Opmerkingen: deze beoordeling is geproduceerd door het hoofd van NIDA, Nora Volkow en haar team. Het laat weinig twijfel bestaan dat chemische verslavingen en gedragsverslavingen dezelfde of vergelijkbare mechanismen en neurale circuits delen. Dit is volkomen logisch, omdat chemische verslavingen het neurale circuit kapen voor binding, seks en eten. Omdat seks twee keer zoveel dopamine afgeeft als het eten van je favoriete eten, en een pornogebruiker kan urenlang dopamine hoog houden, is het gek om te stellen dat pornoverslaving niet kan bestaan.
Curr Top Behav Neurosci. 2011 Oct 21.
Volkow ND, Wang GJ, Fowler JS, Tomasi D, Baler R.
Bron
Nationaal Instituut voor Drugsmisbruik, 6001 Executive Boulevard 6001, Room 5274, Bethesda, MD, 20892, VS, [e-mail beveiligd].
Abstract
Zowel drugsverslaving als obesitas kunnen worden gedefinieerd als stoornissen waarbij de saliency-waarde van een type beloning (respectievelijk drugs en voedsel) abnormaal wordt verhoogd ten opzichte van en ten koste van anderen. Dit model is consistent met het feit dat zowel medicijnen als voedsel krachtige versterkende effecten hebben - gedeeltelijk gemedieerd door dopamine-verhogingen in het limbische systeem - die, onder bepaalde omstandigheden of bij kwetsbare individuen, de homeostatische controlemechanismen van de hersenen zouden kunnen overweldigen. Dergelijke parallellen hebben geleid tot significante interesse in het begrijpen van de gedeelde kwetsbaarheden en trajecten tussen verslaving en obesitas. Nu zijn hersenscan-ontdekkingen begonnen gemeenschappelijke kenmerken tussen deze twee condities te ontdekken en enkele overlappende hersencircuits te schetsen waarvan disfuncties stereotypische en gerelateerde gedragstekorten in menselijke subjecten kunnen verklaren. TDeze resultaten suggereren dat zowel obese als drugsverslaafden lijden aan stoornissen in dopaminerge routes die neuronale systemen reguleren die niet alleen verband houden met beloningsgevoeligheid en stimulerende motivatie, maar ook met conditionering (geheugen / leren), impulscontrole (gedragsinhibitie), stressreactiviteit en interoceptief bewustzijn. Hier integreren we bevindingen die voornamelijk zijn afgeleid van positronemissietomografie die licht werpen op de rol van dopamine bij drugsverslaving en bij obesitas, en stellen een bijgewerkt werkmodel voor om behandelingsstrategieën te identificeren die beide aandoeningen ten goede kunnen komen.
1 Achtergrond
2 De rol van dopamine bij acute beloning voor geneesmiddelen en voedsel
3 Beeldvorming van DA in reactie op geneesmiddelen en geconditioneerde aanwijzingen bij verslaving
4 Het effect van disfunctie in remmende controle
5 Betrokkenheid van motivatiecircuits
6 Betrokkenheid van interoceptieve schakelingen
7 Het schakelsysteem van afkeer
8 Pathologische geneesmiddelen- en voedselbeloning: een aangepast werkmodel
1 Achtergrond
Dopamine (DA) wordt beschouwd als een sleutel tot de lonende effecten van beloningen voor natuurlijke en geneesmiddelen. Zijn rol in het verlies van controle en compulsief gedrag dat geassocieerd wordt met verslaving en obesitas is echter veel minder duidelijk. PET-onderzoeken hebben een cruciale rol gespeeld bij het karakteriseren van de rol van de hersenen van DA-systemen bij verslaving (naast de rol ervan bij geneesmiddelenbeloningen) en bij obesitas. Drugsmisbruik (inclusief alcohol) wordt inderdaad door mensen geconsumeerd of zelf toegediend door proefdieren, omdat ze inherent lonend zijn, een effect dat wordt gemedieerd door hun DA-verhogende eigenschappen in het mesolimbische systeem (Wise 2009). HIn het geval van verslaving hebben beeldvormende onderzoeken echter onthuld dat de stoornis niet alleen de DA-beloningscircuit beïnvloedt, maar ook andere DA-routes die betrokken zijn bij de modulatie van conditionering / gewoonten, motivatie en executieve functies (remmende controle, salience-attributie en beslissing -vorming), en dat DA-deficiënties ook kunnen bijdragen aan de verhoogde stressreactiviteit en verstoring van het interoceptieve bewustzijn in verband met verslaving. Preklinische en klinische studies hebben ook andere neurotransmitters (en neuropeptiden) onthuld die een belangrijke rol spelen bij de beloning en verslaving van geneesmiddelen. (dat wil zeggen cannabinoïden, opioïden) en zijn nauw betrokken bij de neuroplastische veranderingen die zich voordoen na herhaald drugsgebruik (dwz glutamaat, opioïden, GABA, corticotropine-afgevende factor). Het glutamaterge systeem is in dit opzicht in het bijzonder prominent omdat het de verstoringen in zowel langdurige versterking als langdurige depressie medieert die zijn waargenomen in diermodellen van chronisch geneesmiddeltoediening (Thomas et al. 2008). Recensies met betrekking tot deze aanvullende systemen zijn elders te vinden (Kalivas 2009; Koob 1992).
Omdat medicijnen dezelfde beloningssystemen activeren die ten grondslag liggen aan voedselbeloningen, is het niet geheel onverwacht dat, in het algemeen, hersenafbeeldingsstudies het idee hebben ondersteund dat stoornissen in DA-gemoduleerde circuits ook betrokken zijn bij pathologisch, dwangmatig eetgedrag. Voedingscues, zoals drug cues, verhogen striatale extracellulaire DA en stimuleren de motivatie om deel te nemen aan het gedrag dat nodig is om het voedsel te verkrijgen en te eten, bewijsmateriaal voor de betrokkenheid van DA niet alleen in voedselbeloning maar ook in de niet-hedonische motivatie eigenschappen van voedsel (dat wil zeggen calorie-eisen) en de afname van remmende controle waargenomen bij dwangmatige overeten (Avena et al. 2008; Volkow et al. 2008).
Hier bespreken we bevindingen van beeldvormingsstudies die specifiek gericht zijn op de overlappingen in de hersencircuits die verstoord zijn door obesitas en in drugsverslaving. Het is echter de moeite waard om te onthouden dat de regulering van het gedrag van voedselinnames veel complexer is dan de regulatie van de inname van geneesmiddelen. De laatste wordt voornamelijk gemedieerd door de belonende effecten van geneesmiddelen, terwijl de eerste wordt gemoduleerd, niet alleen door de belonende effecten (hedonische factoren), maar ook door meerdere perifere en centrale factoren die nutriëntvereisten in het lichaam aantonen die nodig zijn voor overleving (homeostatische factoren). Interessant is dat er steeds meer aanwijzingen zijn dat homeostatische factoren (bijv. Insuline, leptine, ghreline) de voedselinname gedeeltelijk moduleren door de gevoeligheid van hersenbeloningscircuits voor voedselprikkels te vergroten of te verkleinen (Volkow et al. 2011).
2 De rol van dopamine bij acute beloning voor geneesmiddelen en voedsel
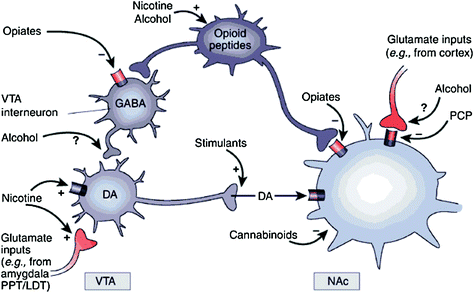
Of ze nu direct of indirect zijn, alle verslavende drugs hebben een vermogen om DA in nucleus accumbens (NAc) te verhogen via specifieke interacties met verschillende moleculaire doelwitten (Nestler 2004) (Fig. 1). De mesolimbische DA-route [DA-cellen in het ventrale tegmentale gebied (VTA) die naar het NAc] projecteren, lijkt cruciaal te zijn voor de beloning van geneesmiddelen (Wise 2009). Echter, zoals hieronder beschreven, dragen ook andere DA-routes [mesostriatale (DA-cellen in substantia nigra die naar dorsale striatum projecteren) en mesocorticale (DA-cellen in VTA die naar de frontale cortex projecteren)]] bij tot geneesmiddelbeloning en -verslaving (Wise 2009). Over het algemeen lijkt het erop dat de belonende en conditionerende effecten van geneesmiddelen voornamelijk worden veroorzaakt door fasisch DA-celsynchronisatie, wat leidt tot grote en voorbijgaande DA-verhogingen. Daarentegen zijn de stroomafwaartse veranderingen in de uitvoerende functie die optreden bij verslaving, gekoppeld aan veranderingen in het vuren van tonische DA-cellen en resulteren in lagere maar stabielere DA-niveaus (Grace 2000; Wanat et al. 2009). Dit wijst op zijn beurt weer naar de D1-receptoren (D1R), die DA-receptoren zijn met lage affiniteit die cyclische AMP-signalering stimuleren, omdat ze zowel betrokken zijn bij acute geneesmiddelenbeloning als bij conditionering, omdat deze worden geassocieerd met de hoge DA-concentraties die noodzakelijk zijn. om D1R te stimuleren. D2R's daarentegen, die cyclische AMP-signalering remmen, worden gestimuleerd door zowel fasische als tonische DA. Merk op dat, vanwege het ontbreken van specifieke radiotracers voor de PET-beeldvorming van DA-receptoren van de D1-, D3-, D4- en D5-typen, de meeste onderzoeken naar de effecten van drugs tegen misbruik en verslaving in het menselijk brein zich hebben gericht op D2R's.
Fig 1 Drugs van misbruik handelen op de beloning en bijkomende circuits door verschillende mechanismen, maar ze leiden allemaal tot vergelijkbare dopaminerge effecten in de VTA en NAc. Aldus stimuleren stimulantia acumbal DA direct, terwijl opiaten dit doen door de remmende toon van GABAergic interneuronen op DA-signalering te verlagen, zowel in de VTA of in daarna NAc. Hoewel de mechanismen van andere drugsmisbruik minder duidelijk zijn, zijn er aanwijzingen dat nicotine VTA DA rechtstreeks kan activeren via nicotine-acetylcholinereceptor (nAChR) op die neuronen en indirect via stimulatie van de receptoren ervan op glutamaterge zenuwuiteinden die de DA-cellen innerveren. Alcohol lijkt GABAergic-terminals in VTA te remmen, wat leidt tot ontremming van DA-neuronen in de VTA. Cannabinoïden werken onder andere door de activering van CB1-receptoren op glutamaterge en GABAergic zenuwuiteinden in het NAc, en op NAc-neuronen zelf. Fencyclidine (PCP) kan werken door postsynaptische NMDA-glutamaatreceptoren in het NAc te remmen. Bovendien is er enig bewijs dat suggereert dat nicotine en alcohol ook een interactie kunnen aangaan met endogene opioïde en cannabinoïde routes (niet getoond). PPT / LDT, peduncular pontine tegmentum / lateral dorsal tegmentum. Overgenomen met toestemming Nestler (2005)
Bij mensen hebben PET-onderzoeken aangetoond dat verschillende geneesmiddelen [stimulerende middelen (Drevets et al. 2001; Volkow et al. 1999b), nicotine (Brody et al. 2009), alcohol (Boileau et al. 2003) en marihuana (Bossong et al. 2009)] verhoog DA in dorsaal en ventraal striatum (waar het NAc zich bevindt). Deze studies maken gebruik van verschillende radiotracers, zoals [11C] raclopride, die binden aan D2R, maar alleen wanneer deze geen bindende endogene DA (niet-bezet) zijn, die onder basiscondities overeenkomt met 85-90% van de striatale D2R (Abi-Dargham et al. 1998). Dus een vergelijking van [11C] raclopride-binding na placebo en na toediening van het geneesmiddel kan ons helpen de dalingen in D2R-beschikbaarheid die door het medicijn wordt veroorzaakt (of andere stimuli die DA kunnen verhogen) te schatten. Deze dalingen in [11C] binding van raclopride is evenredig met de DA-verhogingen (Breier et al. 1997). Deze studies hebben aangetoond dat door geneesmiddelen veroorzaakte DA-verhogingen in striatum evenredig zijn met de intensiteit van de subjectieve ervaring van euforie of "high" [zie recensie (Volkow et al. 2009)] (Fig. 2).
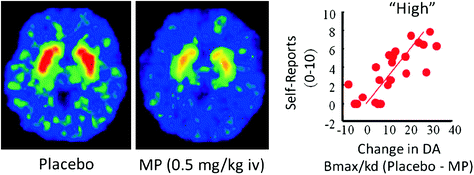
Fig 2 Effecten van intraveneus methylfenidaat (MP) op de binding van raclopride en de relatie tussen striatale DA-verhogingen veroorzaakt door MP in het striatum en de zelfrapportages van "hoog". Gewijzigd van Volkow et al. (1999b)
PET-onderzoeken hebben ook een duidelijke, directe relatie aangetoond tussen het farmacokinetische profiel van een medicijn (dwz de snelheid waarmee het de hersenen binnengaat en verlaat) en de versterkende effecten ervan. In het bijzonder geldt dat hoe sneller een geneesmiddel piekniveaus in de hersenen bereikt, des te intenser de "high" (Volkow et al. 2009). Bijvoorbeeld, voor een gelijkwaardig niveau van cocaïne dat de hersenen bereikte (beoordeeld door middel van PET), toen cocaïne snel in de hersenen terechtkwam (gerookt of iv toediening), lokte het een meer intense "high" uit dan wanneer het in een langzamer tempo binnenkwam (snuif) (Volkow et al. 2000). Dit komt overeen met preklinische studies die een vergelijkbare correlatie laten zien tussen het farmacokinetisch profiel van een medicijn en zijn versterkende eigenschappen (Balster en Schuster 1973). IkHet is redelijk om te veronderstellen dat een dergelijke abrupte en grote DA-toename, veroorzaakt door misbruiktegen drugs, de snelle en grote DA-verhogingen kan nabootsen die het gevolg zijn van fasisch DA-schieten dat in de hersenen is geassocieerd met de verwerking van informatie over beloning en oplettendheid (Schultz 2010). Dergelijke door geneesmiddelen veroorzaakte DA-verhogingen in het NAc kunnen noodzakelijk zijn voor verslaving, maar het feit dat ze ook voorkomen bij niet-verslaafde personen geeft aan dat ze onvoldoende zijn om het impulsieve en compulsieve drugsgebruik dat kenmerkend is voor verslaving te verklaren.
Er zijn nu aanwijzingen dat vergelijkbare dopaminerge reacties verband houden met de voedselbeloning en dat deze mechanismen waarschijnlijk ook een rol spelen bij overmatige voedselconsumptie en obesitas. Het is bekend dat bepaalde voedingsmiddelen, met name die rijk aan suikers en vet, zeer de moeite waard zijn (Lenoir et al. 2007). Hoogcalorisch voedsel kan overeten (eten dat is losgekoppeld van energetische behoeften) bevorderen en geleerde associaties tussen de stimulus en de beloning (conditionering) teweegbrengen. In evolutionaire termen was deze eigenschap van smakelijke voedingsmiddelen voordelig in omgevingen waar voedselbronnen schaars en / of onbetrouwbaar waren, omdat het ervoor zorgde dat voedsel werd gegeten indien beschikbaar, waardoor energie in het lichaam (als vet) kon worden opgeslagen voor toekomstig gebruik . Helaas, in samenlevingen zoals de onze, waar voedsel overvloedig is en constant beschikbaar is, is deze aanpassing een verplichting geworden.
Verschillende neurotransmitters, waaronder DA, cannabinoïden, opioïden en serotonine, evenals hormonen en neuropeptiden die betrokken zijn bij homeostatische regulatie van voedselinname, zoals insuline, orexin, leptine en ghreline, zijn betrokken bij de belonende effecten van voedsel (Atkinson 2008; Cason et al. 2010; Cota et al. 2006). Hiervan is DA het grondigst onderzocht en het best gekarakteriseerd. Experimenten met knaagdieren hebben aangetoond dat, na de eerste blootstelling aan een voedselbeloning, het vuren van DA-neuronen in de VTA toeneemt met een resulterende toename in DA-afgifte in NAc (Norgren et al. 2006). Evenzo is bij gezonde menselijke proefpersonen met normaal gewicht aangetoond dat de inname van smakelijk voedsel DA in het dorsale striatum afgeeft in verhouding tot de beoordelingen van maaltijdplezier (Small et al. 2003) (Fig. 3). Echter, zoals blijkt uit onderzoeken met drugsgebruikers, kan door voedsel veroorzaakte toename van alleen striatum DA het verschil tussen normale voedselinname en overmatige dwangmatige voedselconsumptie niet verklaren, aangezien deze ook voorkomen bij gezonde personen die niet overmatig eten. Dus, zoals het geval is voor verslaving, zijn downstreamaanpassingen waarschijnlijk betrokken bij het verlies van controle over voedselinname.
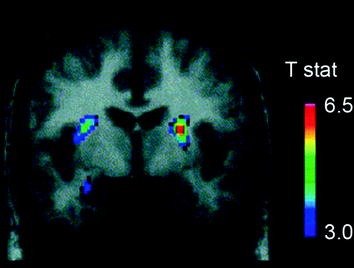
Fig 3 Dopamine-afgifte geïnduceerd door voeding. Coronale sectie van de T-map met statistisch significante verminderingen in [11C] het bindende potentieel van raclopride (BP) na het voeren. De kleurenbalk geeft de t-statistische waarden weer. (Overgenomen met toestemming Small et al. 2003)
3 Beeldvorming van DA in reactie op geneesmiddelen en geconditioneerde aanwijzingen bij verslaving
DA's rol in versterking is complexer dan alleen coderen voor beloning per se (hedonisch plezier); stimulansen die snelle en grote DA-verhogingen induceren, stimuleren bijvoorbeeld ook geconditioneerde responsen en lokken stimuleringsmotivatie uit om deze te verkrijgen (Owesson-White et al. 2009). Dit is belangrijk omdat, door het proces van conditionering, neutrale stimuli die zijn gekoppeld aan de bekrachtiger (of het nu een natuurlijke of een medicijnversterker is) het vermogen verwerven om zelf DA te verhogen in striatum (inclusief NAc) in afwachting van de beloning, waardoor een sterke motivatie om het medicijn te zoeken (Owesson-White et al. 2009). Het loskoppelen van belonings- en conditioneringsmechanismen in het proces van drugsverslaving is echter uitdagender dan voor voedselconsumptie, omdat drugs door hun farmacologische effecten direct DA-neuronen activeren (dwz nicotine) of DA-afgifte (dat wil zeggen amfetamine) verhogen.
Hersenafbeeldingsstudies waarin de DA-verhogingen werden vergeleken die waren geïnduceerd door het stimulerende medicijn methylfenidaat (MP) of amfetamine (AMPH) bij cocaïne-verslaafde proefpersonen versus controles vertoonden een duidelijke verzwakking van MP of AMPH-geïnduceerde DA-verhogingen in striatum (50% lager in gedetoxificeerde misbruikers en 80% in actieve misbruikers) en lagere zelfrapportages van de belonende effecten van het medicijn ten opzichte van niet-medicamenteuze controles (Martinez et al. 2007; Volkow et al. 1997) (Fig. 4). Dit was verrassend, omdat MP en AMPH farmacologisch vergelijkbaar zijn met respectievelijk cocaïne en metamfetamine, en drugsverslaafden geen onderscheid kunnen maken tussen hen wanneer ze intraveneus worden toegediend. Omdat de duidelijke afnames in de door drugs geïnduceerde DA-verhogingen werden waargenomen ongeacht of de cocaïnegebruikers wel of niet waren ontgift, geeft dit aan dat de staat van terugtrekking geen verstorende factor is (Volkow et al. 2011b). Deze en gerelateerde resultaten (Volkow et al. 2009) zijn consistent met de hypothese dat de hedonische respons deficiënt wordt in drugsverslaafden, en de gedachte versterken dat de acute farmacologische DA-verhogende effecten van het geneesmiddel in NAc de verhoogde motivatie om ze te consumeren niet zelf kunnen verklaren.
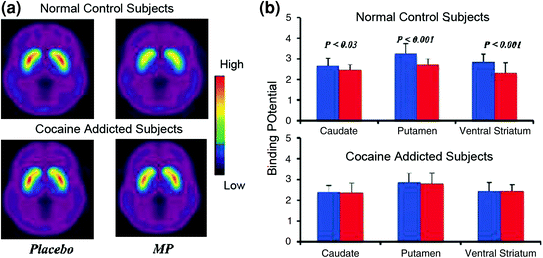
Fig 4 DA-veranderingen geïnduceerd door iv MP in controles en in actieve cocaïne-verslaafde onderwerpen. een gemiddelde niet-plaatsbare biding potential (BPND) -afbeeldingen van [11C] raclopride in actieve cocaïneverslaafden (n = 19) en in controles (n = 24) getest na placebo en na iv MP. b D2R beschikbaarheid (BPND) in caudate, putamen en ventraal striatum na placebo (blauw) en na MP (rood) in controles en in cocaïne-verslaafde onderwerpen. MP verminderde D2R in controles, maar niet in onderwerpen die aan cocaïne waren verslaafd. Merk op dat cocaïnegebruikers zowel afname van baseline striatale beschikbaarheid van D2R (placebo-meting) als afname in DA-afgifte laten zien bij gebruik van iv MP (gemeten als afname van D2R-beschikbaarheid vanaf baseline). Hoewel men de vraag kan stellen in hoeverre de lage striatale beschikbaarheid van D2R in een aan cocaïne verslavend onderwerp het vermogen beperkt om verdere verlagingen van MP te detecteren, wijst het feit dat cocaïneverslaafde personen verlaging van de beschikbaarheid van D2R bij blootstelling aan cocaïne aanwijzingen geven aan dat de verzwakte effecten van MP op [11C] raclopride-binding weerspiegelt verminderde DA-afgifte. Overgenomen met toestemming (Volkow et al. 1997; Wang et al. 2010)
De respons van VTA DA-neuronen op stimulerende stimuli verandert bij herhaalde blootstelling.
Terwijl DA-cellen vuren bij de eerste blootstelling aan een nieuwe beloning, zorgt herhaalde blootstelling aan DA ervoor dat de neuronen stoppen met schieten op beloningsconsumptie en vuur in plaats daarvan wanneer ze worden blootgesteld aan stimuli die de beloning voorspellen (Schultz et al. 1997). Dit zal waarschijnlijk ten grondslag liggen aan de rol van DA in leren en conditionering. Inderdaad, door geneesmiddelen geïnduceerde fasische DA-signalering kan uiteindelijk neuroadaptaties in nevencircuits teweegbrengen die verband houden met gewoontevorming en gedragsconditionering. Tdeze veranderingen worden voornamelijk veroorzaakt door D1R-signalering en synaptische veranderingen in glutamaatgemoduleerde NMDA- en AMPA-receptoren (Luscher en Malenka 2011; Zweifel et al. 2009). Rekrutering van deze circuits is significant in de ziekteprogressie omdat de daaropvolgende geconditioneerde responsen helpen bij het verklaren van de intense behoefte aan het medicijn (verlangen) en het dwangmatige gebruik dat optreedt wanneer verslaafde personen worden blootgesteld aan drugsgerelateerde aanwijzingen. Deze hypothese is consistent met onafhankelijke waarnemingen (Volkow et al. 2006b; Wong et al. 2006) die de kracht van cocaïne-geassocieerde cue-blootstelling laten zien om de DA-niveaus in het dorsale striatum te verhogen en een gelijktijdige toename van de subjectieve ervaring van craving in gedetoxificeerde cocaïne-misbruikers teweeg te brengen (Fig. 5). Omdat het dorsale striatum een rol speelt bij het leren van gewoonten (Belin et al. 2009; Yin et al. 2004), weerspiegelt de associatie waarschijnlijk de versterking van gewoonten naarmate de chroniciteit van de verslaving vordert. Dit suggereert dat een fundamentele verstoring van de verslaving gerelateerd kan zijn aan de DA-getriggerde geconditioneerde responsen die resulteren in gewoonten die leiden tot intense hunkering en dwangmatig drugsgebruik. Interessant is dat bij actief gebruik van cocaïne-verslaafde onderwerpen, de DA-verhogingen veroorzaakt door geconditioneerde aanwijzingen zelfs groter blijken te zijn dan die geproduceerd door het stimulerende medicijn zelf zoals beoordeeld in twee afzonderlijke groepen van individuen (Volkow et al. 2011b, 2006b), wat suggereert dat geconditioneerde reacties de DA-signalering kunnen aanwakkeren die de motivatie om het medicijn te nemen behoudt, zelfs wanneer de farmacologische effecten ervan lijken te worden verminderd. Dus hoewel medicijnen in eerste instantie gevoelens van directe beloning kunnen opwekken door DA-afgifte in het ventrale striatum, bij herhaald gebruik, en als gewoonte zich ontwikkelt, lijkt er een verschuiving van het medicijn naar de geconditioneerde stimulus te zijn. Volgens studies in proefdieren bemiddelen glutamaterge projecties van de prefrontale cortex en van amygdala in VTA / SN en NAc deze geconditioneerde reacties (Kalivas 2009). Op deze manier kan de loutere voorspelling van een beloning uiteindelijk de beloning worden die het gedrag motiveert dat nodig is voor drugs- (of voedsel) consumptie.
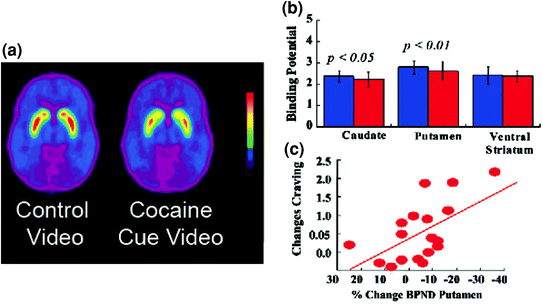
Fig 5 DA-veranderingen geïnduceerd door geconditioneerde aanwijzingen in actieve cocaïne-verslaafde onderwerpen. een gemiddelde niet-plaatsbare bindingspotentiaal (BPND) -afbeeldingen van [11C] -raclopride in aan cocaïne verslaafde personen (n = 17), getest tijdens het bekijken van een neutrale video (natuurtaferelen) en tijdens het bekijken van een video met cocaïne-aanwijzingen (proefpersonen die cocaïne toedienen). b D2R beschikbaarheid (BPND) in caudate, putamen en ventrale striatum voor de neutrale video (blauw) en de cocaïne-cues video (rood). De cocaïne-aanwijzingen verminderden D2R in caudate en putamen. c Correlaties tussen veranderingen in D2R (als gevolg van DA neemt toe) en zelfrapportages van cocaïnecraving geïnduceerd door de video met cocaïne-aanwijzingen. Gewijzigd van ref. (Volkow et al. 2006b)
IIn feite is dit type functionele "switch" ook gerapporteerd voor natuurlijke versterkers, die waarschijnlijk een gelijkwaardige en geleidelijke verschuiving in DA-verhogingen veroorzaken, van ventrale naar meer dorsale gebieden van het striatum tijdens de overgang van een nieuwe stimulus die inherent is belonen met dat van de bijbehorende signalen die het voorspellen. Deze overgang wordt overgebracht door DA-signalering, die lijkt te coderen voor een "beloningsvoorspellingsfout" (Schultz 2010). De uitgebreide glutamaterge afferenten voor DA-neuronen uit regio's die betrokken zijn bij de verwerking van sensorische (insula of primaire gustatory cortex), homeostatische (hypothalamus), beloning (NAc), emotionele (amygdala en hippocampus) en multimodale (orbitofrontale cortex voor opvallende toeschrijving) informatie , moduleren hun activiteit in reactie op beloningen en geconditioneerde aanwijzingen (Geisler en Wise 2008). Meer specifiek zijn projecties van de amygdala en de orbitofrontale cortex (OFC) naar DA-neuronen en naar NAc betrokken bij geconditioneerde reacties op voedsel (Petrovich). 2010). Inderdaad toonden beeldvormende onderzoeken aan dat wanneer niet-zwaarlijvige mannelijke proefpersonen gevraagd werden hun eetlust te verminderen - terwijl ze werden blootgesteld aan voedingsstoffen - ze een verminderde metabole activiteit vertoonden in amygdala en OFC (evenals in de hippocampus), insula en striatum, en dat de afnames in OFC geassocieerd waren met verminderingen van het verlangen naar voedsel (Wang et al. 2009). Een vergelijkbare remming van de metabole activiteit in de OFC (en ook in NAc) is waargenomen bij cocaïne-misbruikers toen hen werd gevraagd hun medicijn hunkeren naar blootstelling aan cocaïne-aanwijzingen (Volkow et al. 2009b).
Toch zou de opkomst van dergelijke krachtige cue-geconditioneerde onbedwingbare trek, die voor voedsel ook voorkomt bij gezonde individuen die niet te veel eten, niet zo verwoestend zijn als ze niet gepaard gaan met groeiende tekorten in het vermogen van de hersenen om slecht aanpasbaar gedrag te remmen.
4 Het effect van disfunctie in remmende controle
Het vermogen om prepotente reacties te remmen, draagt zeker bij aan het vermogen van een individu om te vermijden dat hij zich met ongepast gedrag bezighoudt, zoals het nemen van medicijnen of eten voorbij het punt van verzadiging, en daarmee zijn / haar kwetsbaarheid voor verslaving (of obesitas) te vergroten (Volkow en Fowler) 2000; Volkow et al. 2008).
PET-onderzoeken hebben een significante vermindering van de beschikbaarheid van D2R in het striatum van verslaafde personen aan het licht gebracht die nog maanden na langdurige detoxificatie aanhouden [gereviewed in (Volkow et al. (1998). 2009)]. Evenzo hebben preklinische studies bij knaagdier- en niet-menselijke primaten aangetoond dat herhaalde blootstelling van geneesmiddelen verband houdt met verlagingen van striatale D2R-niveaus (Nader et al. 2006; Thanos et al. 2007; Volkow et al. 2001). In het striatum bemiddelen D2R's signalering in de striatale indirecte route die prefrontale regio's moduleert; en er is aangetoond dat de downregulatie de sensitisatie verbetert voor de effecten van geneesmiddelen in diermodellen (Ferguson et al. 2011). Bij mensen die verslaafd zijn aan geneesmiddelen, is de reductie van striatale D2R geassocieerd met verminderde activiteit van prefrontale regio's, zoals wordt aangetoond door afnames in het basislijn-glucosemetabolisme (een marker van de hersenfunctie) in OFC, anterieure cingulate gyrus (ACC) en dorsolaterale prefrontale cortex (DLPFC) ) (Volkow et al. 2001, 1993, 2007) (Fig. 6). Voor zover OFC, ACC en DLPFC betrokken zijn bij salience-attributie, remmende controle / emotieregulatie en besluitvorming, respectievelijk, er is gepostuleerd dat hun onjuiste regulatie door D2R-gemedieerde DA-signalering in verslaafde onderwerpen ten grondslag zou kunnen liggen aan de verhoogde motivationele waarde van drugs in hun gedrag en het verlies van controle over de inname van geneesmiddelen (Volkow en Fowler 2000). Bovendien, omdat stoornissen in OFC en ACC geassocieerd zijn met dwangmatig gedrag en impulsiviteit (Fineberg et al. 2009), DA's verstoorde modulatie van deze regio's zal waarschijnlijk bijdragen aan de dwangmatige en impulsieve drugsinname die wordt gezien bij verslaving (Goldstein en Volkow 2002). Inderdaad, bij methamfetamine-misbruikers, werd lage striatale D2R geassocieerd met impulsiviteit (Lee et al. 2009), en het voorspelde ook de toediening van compulsieve cocaïne bij knaagdieren (Everitt et al. 2008). Een omgekeerd scenario, waarbij een eerste kwetsbaarheid voor drugsgebruik reeds bestaat in prefrontale regio's, en waarbij herhaalde medicijngebruiken verdere verlagingen van striatale D2R veroorzaken, is ook mogelijk. Inderdaad, een studie uitgevoerd bij proefpersonen die ondanks een hoog risico op alcoholisme (positieve familiegeschiedenis van alcoholisme) geen alcoholisten waren, onthulden een hogere dan normale striatale beschikbaarheid van D2R die werd geassocieerd met een normaal metabolisme in OFC, ACC en DLPFC (Volkow et al. 2006). Dit suggereert dat bij deze proefpersonen met een risico op alcoholisme de normale prefrontale functie gekoppeld was aan verbeterde striatale D2R-signalering, wat hen op hun beurt mogelijk heeft beschermd tegen alcoholmisbruik.
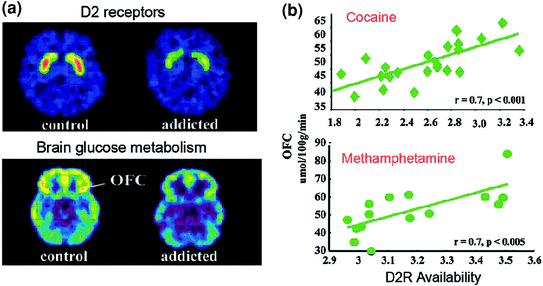
Fig 6 Correlaties tussen striatale beschikbaarheid en metabolisme van D2R in prefrontale hersengebieden. a Axiale hersenbeelden voor een controle en voor een cocaïneverslaafd onderwerp voor basislijnafbeeldingen van D2R beschikbaarheid in striatum (verkregen met [11C] raclopride) en van glucose metabolisme in de hersenen in OFC (verkregen met [18FDG). b Correlaties tussen striatum D2R en metabolisme in OFC bij verslaafden die verslaafd zijn aan cocaïne en methamfetamine. Overgenomen van Volkow et al. (2009) Copyright (2009), met toestemming van Elsevier
Voorspelbaar is dat er aanwijzingen zijn voor ontregeling in regelcircuits bij obese personen. Zowel preklinische als klinische studies hebben bewijs geleverd van verminderde striatale D2R-signalering, die, zoals hierboven vermeld, verband houdt met beloning (NAc) maar ook met het vaststellen van gewoonten en routines (dorsale striatum) bij obesitasy (Geiger et al. 2009; Wang et al. 2001). Belangrijk is dat de verminderde striatale beschikbaarheid van D2R is gekoppeld aan dwangmatige voedselinname bij obese knaagdieren (Johnson en Kenny 2010) en met verminderde metabole activiteit in OFC en ACC bij obese mensen (Volkow et al. 2008b) (Fig. 7a-c). Gezien het feit dat disfunctie in OFC en ACC resulteert in compulsiviteit [zie review (Fineberg et al. 2009)], kan dit onderdeel zijn van het mechanisme waarmee lage-striatale D2R-signalering hyperfagie mogelijk maakt (Davis et al. 2009). Aangezien verminderde D2R-gerelateerde signalering waarschijnlijk ook de gevoeligheid voor andere natuurlijke beloningen zal verminderen, kan dit tekort bij obese personen ook bijdragen aan compenserende te veel eten. (Geiger et al. 2008).
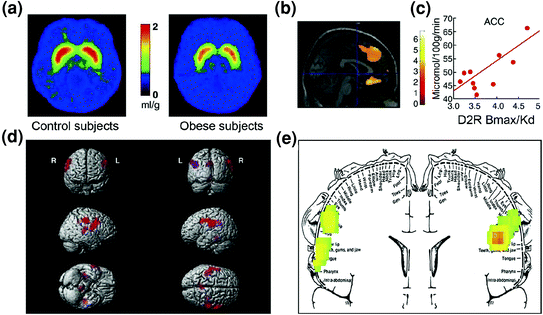
Fig 7 Hyperphagia kan het gevolg zijn van een drive om te compenseren voor een verzwakt beloningscircuit (verwerkt door dopamine-gereguleerde corticostriatale circuits) gecombineerd met een verhoogde gevoeligheid voor smakelijkheid (hedonische eigenschappen van voedsel dat deels door de somatosensorische cortex is verwerkt). a Gemiddelde afbeeldingen voor beschikbaarheid van DA D2-receptor (D2R) bij controles (n = 10) en bij morbide obese proefpersonen (n = 10). b Resultaten van (Statistical Parametric Mapping) SPM die de gebieden in de hersenen identificeerde waar D2R werd geassocieerd met glucosemetabolisme, deze omvatten de mediale OFC, ACC en de dorsolaterale PFC (regio niet getoond). c Regressiehelling tussen striatale D2R en metabolische activiteit bij ACC bij obese proefpersonen. d Driedimensionaal gerenderde SPM-afbeeldingen die de gebieden met een hoger metabolisme tonen bij zwaarlijvige dan bij magere proefpersonen (P <0.003, niet gecorrigeerd). e Kleurgecodeerde SPM-resultaten weergegeven in een coronaal vlak met een gesuperponeerd diagram van de somatosensorische homunculus. De resultaten (z-waarde) worden weergegeven met behulp van de regenboogschaal, waarbij rood> geel> groen. In vergelijking met magere proefpersonen hadden zwaarlijvige proefpersonen een hoger metabolisme bij de baseline in de somatosensorische gebieden waar de mond, lippen en tong vertegenwoordigd zijn en die betrokken zijn bij het verwerken van de smakelijkheid van voedsel. Gewijzigd, met toestemming, van Volkow et al. (2008) (a-c) en Wang et al. (2002) (d, e)
Deze hypothese komt overeen met preklinisch bewijs dat aantoont dat verminderde DA-activiteit in VTA resulteert in een dramatische toename van het gebruik van voedingsmiddelen met een hoog vetgehalte (Stoeckel et al. 2008). Evenzo vertoonden zwaarlijvige personen in vergelijking met mensen met een normaal gewicht foto's van calorierijk voedsel (stimuli waaraan ze werden geconditioneerd) in gebieden die deel uitmaken van belonings- en motivatiecircuits (NAc, dorsal striatum, OFC) verhoogde neurale activering. ACC, amygdala, hippocampus en insula) (Killgore en Yurgelun-Todd 2005). Daarentegen bleek bij normale gewichtscontroles de activering van de ACC en OFC (regio's die betrokken zijn bij salience-attributie die naar het NAc projecteren) tijdens de presentatie van calorierijk voedsel negatief gecorreleerd met hun body mass index (BMI) ( Stice et al. 2008b). Dit suggereert een dynamische interactie tussen de hoeveelheid gegeten voedsel (gedeeltelijk weerspiegeld in de BMI) en de reactiviteit van beloningsregio's tot calorierijk voedsel (weerspiegeld in de activering van OFC en ACC) bij personen met een normaal gewicht, die verloren is gegaan in obesitas.
Verrassend genoeg vertoonden zwaarlijvige personen minder activering van beloningscircuits van de werkelijke voedselconsumptie (aangeduid als beloning met consumptiegoederen) dan magere individuen, terwijl ze een grotere activering van somatosensorische corticale regio's vertoonden die de eetlust verwerken als ze de consumptie verwachtten (Stice et al. 2008b). De laatste observatie correspondeerde met regio's waar een eerdere studie verbeterde activiteit onthulde bij proefpersonen met obesitas getest bij aanvang (niet-stimulering) (Wang et al. 2002) (Fig. 7d, e). Een verhoogde activiteit van regio's die de eetbaarheid verbeteren, kan obese proefpersonen voedsel geven ten opzichte van andere natuurlijke versterkers, terwijl een verminderde activering van dopaminerge doelen door de feitelijke voedselconsumptie kan leiden tot overconsumptie als een middel om te compenseren voor zwakke door D2R gemedieerde signalering (Stice et al. 2008). Deze verminderde respons van het beloningscircuit op voedselconsumptie bij zwaarlijvige proefpersonen doet denken aan de verminderde DA-verhogingen veroorzaakt door drugsgebruik bij verslaafde personen in vergelijking met niet-verslaafde personen.
De prefrontale cortex (PFC) speelt een cruciale rol in de uitvoerende functie, inclusief remmende controle (Miller en Cohen 2001). Deze processen worden gemoduleerd door D1R en D2R (vermoedelijk ook D4R) en dus zal de verminderde activiteit in PFC, zowel bij verslaving als bij obesitas, waarschijnlijk bijdragen aan slechte controle en hoge compulsiviteit. De lager dan normale beschikbaarheid van D2R in het striatum van obese personen, die in verband werd gebracht met verminderde activiteit in PFC en ACC (Volkow et al. 2008b) zal daarom waarschijnlijk bijdragen aan hun gebrekkige controle over voedselinname. Inderdaad, de negatieve correlatie tussen BMI en striatum D2R gerapporteerd bij obesitas (Wang et al. 2001) en bij overgewicht (Haltia et al. 2007) individuen ondersteunen dit. Een beter begrip van de mechanismen die tot een verminderde PFC-functie bij obesitas (of verslaving) leiden, zou de ontwikkeling van strategieën om bepaalde stoornissen in cruciale cognitieve domeinen te verbeteren of misschien zelfs om te keren kunnen vergemakkelijken. Vertragingskorting bijvoorbeeld, die de neiging heeft om een beloning te devalueren als een functie van de tijdelijke vertraging van de toediening, is een van de meest uitgebreid onderzochte cognitieve operaties in relatie tot stoornissen die verband houden met impulsiviteit en compulsiviteit. Vertragingskorting is het meest uitgebreid onderzocht bij drugsgebruikers die een overdreven voorkeur hebben voor kleine maar onmiddellijke over grote maar vertraagde beloningen (Bickel et al. 2007). Echter, de weinige studies uitgevoerd met obese personen hebben ook bewijs blootgelegd van een voorkeur voor hoge, onmiddellijke beloningen, ondanks een verhoogde kans op hogere toekomstige verliezen (Brogan et al. 2010; Weller et al. 2008). En meer recent vond een andere studie een positieve correlatie tussen BMI en hyperbolische discontering, waarbij toekomstige negatieve uitbetalingen minder worden verdisconteerd dan toekomstige positieve uitbetalingen (Ikeda et al. 2010). Interessant is dat uitgestelde discontering afhankelijk lijkt te zijn van de functie van het ventraal striatum (Gregorios-Pippas et al. 2009) en van de PFC, inclusief laterale OFC (Bjork et al. 2009), en is gevoelig voor DA-manipulaties (Pine et al. 2010). Specifiek verhoogde DA-signalering (met L DOPA-behandeling) verhoogde impulsiviteit en temporele verdiscontering.
5 Betrokkenheid van motivatiecircuits
Dopaminerge signalering moduleert ook de motivatie. Gedragstrekken zoals vitaliteit, doorzettingsvermogen en het investeren van een voortdurende inspanning om een doel te bereiken, zijn allemaal onderhevig aan modulatie door DA via verschillende doelregio's, waaronder NAc, ACC, OFC, DLPFC, amygdala, dorsal striatum en ventrale pallidum (Salamone et al. 2007). Dysregulated DA-signalering wordt geassocieerd met een verhoogde motivatie om medicijnen aan te schaffen, een kenmerk van verslaving. Daarom nemen drugsverslaafden vaak extreem gedrag om medicijnen te verkrijgen, zelfs als ze bekende ernstige en nadelige gevolgen met zich meebrengen (Volkow en Li 2005). Omdat het nemen van drugs de belangrijkste drijfveer wordt bij drugsverslaving (Volkow et al. 2003), verslaafde onderwerpen worden gewekt en gemotiveerd door het proces om het medicijn te verkrijgen, maar hebben de neiging zich teruggetrokken en apathisch te voelen wanneer ze worden blootgesteld aan niet-drugsgerelateerde activiteiten. Deze verschuiving is bestudeerd door de hersenactiveringspatronen die optreden bij blootstelling aan geconditioneerde aanwijzingen te vergelijken met die die optreden bij afwezigheid van dergelijke signalen. In tegenstelling tot de afname van prefrontale activiteit gerapporteerd in gedetoxificeerde cocaïneverslaafden wanneer ze niet worden gestimuleerd met drugs- of medicijn-aanwijzingen [zie recensie (Volkow et al. 2009)], worden deze prefrontale regio's geactiveerd wanneer cocaïne misbruikers worden blootgesteld aan hunkering-inducerende stimuli (drugs of cues) (Grant et al. 1996; Volkow et al. 1999; Wang et al. 1999). Dit resultaat doet denken aan de waarneming dat cocaïne-misbruikers, kort na een episode van cocaïnebeking bestudeerd, een toename in metabolische activiteit vertoonden in OFC en ACC (ook dorsale striatum) die geassocieerd was met hunkering (Volkow et al. 1991).
Bovendien, wanneer de responsen op iv MP worden vergeleken tussen cocaïne-verslaafde en niet-verslaafde individuen, reageerde de eerstgenoemde met verhoogd metabolisme in ventrale ACC en mediale OFC (een effect geassocieerd met hunkering), terwijl de laatste de tegenovergestelde reactie vertoonde, namelijk afgenomen metabolisme in deze regio's (Volkow et al. 2005). Dit suggereert dat de activering van deze prefrontale gebieden met blootstelling aan drugs specifiek kan zijn voor verslaving en geassocieerd is met de verhoogde behoefte aan het medicijn. Bovendien toonde een onderzoek dat cocaïneverslaafde proefpersonen aanzette om hunkering te remmen wanneer ze werden blootgesteld aan medicijncues, aan dat die proefpersonen die succesvol waren bij het remmen van hunkering, een verlaagd metabolisme vertoonden in mediale OFC (die de motivatiewaarde van een versterker verwerkt) en NAc (wat voorspelt). beloning) (Volkow et al. 2009b). Deze bevindingen bevestigen verder de betrokkenheid van OFC, ACC en striatum in de verhoogde motivatie om het medicijn bij verslaving te krijgen.
Het is te voorspellen dat de OFC ook betrokken is geweest bij het toekennen van de waarde van opvallendheid aan voedsel (Grabenhorst et al. 2008; Rolls en McCabe 2007), helpen om de verwachte aangenaamheid en smakelijkheid te beoordelen als een functie van de context. PET-onderzoeken met FDG om het glucosemetabolisme bij hersenweefsel te meten bij personen met een normaal gewicht rapporteerden dat blootstelling aan voedsel-cues de metabolische activiteit verhoogde in OFC, wat een effect was dat geassocieerd was met de perceptie van honger en de behoefte aan voedsel (Wang et al. 2004). De verhoogde OFC-activering door de voedselstimulering weerspiegelt waarschijnlijk de stroomafwaartse dopaminerge effecten en neemt deel aan de betrokkenheid van DA bij het streven naar voedselconsumptie. Het OFC speelt een rol bij het leren van stimulusversterkende associaties en conditionering (Cox et al. 2005; Gallagher et al. 1999), ondersteunt geconditioneerde cue uitgelokte voeding (Weingarten 1983) en draagt waarschijnlijk bij aan te veel eten, ongeacht de hongersignalen (Ogden en Wardle 1990). Inderdaad, disfunctie van de OFC is in verband gebracht met overeten (Machado en Bachevalier 2007).
Ondanks enkele inconsistenties tussen studies, ondersteunen gegevens over hersenafbeeldingen ook het idee dat structurele en functionele veranderingen in hersengebieden die betrokken zijn bij executieve functies (inclusief remmende controle) geassocieerd kunnen zijn met hoge BMI bij verder gezonde personen. Een MRI-studie bij oudere vrouwen bijvoorbeeld, waarbij gebruik werd gemaakt van op voxel gebaseerde morfometrie, vond een negatieve correlatie tussen BMI- en grijze-stofvolumes (inclusief frontale regio's), die in de OFC geassocieerd was met een verminderde uitvoerende functie (Walther et al. 2010). Met behulp van PET om het hersen-glucosemetabolisme in gezonde controles te meten, rapporteerden we een negatieve correlatie tussen BMI en metabole activiteit in DLPFC, OFC en ACC. In deze studie voorspelde de metabole activiteit in prefrontale regio's de prestaties van de proefpersonen in tests van executieve functies (Volkow et al. 2009c). Evenzo toonde een nucleaire magnetische resonantie (NMR) spectroscopische studie bij gezonde controles op middelbare leeftijd en bij ouderen aan dat BMI negatief was geassocieerd met de niveaus van N-acetyl-aspartaat (een marker van neuronale integriteit) in de frontale cortex en ACC (Gazdzinski et al. 2008; Volkow et al. 2009c).
Hersenbeeldvormingsstudies waarin obese en magere individuen worden vergeleken, hebben ook een lagere grijze materiedichtheid in frontale gebieden gemeld (frontale operculum en midden frontale gyrus) en in post-centrale gyrus en putamen (Pannacciulli et al. 2006). Een ander onderzoek vond echter geen verschillen in volumes van grijze stoffen tussen obese en magere patiënten. het registreerde wel een positieve correlatie tussen het volume witte stof in basale hersenstructuren en de ratio's tussen taille en heup, een trend die gedeeltelijk werd ongedaan gemaakt door een dieet (Haltia et al. 2007b). Interessant is dat corticale gebieden, zoals de DPFC en OFC die betrokken zijn bij remmende controle, ook zijn geactiveerd bij succesvolle lijners in reactie op maaltijdconsumptie (DelParigi et al. 2007), wat duidt op een potentieel doelwit voor gedrags-omscholing bij de behandeling van obesitas (en ook bij verslaving).
6 Betrokkenheid van interoceptieve schakelingen
Neuroimaging-onderzoeken hebben aangetoond dat de middelste insula een cruciale rol speelt in hunkeren naar voedsel, cocaïne en sigaretten (Bonson et al. 2002; Pelchat et al. 2004; Wang et al. 2007). Het belang van de insula is benadrukt door een onderzoek dat meldde dat rokers met schade aan deze regio (maar niet controle rokers die last hadden van extra-insulaire laesies) gemakkelijk konden stoppen met roken en zonder cravingen of terugval te ervaren (Naqvi et al. . 2007). De insula, in het bijzonder de meer voorste regio's, is wederzijds verbonden met verschillende limbische regio's (bijv. Ventromediale prefrontale cortex, amygdala en ventraal striatum) en lijkt een interoceptieve functie te hebben, waarbij de autonome en viscerale informatie wordt gecombineerd met emotie en motivatie, waardoor bewuste bewustwording van deze driften (Naqvi en Bechara 2009). Onderzoek naar hersenlaesies suggereert inderdaad dat de ventromediale PFC en insula noodzakelijke componenten zijn van de verdeelde circuits die emotionele besluitvorming ondersteunen (Clark et al. 2008). Consistent met deze hypothese tonen beeldvormingsstudies consequent differentiële activering van de insula tijdens hunkering (Brody et al. 2009; Goudriaan et al. 2010; Naqvi en Bechara 2009; Wang et al. 1999). Dienovereenkomstig is gesuggereerd dat de reactiviteit van dit hersengebied kan dienen als een biomarker om terugval te helpen voorspellen (Janes et al. 2010).
De insula is ook een primaire smaakmakerij, die deelneemt aan vele aspecten van eetgedrag, zoals smaak. Bovendien verschaft de rostrale insula (verbonden met de primaire smaakcortex) informatie aan de OFC die van invloed is op de multimodale weergave van de aangenaamheid of de beloningswaarde van binnenkomend voedsel (Rolls 2008). Vanwege de betrokkenheid van de insula in de interoceptieve zin van het lichaam, in emotioneel bewustzijn (Craig 2003) en in motivatie en emotie (Rolls 2008), zou een bijdrage van insulaire stoornissen in obesitas te verwachten zijn. Inderdaad resulteert de uitzetting van de maag in activering van de posterieure insula, die waarschijnlijk zijn rol weerspiegelt in het bewustzijn van lichaamstoestanden (in dit geval van volheid) (Wang et al. 2008). Bovendien resulteerde in mager, maar niet in zwaarlijvige personen de uitzetting van de maag in activering van de amygdala en deactivering van de voorste insula (Tomasi et al. 2009). Het gebrek aan amygdala-respons bij zwaarlijvige personen kan een afgestompt interoceptief bewustzijn van lichaamstoestanden in verband met verzadiging (volle maag) weerspiegelen. Hoewel de modulatie van insulaire activiteit door DA slecht is onderzocht, wordt erkend dat DA betrokken is bij de reacties op het proeven van smakelijke voedingsmiddelen die worden gemedieerd door de insula (Hajnal en Norgren 2005). Humane beeldvormingsstudies hebben aangetoond dat het proeven van smakelijke voedingsmiddelen de gebieden met insula en middenhersenen activeerde (DelParigi et al. 2005; Frank et al. 2008). De DA-signalering kan echter ook nodig zijn voor het detecteren van het caloriegehalte van voedsel. Toen vrouwen met een normaal gewicht bijvoorbeeld een zoetstof proefden met calorieën (sucrose), werden zowel de insula- als dopaminerge middenhersenengebieden geactiveerd, terwijl het proeven van een calorie-vrije zoetstof (sucralose) alleen de insula activeerde (Frank et al. 2008). Obese proefpersonen vertonen een grotere insulaire activatie dan normale controles bij het proeven van een vloeibare maaltijd die bestaat uit suiker en vet (DelParigi et al. 2005). Daarentegen vertonen proefpersonen die hersteld zijn van anorexia nervosa minder activatie in de insula bij het proeven van sucrose en geen associatie van aangename gevoelens met insulaire activatie zoals waargenomen in de normale controles (Wagner et al. 2008). Wanneer ze gecombineerd worden, maken deze resultaten het waarschijnlijk dat ontregeling van de insula als reactie op smaakstimuli betrokken kan zijn bij de verminderde controle van verschillende eetlustgevoelens.
7 Het schakelsysteem van afkeer
Zoals eerder vermeld, leidt training (conditionering) op een cue die beloning voorspelt ertoe dat dopaminerge cellen vuren als reactie op de voorspelling van beloning, en niet op de beloning zelf. Aan de andere kant, en consistent met deze logica, is waargenomen dat dopaminerge cellen minder dan normaal zullen vuren als de verwachte beloning niet uitkomt (Schultz et al. 1997). Cumulatief bewijs (Christoph et al. 1986; Lisoprawski et al. 1980; Matsumoto en Hikosaka 2007; Nishikawa et al. 1986) verwijst naar de habenula als een van de regio's die de afname in het afvuren van dopaminerge cellen in VTA regelt die kan volgen op het niet ontvangen van een verwachte beloning (Kimura et al. 2007). Aldus zou een verhoogde gevoeligheid van de habenula, als een gevolg van blootstelling aan chronische geneesmiddelen, ten grondslag kunnen liggen aan een grotere reactiviteit ten opzichte van geneesmiddelaanwijzingen. Inderdaad, activering van de habenula, in cocaïne-verslaafde personen, is in verband gebracht met gedragsmatige terugval naar het nemen van geneesmiddelen na cue-blootstelling (Brown et al. 2011; Zhang et al. 2005). In het geval van nicotine lijken de α5 nicotinereceptoren in de habenula de aversieve respons op grote doses nicotine te moduleren (Fowler et al. 2011); en de α5- en α2-receptoren in de habenula zijn betrokken bij nicotineontwenning (Salas et al. 2009). Vanwege de tegengestelde respons van de habenula op die van DA-neuronen om te belonen (deactivering) en de activering ervan bij blootstelling aan aversieve stimuli, verwijzen we hier naar de habenula-signalering als een die een "antireward" -invoer overdraagt.
De habenula lijkt een vergelijkbare rol te spelen met betrekking tot voedselbeloning. Een zeer smakelijk voedingsdieet kan bij ratten obesitas veroorzaken, waarbij de gewichtstoename correleert met de toename van de binding van μ-opioïde peptiden in de basolaterale en basomediale amygdala. Interessant genoeg vertoonde de mediale habenula significant hogere μ-opioïde peptidebinding (met ongeveer 40%) na blootstelling aan het smakelijke voedsel in de ratten die aankwamen (die die meer voedsel consumeerden) maar niet in die die dat niet deden (Smith et al. 2002). Dit suggereert dat de habenula betrokken kan zijn bij overeten onder omstandigheden van beschikbaarheid van smakelijk voedsel. Bovendien projecteren neuronen in de rostromediale tegmentale kern, die een belangrijke input van de laterale habenula ontvangen, naar VTA DA-neuronen en worden geactiveerd na voedseldeprivatie (Jhou et al. 2009). Deze bevindingen komen overeen met een rol voor de habenula bij het bemiddelen van reacties op aversieve stimuli of toestanden, zoals die zich voordoen tijdens het volgen van een dieet of het ontwennen van geneesmiddelen.
De betrokkenheid van de habenula als een antireward-knooppunt in emotionele netwerken is consistent met eerdere theoretische verslavingsmodellen die gepersitiseerde anti- beloningsreacties stelden (gemedieerd door verhoogde gevoeligheid van de amygdala en verhoogde signalering door de corticotropine-afgevende factor) als drugsverslaving bij verslaving (Koob en Le Moal 2008). Vergelijkbare antireward-reacties kunnen ook bijdragen aan overmatige voedselconsumptie bij obesitas.
8 Pathologische geneesmiddelen- en voedselbeloning: een aangepast werkmodel
THet vermogen om weerstand te bieden aan de drang om een medicijn te gebruiken of te eten voorbij het punt van verzadiging vereist de juiste werking van de neuronale circuits die betrokken zijn bij top-down controle om de geconditioneerde reacties die de beloning van inname van het voedsel / medicijn en de wens om de food / drugs. Hier hebben we zes van deze circuits belicht: beloning / oplettendheid, conditionering / gewoonten, remmende controle / executieve functie, motivatie / drive, interoceptie en aversie vermijden / stress-reactiviteit (Fig. 8). Op basis van de hier gepresenteerde beeldgegevens postuleren we dat het de discrepantie is tussen de verwachting voor de medicatie / voedseleffecten (geconditioneerde reacties) en de afgestompte neurofysiologische effecten die het innemen van medicijnen of de overconsumptie van voedsel in een poging om de verwachte beloning. Ook, al dan niet getest tijdens vroege of langdurige perioden van onthouding / diëten, vertonen verslaafde / obese personen een lagere D2R in striatum (inclusief NAc), die geassocieerd zijn met verlagingen van baseline-activiteit in frontale hersengebieden die betrokken zijn bij salience-attributie (orbitofrontale cortex) en remmende controle (ACC en DLPFC), waarvan de verstoring resulteert in compulsiviteit en impulsiviteit. Fin feite is er ook bewijs naar voren gekomen over de rol van interoceptieve en aversieve schakelingen in de systemische onevenwichtigheden die resulteren in de dwangmatige consumptie van drugs of voedsel.
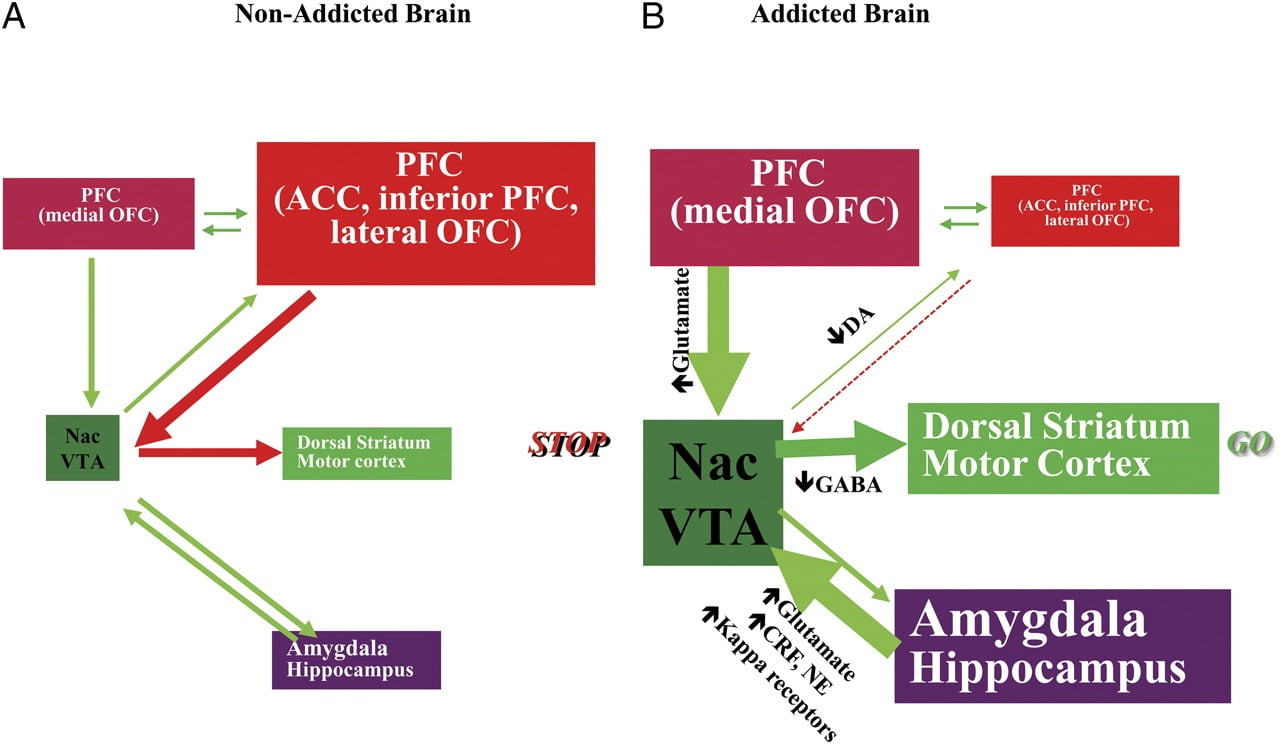
Fig 8 Model met een netwerk van interacterende circuits, verstoringen die bijdragen aan de complexe set stereotypische gedragingen die ten grondslag liggen aan drugsverslaving en chronische overeten: beloning (nucleus accumbens, VTA en ventrale pallidum), conditionering / geheugen (amygdala, mediale OFC voor toewijzing van oprispingen, hippocampus en dorsale striatum voor gewoonten), uitvoerende controle (DLPFC, ACC, inferieure frontale cortex en laterale OFC), motivatie / drive (mediale OFC voor toewijzing van saliency, ventrale ACC, VTA, SN, dorsale striatum en motorische cortex). Nac, nucleus accumbens, interoceptie (Insula en ACC) en afkeer / vermijding (Habenula). a Wanneer deze circuits in balans zijn, resulteert dit in de juiste remmende controle en besluitvorming. b Tijdens verslaving, wanneer de verhoogde verwachtingswaarde van het medicijn in de beloning, motivatie en geheugencircuits het controlecircuit overwint, waarbij een positieve feedbacklus wordt bevorderd die wordt geïnitieerd door de consumptie van het medicijn en bestendigd door de verbeterde activering van de motivatie / drive en geheugencircuits. Deze circuits hebben ook interactie met schakelingen die betrokken zijn bij stemmingsregulatie, waaronder stressreactiviteit (waarbij de amygdala, hypothalamus, habenula) en interoceptie (waarbij de insula en ACC zijn betrokken en bijdraagt tot het bewustzijn van hunkering). Verschillende neurotransmitters zijn betrokken bij deze neuroadaptaties, waaronder glutamaat, GABA, norepinefrine, corticotropine-afgevende factor en opioïde receptoren. CRF, corticotropin-releasing factor; NE, norepinephrine. Gewijzigd met toestemming van Volkow et al. (2011b)
Als gevolg van de opeenvolgende verstoring van deze circuits, kunnen individuen 1 ervaren) een verbeterde motivationele waarde van het medicijn / voedsel (secundair aan geleerde associaties door conditionering en gewoonten) ten koste van andere versterkers (secundair aan verminderde gevoeligheid van het beloningscircuit) ), 2) een verminderd vermogen om de opzettelijke (doelgerichte) acties veroorzaakt door de sterke drang om het medicijn / voedsel (secundair aan een verminderde executieve functie) dat resulteert in dwangmatig innemen van medicijnen en voedsel en 3) te verminderen, verhoogde stressreactiviteit en aversieve vermijding die resulteert in impulsief drugsgebruik om aan de aversieve toestand te ontsnappen.
Dit model suggereert een meervoudige therapeutische benadering van verslaving die ontworpen is om de versterkende eigenschappen van drugs / voedsel te verminderen, de belonende eigenschappen van natuurlijke versterkers te herstellen / verbeteren, geconditioneerde geleerde associaties te remmen, de motivatie voor niet-medicamenteuze / voedselgerelateerde activiteiten te vergroten, stressreactiviteit te verminderen , verbeter de stemming en versterk de algehele remmende controle.
Dankwoord
De auteurs willen de steun van het intramurale programma van de NIAAA van de National Institutes of Health bedanken.
Referenties