説の文脈における摂食障害のフレーミングの有用性を強調しています。 摂食障害に加えて、ミスマッチ仮説は、2型糖尿病、冠動脈疾患などのいくつかの非感染性疾患の現代的な流行についても説明しています(Corbett et al。、2018)および他の多くのメンタルヘルスの問題(Liら、2018; ランタラら、2018).
摂食障害の精神神経免疫学的モデル
薄さのための性内競争と、感覚的に価値がありカロリーが高い食品の豊富さから生じる適応性の問題キング、2013; ロジンとトッド、2015)先進社会に住む女性のinせを追求するためのもっともらしい究極の説明を提供しているようです。 しかし、これらの仮説は、なぜ女性の一部と同性愛の男性だけが摂食障害を発症するのかを説明していません。 さらに、仮説では、なぜ一部の人々が体重を減らすという強い執念を抱いて死ぬほど飢えているのか、他の人々は過食して過体重になるのかを説明していません。 仮説では、非脂肪恐怖ANの存在を説明することもできません(「自己免疫と摂食障害」のセクションを参照)。
科学の進歩は、理論と経験的証拠の間の良好な適合に依存しています(Mathot and Frankenhuis、2018)。 現時点では、進化精神医学の理論と摂食障害に関する臨床的証拠との間にこの適合性が欠けています。 したがって、摂食障害の個人間および個人内の変動を説明する近似メカニズムの存在を仮定し、理論と経験的知見の適合性をさらに改善します。 (1)性内競争の激化が女性のごく一部で摂食障害につながるという発見を説明する新しいモデルを提案します。 (2)女性のこのサブセットは、極端なthinせと肥満の反対の表現型の結果を伴うさまざまな摂食障害を発症する可能性が高い。 (3)患者の診断は、時間の経過とともに摂食障害に移行する可能性があります。
摂食障害および強迫性障害
診断基準によると、摂食障害では身体運動、外見、食物への執着が一般的です(アメリカ精神医学会、2013)。 これらの強迫観念は、感情的な不快感につながり、体重のチェック、運動、パージ、断食などの一連の行動の発達につながります。 摂食障害のこれらの古典的な症状に加えて、疑い、確認、対称性と正確性の必要性など、他の多くの強迫性特性は、精神医学的コントロールグループよりもBNおよびAN患者ではるかに一般的です(Cassidy et al。、1999)。 摂食障害の患者の中には、身体の一部に繰り返し触れる、鏡で自分の体の形を見るなど、視覚的または触覚的チェックの儀式を持っている人がいます(Legenbauer et al。、2014)。 したがって、摂食障害患者の行動は、OCDの行動と多くの類似点を持っています(バスティアニら、1996; ガルシア・ソリアーノ他、2014)。 OCDと診断された19,814人の参加者とAN(8,462%の男性)と診断された6.4人の参加者を含むスウェーデンの多世代家族と双子の研究では、OCDの女性はANの16倍の診断を受け、OCDの男性は37倍のリスク増加(Cederlof et al。、2015)。 ANおよびBNは、完全主義や神経症など、OCDにリンクされた人格特性にも関連付けられています(Cassidy et al。、1999; Anderluh et al。、2003; Halmi et al。、2005; アルトマンとシャンクマン、2009)。 さらに、ANは、一致したコントロールの親族と比較して、OCDのある個人の罹患していない親族でより一般的であり、遺伝的リスク因子の共有を示唆しています(Kayeら、1993)。 したがって、GWASメタ分析により、AN表現型とOCD表現型の間の遺伝的相関が見つかりました(Anttila et al。、2018).
最近のポジトロン放出断層撮影(PET)の研究では、OCD患者の神経炎症が見つかりました。 特に、彼らは彼らの脳のミクログリア活性を高めた(アトウェルズら、2017)。 強迫行動の防止に関連する苦痛は、眼窩前頭皮質の神経炎症と強く相関しています(アトウェルズら、2017)。 神経炎症が生化学的事象のカスケードを引き起こし、その結果、神経ホルモン、神経ペプチド、神経伝達物質の調節不全が起こり、OCD症状を引き起こす可能性があります。 ただし、先行研究(アトウェルズら、2017)OCD患者が神経炎症を起こす理由を説明できませんでした。
OCDの強迫観念は患者に大きなストレスを引き起こしますが、ストレス自体はOCDの発症にも重要な役割を果たすようです(Toro他、1992; Behl et al。、2010; Adamsら、2018)。 ストレスはOCD症状を引き起こし、その頻度と重症度を増加させます(Findley et al。、2003)。 非ヒト動物での実験的研究は、ストレスが神経炎症を増加させ、ミクログリア活性を高めることを示しました Calcia et al。、2016)。 したがって、慢性ストレスは、OCD表現型で発生する神経炎症の原因となる可能性があります。 ストレスとHPA軸の活性化はOCDでこのような重要な役割を果たしているため(Sousa-Lima et al。、2019)、ストレスや摂食障害の症状と同様の関連性が期待できます。
摂食障害とストレス
BNとANを持つ個人は、「美の理想」を満たすために体重を減らし、薄さのための性内競争に固執しようとしています(cf. Abed、1998)。 ANおよびBN患者は、時間とともに慢性化する高ストレスホルモンレベルによって競合に応答します( Soukup et al。、1990; Rojo et al。、2006)。 ストレスは、「完璧な体型」を持つために仲間や社会の圧力によって引き起こされる可能性があります(カステリーニら、2017)、自分の自己イメージに対する恥と罪悪感は、ストレスの悪循環を個人に継続させる可能性があります。 ANの患者の中には、体重についての否定的なコメントがANの引き金となる出来事であると遡及的に特定している人もいます(Dignon et al。、2006)。 特に、低体重が競争要因であるスポーツでは、薄さの要件が摂食障害を引き起こす可能性があります(Joy et al。、2016; Arthur-Cameselle et al。、2017)。 同じことがファッションの世界、ダンス、バレエにも当てはまります(マルケス、2008)。 スポーツやファッションで体重を減らすという要件は、身体の不満や社会的ストレスを引き起こす可能性があります(cf. カステリーニら、2017).
ANおよびBNの患者を対象としたレトロスペクティブ研究により、摂食障害の他の1つのトリガーイベントが特定されました。病気/入院および(2)虐待、性的暴行または近親相姦(Berge et al。、2012)。 これらすべてのトリガーイベントに共通するのは、ストレスを増加させることが知られていることです。 したがって、DSM-5は、ANの発症はしばしばストレスの多い人生の出来事に関連していると述べています(アメリカ精神医学会、2013).
慢性ストレスは免疫系をアップレギュレートすることが知られています( スタントンら、2018; ローレダー、2019)。 人間および他の動物での研究は、社会的ストレッサーが、低悪性度の末梢炎症および神経炎症を促進する炎症性サイトカインの産生の特に強力な引き金であることを示しています。 人間の社会的拒絶は、腫瘍壊死因子-α(TNF-α)およびインターロイキン-6(IL-6)のレベルの増加と関連しています(Slavich et al。、2010)。 したがって、23の研究を含むメタ分析では、AN患者のTNF-αおよびIL-6のレベルが大幅に増加していることがわかりました。Daltonら、2018)。 しかし、研究では減量のためにIL-6の増加が起こる可能性を排除することができませんでした。 IL-6は脂肪分解を刺激するため(Wedell-Neergaard et al。、2019)、IL-6のレベルの増加が栄養失調または炎症、あるいはその両方によって引き起こされるかどうかは明らかではありません。 それにもかかわらず、 ダルトン等。 (2018) ANの患者もIL-15レベルが上昇していることがわかった。 IL-15は神経炎症に関連しています(パン他、2013)、ANと神経炎症の間のリンクを示唆しています。
AN患者と同様(Solmi et al。、2015; Daltonら、2018)、OCDを持つ個人はまたTNF-αおよびIL-6のレベルが増加しています(Konuk et al。、2007)。 ANおよびBN患者はしばしばOCDと診断されるため(そして体重を減らすことは彼らにとって強迫観念となるため)、OCDの場合と同様に、慢性ストレスによって引き起こされる神経炎症はANおよびBNの根底にあると仮定します(cf. アトウェルズら、2017)。 この仮説の間接的な証拠は、ANとBNの患者の74%が片頭痛に苦しんでいるという観察から得られます(ブリューワートンとジョージ、1993; Brewertonら、1993; D'Andrea et al。、2009)、これは神経炎症性疾患です(マルホトラ、2016)。 飢starはストレスホルモンレベルを増加させることが知られているため(Naisbitt and Davies、2017)、自己誘発飢starは、AN患者の体重減少に対するストレス誘発性の強迫観念を強化する可能性があるようです。 このフィードバックループは、停止するのが難しく、生命にかかわるレベルまでエスカレートする可能性のある悪循環を引き起こす可能性があります。 興味深いことに、AN患者で上方制御されるストレスホルモンコルチゾール( Soukup et al。、1990; Rojo et al。、2006)は、人間の糖新生を増加させるホルモンのXNUMXつです。 糖新生は、体内でエネルギーとして使用されるタンパク質または脂肪から体内のグルコースを合成するプロセスです(Khani and Tayek、2001)。 ストレスホルモンレベルを増加させることにより、糖新生は、AN患者の食事中および飢ation中の神経炎症を増加させる可能性があります。
ヒトおよび他の動物の実験的研究により、炎症誘発性サイトカインは食欲を減退させ、病気による食欲不振を引き起こす可能性があることが示されています(ダンツァー、2009)。 食欲の動機付けと食物の快楽的価値を処理する中脳辺縁系報酬システムは、AN患者では健康なコントロールや他の摂食障害のある患者ほど効果的に機能しません(Ceccarini et al。、2016)。 したがって、食事は、影響を受けていない個人の場合と同じように、AN患者にとって快楽的な経験とはならないかもしれません(cf. スタントンら、2018)。 この快楽の低下は、AN患者のダイエットの効率に一部貢献しているかもしれませんが、ほとんどの健康的なダイエットをする人は努力に失敗します( マン他、2007).
過食とストレス
BNおよびANの場合と同様に、多くの場合、BEDは薄さを求める性内競争によって引き起こされると仮定しています。 この仮説の間接的な証拠は、BED患者が低い自尊心、一般的な身体不満を持っていることを示す調査結果によって与えられます(パール他、2014)および心理的苦痛の上昇(カステリーニら、2017; Mustelin et al。、2017)。 彼らは自分の体重を過大評価し、負の光で体の形を見る傾向があります(パール他、2014)。 減量を意図しているにもかかわらず、BED患者は大食いになり、体重が増えて、しばしば肥満になります。 カステリーニ等。 (2017) 過食症は、非臨床的女性集団における機能障害の身体イメージの尊敬と性的苦痛の増加に関連していることが報告されており、BEDは薄さのための性内競争によって引き起こされるという仮説をさらに裏付けています。 この仮説は、BEDの症状がその原因ではなくBEDの影響であることを示すことによって挑戦することができます。 しかし、正常体重のBEDと肥満BEDの個人を比較した研究では、正常体重の人は肥満の人よりも体重を減らす衝動が強いことがわかりました(ゴールドシュミット他、2011)。 形状や体重の過大評価にグループ間差はなく、これらの症状は共存する肥満によって引き起こされたものではないことを示唆しています(ゴールドシュミット他、2011).
多くの人がストレスを感じると食欲を失いますが、軽度の心理的ストレッサーやネガティブな感情的エピソードでさえ、BEDまたはBN患者の過食を引き起こす可能性があります(Masheb et al。、2011)。 通常、ストレスは交感神経系と身体の戦闘または飛行反応を活性化します。 これらの状況下では、副腎皮質刺激ホルモン放出因子(CRF)は、消化器系に影響を与え、空腹感を減少させることにより、食欲を抑制します。 これが、BEDを有する個人が急性ストレス中に激しい欲求と過食を経験しない理由ですが、自宅のプライバシーと、急性ストレス要因がおさまった後の一人でいるときです(Masheb et al。、2011)。 過食はネガティブな嫌悪的な感情状態から「逃げる」方法と見ることができます(バートンとアボット、2019).
BEDとBNの患者には、体重を減らしたいという明確な願望があり、したがって、なぜ彼らが過食に抵抗するのがそれほど難しいのかを理解することが重要です。 理由は彼らのダイエット習慣にあるかもしれません。 カロリー制限されたラットでは、被験者が砂糖と脂肪が多い食品を食べる機会がある場合、心理的ストレスが過食を引き起こすことが示されています(Haganら、2002, 2003)。 同様に、カロリー制限を伴うフットショックストレスは、ラットに通常量のXNUMX倍の食物を消費させます(Boggiano他、2005)。 実験的にストレスを与えられた食物制限ラットは、食物摂取を直接的または間接的に調節する別個の脳領域で炎症を発症します。 これらのラットはまた、過食症のような摂食行動を発症します(Alboni et al。、2017)。 それに対応して、人間の被験者では、非常においしい食物が利用できる場合、心理的ストレスが健康的なダイエット中の暴食を引き起こす可能性があります(Oliver and Wardle、1999も参照してください カステリーニら、2017; Klatzkin et al。、2018).
患者の心理生理学的状態の評価は、BEDについてのさらなる洞察を提供します。 BED患者は、コントロールよりも高いストレス応答性を示します(Klatzkin et al。、2018)。 BED患者は、ストレス応答性が高いため、コントロールよりも簡単に過食症になりやすい場合があります(cf. Klatzkin et al。、2018)。 ストレス応答性が高い理由のXNUMXつは、内臓脂肪組織による炎症(Shields et al。、2017; Krams et al。、2018; ローレダー、2019)。 心理社会的ストレッサーはほとんどの人の生活に存在しますが、自己調節能力はストレスによって頻繁に引き起こされる負の健康結果に対して個人を緩衝します(Evans and Fuller-Rowell、2013; Shields et al。、2017)。 しかし、蓄積された証拠は、炎症が自己調節不全を促進する広範な生物学的行動変化を引き起こす可能性があることを示唆していますShields et al。、2017)。 BED患者は、体重が一致したコントロールよりも88%高い感度のCRP値を示しており、BED患者の体に重度の炎症があることを示唆しています(Succurro et al。、2015)。 したがって、末梢炎症は、自己調節能力を低下させる可能性があります(Shields et al。、2017)BED患者では、ストレス応答性をさらに高めます。 これは、免疫細胞または脂肪細胞によって産生される炎症誘発性サイトカインがHPA軸を刺激することが知られているためです(ヤウとポテンザ、2013)。 この機構的リンクは、肥満につながる悪循環を引き起こす可能性があります(cf. Shields et al。、2017; Milaneschi et al。、2018)そして、私たちが提案するように、BEDに。 末梢炎症は、気分障害がBED患者の間でそれほど一般的である理由を説明する潜在的な原因メカニズムです。たとえば、炎症は、適応性のある気分変化が不適応な臨床的うつ病に変わる可能性を高めます(cf. ルオトら、2018; ランタラら、2018)。 遺伝的要因は、これらの障害の併存疾患をさらに増加させる可能性があります(「摂食障害のリスク要因」のセクションで説明)。
神経性食欲不振症および神経性過食症の神経化学
セロトニン(5-ヒドロキシトリプトファン)は、衝動制御、強迫観念、気分、食欲に影響を与えることが知られています(Bailer and Kaye、2011; Dalley and Roiser、2012; Garcia-Garcia et al。、2017)。 セロトニン作動性活動を上方制御する治療は食物消費を減らす傾向がありますが、セロトニン作動性活動を下方制御する治療は食物消費を増加させ、体重増加を促進します Bailer and Kaye、2011も参照してください Alonso-Pedrero et al。、2019)。 AN患者に関する研究では、セロトニン系の機能障害が報告されています( Bailer and Kaye、2011; Riva、2016)。 興味深いことに、IL-15はAN患者で上方制御されています(Daltonら、2018)、およびマウスに関する研究により、IL-15がセロトニン作動性システムを上方制御することが示されています(Wuら、2011; パン他、2013).
ANの急性期(個人が低体重の場合)では、患者は脳脊髄液中のセロトニン代謝産物のレベルが健康なコントロールよりも著しく低い(Kayeら、1984, 1988)。 彼らはまた、セロトニン活性を有する薬物に対するプロラクチン応答を鈍らせており、 3H-イミプラミン結合、さらにセロトニン作動性活性の低下を示唆(Bailer and Kaye、2011)。 セロトニンはトリプトファンと呼ばれるアミノ酸から合成されるため、食物から得なければならない必須アミノ酸であるため、病気の急性期のAN患者におけるセロトニン代謝の低さのもっともらしい説明は、飢star /ダイエットから生じるということです(Kayeら、2009; ハリーム、2012)。 対照的に、ANから回復した人はセロトニンレベルが上昇しています(Kayeら、1991)。 実験的研究では、食餌性トリプトファンを減らすと、ANの女性の不安と気分が低下するが、対照の女性には影響がなかった(Kayeら、2003).
AN患者は、病前および回復後の両方で、高レベルの不安、強迫観念および危害回避を有することが知られています。 また、病的前にセロトニンのレベルが高くなり、不快な状態になる場合があります(Bailer and Kaye、2011). ケイ等。 (2009) ダイエット/飢starは、脳内のセロトニン作動性活動を低下させることにより、AN患者の気分を良くすることを示唆しました。 これらの個人はまた、より薄い外観について仲間から肯定的なフィードバックを得る場合があり、それはさらに飢starを続ける動機付けとなる。 飢byによって引き起こされるトリプトファンの枯渇の結果として、脳はセロトニン受容体の数を増やすことで応答し、残りのセロトニンをより効率的に利用します(Kayeら、2009)。 これは悪質な恒常性サイクルにつながります(図1)、気分が良くなるために、AN患者はトリプトファンをさらに減らす必要があり、食物消費の削減につながるため(Kayeら、2009)。 患者がトリプトファンを含む食物を食べ始めると、セロトニンのレベルが急激に上昇し、極度の不安と感情的な混乱を引き起こします(Kayeら、2009)。 これにより、AN患者の回復が非常に困難になります(Kayeら、2009)。 トリプトファンの不足による病気の急性期のセロトニンレベルの低下(Riva、2016)ANで典型的な深刻なボディイメージ障害を説明できます。 そのような障害を引き起こす正確な神経生理学的メカニズムは知られていないが、そのメカニズムはおそらくうつ病でしばしば見られる低い自尊心に類似している(cf. Orth and Robins、2013).
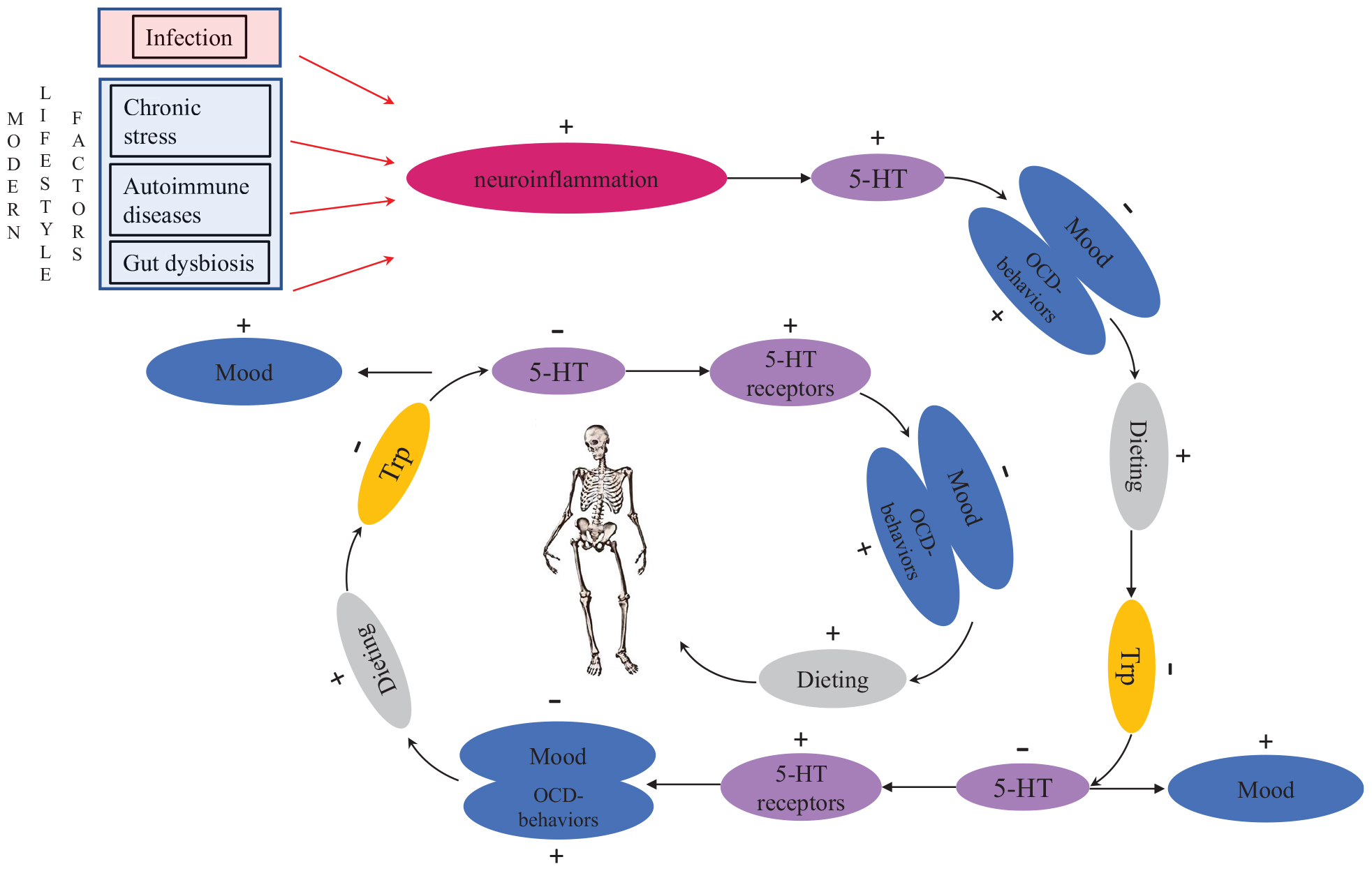 図1。 神経性食欲不振につながる、悪質で潜在的に致命的なセロトニン作動性恒常性ストレス/飢vサイクル。 TRP、トリプトファン。 5-HT、セロトニン。
図1。 神経性食欲不振につながる、悪質で潜在的に致命的なセロトニン作動性恒常性ストレス/飢vサイクル。 TRP、トリプトファン。 5-HT、セロトニン。
ケイ等。 (2009) ANの個人はセロトニン作動性システムに内因性の欠陥があり、思春期の個人化に関する初潮またはストレス関連の問題の間に性腺ステロイドが変化すると、セロトニン作動性システムの活性がさらに変化すると仮定した。 しかし、この説明は、現代社会におけるAN有病率の増加や、男性の摂食障害の発生を説明することはできません。 近接メカニズムを最終的な原因とリンクするには、生殖発達のこの中心期間の周りのthinせに対する性内競争の激化によって潜在的に引き起こされる初経および成人初期のANの発症のタイミングを考慮することが重要です。 性内競争は、競争の激しい個人に慢性的なストレスを引き起こす可能性があります(cf. Vaillancourt、2013)。 重要なことに、慢性ストレスは、人間と人間の両方の脳のセロトニンレベルを増加させることが知られています。 Hale et al。、2012)および他の動物(例、 アデル他、1988; キーニーら、2006; Vindas et al。、2016)。 ラットでの実験的研究は、絶食が脳のセロトニンレベルを低下させることを示しました(Haleem and Haider、1996)。 同じメカニズムは、セロトニン作動性システムがアップレギュレートされているストレスのある人々が、不快な状態を緩和するためにダイエットを開始したときにも発生します。 全体として、これらの調査結果は、慢性的なストレスと摂食障害の間の機械的リンクの一部を説明しています(図1).
神経性過食症のセロトニン作動性システムの機能不全に関する証拠もあります( シェーグレン、2017)が、ANとは異なる方法で。 BN患者のセロトニンレベルは、短時間の空腹時(例、睡眠中)でも健康なコントロールよりも低下し、気分過敏症と過食症エピソードにつながります(シュタイガー他、2001)。 セロトニン系の機能のこれらの異常は回復後も持続し、BNの発症前にすでに存在していた可能性があることを示唆しています(Kayeら、2001)。 AN患者とは対照的に、BN患者のトリプトファン枯渇は気分を低下させ、過食を促す衝動を引き起こす( シェーグレン、2017)。 神経画像研究では、健康なコントロールと比較して、BN患者の5-HT1A結合の増加が発見されました(Galusca et al。、2014)。 セロトニン受容体5-HTの活性化2CRは、ラットモデルで味の良い食物の過食を減らしました(Martin他、1998; フレッチャー他、2010; ヒギンズら、2013; プライス他、2018)。 同様に、SSRIの薬物療法は、過食症の衝動を軽減しました( Tortorella et al。、2014)。 これらの発見は、BN患者が中枢神経系でのセロトニン産生を減少させたという仮説を支持しています。
性内競争への反応における個人差の原因
女性と女性の性内競争に対する反応には大きな個人差があります:一部の個人は暴食をし、他の人はstar死します(図2)。 ほとんどの若い女性は、薄さのために性内競争にさらされていますが、摂食障害を発症するのはごく一部です。 この変動の理由は、ストレス応答性およびセロトニン作動系の機能の個人差に関連しているようです。 摂食障害の病因を理解するためのXNUMXつの中心的な質問は、摂食障害を持つ人々のセロトニン系のストレス応答性と機能障害の主な原因は何ですか?
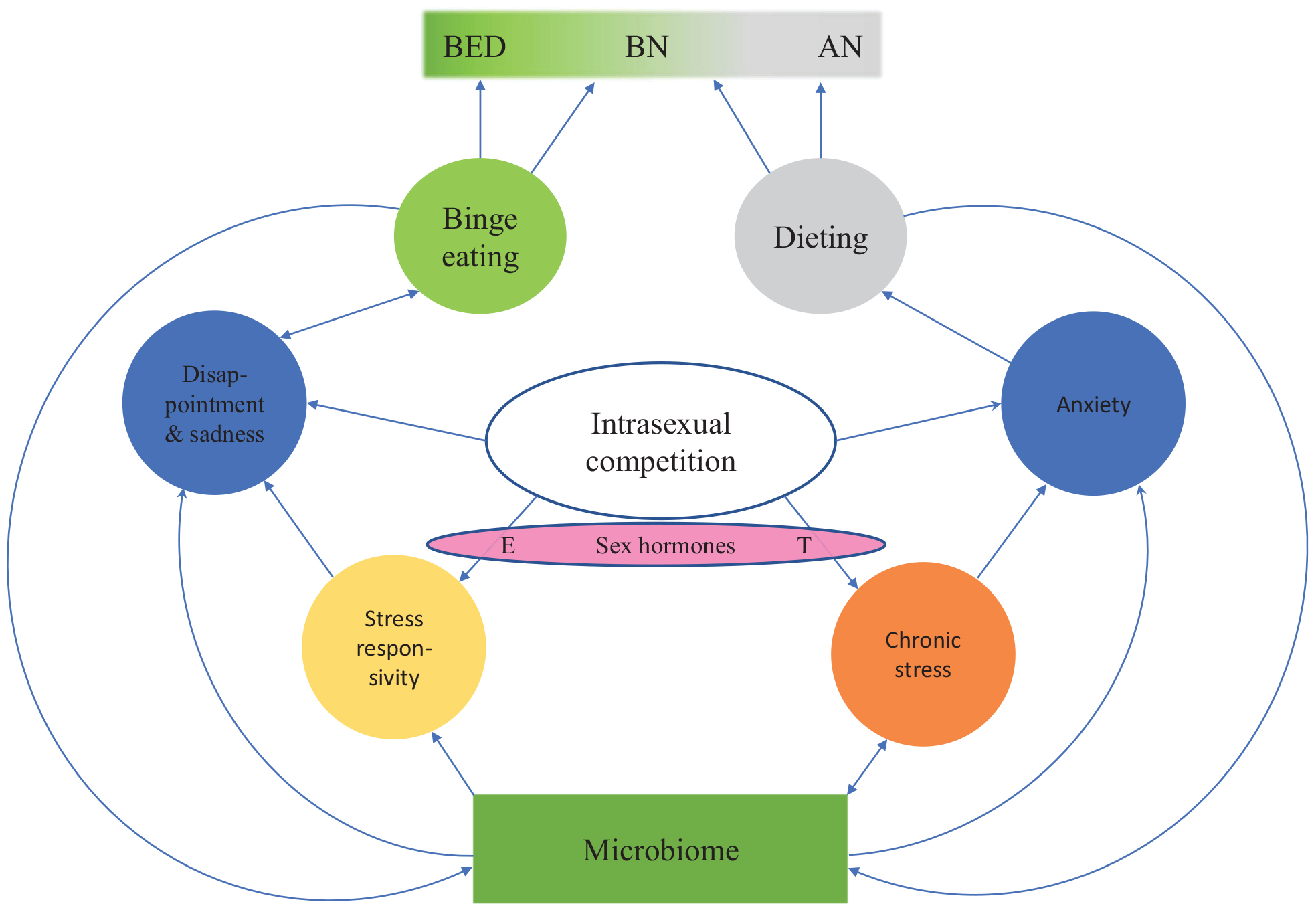 図2。 摂食障害の進化的精神神経免疫学的連続体モデル。 モデルは、薄さのための性内競争がどのようにさまざまな感情状態につながるかを示しています(青い丸)。 これらの感情状態の個人差および結果として生じる摂食行動(緑色と灰色の円)は、出生前の性ホルモン暴露の個人差と、病前および現在の微生物叢構成によって部分的に駆動されます。 したがって、摂食障害の個人間および個人内の変動は、炎症、ストレス、セロトニンレベル、トリプトファン濃度、および微生物叢の動的な変化から生じる可能性があります。 BED、過食症; BN、神経性過食症; AN、神経性食欲不振; E、エストロゲン; T、テストステロン。
図2。 摂食障害の進化的精神神経免疫学的連続体モデル。 モデルは、薄さのための性内競争がどのようにさまざまな感情状態につながるかを示しています(青い丸)。 これらの感情状態の個人差および結果として生じる摂食行動(緑色と灰色の円)は、出生前の性ホルモン暴露の個人差と、病前および現在の微生物叢構成によって部分的に駆動されます。 したがって、摂食障害の個人間および個人内の変動は、炎症、ストレス、セロトニンレベル、トリプトファン濃度、および微生物叢の動的な変化から生じる可能性があります。 BED、過食症; BN、神経性過食症; AN、神経性食欲不振; E、エストロゲン; T、テストステロン。
これらの個人差を引き起こす可能性のある要因は、腸内共生である可能性があります(cf. Temko et al。、2017)。 他の動物におけるかなりの量の経験的証拠は、腸内微生物叢がストレス応答性、不安様行動、および神経内分泌視床下部-下垂体-副腎(HPA)ストレス軸の活性化の設定点に影響を与えることを示唆しています Fosterら、2017も参照してください Molina-Torres et al。、2019)。 腸内共生によって調節されるストレス関連の生理学および行動の変化は、抗生物質への曝露、貧しい食生活、母乳育児の不足、帝王切開による出産、感染、ストレス曝露、および他の多くの環境要因による微生物組成の変化に起因する可能性があります Fosterら、2017).
摂食障害と微生物叢の間の仮説のメカニズムの重要な証拠は、摂食障害のある人の64%が過敏性腸症候群と診断されているという発見によって提供されます(Perkins他、2005)。 最近の研究では、AN患者は糞便微生物叢の豊富さ、多様性、および微生物組成のコントロールから逸脱していることがわかっています( Schwensen et al。、2018)、再給餌後も健康なコントロールとは著しく異なるままです(Kleiman et al。、2015; Mack et al。、2016)。 微生物叢の逸脱の一部はダイエットによって引き起こされる可能性がありますが、微生物の多様性を制限する食事制限があります(昆虫だけでなく人間に関する研究でも見られます: Krams et al。、2017; Stevensら、2019)–これらの個人が病的前に逸脱した微生物叢を有していた可能性もあります。 最近の研究では、ストレスが腸内細菌叢を乱すことが示されています(Gao他、2018; Partrick et al。、2018; Molina-Torres et al。、2019)。 したがって、摂食障害を引き起こした慢性的なストレスは、摂食障害の患者の微生物叢を変化させた可能性があります(cf. Seitz et al。、2019)。 残念ながら、BEDおよびBN患者の微生物組成に関する研究は現在不足しています。 摂食障害患者の病前の微生物叢を試験する研究は特に価値があるだろう。 興味深いことに、大規模なスウェーデンの研究(ヘドマンら、2019)AN診断後217年以内にセリアック病のリスクが2.3%増加することがわかりました。 同様に、クローン病はANでXNUMX倍多く、潰瘍性大腸炎はANでコントロールよりXNUMX倍多かった(Wotton et al。、2016)。 これらの疾患は、共生によって引き起こされるため(例、 Ni et al。、2017)、AN患者における有病率の増加は、共生とANの間のリンクに追加のサポートを提供します。
無菌マウスの研究により、幼若期の微生物叢の欠如は、血漿中のトリプトファン濃度の増加と脳のセロトニン作動性活性の増加をもたらすことが示されています(クラーク他、2013)。 これらの変化は、トリプトファン代謝に影響を与えることが知られているプロバイオティクス細菌をマウスに提供することにより正規化できます(クラーク他、2013)。 総括すると、上記で検討した研究は、摂食障害におけるセロトニン作動性の異常は、腸の微生物叢の逸脱によって少なくとも部分的に引き起こされる可能性があることを示しています。 これは将来の研究にとって重要な手段であると信じています(cf. Seitz et al。、2019)、摂食障害に対するマイクロバイオームベースの治療法を開発する立場になる前に、かなりの量の作業が必要です。
連続体としての摂食障害
これは重要ですが、ANと診断された女性の50%以上がBNを発症するという理論的および臨床的に過小評価されていますBulikら、1997)。 ANがBNに変わる理由のXNUMXつは、個人の栄養状態が改善されると、腸内微生物叢が変化し、それがストレス応答性とセロトニン系の機能に影響を及ぼす可能性があることです。 この仮説は、過食性ANサブタイプ(AN-B)が微生物群集構成(AN-R)制限型と異なることを示す知見によって裏付けられています(Mack et al。、2016)、再給餌により腸内細菌叢が変化する(Kleiman et al。、2015)。 さらに、再給餌によりトリプトファン濃度が増加した後、セロトニン系の恒常性応答は、時間とともにセロトニンレベルが過度に低下する状態につながる可能性があり、その結果、過食症が引き起こされます(cf. シュタイガー他、2001).
興味深いことに、メタ分析により、AN患者の血漿中サイトカイン濃度(特にIL-6およびTNF-α)が増加していることがわかりましたが、BN患者のサイトカインレベルはコントロールと異なりませんでした(Daltonら、2018)。 非ヒト動物での実験的研究では、IL-6サイトカインの投与により、側坐核のセロトニンレベルが上昇し、ドーパミンレベルが低下することがわかりました。Song他、1999)。 これらの調査結果に基づいて、AN-R患者のサイトカインレベルが低下すると、過食の衝動が現れる程度までセロトニンレベルが低下し、その結果、患者の摂食障害の診断がAN-Bに変わると仮定します。 。 この仮説は、AN-B患者はAN-R患者よりも炎症レベルが低いという発見によって裏付けられています(Solmi et al。、2015).
全体として、摂食障害の根底にあるこれらの仮定された共有メカニズムは、摂食障害が別個の障害ではないことを示しています。 代わりに、BEDがスペクトルの一方の端にあり、AN-Rがもう一方の端にある連続体を形成しているようです。 BNおよび過食症型AN(AN-B)は、極値(図2)。 以前は、摂食障害の診断を横断する見方が疑問視されていました(バーミンガムら、2009)、部分的には、摂食障害の病因におけるストレス、神経炎症、腸内共生の役割が理解されていなかったため。 この記事で紹介したモデル(図1, 2)は、炎症、ストレス、セロトニンレベル、トリプトファン濃度、および微生物叢の動的な変化が、摂食障害の変化を引き起こすことを示唆しています。 上記の近接メカニズムを摂食障害の連続体モデルに統合すると、摂食障害の過程で人の症状が頻繁に変化する理由と、その後患者が別の摂食障害と診断される理由を説明できるようになります。
自己免疫および摂食障害
食事の注文のすべてのケースは、thinせを求める性内競争によって引き起こされるとは限りません。 たとえば、体重が増えたり太ったりすることを強く恐れないAN患者がいます。 DSM-V(アメリカ精神医学会、2013)ボディイメージ障害(すなわち、非脂肪恐怖AN)を経験していない低体重患者を説明するために、診断ARFID(回避/制限的食物摂取障害)を導入しました。 ARFIDは、先進国で非常に一般的である疎油性ANよりも発展途上国でより一般的であるようです( ベッカー他、2006)。 同様に、ANおよびその他の摂食障害は、異性愛者より同性愛者の間ではるかに一般的ですが(Liら、2010; カルゾら、2018)、摂食障害は、一部の異性愛者の男性で発生します。 薄さは男性よりも女性の配偶者の好みほど重要ではないため、これは薄さのための性内競争を通じてのみ説明するのは難しいかもしれません(Liら、2010).
神経炎症がANで重要な役割を果たしている場合、重要な問題は、これらの場合に神経炎症を引き起こすものは何ですか? 当然、神経炎症を引き起こす慢性ストレスは、性内競争以外の要因によって引き起こされる可能性があります。 これは、特に非脂肪恐怖症のAN患者に当てはまります。 慢性ストレスに加えて、神経炎症は自己免疫疾患および自己炎症性疾患によって引き起こされる場合もあります(Najjar et al。、2013)。 それに対応して、ANが感染によって引き起こされた多くの症例報告があります:これらの症例は「自己免疫性食欲不振症」(ソコルとグレー、1997; ソコル、2000)。 場合によっては、OCDが感染に続いて観察されることがあります(アメリカ精神医学会、2013)、免疫系の活性化とOCDおよびANの発症との関連性をさらにサポートします(図1)。 脂肪恐怖症のAN患者と同じように、非脂肪恐怖症のAN患者も、ダイエットによって不安を和らげることを学ぶかもしれません。 これは、疎油性ANと同じ悪ストレス/飢starサイクルにつながる可能性があります(図1)。 さらに、我々の精神神経免疫学的モデルは、薄い理想がなかった歴史社会に存在していた非脂肪恐怖神経性食欲不振の歴史的事例についても説明を提供します(cf. アーノルド、2013).
デンマークで実施された大規模な全国規模の人口調査では、 Zerwas et al。 (2017) 自己炎症性疾患または自己免疫疾患により、ANのリスクが36%、BNリスクが73%、摂食障害(EDNOS)リスクが72%増加することがわかりました。 この効果は、女の子よりも男の子のほうが強かった(Zerwas et al。、2017)。 男の子の場合、自己炎症があるとEDNOSのリスクが740%増加しました。 スウェーデンの大規模な研究では、先行する自己免疫疾患によりANリスクが59%増加したことが報告されました(ヘドマンら、2019)。 摂食障害と自己免疫疾患の遺伝的関連を分析する別の大規模な研究では、神経性食欲不振症と自己免疫疾患の遺伝的重複は特定されませんでした(Tylee et al。、2018)。 これは、遺伝的要因ではなく環境的要因がANと自己免疫疾患との関連を引き起こすことを示唆しています。
これらの発見は、摂食障害の精神神経免疫モデルに幅広いサポートを提供するものと解釈します。これには、次の1つの理由があります。(XNUMX)慢性ストレスは自己免疫疾患の原因となるSong他、2018)、(2)多くの自己炎症性疾患および自己免疫疾患は、神経炎症を増加させることが知られています(Najjar et al。、2013)、(3)免疫系の活性化はストレス応答性を高めることが知られています(ヤウとポテンザ、2013、および(4)腸内細菌叢の共生は、自己炎症性疾患の発症につながる可能性があります(Lukens et al。、2014)。 「神経性食欲不振症および神経性過食症の神経化学」のセクションでレビューしたように、摂食障害においても共生は一般的です。
したがって、ARFID(非脂肪恐怖AN)の場合、神経炎症のある人は、食事と空腹が自己免疫反応を低下させるため、食事と空腹が不安や不快感を軽減できることを知るかもしれません(cf. ハフストロムら、1988)およびセロトニン作動性システムのダウンレギュレーション(cf. Kayeら、2009)、悪質なダイエットサイクルにつながり、最終的にはANになります。 この精神神経免疫学的メカニズムにより、異性愛者および無性者の男性がANに苦しむことがある理由を説明できます(cf. Carlat et al。、1997)薄さを求める強い性内競争が異性愛者の女性と同程度に存在しない場合でも(cf. Abed et al。、2012).
摂食障害の併存疾患
摂食障害は通常、他の精神障害との併存疾患率が高い(Keski-RahkonenおよびMustelin、2016)。 たとえば、成人のAN患者の93〜95%に共存気分障害、55〜59%に不安障害、5〜20%に物質関連障害(Blinder et al。、2006)。 BNの精神障害の共存症に関する研究では、成人BN患者の94%が気分障害、55%が不安障害、34%が物質使用障害(Swansonら、2011)。 BNのティーンエイジャーでは併存疾患の頻度は低かった:49.9%が気分障害、66.2%が不安障害、20.1%の薬物乱用がBNティーンエイジャーであり、57.8%が行動障害(Swansonら、2011).
BEDは最近、別の障害として分類されているため、共存する精神障害に関する研究はほとんどありません(cf. Olguin et al。、2017)。 米国のティーンエイジャーに関する大規模な疫学研究では、BEDの個人の45.3%が共存気分障害、65.2%が不安障害、物質乱用がティーンエイジャーの26.8%で発生し、42.6%が行動障害であることがわかりました(Swansonら、2011).
摂食障害と多くの類似点を共有するOCDは、腸内微生物叢の調節不全に関連しています(ターナ他、2017)および脳内のセロトニン活性の変化(Lissemore et al。、2018)。 摂食障害と同様に、OCDは他の精神障害との高い併存疾患に関連しています(Hofmeijer-Sevink et al。、2013)。 腸内微生物叢の調節不全と慢性ストレスは両方とも気分障害と不安障害に関連しているため( Bekhbat and Neigh、2018; Liangら、2018)、これらの併存疾患の発生に関するもっともらしい説明は、OCDおよび摂食障害では、これらの併存疾患は、共生とストレスに対する感受性の高まりによって引き起こされるというものです。
重要なのは、多くのAN症状が次の症状のようです 飢餓、精神障害ではありません。 たとえば、有名なミネソタ州の飢study研究では、36人の健康な男性が6か月間半飢starにさらされました(キー、1950)。 男性はその後、儀式的な摂食、食物へのこだわり、摂食などの摂食障害に似た症状を示し始めました。 飢えた男性はまた、買いだめおよび強迫的な収集行動を発達させ、飢vが強迫性障害(OCD)に似た症状を引き起こしたり強化したりする可能性があることを示唆しています。 飢えた男性の中には、料理の本を読んで、食べ物について夢を見て、常にそれについて話す傾向がありました(キー、1950)。 食物に対する同様の強迫観念がAN患者で一般的に観察されている(クリスプ、1983).
飢star状態にある男性も、いらいらし、不安を感じ、落ち込んでいます。これは、飢vが飢v誘発性鬱病につながったことを示唆しています。 ランタラら、2018)。 多くの被験者では、再給餌後も症状が持続しました。 長期にわたる飢episodeのエピソードは、無関心と社会的引きこもりにつながります(cf. キー、1950)、これも一般的なANの症状です。 ミネソタ州の飢study研究では、飢experiment実験が終了するとすぐに、多くの男性が体重が増えすぎて「だらしない」という懸念を表明したことが示されました。キー、1950)。 同様の種類の例は、飢reports時の症例報告と日記に記載されています(キー、1950)。 衰弱しているにもかかわらず、ほとんどの男性は自分自身を低体重とは見なしませんでした(キー、1950)。 したがって、飢starはANの特徴である歪んだ自己イメージを刺激し、飢itself自体がAN患者に見られる一種の精神病理を引き起こす可能性があると考えられます。
摂食障害の危険因子
遺伝因子と神経炎症
家族、双子および養子縁組の研究は、遺伝的要因が摂食障害に対する感受性の変動に寄与することを一貫して実証しています。 BEDの遺伝率の推定値は41〜57%です。 BNの遺伝率の推定範囲は30〜83%ですが、ANの遺伝率は28〜78%(Thornton et al。、2011)。 摂食障害は家族性です。ANのある人の女性の親ANは、ANのない人の親thanよりも11.3倍発症する可能性が高いです。 BNを持つ個人の女性の親族は、BNを持たない個人の親族より12.3Strober et al。、2000)。 摂食障害について特定の遺伝的危険因子はほとんど特定されていません( Mayhew et al。、2018)、最近の研究では、AN病因の基礎となるXNUMXつの遺伝子座が示されましたが、障害の代謝精神医学的起源が示唆されました(Watsonら、2019)。 双子の研究は、摂食障害と不安障害の間で共通の伝達があることを明らかにしました(キール他、2005)、ANとOCDの間(アルトマンとシャンクマン、2009)およびBNとパニック障害(キール他、2005)。 これらの調査結果の最も可能性の高い説明は、上記のエビデンスで示唆されているように、共有された伝達は神経炎症とストレス応答性に対する脆弱性によって引き起こされるというものです。 神経炎症とストレスに対するこの脆弱性は、GWASの研究がANと統合失調症、大鬱病性障害、双極性障害、自閉症のような他の多くの精神障害との間に遺伝的相関を発見した理由を説明するかもしれませんAnttila et al。、2018; Sullivanら、2018)、それらのすべてで神経炎症が役割を果たすため(Najjar et al。、2013)。 微生物叢の組成の違いとともに、これらの神経炎症に対する遺伝的感受性は、性内競争がBED、BNまたはANにつながるかどうかを部分的に説明するかもしれません(cf. 図2).
小児期の虐待、ストレス、エピジェネティクス、微生物叢
性的、感情的または身体的虐待の形での小児期の虐待は、情緒障害のリスクを高めます( ホッペンとチャーダー、2018)。 小児期の虐待は、摂食障害を発症するリスクをXNUMX倍以上増加させます(Caslini et al。、2016)。 小児期の虐待は、HPA軸遺伝子のDNAメチル化(DNAm)の変化などの後成的メカニズムによって部分的に駆動される結果である、成人期のストレス応答性を増加させます(ブスタマンテら、2016)。 小児期の慢性ストレスは、変化した、次善の微生物叢がストレスを増加させる素因となるような方法で、微生物叢に影響を与える可能性があります(O'Mahony et al。、2016)。 ストレスに対する感受性の増加に加えて、幼年期のストレスはミクログリアを刺激することがあり、これはその後のストレスに対する神経炎症反応の増強につながる可能性があります( Calcia et al。、2016)。 疫学的研究では、AN発症前のXNUMX年以内に慢性ストレスがしばしば報告されています(Rojo et al。、2006)。 AN患者は、健康なコントロールよりも生涯ストレスのレベルが高く、ストレスへの対処が困難であると報告しています(Soukup et al。、1990)。 レトロスペクティブ研究により、ANと対照サンプルでは重度の生活ストレスが異なり、症例の67%でANの発症が予測されることが判明した(シュミット他、2012).
性ホルモンの役割
出生前および現在の性ホルモンのレベルは、薄さをめぐる性内競争が摂食障害につながるかどうかに部分的に影響しているようです。 性ホルモンは、発症した摂食障害のタイプにも影響を与えます(図2)。 エストロゲンはHPA活性を刺激することが知られており、それによりストレス応答性が向上します(クディエルカとキルシュバウム、2005)。 対照的に、アンドロゲンはHPA活性を低下させる傾向があり、それによりストレス応答性を低下させます(クディエルカとキルシュバウム、2005)。 男性は通常、状態関連の状況でより高いHPA活性化を示しますが、女性は社会的拒絶を含む状況でより高いHPA活性化を示します( Del Giudice et al。、2011).
したがって、より男性的な異性愛の女性(Bártováet al。、2020)、すなわち、出生前のアンドロゲン曝露が高い女性または現在のテストステロン濃度が高い女性(ルオトら、2019a, b)–したがって、社会的地位の向上への意欲(cf. Nave et al。、2018)– ANを開発する可能性が高い。 対照的に、社会的拒絶に対してより敏感な女性が多いほど、BEDを発症するリスクが高くなると予想されます。 実際、桁比(2D:4D、すなわち出生前のアンドロゲン曝露のバイオマーカー: ルオトら、2019a)BN患者よりもAN患者の方が男性的であり、コントロールは中間桁比(Quinton et al。、2011)。 これは、AN患者がBN患者および対照よりも高い出生前アンドロゲン曝露を経験した可能性があることを示唆しています(非臨床集団におけるこれらの発達メカニズムの詳細については、 ルオトら、2019a, b)。 女性では、出生前のテストステロンレベルが低く、思春期の卵巣ホルモンレベルが高いと、BEDのリスクが高くなるようです。 男性では、出生前の高いテストステロンレベルがBEDから保護するようです(Klump et al。、2017)。 これらの調査結果は、表現型の変化における性ホルモンの役割を強調しています(図2)および摂食障害の性差。
現在の摂食障害の治療
摂食障害の治療は、他の精神障害の治療よりもはるかに効果が低くなります。 AN患者の46%のみが完全に回復し、20分のXNUMXが部分的に回復し、XNUMX%のAN患者は慢性状態のままです(Arcelus et al。、2011)。 病気の平均期間は6年(シュミット他、2016)。 現在、ANに対する有効な薬理学的治療法はありません。 選択的セロトニン再取り込み阻害薬(SSRI)は、AN(デイビスとアッティア、2017)。 米国またはEU(神経性食欲不振症の治療に承認された薬はありません(ボデルとキール、2010; Starr and Kreipe、2014)。 したがって、AN治療は、さまざまな種類の治療と体重を回復する努力に基づいています(例、 Brockmeyer et al。、2017; Harrisonら、2018 およびその中の参照)。 家族ベースの治療(FBT)の有効性は、精神療法の他の形態(Starr and Kreipe、2014).
BNの主な治療法は認知行動療法(CBT)です。これは、過食症の根底にあるネガティブな思考パターンを変えながら、摂食行動を正常化しようとするものです(フェアバーン、2008)。 SSRIは過食症の行動をわずかに抑制しますが、通常は排除しません(Mitchellら、2013)。 BN患者の抗うつ薬治療のドロップアウト率は約40%(Bacaltchuk and Hay、2003)。 現在のBN治療は特に効果的ではありません:追跡調査により、10年間で患者の50%のみが完全に回復することが示されています(Hay et al。、2009).
抗うつ薬は、短期的には過食症のエピソードに対して中程度の効果しかありませんが、それらの長期的な効果は不明です(McElroyら、2012)。 さらに、それらは体重を減らすのに役立ちませんし、CBTのアンチビンジ摂食効果を高めるようには見えません(McElroyら、2012)。 実際、AN治療で使用される第XNUMX世代の抗精神病薬は、BEDおよびBN患者の過食を誘発または悪化させます(McElroyら、2012; Cuesto et al。、2017)。 二重盲検およびプラセボ対照実験は、抗てんかん剤トピラマートが肥満を伴うBEDの過食症エピソードに対して有効であることを示しています。 トピラマートは高い抗炎症特性を持ち、ラットの神経炎症と酸化ストレスを軽減することが示されています(Pinheiro et al。、2015)。 また、マウスでのストレスによるアルコール消費の増加を減衰させることが示されています(Farook et al。、2009)、ストレス応答性を低下させる可能性があることを示唆しています。 興味深いことに、トピラマートはOCDの衝動を効果的に減らします(ルビオら、2006; Van Ameringen et al。、2006; Mowla et al。、2010; ベルリンら、2011)。 残念ながら、トピラマートは、ANの治療薬としてもANの既往歴のある患者にも適していません。トピラマートは副作用として食欲を減らし、体重減少を促進するためです。 ANの既知の危険因子を持つ人にANを誘発することさえあります(Lebow et al。、2015)。 米国およびカナダでは、中等度および重度のBEDに対して承認されている唯一の薬物はリスデキサフェタミンであり、注意欠陥多動性障害(ADHD)の治療にも使用されています(ホ・ダガン、2017)。 ランダム化二重盲検試験では、リスデキサフェタミンがプラセボよりも過食に対してより効果的であることが示されています。 しかし、リスデキサフェタミンには、口の渇き、頭痛、不眠症などの有害な副作用があり、多くの患者で薬物の中止につながります(ホ・ダガン、2017)。 さらに、摂食障害の原因となる根本的な問題を取り除くのではなく、単に症状を緩和するだけで、最適ではない長期的な解決策を提供します(cf. ランタラら、2017).
精神神経免疫学に基づく摂食障害の治療
私たちが提示した精神神経免疫モデルは、摂食障害の治療の有効性を改善する可能性を秘めています。 このモデルでは、AN患者に家族ベースの治療と心理療法を提供するよりも、神経炎症と慢性ストレスを減らすことで、体重を減らすという強迫観念を減らすことの方が効果的であると仮定しています。 また、進化精神医学に基づいたCBTは、患者の自己イメージと薄い美しさの理想に対する態度を、より健康的な方向に向けることができることを示唆しています。 批判的に、我々の精神神経免疫学的モデルは、神経炎症とストレスを軽減するライフスタイルの変化がAN症状を軽減すると予想されることを示唆していますが、これを経験的に検証するにはさらなる臨床研究が必要です。
神経炎症を標的とする可能性のあるANの薬物療法
最近の研究では、オランザピン(非定型抗精神病薬)治療は、AN(Dold et al。、2015; Himmerich et al。、2017)。 マウスの研究により、オランザピンは神経炎症を軽減することが示されています(Sharon-Granit et al。、2016)。 また、TNF-αおよびIL-6を抑制し、抗炎症性サイトカインであるIL-10レベルを増加させます(杉野ほか、2009)。 したがって、オランザピンがAN患者をどのように支援するかの考えられるメカニズムは、神経炎症を軽減する可能性がありますが、これは経験的に検証されたままです。
ケーススタディは、ANに対する抗TNF-α治療の肯定的な効果を示しています(Solmi et al。、2013). エサラトマネシュ他 (2016) その抗炎症特性で知られているミノサイクリン抗生物質は、有害な副作用を引き起こすことなく、OCD患者のOCD症状を有意に減少させることを発見しました。
亜鉛、拒食症および神経炎症
臨床研究からの証拠は、AN患者の血清亜鉛レベルが低く、尿中亜鉛排泄率が低いことを示しています(Katzら、1987)。 亜鉛欠乏の重症度は、ANの重症度、ならびにより高いレベルのうつ病および不安症と関連しています(Katzら、1987)。 亜鉛補給のいくつかのランダム化比較試験では、被験者の体重の有意な増加が報告されています(サファイ・クッティとクッティ、1986; サファイ・クッティ、1990; バーミンガムら、1994; バーミンガムとグリッツナー、2006)。 亜鉛欠乏は、炎症誘発性サイトカイン、特にTNF-αおよびIL-6の産生の増加と関連していることが知られています(ガンモとリンク、2017)。 したがって、亜鉛の補給とAN症状および体重増加の減少の間の可能なメカニズムは、神経炎症の減少を介して作用する可能性があることを示唆します。 神経炎症の減少は、強迫観念を減少させます。 この解釈は、特に亜鉛錠剤を摂取したAN患者で(プラセボ錠剤を摂取した患者とは対照的に)食事および食物に対する態度がよりポジティブになったと報告したプラセボ対照試験の結果によって裏付けられています(Khademian et al。、2014)。 したがって、亜鉛はAN患者のOCD症状を軽減すると思われる。 興味深いことに、亜鉛サプリメントは、摂食障害のないOCD患者でも症状を軽減します(Sayyah et al。、2012)。 全体として、亜鉛がAN症状に影響を及ぼすメカニズムを理解するには、より多くの研究が必要です。
亜鉛に加えて、他の微量栄養素も神経炎症を減らすために使用できます。 たとえば、AN患者にはビタミンDの欠乏があります(Veronese et al。、2015; Tasegian et al。、2016)ビタミンDの補給は炎症を軽減することが知られています(グロスマンら、2012; Zhangら、2012; Berk et al。、2013)および神経炎症(Koduah et al。、2017).
糞便微生物叢の移植とプロバイオティクス
科学的進歩が、摂食障害の病因における微生物叢の役割に対する経験的サポートを提供し続けている場合、健康な個人からの糞便微生物叢移植が摂食障害の将来の治療的処置の一部になる可能性があります。 最初に公開されたケーススタディ(de Clercq et al。、2019)AN患者に対する糞便微生物叢移植後の有意な体重増加を報告した。 ただし、この分野の研究はまだ始まったばかりです。 糞便微生物叢の移植に加えて、腸内細菌叢はプロバイオティクスまたは他のサプリメントで治療的に操作される可能性があります(Stevensら、2019)。 残念ながら、蓄積された証拠は、プロバイオティクスが大うつ病性障害に見られる炎症性活性化を減らす有望なアジュバント治療であることを示唆していますが Park et al。、2018)、摂食障害の治療としてのプロバイオティクスの有効性に関する研究は現在不足しています。
進化精神医学に基づいた過食症(BED)治療
進化的精神医学に基づいたBED治療は、私たちの見解では、過食症のエピソードを促進するダイエット努力の悪循環を止めることに焦点を当てるべきです。 患者は、健康的な食事と運動を通して体重を減らすよう努めるべきです(cf. リンデバーグ、2010; Temko et al。、2017; Leoneら、2018)断食によってカロリー摂取量を減らそうとするよりも。 これは、現在のBEDのCBT治療と非常によく似たアプローチです(cf. ヒルベルト、2013)。 健康的な食事は、BED患者のストレス応答性の低下にも役立つ可能性があります:動物での実験的研究は、腸内微生物叢がストレス応答性に影響することを示しています(Bravo et al。、2011)。 さらに、健康的な食事、特にオメガ3脂肪酸とポリフェノールは炎症を軽減します(リコルディら、2015)、HPA活性を刺激することが知られています(ヤウとポテンザ、2013; ローレダー、2019)自主規制能力の低下(Shields et al。、2017)。 また、運動はストレス応答性を低下させるのに役立ちます(cf. Zschucke et al。、2015)。 これらのBED治療オプションの有効性は、将来の臨床研究で決定されるべきです。
摂食障害の究極レベルの予防
上記の治療は、摂食障害の根底にある近接メカニズムに焦点を当てています。 しかし、どの治療法も実質的な長期効果を発揮するには、摂食障害の根本的な原因にも対処する必要があります。 メディアリテラシープログラムは、摂食障害の効果的な予防手段となる可能性があります(Liら、2014)メディアにおけるスリムな女性と筋肉質の男性の理想化された画像が、性内競争の動機を過剰に活性化する限り(ブースロイド他、2016; Borau and Bonnefon、2017; サンダースとイートン、2018)、そして若者がメディアへの全体的な露出を減らすことが難しい限り。 メディアリテラシープログラムは、参加者にメディアの消費者に情報を提供することを目指しています。 心理教育的コンポーネントが含まれ、写真編集ソフトウェアを使用して画像をより「完璧」に見せるための方法を示しています(Liら、2014)。 メディアリテラシープログラムは、若者に広告やメディア画像に関する事実を提供することで、細い身体の理想を内在化する可能性が低く、体重にあまり関心がないという前提に基づいています:結果として、摂食障害に関連する行動が軽減されます(Liら、2014).
メディアリテラシープログラムは、摂食障害に関連する認知状態の軽減に効果的です。 学部生に関する研究では、4週間のメディアリテラシー介入により、身体の不満と薄さの社会文化的理想の内在化が減少した(ワトソンとヴォーグン、2006)。 別の研究では、30セッションのメディアリテラシープログラムに参加することで、思春期の少女の体型、体重、ダイエットの懸念や身体の不満が軽減されることがわかりました。効果はXNUMXか月のフォローアップでも持続し、長期的な改善を示唆しています(Wilksch and Wade、2009)。 メディアリテラシーに加えて、参加者が薄い美しさの理想に対して反姿勢的な姿勢を取るように訓練される不協和に基づくアプローチは、摂食障害に関連する行動や思考を減らすのに効果的です(Sticeら、2001, 2006; ベッカー他、2006; イェーガーとオデア、2008). リーら (2014) メディアリテラシープログラムと不協和音アプローチの組み合わせが、若者の摂食障害を防ぐ効果的な方法である可能性があることを示唆しました。
結論
収evidenceする証拠は、摂食障害はしばしば、薄さを求める性内競争に対する不適応な反応であることを示しています。 「摂食障害の不一致仮説」で概説した不一致仮説は、食物の豊富さと座りがちな生活様式の進化的に新しい条件が、 適応メタ問題 食物摂取の心理的メカニズムが交配関連の心理的メカニズムと衝突する。 したがって、現代の人間における摂食障害の大規模な流行は進化の目新しさです:他の種のメンバーは知られていません 飢え死に 食物が豊富だからです これは、進化的情報に基づいた精神医学の驚くべき実現です。
この記事でレビューした証拠は、摂食障害を個別の状態として概念化するのではなく、連続的に見るべきだと示唆するようになりました。 この記事で紹介した精神神経免疫モデルによると、摂食障害の変動は、腸内細菌叢とストレス応答性の個人差から生じる可能性があります(図2)、神経炎症とセロトニン系に影響を与えます(図1)。 私たちの合成モデルは、(1)診断症状と関連する行動が摂食障害の範囲全体で実質的に重複する理由、(2)摂食障害の診断が難しい理由、(3)患者の診断が摂食障害間で移行する理由、4つの永続的な質問に対する答えを提供します(XNUMX)ANがXNUMXつの形式で存在する理由:脂肪恐怖ANと非脂肪恐怖AN。 このモデルによって導かれる将来の実証研究は、摂食障害の一般的な生物心理社会的理解をさらに発展させることが期待されています。
このレビュー記事は、摂食障害の違いが、ストレス応答性と慢性ストレスによって引き起こされる神経炎症の変動と共変動によって媒介される可能性があることを示唆しています。 ストレスの程度と(その後の)ストレス応答性と神経炎症が変化すると、モデルは患者の症状と摂食障害の診断がそれに応じて変化すると予測します(図2)。 我々がレビューした証拠は、BN表現型とAN表現型の違いは慢性ストレスによって引き起こされる神経炎症の程度から生じ、AN患者はBN患者より強い神経炎症を持っていることを示唆しています。 したがって、摂食障害の連続体における患者の位置(図2)は、ストレス応答性と神経炎症によって決定されますが、どちらもストレスの慢性化の影響を受けます。
この記事でレビューした証拠に照らして、神経炎症は摂食障害患者の体重を減らすという強迫観念を維持し、ANで最高でBED患者で最低であると考えられます。 BED患者はpatients清せず、BN患者よりも体重を減らすという執着が弱いことを示唆しています。 この強迫観念は、これらの問題に対処することを目的としたOCDのような行動だけでなく、食物、体重、食事、運動および外見に関する広範な精神的侵入によって媒介されます。 AN患者の神経炎症が強いほど、体重を減らすことへの強迫観念と体重増加の恐怖が強くなり、OCDのような行動がより持続的かつ極端になると仮定します(図1)。 このモデルのさらなる証拠は、オランザピンの有効性に関する調査結果によって示されています(Dold et al。、2015; Himmerich et al。、2017)およびANの治療薬としての亜鉛(サファイ・クッティとクッティ、1986; サファイ・クッティ、1990; バーミンガムら、1994; バーミンガムとグリッツナー、2006)。 結局のところ、両方とも抗炎症特性を持っていることが知られています。 ANおよびBNの治療オプションとしての別の抗炎症剤であるミノサイクリンの有効性について、将来の研究を行うことができます(cf. Esalatmanesh et al。、2016).
神経性食欲不振症の患者は神経炎症を持っていることを収束する(間接的ではあるが)証拠が示しているため、神経炎症が摂食障害のスペクトルの根底にある生物学的メカニズムであるという仮説をさらに裏付けるには、陽電子放射断層撮影スキャン(PET)研究が必要です。 さらに、ストレスホルモンレベル、ストレス応答性、セロトニンレベル、神経炎症および腸内細菌叢の組成が患者から測定される追跡調査 コースで 摂食障害のモデルは、モデルから生じる予測に従って症状が変化するかどうかを明らかにします。 最終的に、ここで紹介する進化的精神神経免疫モデルがさらなる経験的作業を促進し、摂食障害の治療と薬物の大幅な改善を提供し、最終的に摂食障害によってひどく衰弱した生活を送る何百万人もの人々の実用性を証明することを願っています。
著者寄稿
MRは原稿を起草しました。 MRは、精神神経免疫モデルを概念化しました。 SLはミスマッチ仮説を概念化しました。 SL、TK、およびIKは、知的コンテンツの原稿を批判的にレビューしました。 MRとSLが図を作成しました。 すべての著者が原稿の最終版に貢献し、承認しました。
資金調達
SLはEmil Aaltonen Foundationからの助成金によってサポートされました(SLの博士研究用)。 IKは、エストニア研究評議会(PUT-1223)およびラトビア科学評議会(lzp-2018 / 1-0393)によってサポートされていました。 TKは、ラトビア科学評議会(lzp-2018 / 2-0057)によってサポートされていました。 資金源は原稿の執筆に影響を与えませんでした。
利害の衝突
著者らは、潜在的な利益相反として解釈される可能性がある商業的または金銭的関係がない状態で研究が行われたと宣言しています。
参考情報
Abed、R.、Mehta、S.、Figueredo、AJ、Aldridge、S.、Balson、H.、Meyer、C.、et al。 (2012)。 摂食障害と性内競争:若い女性の進化仮説を検証します。 科学 ワールドJ 2012:290813. doi: 10.1100/2012/290813
Adams、TG、Kelmendi、B.、Brake、C.、Gruner、P。、およびBadour、C。(2018)。 強迫性障害の病因と維持におけるストレスの役割。 慢性ストレス 2:2470547018758043。 土井:10.1177 / 2470547018758043
アデル、A。、ガルシアマルケス、C。、アルマリオ、A。、およびゲルピ、E。(1988)。 慢性ストレスは、ラットの脳のセロトニンとノルアドレナリンを増加させ、さらなる急性ストレスに対する反応を敏感にします。 J.Neurochem。 50, 1678–1681. doi: 10.1111/j.1471-4159.1988.tb02462.x
Alboni、S.、Di Bonaventura、MVM、Benatti、C.、Giusepponi、ME、Brunello、N。、およびCifani、C.(2017)。 過食の動物モデルにおける炎症性メディエーターの視床下部発現。 Behav。 Brain Res。 320、420〜430。 土井:10.1016 / j.bbr.2016.10.044
アロンソペドロ、L.、Bes-Rastrollo、M.、Marti、A。(2019)。 抗うつ薬および抗精神病薬の使用が体重増加に及ぼす影響:系統的レビュー。 オベス 牧師 doi:10.1111 / obr.12934 [印刷前にEpub]。
アルトマン、SE、シャンクマン、SA(2009)。 強迫性障害と摂食障害の関係は何ですか? クリン サイコロル。 牧師 29、638〜646。 土井:10.1016 / j.cpr.2009.08.001
Anderluh、MB、Tchanturia、K.、Rabe-Hesketh、S。、およびTreasure、J.(2003)。 摂食障害のある成人女性における小児期の強迫性人格特性:より広範な摂食障害の表現型の定義。 あ。 精神医学 160、242〜247。 土井:10.1176 / appi.ajp.160.2.242
Andrews、TM、Lukaszewski、AW、Simmons、ZL、およびBleske-Rechek、A。(2017)。 生殖価値のキューベースの推定値は、女性の身体の魅力を説明します。 進化。 ハム。 行動。 38、461–467。 土井:10.1016 / j.evolhumbehav.2017.04.002
Anttila、V.、Bulik-Sullivan、B.、Finucane、HK、Walters、RK、Bras、J.、Duncan、L.、et al。 (2018)。 脳の一般的な障害における共通の遺伝率の分析。 科学 360、eaa8757。 土井:10.1126 / science.aap8757
Arcelus、J.、Mitchell、AJ、Wales、J。、およびNielsen、S.(2011)。 神経性食欲不振症およびその他の摂食障害患者の死亡率は、36件の研究のメタ分析です。 アーチ。 一般精神医学 68、724〜731。 土井:10.1001 / archgenpsychiatry.2011.74
Arthur-Cameselle、J.、Sossin、K。、およびQuatromoni、P.(2017)。 女性の大学のアスリートおよび非アスリートにおける摂食障害の発症に関連する要因の定性分析。 食べる。 無理。 25、199〜215。 土井:10.1080 / 10640266.2016.1258940
Attwells、S.、Setiawan、E.、Wilson、AA、Rusjan、PM、Mizrahi、R.、Miler、L.、et al。 (2017)。 強迫性障害の神経回路における炎症。 Biol。 精神医学 81、S97〜S97。 土井:10.1001 / jamapsychiatry.2017.1567
フロリダ州ベイラー、WH(2011年)。 「セロトニン:摂食障害の画像所見」 摂食障害の行動神経生物学、Vol。 6、eds RAH Adan、およびWH Kaye(ベルリン:Springer-Verlag Berlin)、59–79。 土井:10.1007 / 7854_2010_78
バルトヴァー、K。、シュトルボヴァー、Z。、ヴァレラ、MAC、およびバレントバ、JV(2020)。 男性の女性らしさと女性の男らしさは、社会的性欲と正に関連しています。 皆さん。 個別。 異なる。 152:109575。 土井:10.1016 / j.paid.2019.109575
バスティアニ、AM、アルテムス、M。、ピゴット、TA、ルーベンスタイン、C。、ウェルジン、TE、およびケイ、WH(1996)。 神経性食欲不振症および強迫性障害の患者における強迫観念と強迫の比較。 Biol。 精神医学 39, 966–969. doi: 10.1016/0006-3223(95)00306-1
ベイトソン、P。、およびラランド、KN(2013)。 ティンバーゲンのXNUMXつの質問:感謝と更新。 トレンドEcol。 進化。 28、712〜718。 土井:10.1016 / j.tree.2013.09.013
Baumeister、RF、Reynolds、T.、Winegard、B。、およびVohs、KD(2017)。 愛のために競う:性的経済学理論を交配コンテストに適用します。 J.イーコン。 サイコ。 63、230〜241。 土井:10.1016 / j.joep.2017.07.009
ベッカー、CB、スミス、LM、およびチャオ、AC(2006)。 ピアが促進する摂食障害の予防:認知的不協和とメディア擁護のランダム化された有効性試験。 J.Couns。 サイコロル。 53、550〜555。 土井:10.1037 / 0022-0167.53.4.550
Behl、A.、Swami、G.、Sircar、SS、Bhatia、MS、およびBanerjee、BD(2010年)。 強迫性障害におけるストレス関連生化学マーカーの可能性と酸化/抗酸化状態との関係。 神経心理生物学 61、210〜214。 土井:10.1159 / 000306591
Bekhbat、M。、およびNeigh、GN(2018)。 ストレスの神経免疫結果の性差:うつ病と不安に焦点を当てます。 脳の行動。 イミュン。 67、1–12。 土井:10.1016 / j.bbi.2017.02.006
Berge、JM、Loth、K.、Hanson、C.、Croll-Lampert、J。、およびNeumark-Sztainer、D.(2012)。 家族のライフサイクルの移行と摂食障害の発症:遡及的根拠のある理論的アプローチ。 J. Clin。 看護 21、1355〜1363。 土井:10.1111 / j.1365-2702.2011.03762.x
バーク、M。、ウィリアムズ、LJ、ジャッカ、FN、オニール、A。、パスコ、JA、モイラン、S。 (2013)。 うつ病は炎症性疾患ですが、炎症はどこから来ますか? BMC Med。 11:200. doi: 10.1186/1741-7015-11-200
ベルリン、HA、コーラン、LM、ジェニケ、MA、シャピラ、NA、チャップリン、W。、パランティ、S。 (2011)。 治療抵抗性強迫性障害におけるトピラメート増強の二重盲検プラセボ対照試験。 J.Clin。 精神科 72、716–721。 土井:10.4088 / JCP.09m05266gre
バーミンガム、C。、およびグリッツナー、S。(2006)。 亜鉛補給は、拒食症にどのような利点がありますか? 食べる。 体重の無駄。 11、e109〜e111。 土井:10.1007 / bf03327573
バーミンガム、CL、Touyz、S。、およびHarbottle、J.(2009)。 神経性食欲不振症と神経性過食症は別々の障害ですか? 「診断横断的」に挑戦する。 理論。 食べる。 不協和音。 ユーロ。 食べる。 不調和。 牧師 17、2–13。 土井:10.1002 / erv.896
Blinder、BJ、Cumella、EJ、およびSanathara、VA(2006)。 摂食障害のある女性入院患者の精神医学的併存疾患。 サイコソム メド。 68, 454–462. doi: 10.1097/01.psy.0000221524.77675.f5
ボデル、LP、キール、PK(2010)。 神経性食欲不振の現在の治療:有効性、安全性、および遵守。 サイコロル。 結果 ふるまい。 マナグ。 3、91〜108。 土井:10.2147 / PRBM.S13814
Boggiano、MA、Chandler、PC、Viana、JB、Oswald、KD、Maldonado、CR、Wauford、PK(2005)。 ダイエットとストレスを組み合わせることで、過食症ラットのオピオイドに対する反応が誇張されます。 ふるまい。 神経症。 119、1207〜1214。 土井:10.1037 / 0735-7044.119.5.1207
Boothroyd、LG、Jucker、JL、Thornborrow、T.、Jamieson、MA、Burt、DM、Barton、RAなど (2016)。 テレビの露出は、ニカラグアの田舎の理想的な体の大きさを予測します。 Br。 J.サイコ。 107、752–767。 土井:10.1111 / bjop.12184
Borau、S.およびBonnefon、J.(2017)。 架空の性内競争:挑発的な女性モデルを取り上げた広告は、女性が間接的な攻撃に関与するきっかけとなります。 J.バス。 倫理 157:45. doi: 10.1007/s10551-017-3643-y
ボールド、H。、デスタボラ、B。、マグナソン、C。、ミカリ、N。、ダル、H。、エバンス、J。 (2016)。 女子が摂食障害を発症するかどうかに対する学校の影響。 Int。 J.エピデミオール。 45、480–488。 土井:10.1093 / ije / dyw037
ボベット、J。(2019)。 進化論と女性のウエストヒップ比に対する男性の好み:どの仮説が残っていますか? 前面。 サイコロル。 10:1221 土井:10.3389 / fpsyg.2019.01221
Bravo、JA、Forsythe、P.、Chew、MV、Escaravage、E.、Savignac、HM、Dinan、TG、et al。 (2011)。 乳酸oba菌株の摂取は、迷走神経を介してマウスの感情的行動と中枢GABA受容体発現を調節します。 Proc。 Natl。 Acad。 Sci。 アメリカ合衆国 108、16050〜16055。 土井:10.1073 / pnas.1102999108
Brewerton、TD、およびGeorge、MS(1993)。 片頭痛は摂食障害に関連していますか? Int。 J.Eat。 無理。 14, 75–79. doi: 10.1002/1098-108x(199307)14:1<75::aid-eat2260140110>3.0.co;2-d
TDのBrewerton、MSのGeorge、RNのHarden(1993)。 片頭痛と摂食障害。 Psychiatry Res。 46, 201–202. doi: 10.1016/0165-1781(93)90020-h
Brockmeyer、T.、Friederich、H。、およびSchmidt、U。(2017)。 神経性食欲不振の治療の進歩:確立された新たな介入のレビュー。 サイコロ。 メド。 11、1〜37。 土井:10.1017 / S0033291717002604
Bulik、CM、Sullivan、PF、Fear、J。、およびPickering、A.(1997)。 神経性食欲不振症の女性における神経性過食症の発生の予測因子。 J.Nerv。 そうだ Dis。 185, 704–707. doi: 10.1097/00005053-199711000-00009
バートン、A。、およびアボット、M。(2019)。 過食症のプロセスと経路:過食症の統合された認知および行動モデルの開発。 J. Eat。 不調和。 7:18. doi: 10.1186/s40337-019-0248-0
ブスタマンテ、AC、アイエロ、AE、ガレア、S。、ラタナタラソーン、A。、ノローニャ、C。、ワイルドマン、DE、他 (2016)。 グルココルチコイド受容体のDNAメチル化、小児期の虐待および大鬱病。 J.Affect。 無理。 206、181–188。 doi:10.1016 / j.jad.2016.07.038
Butovskaya、M.、Sorokowska、A.、Karwowski、M.、Sabiniewicz、A.、Fedenok、J.、Dronova、D.、et al。 (2017)。 XNUMXつの伝統的な社会におけるウエストヒップ比、ボディマスインデックス、年齢、子供の数。 サイ。 担当者 7:1622. doi: 10.1038/s41598-017-01916-9
カルシア、マサチューセッツ州、ボンソール、DR、ブルームフィールド、PS、セルバラジ、S。、バリチェロ、T。、ハウズ、OD(2016)。 ストレスと神経炎症:ミクログリアに対するストレスの影響と精神疾患への影響の系統的レビュー。 精神薬理学 233, 1637–1650. doi: 10.1007/s00213-016-4218-9
カルゾ、J。、オースティン、S。、およびミカリ、N。(2018)。 英国の思春期の少年少女の摂食障害症状における性的指向の格差。 ユーロ。 子アデレスク。 精神科 27, 1483–1490. doi: 10.1007/s00787-018-1145-9
Carlat、DJ、Camargo、CA、およびHerzog、DB(1997)。 男性の摂食障害:135人の患者に関する報告。 Am。 J.精神医学 154、1127〜1132。 土井:10.1176 / ajp.154.8.1127
Caslini、M.、Bartoli、F.、Crocamo、C.、Dakanalis、A.、Clerici、M.、Carra、G.(2016)。 児童虐待と摂食障害との関連を解き明かす:系統的レビューとメタ分析。 サイコソム メド。 78、79〜90。 土井:10.1097 / psy.0000000000000233
キャシディ、E。、オールソップ、M。、およびウィリアムズ、T。(1999)。 青年期の摂食障害の初期症状での強迫症状。 ユーロ。 子アデレスク。 精神医学 8、193〜199。 土井:10.1007 / s007870050129
カステリーニ、G。、ロサウロ、C。、リッカ、V。、およびレリニ、AH(2017)。 女性の間での過食や性的不満への傾向の一般的な要因としての身体的自尊心:性別中の解離とストレス反応の役割。 J.セックス。 メド。 14、1036〜1045。 土井:10.1016 / j.jsxm.2017.06.001
Caudle、H.、Pang、C.、Mancuso、S.、Castle、D。、およびNewton、R.(2015)。 オーストラリア、ビクトリア州の摂食障害の診断に対するDSM-5の影響に関するレトロスペクティブ研究。 J. Eat。 不調和。 3:35. doi: 10.1186/s40337-015-0072-0
Ceccarini、J.、Weltens、N.、Ly、HG、Tack、J.、Van Oudenhove、L。、およびVan Laere、K.(2016)。 大脳カンナビノイド1受容体の可用性と食物摂取障害患者および健康な被験者の肥満度指数との関係:F-18 MK-9470 PET研究。 トランス 精神医学 6:8。 土井:10.1038 / tp.2016.118
Cederlof、M.、Thornton、LM、Baker、J.、Lichtenstein、P.、Larsson、H.、Ruck、C.、et al。 (2015)。 強迫性障害と神経性食欲不振の間の病因学的重複:縦断的コホート、多世代家族および双生児研究。 世界精神医学 14、333–338。 土井:10.1002 / wps.20251
チャクラバルシー、MV、およびブース、FW(2004)。 食べる、運動する、そして「th約」遺伝子型:現代の慢性疾患の進化的理解に向けて点をつなぐ。 J. Appl。 生理。 96、3–10。 土井:10.1152 / japplphysiol.00757.2003
クラーク、G。、グレナム、S。、スカリー、P。、フィッツジェラルド、P。、モロニー、RD、シャナハン、F。 (2013)。 幼少期のミクロビオーム-腸-脳軸は、性依存的に海馬のセロトニン系を調節します。 Mol。 精神医学 18、666〜673。 土井:10.1038 / mp.2012.77
Corbett、S.、Courtiol、A.、Lummaa、V.、Moorad、J.、およびStearns、S.(2018)。 現代性と慢性疾患への移行:ミスマッチと自然selection。 Nat。 Rev. Genet。 19, 419–430. doi: 10.1038/s41576-018-0012-3
クリスプ、A。(1983)。 「神経性食欲不振症の精神病理学のいくつかの側面」 神経性食欲不振:研究の最近の進展、eds P. Darby、P。Garfinkel、D。Garner、およびD. Coscina、(ニューヨーク、NY:Guildford Press。)、15–28。
Cuesto、G.、Everaerts、C.、Leon、LG、およびAcebes、A.(2017)。 神経性食欲不振症、神経性過食症、過食症の分子基盤:闇に光を当てる。 J. Neurogenet。 31、266〜287。 土井:10.1080 / 01677063.2017.1353092
D'Andrea、G.、Ostuzzi、R.、Francesconi、F.、Musco、F.、Bolner、A.、d'Onofrio、F.、et al。 (2009)。 摂食障害における片頭痛有病率と病態生理学的相関。 Neurol。 サイ。 30, S55–S59. doi: 10.1007/s10072-009-0070-6
JWのDalleyおよびJP(2012年)のRoiser。 ドーパミン、セロトニン、衝動性。 神経科学 215、42〜58。 土井:10.1016 / j.neuroscience.2012.03.065
ダルトン、B。、バーソルディ、S。、ロビンソン、L。、ソルミ、M。、イブラヒム、MAA、ブリーン、G。、他 (2018)。 摂食障害におけるサイトカイン濃度のメタ分析。 J.精神医学 103、252〜264。 土井:10.1016 / j.jpsychires.2018.06.002
Dantzer、R.(2009)。 サイトカイン、病気の行動、およびうつ病。 Immunol。 アレルギー治療室。 北アム。 29、247–264。 土井:10.1016 / j.iac.2009.02.002
de Clercq、NC、Frissen、MN、Davids、M.、Groen、AK、およびNieuwdorp、M.(2019)。 神経性食欲不振からの臨床的回復後の低体重の再発患者における糞便微生物叢移植後の体重増加。 サイコザー。 サイコサム。 88、58〜60。 土井:10.1159 / 000495044
デルジュディツェ、M。、エリス、BJ、およびシャツクリフ、EA(2011)。 ストレス応答性の適応キャリブレーションモデル。 神経科学。 バイオ行動。 牧師 35、1562〜1592。 土井:10.1016 / j.neubiorev.2010.11.007
デル・ゾット、M。、ペグナ、AJ(2017)。 ウエストとヒップの比率が異なる人間の女性の身体の知覚された性的魅力の電気生理学的証拠。 コグ 影響します。 ふるまい。 神経症。 17, 577–591. doi: 10.3758/s13415-017-0498-8
Dignon、A.、Beardsmore、A.、Spain、S。、およびKuan、A。(2006)。 「なぜ私は食べないのか」–彼らの障害の原因に関する15の食欲不振からの患者の証言。 J. Health Psychol。 11、942〜956。 土井:10.1177 / 1359105306069097
Dold、M.、Aigner、M.、Klabunde、M.、Treasure、J。、およびKasper、S.(2015)。 神経性食欲不振症の第二世代の抗精神病薬:ランダム化比較試験のメタ分析。 サイコザー。 サイコサム。 84、110〜116。 土井:10.1159 / 000369978
Esalatmanesh、S.、Abrishami、Z.、Zeinoddini、A.、Rahiminejad、F.、Sadeghi、M.、Najarzadegan、MR、et al。 (2016)。 中等度から重度の強迫性障害におけるミノサイクリンとフルボキサミンの併用療法:プラセボ対照、二重盲検、無作為化試験。 精神科クリニック。 Neurosci。 70、517–526。 doi:10.1111 / pcn.12430
エバンス、GW、およびフラー・ローウェル、TE(2013)。 小児期の貧困、慢性的ストレス、および若年成人の作業記憶:自己調節能力の保護的役割。 開発。 科学 16、688–696。 土井:10.1111 / desc.12082
Faer、LM、Hendriks、A.、Abed、RT、and Figueredo、AJ(2005)。 摂食障害の進化心理学:配偶者または地位をめぐる女性の競争? サイコ。 サイコザー。 理論解像度 練習。 78、397–417。 土井:10.1348 / 147608305×42929
Farook、JM、Lewis、B.、Littleton、JM、およびBarron、S.(2009)。 トピラマートは、オスのC57BL / 6Jマウスのストレスによるアルコール消費の増加と嗜好を軽減します。 フィジオル。 ふるまい 96、189〜193。 土井:10.1016 / j.physbeh.2008.08.011
Findley、DB、Leckman、JF、Katsovich、L.、Lin、H.、Zhang、H.、Grantz、H.、et al。 (2003)。 エールの子供のグローバルストレスインデックス(YCGSI)の開発と、トゥレット症候群および強迫性障害の子供および青年におけるその適用。 J. Am。 Acad。 子アデレスク。 精神医学 42、450〜457。 土井:10.1097 / 01.chi.0000046816.95464.ef
フレッチャー、PJ、シンヤード、J。、およびヒギンズ、ジョージア(2010)。 5-HT2C受容体の活性化は抑制ではなく活性化であるという遺伝的および薬理学的証拠は、強化の漸進的比率スケジュールの下で摂食する動機付けに影響します。 Pharmacol。 Biochem。 Behav。 97、170〜178。 土井:10.1016 / j.pbb.2010.07.002
Foster、JA、Rinaman、L。、およびCryan、JF(2017)。 ストレスと脳腸軸:マイクロバイオームによる調節。 ニューロバイオオール。 ストレス 7、124〜136。 土井:10.1016 / j.ynstr.2017.03.001
フランク、GKW、デグズマン、MC、およびショット、ME(2019)。 食べる動機と食べない動機–神経性食欲不振症における心理生物学的葛藤。 フィジオル。 ふるまい 206、185〜190。 土井:10.1016 / j.physbeh.2019.04.007
フレデリック、DA、およびハセルトン、MG(2007)。 なぜ筋肉質はセクシーなのですか? フィットネスインジケーター仮説の検定。 パーソナル。 Soc。 サイコ。 ブル。 33、1167〜1183。 土井:10.1177 / 0146167207303022
Furnham、A.、およびBaguma、P.(1994)。 男性と女性の身体形状の評価における異文化間の違い。 Int ,. J. Eat。 不調和。 15, 81–89. doi: 10.1002/1098-108x(199401)15:1<81::aid-eat2260150110>3.0.co;2-d
Galusca、B.、Sigaud、T.、Costes、N.、Redoute、J.、Massoubre、C。、およびEstour、B.(2014)。 脳セロトニン作動性活動の広範な障害が、神経性過食症患者における個人間の不均一性:パイロットF-18 MPPF / PET研究。 Word J. Biol。 精神科 15、599〜608。 土井:10.3109 / 15622975.2014.942358
Gao、XH、Cao、QH、Cheng、Y.、Zhao、DD、Wang、Z.、Yang、HBなど (2018)。 慢性ストレスは、腸内細菌叢を乱し、免疫系の反応を引き起こすことにより大腸炎を促進します(vol 115、pg E2960、2018)。 Proc。 Natl。 Acad。 Sci。 アメリカ合衆国 115、E4542–E4542。 土井:10.1073 / pnas.1806622115
ガルシア-ガルシア、アラバマ州、メン、QY、カネッタ、S。、ガルディア、AM、ギヤード、BP、ケレンドンク、C。 (2017)。 思春期の前頭前野5-HT1A受容体を介したセロトニンシグナル伝達は、ベースラインの気分関連行動を決定することができます。 細胞代表 18、1144–1156。 土井:10.1016 / j.celrep.2017.01.021
ガルシア-ソリアーノ、G。、ロンセロ、M。、ペルピナ、C。、およびベロック、A。(2014)。 強迫性障害および摂食障害患者における侵入的思考:差異分析。 ユーロ。 食べる。 無理。 牧師 22、191–199。 土井:10.1002 / erv.2285
Goldschmidt、AB、Le Grange、D.、Powers、P.、Crow、SJ、Hill、LL、Peterson、CB、他 (2011)。 通常の体重と過食症の肥満患者の摂食障害の症候学。 肥満 19、1515〜1518。 土井:10.1038 / oby.2011.24
グロスマン、RE、ズガイエ、SM、リュー、S。、ライルズ、RH、およびタンプリチャ、V(2012)。 肺の増悪で入院した嚢胞性線維症の成人の炎症マーカーに対するビタミンD補給の影響。 ユーロ。 J. Clin。 ナッツ 66、1072〜1074。 土井:10.1038 / ejcn.2012.82
Guisinger、S.(2003)。 飢amineからの逃避に適応:神経性食欲不振症の進化的展望を追加。 サイコロル。 牧師 110, 745–761. doi: 10.1037/0033-295x.110.4.745
ハフストロム、I。、リンガーツ、B。、ギレンハンマール、H。、パームブラッド、J。、およびハームスリングダール、M。(1988)。 関節リウマチ患者の疾患活動性、好中球機能、脂肪酸組成、ロイコトリエン生合成に対する絶食の影響。 関節炎のRheum。 31、585〜592。 土井:10.1002 / art.1780310502
Hagan、MM、Chandler、PC、Wauford、PK、Rybak、RJ、およびOswald、KD(2003)。 ストレス誘発性の過食の動物モデルにおける引き金となる要因としての美味な食物と空腹の役割。 Int。 J.Eat。 無理。 34、183〜197。 土井:10.1002 / eat.10168
Hagan、MM、Wauford、PK、Chandler、PC、Jarrett、LA、Rybak、RJ、およびBlackburn、K.(2002)。 過食症の新しい動物モデル:過去のカロリー制限とストレスの重要な相乗的役割。 フィジオル。 ふるまい 77, 45–54. doi: 10.1016/s0031-9384(02)00809-0
メリーランド州ヘイル、A。シェカー、カリフォルニア州ローリー(2012)。 ストレス関連セロトニン系:不安および情動障害の症候学への影響。 細胞。 Mol。 Neurobiol。 32, 695–708. doi: 10.1007/s10571-012-9827-1
ハリーム、DJ(2012)。 神経性食欲不振症におけるセロトニン神経伝達。 ふるまい。 Pharmacol。 23, 478–495. doi: 10.1097/FBP.0b013e328357440d
Haleem、DJ、およびHaider、S.(1996)。 食物制限により、視床下部におけるセロトニンとその合成速度が低下します。 神経報告 7, 1153–1156. doi: 10.1097/00001756-199604260-00011
Halmi、KA、Tozzi、F.、Thornton、LM、Crow、S.、Fichter、MM、Kaplan、ASなど (2005)。 摂食障害のある個人における完全主義、強迫性人格障害および強迫性障害の関係。 Int。 J.Eat。 無理。 38、371〜374。 土井:10.1002 / eat.20190
ハリソン、A。、スタブリ、P。、オーモンド、L。、マケネミー、F.、Akyol、D。、およびAl-Khairulla、H。(2018)。 重度で複雑な神経性食欲不振の思春期入院患者に対する認知改善療法:治療試験。 ユーロ。 食べる。 無理。 牧師 26、230–240。 土井:10.1002 / erv.2584
Hay、PPJ、Bacaltchuk、J.、Stefano、S。、およびKashyap、P.(2009)。 神経性過食症および過敏症の心理的治療。 Cochrane Database Syst。 牧師 7:CD000562. doi: 10.1002/14651858.CD000562.pub3
Hedman、A.、Breithaupt、L.、Hübel、C.、Thornton、LM、Tillander、A.、Norring、C.、et al。 (2019)。 摂食障害と自己免疫疾患の双方向の関係。 J.チャイルドサイコ。 精神医学 60、803–812。 土井:10.1111 / jcpp.12958
ヒギンズ、ジョージア州、シレニエクス、LB、ラウ、W。、デラノイ、IAM、リー、DKH、イザコワ、J。 (2013)。 化学的に多様な5-HT2C受容体アゴニストの、食物とニコチンによる動機付けおよび副作用プロファイルに関する評価。 精神薬理学 226, 475–490. doi: 10.1007/s00213-012-2919-2
Himmerich、H.、Au、K.、Dornik、J.、Bentley、J.、Schmidt、U。、およびTreasure、J.(2017)。 神経性食欲不振症患者に対するオランザピン治療。 できる。 J.精神医学、レビュー缶。 精神科医 62、506〜507。 土井:10.1177 / 0706743717709967
Hofmeijer-Sevink、MK、van Oppen、P.、van Megen、HJ、Batelaan、NM、Cath、DC、van der Wee、NJA、et al。 (2013)。 強迫性障害における併存疾患の臨床的関連性:オランダOCD協会の研究。 J.Affect。 無理。 150、847–854。 doi:10.1016 / j.jad.2013.03.014
Hoppen、T.、およびChalder、T.(2018)。 成人期における情動障害の診断横断的危険因子としての小児期の逆境:生物心理社会的調整および媒介変数に焦点を当てた系統的レビュー。 クリン サイコロル。 牧師 65、81〜151。 土井:10.1016 / j.cpr.2018.08.002
Joy、E.、Kussman、A。、およびNattiv、A。(2016)。 アスリートの摂食障害に関する2016年の更新:臨床評価と管理に焦点を当てた包括的な物語のレビュー。 Br。 J.スポーツ医学 50、154–162。 土井:10.1136 / bjsports-2015-095735
カラジア、BT、ムルネン、SK、およびティルカ、TL(2017)。 身体の不満は時間とともに変化しますか? 時間をまたいだメタ分析。 サイコロ。 ブル。 143、293–320。 土井:10.1037 / bul0000081
Katz、RL、Keen、CL、Litt、IF、Hurley、LS、Kellamsharrison、KM、およびGlader、LJ(1987)。 神経性食欲不振症における亜鉛欠乏。 J.Adolesc。 健康 8, 400–406. doi: 10.1016/0197-0070(87)90227-0
Kaye、WH、Barbarich、NC、Putnam、K.、Gendall、KA、Fernstrom、J.、Fernstrom、M.、et al。 (2003)。 神経性食欲不振症における急性トリプトファン枯渇の抗不安作用。 Int。 J.Eat。 無理。 33、257〜267。 土井:10.1002 / eat.10135
ケイ、WH、フランク、GK、メルツァー、CC、プライス、JC、マコナハ、CW、クロッサン、PJ、他 (2001)。 神経性過食症から回復した女性のセロトニン2A受容体活性の変化。 Am。 J.精神医学 158、1152〜1155。 土井:10.1176 / appi.ajp.158.7.1152
ケイ、WH、ファッジ、JL、およびパウルス、M。(2009)。 神経性食欲不振の症状と神経回路機能に対する新しい洞察。 Nat。 Neurosci教授。 10、573〜584。 土井:10.1038 / nrn2682
ケイ、WH、Gwirtsman、HE、George、DT、およびEbert、MH(1991)。 長期体重回復後の神経性食欲不振症におけるセロトニン活性の変化:脳脊髄液5-ヒドロキシインドール酢酸レベルの上昇は硬直的および強迫行動と相関するか? アーチ。 一般精神医学 48、556-562。
ケイ、WH、Gwirtsman、HE、George、DT、Jimerson、DC、およびEbert、MH(1988)。 神経性食欲不振症におけるCSF 5-HIAA濃度:体重が減少した被験者の体重減少は体重増加後に正常化する。 Biol。 精神医学 23, 102–105. doi: 10.1016/0006-3223(88)90113-8
ケイ、WH、Weltzin、T。、およびHsu、LKG(1993)。 神経性食欲不振と強迫行動および強迫行動との関係。 精神医学アン。 23, 365–373. doi: 10.3928/0048-5713-19930701-07
キール、PK、Klump、KL、Miller、KB、McGue、M。、およびIacono、WG(2005)。 摂食障害および不安障害の共有伝播。 Int。 J.Eat。 無理。 38、99〜105。 土井:10.1002 / eat.20168
Keeney、A.、Jessop、DS、Harbuz、MS、Marsden、CA、Hogg、S。、およびBlackburn-Munro、RE(2006)。 マウスの視床下部-下垂体-副腎系機能および海馬セロトニン放出に対する急性および慢性社会的敗北ストレスの異なる効果。 J.Neuro endocrinol。 18、330〜338。 土井:10.1111 / j.1365-2826.2006.01422.x
Keski-Rahkonen、A。、およびMustelin、L.(2016)。 ヨーロッパの摂食障害の疫学:有病率、発生率、併存疾患、経過、結果、および危険因子。 Curr。 意見 精神科 29、340〜345。 土井:10.1097 / yco.0000000000000278
Khademian、M.、Farhangpajouh、N.、Shahsanaee、A.、Bahreynian、M.、Mirshamsi、M。、およびKelishadi、R.(2014)。 小児の食欲不振のサブスケールに対する亜鉛補給の効果:ランダム化比較試験。 パキスタンJ. Med。 科学 30、1213–1217。 土井:10.12669 / pjms.306.6377
Khani、S。、およびTayek、JA(2001)。 コルチゾールは、ヒトの糖新生を増加させる:メタボリックシンドロームにおけるその役割。 臨床。 科学 101、739–747。 土井:10.1042 / cs20010180
Klatzkin、RR、Gaffney、S.、Cyrus、K.、Bigus、E。、およびBrownley、KA(2018)。 過食症と肥満の女性のストレス誘発性食。 Biol。 サイコロ。 131、96〜106。 土井:10.1016 / j.biopsycho.2016.11.002
Kleiman、SC、Watson、HJ、Bulik-Sullivan、EC、Huh、EY、Tarantino、LM、Bulik、CMなど (2015)。 急性神経性食欲不振症および栄養回復期の腸内微生物叢:うつ病との関係。 不安、および摂食障害の精神病理学。 サイコソム メド。 77、969〜981。 土井:10.1097 / psy.0000000000000247
Klump、KL、Culbert、KM、Sisk、CL(2017)。 過食症の性差:発達中の性ホルモンの影響。 アンヌ。 牧師クリニック サイコロル。 13、183–207。 土井:10.1146 / annurev-clinpsy-032816-045309
Koduah、P.、Paul、F。、およびDorr、JM(2017)。 神経変性および神経炎症性疾患の予防、予測、治療におけるビタミンD。 エプマ・J 8, 313–325. doi: 10.1007/s13167-017-0120-8
Konuk、N.、Tekin、IO、Ozturk、U.、Atik、L.、Atasoy、N.、Bektas、S。、他 (2007)。 強迫性障害における腫瘍壊死因子αおよびインターロイキン-6の血漿レベル。 中 炎症。 2007:65704。
Krams、I.、Rantala、MJ、Luoto、S。、およびKrama、T.(2018)。 脂肪は単なるエネルギー貯蔵庫ではありません。 J. Exp。 バイオ 221(Pt 12):jeb183756。 土井:10.1242 / jeb.183756
クラムズ、IA、ケッコ、S.、Joers、P.、Trakimas、G.、Elferts、D.、Krams、R。、他 (2017)。 ミクロビオーム共生生物と食餌の多様性は、昆虫の幼虫の免疫系にコストをかけます。 J. Exp。 バイオ 220、4204-4212。 土井:10.1242 / jeb.169227
クディエルカ、BM、およびキルシュバウム、C。(2005)。 ストレスに対するHPA軸反応の性差:レビュー。 Biol。 サイコロ。 69、113〜132。 土井:10.1016 / j.biopsycho.2004.11.009
ラセック、WD、およびガウリン、S。(2019)。 人間の女性の肉体的魅力の鍵としての不安定性と再現性のある価値を裏付ける証拠。 進化。 ハム。 行動.. doi:10.1016 / j.evolhumbehav.2019.05.001 [印刷前にEpub]。
Lebow、J.、Chuy、JA、Cedermark、K.、Cook、K.、およびSim、LA(2015)。 トピラメート開始後の摂食障害症状の発症または悪化。 小児科 135、E1312–E1316。 土井:10.1542 / peds.2014-3413
Legenbauer、T.、Thiemann、P。、およびVocks、S。(2014)。 摂食障害のある小児および青年の身体イメージ障害現在の証拠と将来の方向。 Z.キンダーユーゲント精神科医。 サイコザー。 42, 51–59. doi: 10.1024/1422-4917/a000269
Leone、A.、Martinez-Gonzalez、MA、Lahortiga-Ramos、F.、Santos、PM、Bertoli、S.、Batttezzati、A。など (2018)。 地中海の食事パターンの順守と、女性における食欲不振および神経性過食症の発生率:SUNコホート。 栄養 54、19〜25。 土井:10.1016 / j.nut.2018.02.008
Lewis、DMG、Al-Shawaf、L.、Conroy-Beam、D.、Asao、K。、およびBuss、DM(2017)。 進化心理学:ハウツーガイド。 午前 サイコロル。 72、353〜373。 土井:10.1037 / a0040409
Li、NP、Smith、AR、Griskevicius、V.、Cason、MJ、およびBryan、A。(2010)。 異性愛者および同性愛者における性内競争および摂食制限。 進化。 ハム。 行動。 31、365–372。 土井:10.1016 / j.evolhumbehav.2010.05.004
Li、NP、Smith、AR、Yong、JC、およびBrown、TA(2014)。 「性内競争および摂食制限に関するその他の理論」 人間の性的心理学と行動に関する進化的展望。 進化心理学、V。Weekes-Shackelford編、T。Shackelford編(ニューヨーク、NY:Springer)。 土井:10.1016 / j.evolhumbehav.2010.05.004
Li、NP、van Vugt、M。、およびColarelli、SM(2018)。 進化的ミスマッチ仮説:心理学への影響。 Curr。 ええ。 サイコロ。 サイ。 27、38〜44。 土井:10.1177 / 0963721417731378
Liang、S.、Wu、XL、およびJin、F.(2018)。 腸脳心理学:微生物叢-腸脳軸から心理学を再考する。 前面。 統合。 神経科学。 12:24。 土井:10.3389 / fnint.2018.00033
Lissemore、JI、Sookman、D.、Gravel、P.、Berney、A.、Barsoum、A.、Diksic、M。など (2018)。 強迫性障害における脳セロトニン合成能力:認知行動療法とセルトラリンの効果。 トランス 精神医学 8:82. doi: 10.1038/s41398-018-0128-4
ラブ、H。、スリコウスキー、D。(2018)。 肉と男性について:肉に対する暗黙的および明示的な態度の性差。 前面。 サイコロル。 9:559 土井:10.3389 / fpsyg.2018.00559
Lukens、JR、Gurung、P.、Vogel、P.、Johnson、GR、Carter、RA、McGoldrick、DJなど (2014)。 ミクロビオームの食事調節は、自己炎症性疾患に影響します。 自然 516、246〜249。 土井:10.1038 / nature13788
Luoto、S.、Karlsson、H.、Krams、I。、およびRantala、M.(2018)。 進化的精神医学に基づいたうつ病のサブタイピング:反応性短期気分の変化からうつ病まで。 脳の行動。 イミュン。 69:630。 土井:10.1016 / j.bbi.2017.10.012
ルオト、S。(2019a)。 人間の性選択の最新の理論的枠組み:生態学、遺伝学、生活史から表現型の拡張まで。 適応する。 ハム。 行動。 生理。 5, 48–102. doi: 10.1007/s40750-018-0103-6
ルオト、S。(2019b)。 解説への応答:生活史遺伝学、流体知能、および拡張表現型。 適応する。 ハム。 行動。 生理。 5, 112–115. doi: 10.1007/s40750-019-0109-8
Luoto、S.、I。Krams、およびMJ Rantala(2019a)。 女性の性的指向スペクトルへのライフヒストリーアプローチ:進化、発達、原因メカニズム、および健康。 アーチ。 セックス。 ベハ。 48, 1273–1308. doi: 10.1007/s10508-018-1261-0
Luoto、S.、Krams、I。、およびRantala、MJ(2019b)。 解説への反応:生活史の進化、原因メカニズム、女性の性的指向。 アーチ。 セックス ふるまい。 48, 1335–1347. doi: 10.1007/s10508-019-1439-0
マチャド、PPP、Goncalves、S。、およびHoek、HW(2013)。 DSM-5は、ednosケースの割合を減らします。コミュニティのサンプルからの証拠です。 Int。 J.Eat。 無理。 46、60〜65。 土井:10.1002 / eat.22040
Mack、I.、Cuntz、U.、Gramer、C.、Niedermaier、S.、Pohl、C.、Schwiertz、A.、et al。 (2016)。 神経性食欲不振症の体重増加は、糞便微生物叢、分岐鎖脂肪酸プロファイル、および胃腸のcomplaint訴を改善しません。 サイ。 担当者 6、26752. doi:10.1038 / srep26752
Malhotra、R.(2016)。 片頭痛の理解:神経性炎症の潜在的な役割。 アン。 Indian Acad。 神経細胞 19、175〜182。 土井:10.4103 / 0972-2327.182302
Mancuso、SG、Newton、JR、Bosanac、P.、Rossell、SL、Nesci、JB、Castle、DJ(2015)。 摂食障害の分類:DSM-IVおよびDSM-5基準を使用した相対有病率の比較。 Br。 J.精神医学 206、519–520。 doi:10.1192 / bjp.bp.113.143461
Mann、T.、Tomiyama、AJ、Westling、E.、Lew、AM、Samuels、B。、およびChatman、J。(2007)。 効果的な肥満治療のためのメディケアの探求–ダイエットは答えではありません。 午前 サイコロル。 62, 220–233. doi: 10.1037/0003-066x.62.3.220
Martin、JR、Bos、M.、Jenck、F.、Moreau、JL、Mutel、V.、Sleight、AJ、et al。 (1998)。 5-HT2C受容体アゴニスト:薬理学的特性と治療の可能性。 J. Pharmacol。 Exp。 Ther。 286、913-924。
マシュエブ、RM、グリロ、CM、ホワイト、MA(2011)。 神経性過食症と過食症のある地域の女性の食事パターンの検査。 Int。 J.Eat。 無理。 44、618〜624。 土井:10.1002 / eat.20853
Mayhew、AJ、Pigeyre、M.、Couturier、J。、およびMeyre、D.(2018)。 摂食障害の進化的遺伝的展望。 神経内分泌学 106、292〜306。 土井:10.1159 / 000484525
McElroy、SL、Guerdjikova、AI、Mori、N。、およびO'Melia、AM(2012)。 過食症の薬理学的管理:現在および今後の治療選択肢。 サー 臨床。 リスク管理。 8、219〜241。 土井:10.2147 / tcrm.s25574
ミラネースキ、Y。、シモンズ、WK、ヴァンロッサム、EFC、およびペニンクス、BW(2018)。 うつ病と肥満:生物学的メカニズムの共有の証拠。 Mol。 精神医学 24, 18–33. doi: 10.1038/s41380-018-0017-5
Mitchell、JE、Roerig、J.およびSteffen、K.(2013)。 Biological therapies for eating disorders.摂食障害のための生物学的療法。 Int。 J.Eat。 無理。 46、470〜477。 土井:10.1002 / eat.22104
Molina-Torres、G.、Rodriguez-Arrastia、M.、Roman、P.、Sanchez-Labraca、N.、およびCardona、D.(2019)。 Stress and the gut microbiota-brain axis.ストレスと腸内細菌叢-脳軸。 ふるまい。 Pharmacol。 30、187〜200。 doi: 10.1097/FBP.0000000000000478土井:XNUMX / FBP.XNUMX
Mowla、A.、Khajeian、AM、Sahraian、A.、Chohedri、AH、and Kashkoli、F.(2010)。 Topiramate augmentation in resistant OCD: a double-blind placebo-controlled clinical trial.耐性OCDにおけるトピラマートの増強:二重盲検プラセボ対照臨床試験。 CNSスペクトラム 15、613〜617。 土井:10.1017 / s1092852912000065
Mueller, AS, Pearson, J., Muller, C., Frank, K., and Turner, A. (2010).ミューラー、AS、ピアソン、J。、ミュラー、C。、フランク、K。、およびターナー、A。(XNUMX)。 Sizing up peers: adolescent girls' weight control and social comparison in the school context.仲間のサイジング:学校における青年期の女の子の体重管理と社会的比較。 J. Health Soc。 行動。 51、64〜78。 土井:10.1177 / 0022146509361191
Murray, SB, Quintana, DS, Loeb, KL, Griffiths, S., and Le Grange, D. (2019).マレー、SB、キンタナ、DS、ローブ、KL、グリフィス、S、およびルグランジ、D(XNUMX)。 Treatment outcomes for anorexia nervosa: a systematic review and meta-analysis of randomized controlled trials.神経性食欲不振の治療結果:ランダム化比較試験の系統的レビューとメタ分析。 サイコロ。 メド。 49、535〜544。 土井:10.1017 / S0033291718002088
Mustelin、L.、Bulik、CM、Kaprio、J.、and Keski-Rahkonen、A.(2017) Prevalence and correlates of binge eating disorder related features in the community.コミュニティにおける過食症関連の特徴の有病率と相関。 食欲 109、165〜171。 土井:10.1016 / j.appet.2016.11.032
Naisbitt、C。、およびDavies、S。(2017)。 Starvation, exercise and the stress response.飢餓、運動、ストレス反応。 Anaesth.アナエス。 Intensive Care Med.集中治療メッド。 18、508〜512。 doi: 10.1016/j.mpaic.2017.06.020土井:XNUMX / j.mpaic.XNUMX
Najjar、S.、Pearlman、DM、Alper、K.、Najjar、A。、およびDevinsky、O。(2013)。 Neuroinflammation and psychiatric illness.神経炎症および精神疾患。 J. Neuroinflamm。 10:43. doi: 10.1186/1742-2094-10-43
Nave、G.、Nadler、A.、Dubois、D.、Zava、D.、Camerer、C.およびPlassmann、H.(2018)。 Single-dose testosterone administration increases men's preference for status goods.単回投与のテストステロン投与は、ステータス商品に対する男性の好みを高めます。 Nat。 コミュニ 9:2433. doi: 10.1038/s41467-018-04923-0
Nettersheim、J.、Gerlach、G.、Herpertz、S.、Abed、R.、Figueredo、A.およびBrüne、M.(2018)。 Evolutionary psychology of eating disorders: an explorative study in patients with anorexia nervosa and bulimia nervosa.摂食障害の進化心理学:神経性食欲不振および神経性過食症の患者における探索的研究。 前面。 サイコロル。 9:2122 土井:10.3389 / fpsyg.2018.02122
Ni、J、Shen、TCD、Chen、EZ、Bittinger、K.、Bailey、A.、Roggiani、M。他(2017)。 A role for bacterial urease in gut dysbiosis and Crohn's disease.腸内細菌叢異常症およびクローン病における細菌ウレアーゼの役割。 Sci.サイエンス。 Trans.トランス。 Med.中 9:eaah6888。 doi: 10.1126/scitranslmed.aah6888土井:XNUMX / scitranslmed.aahXNUMX
O'Mahony、SM、Neufeld、KAM、Waworuntu、RV、Berg、BM、Dinan、TG、およびCryan、JF(2016)。 A combination of dietary prebiotics and the probiotic LGG modulate behavioural and cognitive reponses to early life stress.食事のプレバイオティクスとプロバイオティクスLGGの組み合わせは、初期の生活ストレスに対する行動反応と認知反応を調節します。 Neurogastroenterol。 Motil.モチル。 28, 13–13. doi: 10.1111/j.2042-7166.2005.tb00466.x
Olguin、P.、Fuentes、M.、Gabler、G.、Guerdjikova、AI、Keck、PE、およびMcElroy、SL(2017)。 Medical comorbidity of binge eating disorder.過食症の医学的併存症。 食べる。 体重の無駄。 22, 13–26. doi: 10.1007/s40519-016-0313-5
Oliver、G。、およびWardle、J。(1999)。 Perceived effects of stress on food choice.食物選択に対するストレスの知覚された影響。 フィジオル。 ふるまい 66, 511–515. doi: 10.1016/s0031-9384(98)00322-9
Orth、U。、およびRobins、RW(2013)。 Understanding the link between low self-esteem and depression.低い自尊心とうつ病との関連を理解する。 Curr。 ええ。 サイコロ。 サイ。 22、455〜460。 土井:10.1177 / 0963721413492763
Pan、WH、Wu、XJ、He、Y.、Hung、HC、Huang、EYK、Mishra、PK、etal。 (2013)。 Brain interleukin-15 in neuroinflammation and behavior.神経炎症および行動における脳インターロイキン-XNUMX。 Neurosci。 Biobehav。 Rev. 37、184〜192。 土井:10.1016 / j.neubiorev.2012.11.009
Park、C.、Brietzke、E.、Rosenblat、JD、Musial、N.、Zuckerman、H.、Ragguett、RM、etal。 (2018)。 Probiotics for the treatment of depressive symptoms: an anti-inflammatory mechanism?うつ症状の治療のためのプロバイオティクス:抗炎症メカニズム? 脳の行動。 イミュン。 73、115–124。 土井:10.1016 / j.bbi.2018.07.006
Partrick、KA、Chassaing、B.、Beach、LQ、McCann、KE、Gewirtz、AT、およびHuhman、KL(2018)。 Acute and repeated exposure to social stress reduces gut microbiota diversity in Syrian hamsters (vol 345, pg 39, 2018).社会的ストレスへの急性および反復暴露は、シリアのハムスターの腸内細菌叢の多様性を減少させます(vol XNUMX、pg XNUMX、XNUMX)。 Behavi.行動。 Brain Res.脳の解像度。 348、277〜277。 土井:10.1016 / j.bbr.2018.03.044
Pearl, RL, White, MA, and Grilo, CM (2014).パール、RL、ホワイト、MA、およびグリロ、CM(XNUMX)。 Overvaluation of shape and weight as a mediator between self-esteem and weight bias internalization among patients with binge eating disorder.過食症患者の自尊心と体重バイアスの内面化の間の仲介者としての形状と体重の過大評価。 食べる。 ふるまい。 15、259〜261。 土井:10.1016 / j.eatbeh.2014.03.005
Perkins、SJ、Keville、S.、Schmidt、U。、およびChalder、T。(2005)。 Eating disorders and irritable bowel syndrome: is there a link?摂食障害と過敏性腸症候群:関連性はありますか? J.サイコソム。 解像度 59、57–64。 doi: 10.1016/j.jpsychores.2004.04.375土井:XNUMX / j.jpsychores.XNUMX
Pinheiro、RMC、de Lima、MNM、Portal、BCD、Busato、SB、Falavigna、L.、Ferreira、RD、etal。 (2015)。 Long-lasting recognition memory impairment and alterations in brain levels of cytokines and BDNF induced by maternal deprivation: effects of valproic acid and topiramate.長期にわたる認識記憶障害および母体の剥奪によって誘発されるサイトカインおよびBDNFの脳レベルの変化:バルプロ酸およびトピラメートの影響。 J.神経伝達 122, 709–719. doi: 10.1007/s00702-014-1303-2
Power, ML (2012).パワー、ML(XNUMX)。 The human obesity epidemic, the mismatch paradigm, and our modern “captive” environment.人間の肥満の蔓延、ミスマッチのパラダイム、そして現代の「捕らえられた」環境。 J.ハム。 Biol。 24、116〜122。 doi: 10.1002/ajhb.22236土井:XNUMX / ajhb.XNUMX
Price, AE, Anastasio, NC, Stutz, SJ, Hommel, JD, and Cunningham, KA (2018).価格、AE、アナスタシオ、ノースカロライナ、スタッツ、SJ、オメル、JD、およびカニンガム、KA(5)。 Serotonin 2-HTXNUMXc receptor activation suppresses binge intake and the reinforcing and motivational properties of high-fat food.セロトニンXNUMX-HTXNUMXc受容体の活性化は、高脂肪食品のビンジ摂取と強化および動機付けの特性を抑制します。 前面。 薬理学。 9:821。 doi:10.3389 / fphar.2018.00821
Quinton、SJ、Smith、AR、およびJoiner、T。(2011)。 The 2nd to 4th digit ratio (2D:4D) and eating disorder diagnosis in women.女性のXNUMX〜XNUMX桁の比率(XNUMXD:XNUMXD)と摂食障害の診断。 パーソナル。 個別。 異なる。 51、402–405。 doi:10.1016 / j.paid.2010.07.024
Rantala、M.、Luoto、S。、およびKrams、I。(2017)。 An evolutionary approach to clinical pharmacopsychology.臨床薬理心理学への進化的アプローチ。 サイコザー。 サイコサム。 86、370〜371。 土井:10.1159 / 000480709
Rantala、M.、Luoto、S.、Krams、I。、およびKarlsson、H。(2018)。 Depression subtyping based on evolutionary psychiatry: proximate mechanisms and ultimate functions.進化精神医学に基づくうつ病のサブタイピング:近接メカニズムと究極の機能。 Brain, Behav.脳、行動。 Immun.イミュン。 69、603–617。 土井:10.1016 / j.bbi.2017.10.012
Rohleder、N。(2019)。 Stress and inflammation – the need to address the gap in the transition between acute and chronic stress effect.ストレスと炎症–急性ストレス効果と慢性ストレス効果の間の移行におけるギャップに対処する必要性。 精神神経内分泌学 105、164〜171。 土井:10.1016 / j.psyneuen.2019.02.021
Ricordi、C.、Garcia-Contreras、M。、およびFarnetti、S。(2015)。 Diet and inflammation: possible effects on immunity.食事と炎症:免疫への影響の可能性。 chronic diseases, and life span.慢性疾患、および寿命。 Coll。 Nutrit.栄養素。 34、10〜13。 土井:10.1080 / 07315724.2015.1080101
Riva、G。(2016)。 Neurobiology of anorexia nervosa: serotonin dysfunctions link self-starvation with body image disturbances through an impaired body memory.神経性食欲不振症の神経生物学:セロトニン機能障害は、身体記憶障害を介して自己飢餓と身体イメージ障害を結びつけます。 前面。 ハム。 神経症。 10:600。 土井:10.3389 / fnhum.2016.00600
Rojo、L.、Conesa、L.、Bermudez、O。、およびLivianos、L。(2006)。 Influence of stress in the onset of eating disorders: data from a two-stage epidemiologic controlled study.摂食障害の発症におけるストレスの影響:XNUMX段階の疫学的対照研究からのデータ。 サイコソム メド。 68、628〜635。 doi: 10.1097/01.psy.0000227749.58726.41土井:XNUMX / XNUMX.psy.XNUMX
Rolls, BJ (2017).ロールス、BJ(XNUMX)。 Dietary energy density: applying behavioural science to weight management.食事エネルギー密度:行動科学を体重管理に適用する。 Nutrit.栄養素。 Bull.ブル。 42、246–253。 doi: 10.1111/nbu.12280土井:XNUMX / nbu.XNUMX
Rubio、G.、Jimenez-Arriero、MA、Martinez-Gras、I.、Manzanares、J。、およびPalomo、T。(2006)。 The effects of topiramate adjunctive treatment added to antidepressants in patients with resistant obsessive-compulsive disorder.抵抗性強迫性障害患者の抗うつ薬に追加されたトピラマート補助療法の効果。 J.Clin。 精神病薬。 26、341〜344。 doi: 10.1097/01.jcp.0000220524.44905.9f土井:XNUMX / XNUMX.jcp.XNUMXf
Safai-Kutti、S。(1990)。 Oral zinc supplementation in anorexia nervosa.神経性食欲不振症における経口亜鉛補給。 Acta Psychiatr。 スキャンします。 82, 14–17. doi: 10.1111/j.1600-0447.1990.tb10747.x
Safai-Kutti、S。、およびKutti、J。(1986)。 Zinc supplementation in anorexia nervosa.神経性食欲不振症における亜鉛補給。 J.Clin。 Nutrit.栄養素。 44、581〜582。 doi: 10.1093/ajcn/44.4.581土井:XNUMX / ajcn / XNUMX
Saunders、JF、およびEaton、AA(2018)。 Snaps, selfies, and shares: how three popular social media platforms contribute to the sociocultural model of disordered eating among young women.スナップ、自撮り写真、共有:XNUMXつの人気のソーシャルメディアプラットフォームが、若い女性の摂食障害の社会文化モデルにどのように貢献しているか。 Cyberpsychol。、Behav。 Soc。 Network.通信網。 21、343〜354。 土井:10.1089 / cyber.2017.0713
Sayyah、M.、Olapour、A.、Saeedabad、YS、Parast、RY、and Malayeri、A。(2012) Evaluation of oral zinc sulfate effect on obsessive-compulsive disorder: a randomized placebo-controlled clinical trial.強迫性障害に対する経口硫酸亜鉛効果の評価:無作為化プラセボ対照臨床試験。 栄養 28、892〜895。 土井:10.1016 / j.nut.2011.11.027
Schmidt、U.、Adan、R.、Bohm、I.、Campbell、IC、Dingemans、A.、Ehrlich、S.、etal。 (2016)。 Eating disorders: the big issue.摂食障害:大きな問題。 ランセット精神医学 3, 313–315. doi: 10.1016/s2215-0366(16)00081-x
Schmidt、U.、Oldershaw、A.、Jichi、F.、Sternheim、L.、Startup、H.、McIntosh、V.、etal。 (2012)。 Out-patient psychological therapies for adults with anorexia nervosa: randomised controlled trial.神経性食欲不振症の成人に対する外来患者の心理療法:ランダム化比較試験。 Br。 J.精神医学 201、392–399。 doi:10.1192 / bjp.bp.112.112078
Schwensen、HF、Kan、C.、Treasure、J.、Hoiby、N。、およびSjogren、M。(2018)。 A systematic review of studies on the faecal microbiota in anorexia nervosa: future research may need to include microbiota from the small intestine.神経性食欲不振症の糞便微生物叢に関する研究の系統的レビュー:将来の研究では、小腸からの微生物叢を含める必要があるかもしれません。 食べる。 体重の無駄。 23, 399–418. doi: 10.1007/s40519-018-0499-9
Seitz、J.、Belheouane、M.、Schulz、N.、Dempfle、A.、Baines、JF、およびHerpertz-Dahlmann、B。(2019)。 The impact of starvation on the microbiome and gut-brain interaction in anorexia nervosa.神経性食欲不振症におけるミクロビオームと腸と脳の相互作用に対する飢餓の影響。 前面。 エンドクリノール。 10:41 土井:10.3389 / fendo.2019.00041
Sell、A.、Lukazsweski、AW、およびTownsley、M。(2017)。 Cues of upper body strength account for most of the variance in men's bodily attractiveness.上半身の強さの手がかりは、男性の身体的魅力の変動の大部分を占めています。 手続き R. Soc。 Bバイオル。 サイエンス。 284:20171819。 土井:10.1098 / rspb.2017.1819
Sharon-Granit、Y.、Nassar、A.、Azab、AN、およびKaplanski、J。(2016)。 Effects of olanzapine and valproate on brain inflammation in lipopolysaccharide-treated rats.リポ多糖で治療したラットの脳の炎症に対するオランザピンとバルプロ酸の効果。 Int。 J.Neurophyschopharmacol。 19、64-65。
Shields、GS、Moons、WG、およびSlavich、GM(2017)。 Inflammation, self-regulation, and health: an immunologic model of self-regulatory failure.炎症、自己調節、および健康:自己調節障害の免疫学的モデル。 見通し。 サイコ。 科学 12、588〜612。 土井:10.1177 / 1745691616689091
Sjögren, M. (2017).シェーグレン、M。(XNUMX)。 An update on genetic and serotoneric biomarker findings in Bulimia Nervosa.神経性過食症における遺伝的および血清学的バイオマーカーの所見に関する最新情報。 ECNeurol。 7、107-116。
Slavich、GM、Way、BM、Eisenberger、NI、およびTaylor、SE(2010)。 Neural sensitivity to social rejection is associated with inflammatory responses to social stress.社会的拒絶に対する神経の感受性は、社会的ストレスに対する炎症反応と関連しています。 Proc。 Natl。 Acad。 Sci。 アメリカ合衆国 107、14817〜14822。 土井:10.1073 / pnas.1009164107
Sohn、K。(2016)。 Men's revealed preferences regarding women's ages: evidence from prostitution.女性の年齢に関する男性の明らかにされた好み:売春からの証拠。 進化。 ハム。 行動。 37、272–280。 土井:10.1016 / j.evolhumbehav.2016.01.002
Sokol, MS (2000).ソコル、MS(XNUMX)。 Infection-triggered anorexia nervosa in children: clinical description of four cases.小児における感染誘発性神経性食欲不振症:XNUMX症例の臨床的説明。 J.チャイルドアドレスク。 Psychopharmacol.精神薬理学。 10、133〜145。 doi: 10.1089/cap.2000.10.133土井:XNUMX / cap.XNUMX
Sokol、MS、およびGray、NS(1997)。 Case study: an infection-triggered, autoimmune subtype of anorexia nervosa.ケーススタディ:神経性食欲不振症の感染誘発性自己免疫サブタイプ。 J. Am。 Acad。 子アデレスク。 精神医学 36, 1128–1133. doi: 10.1097/00004583-199708000-00021
Solmi、M.、Santonastaso、P.、Caccaro、R。、およびFavaro、A。(2013)。 A case of anorexia nervosa with comorbid Crohn's disease: beneficial effects of anti-TNF-alpha therapy?クローン病を併発した神経性食欲不振症の症例:抗TNF-α療法の有益な効果? Int。 J.Eat。 無理。 46、639〜641。 土井:10.1002 / eat.22153
Solmi、M.、Veronese、N.、Favaro、A.、Santonastaso、P.、Manzato、E.、Sergi、G.、etal。 (2015)。 Inflammatory cytokines and anorexia nervosa: a meta-analysis of cross-sectional and longitudinal studies.炎症性サイトカインと神経性食欲不振症:横断的および縦断的研究のメタアナリシス。 精神神経内分泌学 51、237〜252。 土井:10.1016 / j.psyneuen.2014.09.031
Song、C.、Merali、Z。、およびAnisman、H。(1999)。 Variations of nucleus accumbens dopamine and serotonin following systemic interleukin-1, interleukin-2 or interleukin-6 treatment.全身インターロイキン-XNUMX、インターロイキン-XNUMXまたはインターロイキン-XNUMX治療後の側坐核ドーパミンおよびセロトニンの変動。 神経科学 88, 823–836. doi: 10.1016/s0306-4522(98)00271-1
Song、H.、Fang、F.、Tomasson、G.、Arnberg、FK、Mataix-Cols、D.、Fernandez de la Cruz、L.、etal。 (2018)。 Association of stress-related disorders with subsequent autoimmune disease.ストレス関連障害とその後の自己免疫疾患との関連。 Med。 Assoc.協会 319、2388〜2400。 土井:10.1001 / jama.2018.7028
Soukup、VM、Beiler、ME、およびTerrell、F。(1990)。 Stress, coping style, and problem solving ability among eating-disordered inpatients.摂食障害入院患者のストレス、対処スタイル、問題解決能力。 J.Clin。 サイコロ。 46、592-599。
Sousa-Lima、J.、Moreira、PS、Raposo-Lima、C.、Sousa、N。、およびMorgado、P。(2019)。 Relationship between obsessive compulsive disorder and cortisol: Systematic review and meta-analysis.強迫性障害とコルチゾールの関係:系統的レビューとメタアナリシス。 ユーロ。 Neuropsychopharmacol。 doi:10.1016 / j.euroneuro.2019.09.001 [印刷前のEpub]。
Speakman, JR (2018).スピークマン、JR(XNUMX)。 The evolution of body fatness: trading off disease and predation risk.体脂肪の進化:病気と捕食リスクのトレードオフ。 J. Exp。 バイオ 221(Pt。Suppl。1):jeb167254。 doi: 10.1242/jeb.167254土井:XNUMX / jeb.XNUMX
Speakman、JR、Levitsky、DA、Allison、DB、Bray、MS、de Castro、JM、Clegg、DJなど。 (2011)。 Set points, settling points and some alternative models: theoretical options to understand how genes and environments combine to regulate body adiposity.セットポイント、セトリングポイントおよびいくつかの代替モデル:遺伝子と環境がどのように組み合わされて体脂肪を調節するかを理解するための理論的オプション。 Dis。 Models Mech.モデルメカ。 4、733–745。 doi: 10.1242/dmm.008698土井:XNUMX / dmm.XNUMX
Stanton、C.、Holmes、A.、Chang、S。、およびJoormann、J。(2018)。 From stress to anhedonia: molecular processes through functional circuits.ストレスから無快感症へ:機能回路を介した分子プロセス。 トレンドニューロサイエンス。 42、23-42。
Starr、TB、およびKreipe、RE(2014)。 Anorexia nervosa and bulimia nervosa: brains.神経性食欲不振症および神経性過食症:脳。 Curr。 Psychiatry Rep.精神科担当者 16:11. doi: 10.1007/s11920-014-0441-4
Steiger、H.、Young、SN、Kin、N.、Koerner、N.、Israel、M.、Lageix、P.、etal。 (2001)。 Implications of impulsive and affective symptoms for serotonin function in bulimia nervosa.神経性過食症のセロトニン機能に対する衝動的で感情的な症状の意味。 サイコロ。 メド。 31、85〜95。 doi: 10.1017/s003329179900313x土井:XNUMX / sXNUMXx
Stevens、A.、Purcell、R.、Darling、K.、Eggleston、M.、Kennedy、M。、およびRucklidge、J。(2019)。 Human gut microbiome changes during a 10 week randomised control Trial for micronutrient supplementation in children with attention deficit hyperactivity disorder.注意欠陥多動性障害の子供を対象とした微量栄養素補給のXNUMX週間のランダム化比較試験中のヒト腸内微生物叢の変化。 サイ。 担当者 9:10128. doi: 10.1038/s41598-019-46146-3
Stice、E.、Chase、A.、Stormer、S。、およびAppel、A。(2001)。 A randomized trial of a dissonance-based eating disorder prevention program.不協和に基づく摂食障害予防プログラムのランダム化試験。 Int。 J.Eat。 無理。 29、247〜262。 土井:10.1002 / eat.1016
Stice、E.、Shaw、H.、Burton、E。、およびWade、E。(2006)。 Dissonance and healthy weight eating disorder prevention programs: a randomized efficacy trial.不協和と健康的な体重摂食障害予防プログラム:無作為化有効性試験。 J.相談してください。 クリン サイコロ。 74, 263–275. doi: 10.1037/0022-006x.74.2.263
Strober、M.、Freeman、R.、Lampert、C.、Diamond、J。、およびKaye、W。(2000)。 Controlled family study of anorexia nervosa and bulimia nervosa: evidence of shared liability and transmission of partial syndromes.神経性食欲不振症と神経性過食症の管理された家族研究:責任の共有と部分的症候群の伝播の証拠。 Am。 J.精神医学 157、393〜401。 土井:10.1176 / appi.ajp.157.3.393
Succurro、E.、Segura-Garcia、C.、Ruffo、M.、Caroleo、M.、Rania、M.、Aloi、M.、etal。 (2015)。 Obese patients with a binge eating disorder have an unfavorable metabolic and inflammatory profile.過食症の肥満患者は、代謝および炎症プロファイルが好ましくありません。 医療 94:e2098。 doi: 10.1097/md.0000000000002098土井:XNUMX / md.XNUMX
Sugino, H., Futamura, T., Mitsumoto, Y., Maeda, K., and Marunaka, Y. (2009).杉野秀樹、二村徹、光本恭子、前田健一、丸中恭子(10)。 Atypical antipsychotics suppress production of proinflammatory cytokines and up-regulate interleukin-XNUMX in lipopolysaccharide-treated mice.非定型抗精神病薬は、炎症性サイトカインの産生を抑制し、リポ多糖で治療したマウスのインターロイキン-XNUMXをアップレギュレートします。 プログレ。 神経精神薬。 バイオ 精神医学 33、303〜307。 土井:10.1016 / j.pnpbp.2008.12.006
杉山亮(2015)。 Physical Attractiveness: An Adaptationist Perspective.肉体的な魅力:適応主義者の視点。 Inに、第2版。 Hoboken, NJ: Wiley Online Library., 317–384.ニュージャージー州ホーボーケン:Wiley Online Library。、XNUMX–XNUMX。
Sullivan、PF、Agrawal、A.、Bulik、CM、Andreassen、OA、Borglum、AD、Breen、G.、etal。 (2018)。 Psychiatric genomics: an update and an agenda.精神医学ゲノミクス:最新情報と議題。 Am。 J.精神医学 175、15〜27。 土井:10.1176 / appi.ajp.2017.17030283
Surbey, MK (1987).サーベイ、MK(XNUMX)。 Anorexia nervosa, amenorrhea, and adaptation.神経性食欲不振症、無月経、および適応。 Ethol.エソール。 Sociobiol.ソシオビオール。 8、S47〜S61。
Swami、V.、Frederick、DA、Aavik、T.、Alcalay、L.、Allik、J.、Anderson、D.、etal。 (2010)。 The attractive female body weight and female body dissatisfaction in 26 countries across 10 world regions: results of the international body project I.世界XNUMX地域のXNUMXか国における魅力的な女性の体重と女性の身体の不満:国際的な身体プロジェクトIの結果。 パーソナル。 Soc。 サイコ。 ブル。 36、309〜325。 土井:10.1177 / 0146167209359702
Swanson、SA、Crow、SJ、Le Grange、D.、Swendsen、J。、およびMerikangas、KR(2011)。 Prevalence and correlates of eating disorders in adolescents results from the national comorbidity survey replication adolescent supplement.青年期の摂食障害の有病率と相関関係は、全国的な併存疾患調査の複製青年期サプリメントから得られます。 アーチ。 一般精神医学 68、714〜723。 土井:10.1001 / archgenpsychiatry.2011.22
Tasegian、A.、Curcio、F.、Dalla Ragione、L.、Rossetti、F.、Cataldi、S.、Codini、M.、etal。 (2016)。 Hypovitaminosis D3, leukopenia, and human serotonin transporter polymorphism in anorexia nervosa and bulimia nervosa.神経性食欲不振症および神経性過食症におけるビタミンDXNUMX欠乏症、白血球減少症、およびヒトセロトニントランスポーター多型。 Mediat.仲介。 Inflamm.炎症。 2016:8046479. doi: 10.1155/2016/8046479
Temko、JE、Bouhlal、S.、Farokhnia、M.、Lee、MR、Cryan、JF、and Leggio、L。(2017) The microbiota, the gut and the brain in eating and alcohol use disorders: a 'M,nage A Trois'?摂食およびアルコール使用障害における微生物叢、腸および脳:「M、nage A Trois」? Alcohol.アルコール。 Alcohol.アルコール。 52、403–413。 doi: 10.1093/alcalc/agx024土井:XNUMX / alcalc / agxXNUMX
Thornton、LM、Mazzeo、SE、およびBulik、CM(2011)。 The heritability of eating disorders: methods and current findings.摂食障害の遺伝率:方法と現在の発見。 Neurobiol。 Eat.食べる。 Disord.不和。 6, 141–156. doi: 10.1007/7854_2010_91
Toro、J.、Cervera、M.、Osejo、E。、およびSalamero、M。(1992)。 Obsessive-compulsive disorder in childhood and adolescence: a clinical study.小児期および青年期の強迫性障害:臨床試験。 J.チャイルドサイコ。 精神医学 33, 1025–1037. doi: 10.1111/j.1469-7610.1992.tb00923.x
Tortorella、A.、Fabrazzo、M.、Monteleone、AM、Steardo、L。、およびMonteleone、P。(2014)。 The role of drug therapies in the treatment of anorexia and bulimia nervosa: a review of the literature.拒食症および神経性過食症の治療における薬物療法の役割:文献のレビュー。 J.サイコパトール。 Giornale Psicopatol.ジョルナーレサイコパトール。 20、50-65。
Tovee、MJ、Swami、V.、Furnham、A。、およびMangalparsad、R。(2006)。 Changing perceptions of attractiveness as observers are exposed to a different culture.オブザーバーが異なる文化にさらされるにつれて、魅力に対する認識が変化します。 進化。 ハム。 行動。 27、443–456。 土井:10.1016 / j.evolhumbehav.2006.05.004
Turna、J.、Patterson、B。、およびVan Ameringen、M。(2017)。 An update on the relationship between the gut microbiome and obsessive-compulsive disorder.腸内細菌叢と強迫性障害との関係に関する最新情報。 精神医学アン。 47, 542–551. doi: 10.3928/00485713-20171013-01
Tylee、DS、Sun、JY、Hess、JL、Tahir、MA、Sharma、E.、Malik、R.、etal。 (2018)。 Genetic correlations among psychiatric and immune-related phenotypes based on genome-wide association data.ゲノムワイド関連データに基づく精神医学的表現型と免疫関連表現型の間の遺伝的相関。 J.Med。 Genet.ジェネット。 Part B Neuropsychiatric Genet.パートB神経精神医学的遺伝子。 177、641〜657。 doi: 10.1002/ajmg.b.32652土井:XNUMX / ajmg.b.XNUMX
Vaillancourt、T。(2013)。 Do human females use indirect aggression as an intrasexual competition strategy?人間の女性は、性内競争戦略として間接的な攻撃性を使用していますか? フィロス。 トランス R.Soc。 B Biol。 サイ。 368:20130080。 土井:10.1098 / rstb.2013.0080
Van Ameringen、M.、Mancini、C.、Patterson、B。、およびBennett、M。(2006)。 Topiramate augmentation in treatment-resistant obsessive-compulsive disorder: a retrospective, open-label case series.治療抵抗性の強迫性障害におけるトピラマートの増強:遡及的、非盲検症例シリーズ。 落ち込んでください。 不安 23、1〜5。 土井:10.1002 / da.20118
van Furth、EF、van der Meer、A。、およびCowan、K。(2016)。 Top 10 research priorities for eating disorders.摂食障害の研究の優先事項トップXNUMX。 ランセット精神医学 3、706-707。
(2015)。 Vitamin D status in anorexia nervosa: a meta-analysis.神経性食欲不振症におけるビタミンDの状態:メタアナリシス。 Int。 J.Eat。 無理。 48、803〜813。 土井:10.1002 / eat.22370
Vindas、MA、Johansen、IB、Folkedal、O.、Hoglund、E.、Gorissen、M.、Flik、G.、etal。 (2016)。 Brain serotonergic activation in growth-stunted farmed salmon: adaption versus pathology.成長が阻害された養殖サケにおける脳のセロトニン作動性活性化:適応対病理学 R. Soc。 Sciを開きます。 3:160030。 doi: 10.1098/rsos.160030土井:XNUMX / rsos.XNUMX
Voland、E。、およびVoland、R。(1989)。 Evolutionary biology and psychiatry: the case of anorexia nervosa.進化生物学と精神医学:神経性食欲不振症の場合。 Ethol.エソール。 Sociobiol.ソシオビオール。 10, 223–240. doi: 10.1016/0162-3095(89)90001-0
Wasser、SK、およびBarash、DP(1983)。 Reproductive suppression among female mammals: implications for biomedicine and sexual selection theory.雌の哺乳類の生殖抑制:生物医学および性淘汰理論への影響。 Q. Rev.Biol。 58、513〜538。 土井:10.1086 / 413545
Watson、HJ、Yilmaz、Z.、Thornton、LM、Hübel、C。、Coleman、JR、Gaspar、HA、etal。 (2019)。 Genome-wide association study identifies eight risk loci and implicates metabo-psychiatric origins for anorexia nervosa.ゲノムワイド関連研究は、XNUMXつのリスク遺伝子座を特定し、神経性食欲不振症のメタボ精神医学的起源を示唆しています。 Nat。 Genet 51, 1207–1214. doi: 10.1038/s41588-019-0439-2
Watson、R。、およびVaugn、L。(2006)。 Limiting the effects of the media on body image: does the length of the intervention make a difference?身体イメージに対するメディアの影響を制限する:介入の長さは違いを生むか? 食べる。 無理。 14、384-400。
Wedell-Neergaard、AS、Lehrskov、LL、Christensen、RH、Legaard、GE、Dorph、E.、Larsen、MK、etal。 (2019)。 Exercise-induced changes in visceral adipose tissue mass are regulated by IL-6 signaling: a randomized controlled trial.内臓脂肪組織量の運動誘発性変化は、IL-XNUMXシグナル伝達によって調節されています:ランダム化比較試験。 セルメタ 29、844〜855。 doi: 10.1016/j.cmet.2018.12.007土井:XNUMX / j.cmet.XNUMX
Wells, JCK (2006).ウェルズ、JCK(XNUMX)。 The evolution of human fatness and susceptibility to obesity: an ethological approach.人間の肥満と肥満への感受性の進化:動物行動学的アプローチ。 バイオル。 牧師 81、183〜205。 土井:10.1017 / s1464793105006974
Wilksch、SM、およびWade、TD(2009)。 Reduction of shape and weight concern in young adolescents: a 30-month controlled evaluation of a media literacy Program.若い青年における形と体重の懸念の軽減:メディアリテラシープログラムのXNUMXか月の管理された評価。 J. Am。 Acad。 子アデレスク。 精神医学 48, 652–661. doi: 10.1097/CHI.0b013e3181a1f559
Williams, ACDC (2019).ウィリアムズ、ACDC(XNUMX)。 Persistence of pain in humans and other mammals.人間や他の哺乳類の痛みの持続。 フィロス。 トランス R.Soc。 B 374:20190276。 土井:10.1098 / rstb.2019.0276
Wotton、CJ、James、A。、およびGoldacre、MJ(2016)。 Coexistence of eating disorders and autoimmune diseases: record linkage cohort study.摂食障害と自己免疫疾患の共存:記録リンケージコホート研究。 Int。 J.Eat。 無理。 49、663〜672。 土井:10.1002 / eat.22544
Wu、XJ、Hung、HC、Kastin、AJ、He、Y.、Khan、RS、Stone、KP、etal。 (2011)。 Interleukin-15 affects serotonin system and exerts antidepressive effects through IL15R alpha receptor.インターロイキン-XNUMXはセロトニンシステムに影響を与え、ILXNUMXRアルファ受容体を介して抗うつ効果を発揮します。 精神神経内分泌学 36、266〜278。 土井:10.1016 / j.psyneuen.2010.07.017
Yager、Z。、およびO'Dea、JA(2008)。 Prevention programs for body image and eating disorders on University campuses: a review of large, controlled interventions.大学のキャンパスにおけるボディイメージと摂食障害の予防プログラム:大規模で管理された介入のレビュー。 Int。 23、173〜189。 doi: 10.1093/heapro/dan004土井:XNUMX / heapro / danXNUMX
Zerwas、S.、Larsen、JT、Petersen、L.、Thornton、LM、Quaranta、M.、Koch、SV、etal。 (2017)。 Eating disorders, autoimmune, and autoinflammatory disease.摂食障害、自己免疫、および自己炎症性疾患。 小児科 140:e20162089. doi: 10.1542/peds.2016-2089
Zhang、Y.、Leung、DYM、Richers、BN、Liu、YS、Remigio、LK、and Riches、DW(2012) Vitamin D inhibits monocyte/macrophage proinflammatory cytokine production by targeting MAPK phosphatase-1.ビタミンDは、MAPKホスファターゼ-XNUMXを標的とすることにより、単球/マクロファージの炎症性サイトカインの産生を阻害します。 J.Immunol。 188、2127〜2135。 doi: 10.4049/jimmunol.1102412土井:XNUMX / jimmunol.XNUMX
キーワード:神経性食欲不振症、過食症、神経性過食症、進化精神医学、神経炎症、ストレス応答性、ミスマッチ仮説、適応メタ問題
引用:Rantala MJ、Luoto S、Krama T、Krams I(2019)摂食障害:進化的精神神経免疫学的アプローチ。 前面。 サイコロル。 10:2200 土井:10.3389 / fpsyg.2019.02200
受信:05 3月2019; 受け入れ:12 9月2019;
公開:29 10月2019。
によって編集:
ヤン・アントフォーク、オーボアカデミ大学、フィンランド
したレビュー:
モニカアルガース、オーボアカデミ大学、フィンランド
ジェフリー・ベッドウェル、セントラルフロリダ大学、アメリカ合衆国
Copyright©2019Rantala、Luoto、Krama andKrams。 This is an open-access article distributed under the terms of theこれは、の条件の下で配布されるオープンアクセスの記事です。 クリエイティブコモンズ表示ライセンス(CC BY)。 原著者と著作権者が信用され、この雑誌の原著論文が認められた学術慣習に従って引用されていることを条件として、他のフォーラムでの使用、配布または複製は許可されています。 これらの条件に従わない使用、配布、複製は許可されていません。
*対応:Markus J. Rantala、 [メール保護]