KOMMENTARER: Kontroversiell delen av dette papiret inneholder følgende:
Det finnes en rekke elektrofysiologiske baserte studier fra Prauses gruppe som gir bevis på at seksuell lyst, ikke hyperseksualitet, forutsier selvregulering av seksuell opphisselse. [92]. Disse etterforskerne antyder i annet arbeid at personer som rapporterte problemer som regulerer deres syn på visuell seksuell stimuli (VSS), som også rapporterte høyere seksuell lyst, viste lavere sent-positive potensialer (LPP) som svar på VSS. Forfatterne foreslår at dette mønsteret fremstår annerledes enn stoffavhengighetsmodeller [93].
De to EEG-papirene fra "Prauses gruppe" har blitt kritisert grundig i fagfellevurderte tidsskrifter og av andre (inkludert YBOP). I virkeligheten er funnene i begge papirene i samsvar med avhengighetsmodellen, til tross for forfatternes tvilsomme konklusjoner. For EEG-studien fra 2013, se:
- Ingenting korrelerer med ingenting i SPAN Labs nye pornostudie (2013)
- Fagfellevurdert kritikk: 'High Desire', eller 'Bare' An Addiction? A Response to Steele et al. (2014) av Donald L. Hilton, Jr., MD,
- Fagfellevurdert kritikk: "Neuroscience of Internet Pornography Addiction: A Review and Update" (2015)
- Fagfellevurdert analyse: “Nevrale korrelater av seksuell køreaktivitet hos personer med og uten tvangsmessig seksuell atferd” (2014)
For 2015 EEG-studien se:
- Kritikk av "Modulering av sen positive potensialer av seksuelle bilder hos brukere av brukere og kontroller som ikke er i samsvar med 'Pornavhengighet' (2015)"
- Fagfellevurdert kritikk: Neuroscience of Internet Pornography Addiction: A Review and Update (2015)
Ville det ikke vært bra om forskere faktisk leste studiene de siterer i stedet for bare å papegøye hovedforfatterens overskrifter?
LINK TIL PUBLIKASJON
Publisert: Oktober 12, 2015 (se historie)
DOI: 10.7759 / cureus.348
Oppgi denne artikkelen som: Blum K, Badgaiyan RD, Gold MS (October 12, 2015) Hyperseksualitetsavhengighet og tilbaketrekning: Fenomenologi, nevrogenetikk og epigenetikk. Cureus 7 (10): e348. doi: 10.7759 / cureus.348
Abstrakt
Hyperseksualitet har blitt definert som unormalt økt seksuell aktivitet. Epidemiologiske og kliniske studier har vist at denne ikke-parafile tilstanden består av "overdreven" seksuell atferd og lidelser ledsaget av personlig nød og sosial og medisinsk sykelighet. Det er et veldig kontroversielt og politisk tema når det gjelder hvordan man best kan kategorisere det som lik eller ikke lik vanedannende atferd, inkludert rusmisbruk. Hyperseksuell lidelse er konseptualisert som en ikke-parafil seksuell lystlidelse med impulsivitet. Patofysiologiske perspektiver inkluderer dysregulering av seksuell opphisselse og lyst, seksuell impulsivitet og seksuell kompulsivitet. Kjernen accumbens, som ligger i ventral striatum, formidler de forsterkende effektene av misbruksdroger, som kokain, alkohol, nikotin og mat samt musikk. Det antas at denne strukturen mandater atferd fremkalt av insentivstimuli. Disse atferdene inkluderer naturlige belønninger som fôring, drikking, seksuell oppførsel og utforskende bevegelse. En viktig regel for positiv forsterkning er at motoriske responser vil øke i styrke og styrke hvis de følges av en givende hendelse. Her antar vi at det er en felles virkningsmekanisme (MOA) for de kraftige effektene narkotika, musikk, mat og sex har på menneskelig motivasjon. Menneskets drivkraft for de tre nødvendige motivasjonsatferdene "sult, tørst og sex" kan alle ha vanlige molekylære genetiske antecedenter som, hvis de er svekket, fører til avvikende atferd. Vi antar at basert på en mengde vitenskapelig støtte hyperseksuell aktivitet faktisk er som narkotika, mat og musikk som aktiverer hjernens mesolimbiske belønningskretser. Videre påvirker dopaminerge genet og muligens andre kandidat neurotransmitterrelaterte genpolymorfier både hedoniske og anhedoniske atferdsmessige resultater. Det er lite kjent om både genetikk og epigenetikk av hyperseksualitet i dagens litteratur. Vi forventer imidlertid at fremtidige studier basert på vurderinger med kliniske instrumenter kombinert med genotyping av sexmisbrukere vil gi bevis for spesifikk gruppering av seksuelle typologier med polymorfe assosiasjoner. Det har vært noen studier som bruker elektrofysiologiske teknikker som ikke støtter synspunktet om at hyperseksualitet faktisk ligner på rusmisbruk og andre atferdsmisbruk. Forfatterne oppfordrer også både kliniske og akademiske forskere til å gå i gang med forskning ved hjelp av nevroavbildningsverktøy for å undersøke naturlige dopaminerge agonistiske midler rettet mot spesifikke genpolymorfier for å "normalisere" hyperseksuell oppførsel.
Introduksjon og bakgrunn
Det er klart at hyperseksuell atferd er dokumentert innen kliniske og forskningsmessige omgivelser det siste tiåret [1]. Benjamin Rush, lege, en av USAs grunnleggende far, dokumenterte klinisk overdreven seksuell atferd [2] Richard von Krafft-Ebing, en vesteuropeisk pioner-sexolog fra det 19. århundre, og Hirshfeld i 19 fortsatte begge arbeidet [3-4]. Den grunnleggende leietakeren til disse etterforskerne antydet at hyperseksualitet utgjorde vedvarende sosialt avvikende seksuell atferd hos menn og kvinner, med overdreven seksuell matlyst. I 1975 karakteriserte barnevognen tilstanden som Don Juanism [5]. I 1969 foreslo Allen satyriasis for menn og nymfomani hos kvinner støttet av Ellis og Sagarin [6-7]. Selv om hyperseksualitet ikke er inkludert som en psykiatrisk diagnose i DSM, kan arbeid fra en rekke samtidige etterforskere, inkludert Kafka, Reid, Bancroft, deres kolleger og tilbøyeligheten til Verdens helseorganisasjon føre til inkludering av denne tilstanden som en egen diagnostisk enhet [8-11].
Anmeldelse
Litteraturmetodikk
Medline-databasen, per 12. juli 2015, ble brukt til å utføre et internettbasert litteratursøk. Følgende termer ble inkludert: hypersexual (170), hypersexuality (479), seksuell avhengighet (1,652), sexmisbruker (1,842), seksuell impulsivitet (989), compulsive sexual (946), compulsive sex (1,512 782), seksuell tvang (234) ), parafili-relatert lidelse (857) og overdreven seksuell (17). Siden denne artikkelen er en kort gjennomgang i stedet for en metaanalyse, er den basert på et representativt utvalg av disse studiene som har relevans for de underemnene som dekkes. Manglende inkludering av en bestemt studie negerer ikke dens betydning for feltet. Det er forståelig nok de som ikke er enige i konseptet om at sexavhengighet faktisk er en reell lidelse, og som til og med kan ha bevis for at de er på solid grunn. Vi argumenterer imidlertid for at det er rikelig med bevis for at seksuell avhengighet eksisterer, og studier som involverer nevroavbildning, nevrogenetikk og til og med epigenetikk støtter forestillingen om at tvangsmessig sexavhengighet samt hyperseksualitet kan betraktes som en vanedannende lidelse. Selv om vi er klar over dette avviket, legger vi henholdsvis vårt synspunkt for å generere ytterligere vitenskapelig gransking og ikke for å fjerne vitenskapelig retorikk om dette viktige emnet. For å belyse emnet ytterligere har vi søkt i litteraturen for å få enighet. 2015. august 46 søkte vi i PUBMED Central med følgende ord - "Er sex en reell avhengighet?" og hentet XNUMX artikler.
Definisjon av seksuell avhengighet
Seksuell avhengighet er definert som enhver tvangsmessig seksuell atferd som forstyrrer det normale levebrødet og forårsaker alvorlig stress på familien, venner, kjære og ens arbeidsmiljø. Seksuell avhengighet har blitt kalt seksuell avhengighet, hyperseksualitet og seksuell tvangsmessighet. Uansett hvilket som helst navn, er det en tvangsmessig oppførsel som fullstendig dominerer den narkomanes liv. Seksuelle rusavhengige gjør sex til en prioritet viktigere enn familie, venner og arbeid. Sex blir det organiserende prinsippet i narkomanes liv. De er villige til å ofre det de verdsetter mest for å bevare og fortsette sin usunne oppførsel [12]. Hyperseksuell lyst er blitt avgrenset som ønske basert på en livslang vurdering av hyppigheten av seksuell atferd og tid brukt i tilhørende seksuelle fantasier. Hos menn ble en evaluering av hyperseksuell lyst definert av Kafka og Hannen som den høyeste vedvarende perioden (minst seks måneder minimum varighet) av vedvarende vedtatt seksuell atferd (total seksuell utløp / Uke etter alder 15). Faktisk ble en longitudinell historie med hyperseksuell lyst, operasjonelt definert som ovenfor, identifisert hos 72-80% av menn som søker behandling for parafilier og parafili-relaterte lidelser [13].
Hyperseksualitet og kjønnsforskjeller
Det er veletablert at i det menneskelige seksuelle samfunnet og litteraturen er seksuell lyst kapitulert som tilstedeværelsen av seksuelle fantasier, aktiviteter eller trang, og motivasjon fra mennesket til å delta i seksuell atferd. Det er både interne og eksterne relevante signaler [14]. Tilhengere av evolusjonsteorien har hevdet at menn og kvinner har forskjellige agendaer når det gjelder seksuell aktivitet [15]. Tallrike studier avdekker tydelige forskjeller mellom menn og kvinner. Menn har økt seksuell fantasi [16], økt frekvens av onani [17], økt tilbøyelighet for eksternt generert visuell seksuell opphisselse [18] tillatte holdninger til tilfeldig sex [19], letthet med opphisselse [20], og iboende motivasjon [21]. I kontrast viser kvinner et annet seksuelt landskap med seksuell motivasjon, seksuell opphisselse og seksuell atferd formet av evolusjonsfaktorer [22] og større biologisk, emosjonell og tidsmessig investering i reproduksjon og barneoppdragelse [23]. Kvinner er mindre sårbare for hyperseksualitet [24] og tilpasset for å fremme tilknytningsrelasjoner og langsiktig partnerengasjement [25]. Mens seksuell avhengighet anslås å plage opptil 3% til 6% av befolkningen, er den klare forståelsen av de nevrobiologiske antecedentene begrenset [26] samt kliniske vurderinger [27]. Vi oppfordrer til videre lesning om seksuell tvang, tilknytning og seksuell legning [28], og kjønnsforskjeller i svar på seksuelle stimuli [29-30].
Det er bemerkelsesverdig at Kafka og Hennen [13], fant at gjennomsnittsalderen for begynnelse av vedvarende hyperseksuell atferd var 18.7 ± 7.2 år hos seksuelt aktive menn og aldersområdet for begynnelse av hyperseksuell atferd var alder 7-46. Gjennomsnittlig varighet av denne høyeste konsekvent vedlikeholdte frekvensen av seksuell appetittlig oppførsel var 12.3 ± 10.1 år. Imidlertid var middelalderen for disse aktive seksuelle menn som var hyperseksuell som søkte behandling, 37 ± 9 år. Hanson, et al. vurderte også hyperseksualitet hos lovbrytere og fant at lave lovbrytere hadde lavere tilbakefall enn lovbrytere med høy risiko. [31].
Hyperseksualitet og co-morbid stoffmisbruk
Det er en høy komorbiditet mellom hyperseksuell lidelse og andre avhengighet, som for eksempel rusmiddelforstyrrelse [32-33]. Spesifikt foreslo Garcia og Thibaut at fenomenologien til overdreven ikke-parafilisk seksuell forstyrrelse skulle klassifiseres som en vanedannende atferd, snarere enn en tvangstanker eller en impulsstyringsforstyrrelse [34]. De påpeker riktig at kriteriene er ganske nær kriteriene for avhengighetsforstyrrelser som også foreslått av andre [35]. Disse etterforskerne har gitt drivkraft for fortsatt forskning på dette området og mulig fremtidig inkludering av hyperseksuell lidelse i DSM-6. Narkotikamisbruk, rock 'n' roll og sex forekommer samtidig, og hele festivaler har blitt bygget rundt disse kombinasjonene fra Woodstock til i dag.
Litteraturen avslører at brukere av metamfetamin rapporterer at dette stimulerende stoffet øker seksuell lyst, spesielt risikabel atferd. Imidlertid har amfetamin vist seg å redusere den seksuelle aktiviteten til hunnrotter. Med dette i bakhodet holder Holder, et al. evaluert metamfetaminets rolle i hunnrotter [36]. De fant ut at tvert imot, metamfetamin letter kvinnelig seksuell atferd, og denne effekten skyldes forbedring av dopaminerg overføring og til og med mulig neurotransmisjon på grunn av kombinasjonen av eggstokkhormoner og metamfetamin. Spesielt fant de en forbedring av seksuell motivasjon kombinert med aktivering av nevronaktivitet i den mediale amygdala og ventromedialkjernen i hypothalamus.
Videre har forskere fra Nederland studert co-morbiditeten av stoffmisbruk hos selvidentifiserte swingers [37]. I denne studien har Spauwen, et al. konkluderte med at 79% av swingers rapporterte fritidsbruk av medisiner (inkludert alkohol og bruk av erektil dysfunksjonsmedisiner); 46% av dem rapporterte om flere legemiddelbruk. Faktisk var bruk av fritidsbruk (unntatt alkohol og medisiner med erektil dysfunksjon) betydelig forbundet med seksuell atferd hos menn og kvinner. Medikamentbruk ble også uavhengig assosiert med seksuelt overførbare infeksjoner (STI) hos kvinnelige swingers, spesielt de som deltar i gruppesex.
Castelo-Branco, et al. rapporterte at unge voksne kvinner oppfatter at seksualitet er en viktig del av livet, men ikke en primær bekymring (77.6%) [38]. De rapporterte også at alkohol fjerner hindringene for å ha sex (62.3%). Viktigere, de fant også ut at alkoholmisbruk var en prediktiv variabel for å øke risikofylt atferd uavhengig av kvinnens alder.
Det er bemerkelsesverdig at Jia, et al. rapporterte om farlig seksuell atferd blant psykostimulerende og heroinovergripere, inkludert flere seksuelle omganger, tilfeldige seksuelle partnere, homoseksuelle partnere og aldri eller noen ganger praktisere sikker sex [39].
Vår viktigste leietaker er at medikamenter, som metamfetamin, kokain, heroin og alkohol, kan stimulere seksuell lyst hos ikke-rusavhengige. Hos rusavhengige er det ganske annerledes; de samme medisinene kan forårsake anhedoni på kronisk basis. Imidlertid har man i mange tilfeller blitt observert afrodisiakumlignende atferd etter avhengighet i utvinningsfasen.
Hyperseksualitet og tilbaketrekning
Et PubMed-søk (7-19-15) med betegnelsen "hyperseksualitet og abstinenssymptomer" resulterte i bare fem artikler, hvorav ingen beskrev "abstinenssymptomatologi." Et alternativt søk med begrepene "abstinenssymptomer med høy seksuell aktivitet" resulterte imidlertid i 25 oppførte artikler.
Rusavhengige i bedring rapporterer om økning i spising og appetitt for visse matvarer og misbruk av sigaretter under langvarig avholdenhet. Vektøkning er også påvist og dokumentert hos nylig og langvarige avholdsrike dyr og mennesker [39]. Bruijnzeel gjorde den interessante observasjonen at akutt tilbaketrekning av opiat kan resultere i spontane orgasmer [40]. Det er viktig at Bruijnzeel i artikkelen foreslo at abstinenssymtomatologi fra medisiner og muligens kronisk intensiv seksuell aktivitet kan skyldes den uopplagte funksjonen til kappa opioidreseptorsignalering som hemmer frigjøring av dopamin mens økning av noradrenalin i hjernebelønningskretser.
Artiklene i litteraturen led av mangel på strenghet angående akutt og langvarig abstinens og avholdenhet bekreftet ved urintesting. Kliniske behandlingsprogrammer har i økende grad beveget seg fra å behandle begge kjønn til å isolere de to kjønnene, og tilby kjønnsspesifikke behandlingsprogrammer. De tilbyr også utdanning for å møte de tidlige og langvarige økningene i seksuell interesse og aktivitet, og forholdet mellom utvinning til mat og overspising.
Hyperseksualitetsinduserte abstinenssymptomer er rapportert av noen etterforskere med ulik grad av alvorlighetsgrad og co-sykelig stoffmisbruk [41-45]. Som et resultat av dette søket fant vi ikke en eneste artikkel som beskrev faktiske abstinenssymptomer forbundet med avholdenhet fra aktive seksuelle møter. De fleste av de sitater som ble sitert involverte effekten av abstinens fra misbruksmedisiner, som opioider, nikotin, amfetamin og kokain, noe som kan svekke seksuell aktivitet.
Hyperseksualitet og nevrogenetikk
Et PubMed-søk (7-19-15) avslørte bare seks oppførte artikler med begrepet "gener og hyperseksualitet", med hovedvekt på artikler relatert til Kleine-Levin syndrom (KLS), en svært sjelden sykdom der hyperseksualitet kan vare opptil 27 år. I en studie ble det funnet at en immunresponsiv HLA-DQBl, DQBl * 0602 ble påvist i betydelige mengder hos pasienter med KLS og kunne øke risikoen for KLS [46-47].
Imidlertid, når vi brukte begrepene "seksuell aktivitet og gener", ble 2,826 XNUMX artikler oppført, og vi gir en kort sammendrag av noen få viktige nevrogenetiske aspekter. Det er vår hypotese at både hedonisk og anhedonisk oppførsel er et resultat av en persons risikoalleler for denne oppførselen, og at behandlingen består av å målrette disse identifiserte polymorfismene riktig. Videre avhenger behandlingsrespons også av disse risikoallelene og gir en viktig begrunnelse for farmakogenetisk testing og farmakogenomiske / nutrigenomiske løsninger.
Etter det kontroversielle innledende funnet av Blum, et al. i 1990 av det første beviset for en assosiasjon mellom DRD2 Al-allelen og alvorlig alkoholisme, har det vært 3,938-artikler i PubMed (7-19-15) [48]. Studiene dekker den psykiatriske genpolymorfismen, DRD2 Al-allelet, og tilhørende atferd og fysiologi. Det er imidlertid mangel på data som knytter seksuell aktivitet til denne og andre beslektede gener til tross for det overveldende beviset for mesolimbisk aktivering, spesielt i dopaminergiske veier og neuronale loki relatert til seksuell stimuli og aktivitet. Det er bemerkelsesverdig at Blum og Noble korrekt klassifiserte DRD2-genet som et generalisert belønningsgen som er ansvarlig for all belønningsmangel syndrom (RDS) atferd. Faktisk vil bruk av Bayesianske teoremanalysebærere av Taq Al-allelen i løpet av deres levetid ha 74% sjanse for at de vil møte med ett eller flere belønningsmangel syndrom (RDS) oppførsel [49].
Den første assosiasjonen av gen-polymorfisme og seksuell aktivitet forekom først før 1999 da Miller, et al. evaluert noen dopaminerge gener [50]. Det grunnleggende funnet er at det dopaminerge systemet i hjernen ser ut til å spille en viktig rolle i reguleringen av seksuell atferd. Forholdet mellom gener for Dl-, D2- og D4-dopaminreseptorene og alder ved første seksuell omgang (AFSI) ble undersøkt i en prøve av 414 ikke-spanske, europeisk-amerikanske menn og kvinner. En signifikant assosiasjon ble observert mellom et DRD2-allel og AFSI og en enda sterkere assosiasjon når DRD2-allelet ble interaktert med et DRDl-allel. En begrenset regresjonsmodell ble konstruert for å forutsi AFSI ved bruk av sex og en gruppe på ni psykososiale variabler som prediktorer. Å legge til DRD2 og DRD2-for-DRD1 prediktorer til denne modellen økte den forklarte variansen med henholdsvis 23% og 55%. At disse funnene tyder på en sterkere tilknytning blant menn enn blant kvinner, er i samsvar med andres nyere arbeid som viser høyere seksuell stimuli-respons hos menn enn hos kvinner. [51]. Så kanskje "menn er fra Mars og kvinner fra Venus", og dette kan til og med være sant for kokainmisbruk [52].
Spesielt har både prekliniske og kliniske studier vist seksuelt dimorfe mønstre i atferdsresponser mot kokain i alle faser av kokainavhengighetsprosessen (induksjon, vedlikehold og tilbakefall). Dermed dukker det opp et klart bilde som antyder at det er et biologisk grunnlag av kjønnsspesifikke forskjeller i kokainavhengighet. Disse forskjellene er resultatet av forskjellig regulering av CNS ved mannlige og kvinnelige gonadale hormoner og kan bli forutsagt av tilstedeværelsen av DRD2 genpolymorfismer [53]. Videre er det kjent at genetiske assosiasjoner mellom COMT og forskjellige psykiatriske fenotyper ofte viser forskjeller mellom menn og kvinner. Disse inkluderer den funksjonelle Val (158) Met polymorfisme i COMT som er assosiert med tvangslidelser hos menn og med angstfenotyper hos kvinner. I tillegg har Val (158) Met polymorfisme i COMT større innvirkning på kognitiv funksjon hos gutter enn jenter [54].
Miller, et al. fant ikke en assosiasjon av polymorfismer knyttet til DRD4-genet og alder for første samleie [50]. Andre fant imidlertid en betydelig tilknytning i visse etniske grupper. Spesielt indikerer deres analyse av polymorfismer i DRD4 at de med hvilken som helst - 3R genotype opplevde en risiko for første samleie høyere enn de med annen (eller hvilken som helst - 4R) genotype i alletnisiteter (n = 2,552). Interessant nok er ikke risikoen for første kjønn forskjellig mellom de to genotypene i det afroamerikanske utvalget, noe som reiser spørsmålet om kulturell oppdragelse [55].
Den seksuelle opplevelsen, som gjentatt medikamentbruk, gir langsiktige endringer, inkludert sensibilisering i nucleus accumbens (NAc) og ryggstriatum. Bradley et al. ved hjelp av mikroarray-analyse for å studere hamstere funnet for første gang at den seksuelle opplevelsen hos enten hann- eller hunndyr forskjellig opp eller ned regulerer mRNA-ekspresjon av en serie gener i NAc [56]. De fant at sammenlignet med seksuelt naive dyr, viste seksuelt erfarne hamstere som fikk en stimulushane på uke 7 en økning i et stort antall gener. Motsatt viste seksuelt erfarne kvinnelige hamstere som ikke fikk en stimulushane på uke 7, en reduksjon i uttrykket til mange gener. I følge forfatterne kan denne første genprofilen noensinne i kvinnelige hamstere gi et innblikk i mekanismene som både atferd (kjønn) og misbruksmedisiner induserer langsiktige endringer i de mesolimbiske og nigrostriatale dopaminveiene.
Bipolare elektroder, implantert bilateralt i lateral hypothalamus og substantia nigraventral tegmental område, ble stereotaksisk brukt for å gi kronisk selvstimulerende belønningsopplevelser som ligner på seksuell atferd. Denne typen stimulering har vist seg å indusere en betydelig økning i antall synapser i CA3-regionen i hippocampus og det molekylære laget av motorisk cortex hos rotter. I hovedsak induserte kronisk hjernestimulering langsiktig potensiering (LTP), som er kjent for å øke nye synaptiske forbindelser [57]. En enkelt eksponering for kokain hos naive dyr er tilstrekkelig til å utløse vedvarende forandringer i ventral tegmental area (VTA) glutamatergiske synapser som ligner aktivitetsavhengig LTP i andre hjerneregioner. Denne kokaininduserte LTP ser ut til å være mediert via dopamin D5-reseptoraktivering av N-metyl-D-aspartat (NMDA) reseptorer og å kreve proteinsyntese [58], støtter nok en gang vårt premiss her om at medisiner og sex kan ha vanlige nevrokjemiske underlag.
Empirisk forskning har avdekket et positivt forhold mellom antall sexpartnere og involvering i antisosial atferd [59]. De fleste forsøk på å forklare denne assosiasjonen har tatt et evolusjonært perspektiv. Fra evolusjonsperspektivet er de samme egenskapene, for eksempel impulsivitet, kortsiktighet og aggressivitet, som er relatert til et stort antall sexpartnere, også relatert til kriminelt engasjement. Det er imidlertid også grunn til å tro at samvariasjonen mellom sexpartnere og kriminalitetsatferd delvis kan forklares med en felles genetisk vei, der gener som er relatert til sexpartnerantall også er relatert til antisosial oppførsel. Ved å bruke den ovenfor beskrevne begrunnelsen, Beaver et al. fant en sterk positiv sammenheng mellom sexpartnere og antisosial atferd og polymorfisme av dopamintransportergenet (DAT1) forklarer variasjon i både antall seksuelle partnere og kriminell oppførsel for menn [59]. Den polymorfe effekten av DATl-genet og antall seksuelle partnere kan skyldes en assosiasjon funnet mellom visse polymorfismer og for tidlig mannlig utløsning av penis. Bærere av 1OR / 1OR-genotypen hadde score som indikerte en lavere terskel for å ejakulere på hver av indikatorene sammenlignet med den kombinerte 9R9R / 9R10R (9R høyere aktivitet lavere dopamin-tilgjengelighet) bærergruppe [60]. Polymorfismer av DATl-genet, spesifikt 10R / 10R-genotypen, er funnet hos ungdomskriminister som deltok på Brown School (San Marcos, Texas) for patologisk aggressiv atferd, inkludert anti-sosial atferd [61]. En positiv sammenheng mellom både DRD2 og DATl-polymorfismer ble observert med patologisk vold hos ungdommer i en blindet klinisk studie. Selv om den opprinnelig ble konseptualisert som følge av jevnaldrende imitasjon av ungdomsutbrudd eller livsløp-vedvarende ungdom, er det feste bevis fra tvillingstudier om at ungdomsstart eller ungdomsbegrenset antisosial atferd også kan være genetisk påvirket. Burt og Mikolajewski bekreftet ikke bare disse funnene med DATl-genet, men utvidet disse funnene til også å omfatte His452Tyr-varianten av genet som koder for 5-HT2A-reseptoren også [62]Mer nylig, Jozkow et al. rapporterte en sammenheng mellom den seksuelle dimensjonen av aldrende hannes symptomer (AMS) og genetiske varianter av 5-HTRlB G861C [63]. Dessuten har Sales et al. funnet gjennom multivariabel logistisk regresjonsanalyse, en interaksjon mellom misbruk og 5-HTTLPR-gruppen hvor det var en betydelig tilknytning til ikke-endringsstatus, sammen med partnerkommunikasjonsfrekvensscore ved oppfølging [64] Å ha en historie med misbruk var betydelig assosiert med større sjanser for ikke-endring i kondombruk etter intervensjon for bare de med s-allelen.
Det er kjent at polymorfismer i ikke-kodende regioner av vasopressin la reseptorgenet (Avpr la) er knyttet til sosio-emosjonelle egenskaper hos mennesker, sjimpanser og volder, og kan skyldes en stedsspesifikk variasjon i genuttrykk. I følge Barrett, et al., Tilbyr den sosialt monogame prærievollen en unik mulighet til å studere nevrobiologien til monogami [65]. Faktisk er vasopressin la reseptor (VlaR) signalering nødvendig for dannelsen av parbindingen hos menn. Interessant nok viser sosiale prairievolder større VlaR-binding i belønningsbehandlingen ventral pallidum enn asosiale volder av samme slekt. Barrett, et al. fant at nedregulering av pallidal VlaR-tetthet resulterte i en betydelig svekkelse av preferansen for en parret kvinnelig partner og en reduksjon i angstlignende oppførsel i voksen alder [65]. Annet arbeid av Garcia, et al. avslørte at individer med minst en 7-repeterende allel (7R +) av DRD4 rapporterer om en større kategorisk frekvens av promiskuøs seksuell oppførsel, inkludert å ha hatt en "one-night stand", og rapporterer en mer enn 50% økning i tilfeller av seksuell utroskap [66].
Det er viktig at Daw og Guo rapporterte at individer som bærer genotypene DRD2 * Al / A2, DRD2 * A2 / A2, DATl * 9R / 10R og MAOA * 2R / er assosiert med høyere odds for ubeskyttet seksuell omgang enn andre genotyper. [67]. DRD2-foreningene gjelder både menn og kvinner, mens de andre linkene bare gjelder kvinner. Til slutt, Emanuele, et al. rapporterte en betydelig sammenheng mellom DRD2 TaqI A-genotyper og “Eros · (en kjærlig stil preget av en tendens til å utvikle intense følelsesmessige opplevelser basert på den fysiske tiltrekningen til partneren), så vel som mellom C516T 5HT2A polymorfisme og“ mani ”( en besittende og avhengig romantisk tilknytning, preget av selvdestruktive følelser) [68].
Epigenetikk og seksuell aktivitet
En gjennomgang av litteraturen avslører at en rekke nylige artikler påpeker viktigheten av epigenetiske effekter på seksuell aktivitet. For eksempel gjennomgikk Matsuda de epigenetiske endringene av østrogenreseptoren a (ERalpha) og påvirkning på sosiosexual atferd [69]. Faktisk endrer endring av ER alfa-genaktivitet formidlet av epigenetiske mekanismer, som histonmodifikasjoner og DNA-metylering, ens seksuelle oppførsel. Når det gjelder homofili, har Rice et al. utviklet en modell som kan forklare kanalisering (konvertering) homofili seksuell utvikling [70]. De forklarer at denne modellen er basert på epigenetiske merker lagt ned som svar på XX vs. XY-karyotypen i embryonale stamceller. Følgelig øker disse merkene følsomheten for testosteron hos XY-fostre og senker den i XX-fostre, og kanaliserer den seksuelle utviklingen. Det er blitt postulert at en delmengde av disse konverterende epigenetiske merkene kvantitativt kan overføre transgenetisk, og kan føre til mosaikk for seksuell utvikling hos avstammende kjønn avkom - den homoseksuelle fenotypen.
I den sosialt monogame prærievollen (Microtus ochrogaster) induserer parring varige parbindinger som initieres av partner-preferansedannelse og reguleres av en rekke nevrotransmittere, inkludert oksytocin, vasopressin og dopamin. Arbeid av Gundersen [71], og Wang, et al. [72] antyder at histondeacetylase kan lette partnerdannelse i kvinnelige prærievolum som kan ha relevans for mennesker. Spesifikt Wang, et al. fant at histonedeacetylase-hemmere-natriumbutyrat og trikostatin A (TSA) forbedret partnepreferansedannelse i kvinnelige prærievolum [72]. Denne partnerpreferansedannelsen var assosiert med en oppregulering av oksytocinreseptor (OTR, oxtr) og vasopressin V la reseptor (VlaR, avprla) i NAc, gjennom en økning i histonacetylering hos deres respektive promotorer.
Det er interesse for økende bevis som indikerer at kvinner aktivt driver med polyandry enten for å unngå genetisk inkompatibilitet eller for å skjev farskap til fordel for genetisk overlegne menn. Det er muligheten for at valg av overlegen mannlig egnethet kan være på grunn av epigenetiske effekter. I følge Zeh og Zeh, i motsetning til DNA-sekvensbasert variasjon, kan epigenetisk variasjon bli sterkt påvirket av miljømessige og stokastiske effekter opplevd i løpet av et individ [73]. De antyder at epigenetisk variasjon kan være viktig for den seksuelle seleksjonen etter kopulatoriet og kan redegjøre for funn som knytter sædkonkurransevnen til avkomens kondisjon.
Genetisk og meme evolusjon: menneskelig formering
Eysenck foreslo en positiv sammenheng mellom ekstraversjon og intensivert seksuell atferd og mellom nevrotisme og problemer i seksuell atferd (antisosial atferd). En tidligere studie med gifte viste ingen av disse sammenhengene. Det ble antatt at denne forbindelsen bare eksisterer for ugifte personer som ikke er i langvarige forhold fordi kvaliteten på forholdet bestemmer den seksuelle interaksjonen. I et utvalg av unge ugifte menn var det en positiv sammenheng mellom ekstraversjon og gjenstander der personen beskrev tidligere seksuell aktivitet med flere individer og i høyere frekvens. Det ble ikke funnet noen sammenheng med nevrotisme. Det var også små sammenhenger med andre personlighets- og sosiale holdningsskalaer. På grunn av sammenhengen med en utøvende personlighetsskala ble funnene tolket fra et sosial-psykologisk perspektiv. I dagens samfunn forventes det at den unge hannen tar initiativ til en seksuell interaksjon som en ekstravert ung mann kan forstå bedre enn en som er introvert. [74]. Dette perspektivet er i direkte samsvar med Richard Brodies idé om egoistiske sinnsgener [75]. Fra DNA-synspunktet er selvfølgelig antropologer enige i at «vi er fortsatt her bare av en grunn; å gå ut og formere seg. ” Mens evolusjonære fremskritt er sakte, ett skritt hvert 20. år eller så, sammenlignet med "meme-evolusjon, muterer en idé i tiden det tar å lese en setning." Hjernen vår har ingenting med genetisk utvikling å gjøre, bortsett fra at den er relatert til smarte mennesker som har færre babyer. Faktisk, hvis det er gener som gir folk en tendens til å ta på seg memer som begrenser antall avkom, vil de dø ut i løpet av få generasjoner til fordel for gener som gir mennesker en tendens til å tilegne seg barn. Selv om det er noe kontroversielt, tyder dessverre en rekke studier på at Homo sapiens de siste 42,000 årene har senket IQ-ene på grunn av selektiv parring [76].
Ekstraordinært viser det seg at siden ekstraversjon er knyttet til økt seksuell aktivitet, spesielt hos menn, estimerer kvantitativ genetiker arveligheten til den utadvendte personligheten til å være rundt 40-60%. Smillie og medarbeidere studerte og fant at en kopi av DRD2-genet Al-allelen var assosiert med betydelig høyere ekstraversjon [77]. Denne foreningen reiser et interessant spørsmål når det gjelder menneskelig forplantning. Comings antydet at på grunn av deres markante effekt på reproduktiv atferd, læringsforstyrrelser og andre impulsive, kompulsive, aggressive og vanedannende lidelser, har bærere av DRD2 Al potensial til å forårsake progressive og permanente endringer i frekvensen av DRD2Al-allelet som fører til den genetiske smeltingen av arten ” [78].
I sin bok gir Comings bevis på at personer med vanedannende og forstyrrende atferd har barn tidligere, og dette påvirker utvalget av avhengighetsgener som DRD2 Al-allelen [79]. Han antyder at individer som bærer denne forstyrrende risikoallelen, vil ha barn, la oss si i 20-årsalderen, og individer uten denne allelen vil få barn på 25 år. Som et resultat vil det mutante genet reprodusere raskere, nemlig hvert 20. år, mens den normale formen for genet vil reprodusere hvert 25. år. Forholdet 25/20 er 1.25. Dermed vil hastigheten med hvilket et gen som har en 1.25 ganger selektiv fordel øke i frekvens fra generasjon til generasjon. En forskjell på fem år i alderen til mødre eller fedre når de får sine første barn, er tilstrekkelig til å resultere i et betydelig og relativt raskt utvalg for gener som bæres av en gruppe som initierer fødsel i en tidligere alder. Økninger i noen RDS-atferd er dokumentert fra 1955 til i dag. Disse økningene inkluderer atferdssyndrom (narkotika, kjønn, tenåringsgraviditet og forseelse, røyking), oppførselsforstyrrelse, kriminalitet, narkotikamisbruk, alkoholisme, ubeskyttet seksuell oppførsel, ugifte mødre, velferd, skolevist og skolefrafall, så vel som en samtidig reduksjon i IQ [80]. Disse resultatene er basert på Berkeley-studien som benytter langsgående data fra Barnehelse- og utviklingsstudiene og de nasjonale longitudinelle undersøkelsene av ungdom eller NLYS [81]. Ved å benytte seg av denne informasjonen spådde Comings at fra 1955 til 2015 vil det være en dobling av frekvensen av for eksempel DRD2 Al-allelen, og dermed øke utbredelsen av RDS-oppførsel, inkludert forhåndsdrevet samleie. [50]. Vi oppfordrer til en oppfølging av denne interessante spådommen.
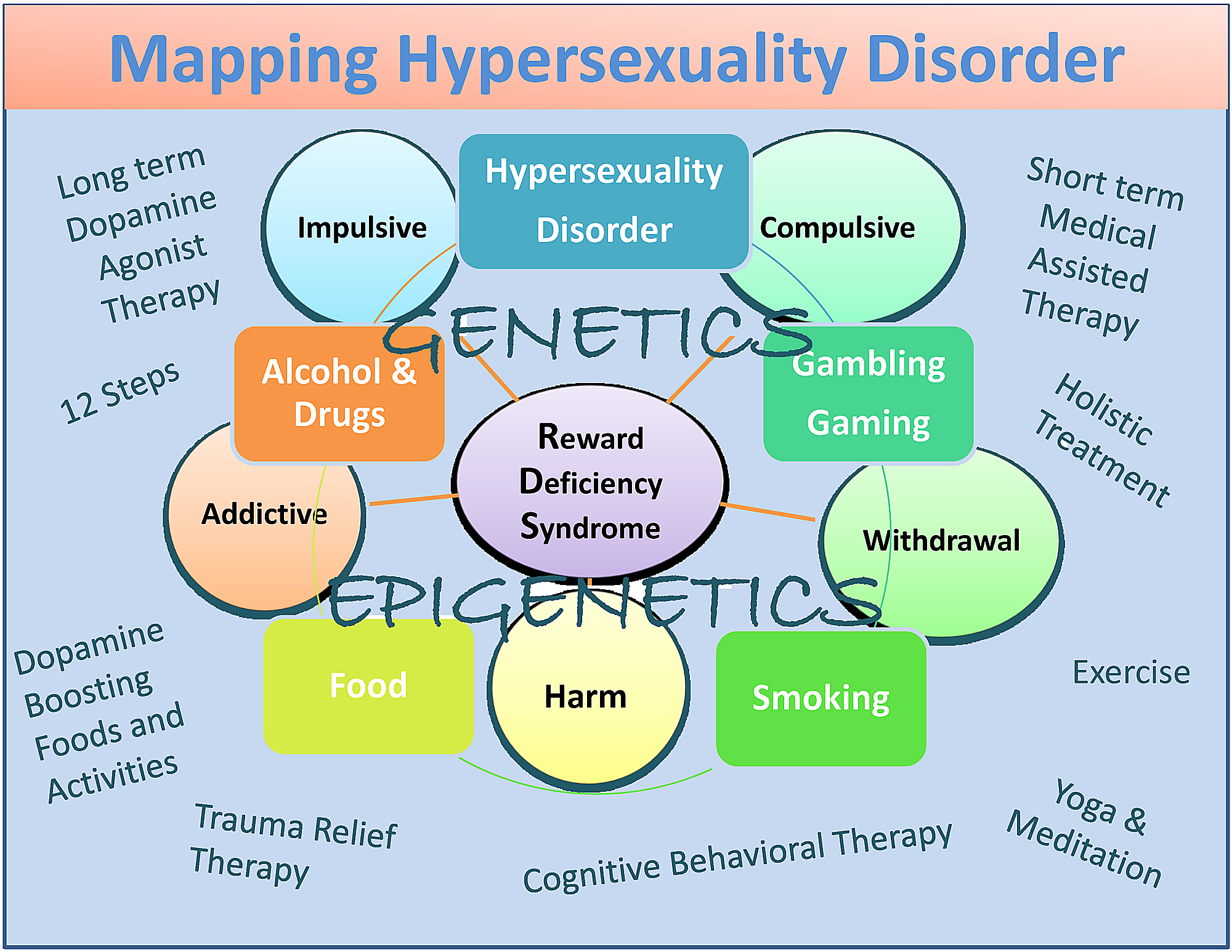
Til tross for en viss uenighet, foreslår vi hyperseksualitetsforstyrrelse som en subtype av RDS som deler kjennetegn med substans- og ikke-rusavhengighetsadferd, der dets kliniske uttrykk delvis påvirkes av både genetikk og epigenetikk. Selv om vi ikke er testet på dette tidspunktet, foreslår vi også kortvarige FDA-godkjente medisinerassistert behandling (MAT) som favoriserer blokkering av dopaminfunksjon etterfulgt av forsiktig aktivering av dopaminergiske veier som fører til dopaminhomeostase på lang sikt. Det siste kan oppnås ved noen modaliteter som kan hjelpe til med utvinning.
Om enn potensiell skjevhet inkluderer de dopaminagonistterapi-ernæringsmidler (KB220), 12 Step-programmet og tradisjonen, helhetlig behandling, kognitiv atferdsterapi (CBT) og traumelindringsterapi (TRT) samt dopamin-boostende aktiviteter og mat (figur) 1) [82].
Kontroversen
Selv om vi er overbevist om at hyperseksualitetsforstyrrelse bør inkluderes i fremtidige utgaver av DSM, er vi litt forvirrede at det er så lite kjent om denne lidelsen når det gjelder nevrogenetikk og epigenetikk og til og med abstinenssymtomatologi og generell fenomenologi [83]. Den viktigste hjemmeddelingen er at vi nå oppfordrer det vitenskapelige samfunnet til å utføre eksperimenter, spesielt innen neuroimaging og neurogenetics, inkludert epigenetika som er spesifikke for gener, for eksempel oxytocin-vasopressin-orexin-dopamin samt andre belønningsgener. Denne tilstanden kan muligens dra nytte av behandling som mål belønner genpolymorfismer for å hjelpe til med å fremme dopaminhomeostase [84-89]. En rekke anmeldelser av Joranby, et al. og Edge and Gold støtter vanlige behandlingsmuligheter relatert til delte nevrokjemiske mekanismer i hjernebelønningskretsløp slik det ble uttalt tidligere i RDS-konseptet [90-91].
Historisk var "sexavhengighet" inkludert i DSM - III, men den ble fjernet fra DSM-1V fordi konsensus fra forfatterne av DSM-1V mente at det ikke var tilstrekkelig bevis for å fortjene konklusjonen. Denne beslutningen var fylt med enorme følelser av ledere i feltet. Etter denne hendelsen bestemte en rekke forskere, inkludert Kafka, Reid, Prause og andre, å ramme “Hypersexuality” ikke som en sexavhengighet, men som en frittstående mental lidelse og ikke som en avhengighet per se. Mens deres tidligere arbeid i 2010 antydet som referert til her at "hyperseksualitet" var lik sexavhengighet og muligens annet avhengighet, inkludert rusmisbruk, støtter deres nyere arbeid seg fra denne påstanden. Nyere arbeid på dette området avslører fortsatt kontrovers. Det er en rekke elektrofysiologiske studier fra Prauses gruppe som gir bevis for at seksuell lyst, ikke hyperseksualitet, forutsier selvregulering av seksuell opphisselse [92]. Disse etterforskerne antyder i annet arbeid at personer som rapporterte problemer som regulerer deres syn på visuell seksuell stimuli (VSS), som også rapporterte høyere seksuell lyst, viste lavere sent-positive potensialer (LPP) som svar på VSS. Forfatterne foreslår at dette mønsteret fremstår annerledes enn stoffavhengighetsmodeller [93]. Selv om de ikke involverte personer med hyperseksuell lidelse, har arbeid fra Voon's gruppe imidlertid vist at hos tvangsmessige-seksuelle atferd, utsatt for seksuelt eksplisitte videoer, en større aktivitet i det nevrale nettverket ligner det som ble observert i studier om medikamentell reaksjon. [94]. Større lyst eller ønsker fremfor å like var videre assosiert med aktivitet i dette nevrale nettverket. Dette arbeidet samsvarer med teorier om insentivmotivasjon [95].
Vi, forfatterne av den nåværende artikkelen, innrømmer at vi ikke har vært fortrolige med alle de viktige interaksjonene som har skjedd mellom forkjemperne for "Hypersexuality Disorder" og deres ærlige hensikt å ha denne lidelsen inkludert i dagens DSM-5. Selv om den mislyktes i den såkalte "acid-test", er det all grunn til å tro at den vil bli inkludert i fremtidige utgaver av DSM. Det er bemerkelsesverdig at Steven Hyman, den nåværende NIH-direktøren, riktig hevdet at “DSM er et dårlig speil av kliniske og biologiske virkeligheter; en grunnleggende ny tilnærming til diagnostisk klassifisering er nødvendig når forskere avdekker nye måter å studere og forstå psykiske lidelser på. [96]. Dessuten har Casey, et al. foreslått at selv om DSM anser forskjellige lidelser som forskjellige enheter, er "grensene mellom lidelsene ofte ikke så strenge som DSM antyder" [97].
I 2014 har Karila, et al. antydet at seksuell avhengighet, også kjent som hyperseksuell lidelse, har blitt ignorert av noen psykiatere, til tross for at tilstanden forårsaker alvorlige psykososiale problemer. I følge disse forfatterne foreslår de at seksuell avhengighet eller hyperseksuell lidelse representerer forskjellige vilkår for det samme problemet. De peker på at forekomsten av seksuelle avhengighetsrelaterte lidelser varierer fra 3% til 6%. Følgelig viser konstruksjonen av seksuell avhengighet / hyperseksuell lidelse problematisk atferd inkludert: overdreven onani, cybersex, pornografibruk, avvikende seksuell atferd med samtykkende voksne, telefon sex, besøk i stripklubber og annen vanedannende atferd [98]. Vi er helt sikkert enige om at det kan være tydelige forskjeller mellom seksuell avhengighet og hyperseksualitet som bemerket av Carvalho, et al. [99], Rettenberger, et al. [100], Kor, et al. [1], Reid, et al. [9], Kafka og Hennen [13], og Prause, et al. [93-94] blant andre.
Oppsummert har vi foreslått at selv om det er noen forskjeller mellom hyperseksualitet og sexavhengighet, er det nødvendig med mer forskning for å kategorisere disse svært viktige forholdene. Vi er enige i arbeidet til Walters, et al. [101] som antydet at individuelle forskjeller i hyperseksualitet er kvantitative snarere enn kvalitative. De antyder også at hyperseksualitet er organisert langs et kontinuum som faller i øvre ende av kontinuumet (Figur 1).

Figur 1: Et beskrivende kart over hyperseksualitetsforstyrrelse som en subtype av RDS
Figuren illustrerer interaktive neurogenetiske og epigenetiske effekter. Både kortsiktige dopaminblokkerende og langsiktige “dopaminerge-homeostase” -baserte behandlinger og dopaminfremmende terapier og daglige aktiviteter er oppført. Sirkler angir RDS-egenskaper og ruter angir RDS-atferd.
{kind=link}
Konklusjoner
Mens vi anerkjenner kontroversen, foreslår vi at mulige forskjeller og likheter mellom hyperseksualitetsforstyrrelse og sexavhengighet bør undersøkes tilstrekkelig ved hjelp av nevroavbildning (fMRI, PET, SPECT), optogenetikk, kandidat- og mikroarrayanalyse og epigenetiske teknikker. Vi tror at disse undersøkelsene vil gi grunnlag for å inkludere hyperseksualitet som en lidelse i fremtidige utgaver av DSM.
Referanser
- Kor A, Fogel Y, Reid RC, Potenza MN: Bør hyperseksuell lidelse klassifiseres som en avhengighet?. Sex Addict Compulsivity. 2013, 20:http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3836191/. 10.1080/10720162.2013.768132
- Rush B: Medisinske henvendelser og observasjoner ved sykdom i sinnet. Rush B (red): Gryphon Editions Ltd., Birmingham, AL; (Originalverk publisert 1812) gjennomgått i 1979.
- von Krafft-Ebing R: Psychopathia Sexualis. Klaf FS (red): Stein og Day, New York; (originalt arbeid utgitt 1886) gjennomgått i 1965.
- Hirshfeld M: Seksuelle anomalier: Opprinnelse, art og behandling av seksuelle lidelser. Hirshfeld M (red): Emerson Books, New York; 1948.
- Barnevogn RJ: Perversjon: Den erotiske formen for hat. Pantheon Books, New York; 1975.
- Allen CA: En lærebok om psykoseksuelle lidelser. Oxford University Press, London; 1962.
- Ellis A, Sagarin E: Nymfomani: En studie av den overexse kvinnen. Gilbert Press, New York; 1964.
- Kafka MP: Hva skjedde med hyperseksuell lidelse?. Arch Sex Behav. 2014, 43: 1259-1261. 10.1007 / s10508-014-0326-y
- Reid RC, Temko J, Moghaddam JF, Fong TW: Skam, drøvtyggelse og selvfølelse hos menn vurdert for hyperseksuell lidelse. J Psychiatr Practice. 2014, 20: 260 – 268. 10.1097 / 01.pra.0000452562.98286.c5
- Bancroft J: Menneskelig seksualitet og dens problemer. Tredje utgave. Elsevier, Oxford, England; 2009.
- Internasjonal statistisk klassifisering av sykdommer og relaterte helseproblemer, 10th revisjon . (2007). Tilgang: juli 23, 2015: http://apps.who.int/classifications/apps/icd/icd10online2007/.
- Singer B, Toates FM: Seksuell motivasjon. J Sexforskning. 1987, 23: 481 – 501. 10.1080/00224498709551386
- Kafka MP, Hennen J: Hyperseksuell lyst hos menn: er menn med parafili forskjellig fra hanner med parafili-relaterte lidelser?. Sexmisbruk. 2003, 15: 307 – 321. 10.1023 / A: 1025000227956
- Buss DM, Schmitt DP: Teorier om seksuelle strategier: et evolusjonært perspektiv på menneskelig parring. Psychol Rev. 1993, 100: 204 – 232. 10.1037 / 0033-295X.100.2.204
- Corbett-Detig RB, Hartl DL, Sackton TB: Naturlig utvalg begrenser nøytralt mangfold over en lang rekke arter. Psychol Bull. 1995, 13: e1002112.- 469-496. doi: 10.1371 / journal.pbio.1002112
- Laumann EO, Michael RT, Gagnon JH: En politisk historie om den nasjonale sexundersøkelsen av voksne. Fam Plann Perspect. 1994, 26: 34 – 38. 10.2307/2136095
- Jones JC, Barlow DH: Selvrapportert hyppighet av seksuelle trang, fantasier og onanerende fantasier hos heterofile menn og kvinner. Arch Sex Behav. 1990, 19: 269 – 79. 10.1007 / BF01541552
- Oliver MB, Hyde JS: Kjønnsforskjeller i seksualitet: en metaanalyse. Psychol Bull. 1993, 114: 29 – 51. 10.1037 / 0033-2909.114.1.29
- Okami P, Shackelford TK: Menneskelige kjønnsforskjeller i seksuell psykologi og atferd. Annu Rev Sex Res. 2001, 12: 186–241. 10.1080/10532528.2001.10559798
- Leitenberg H, Detzer MJ, Srebnik D: Kjønnsforskjeller i onani og forholdet til onaniopplevelse i preadolescens og / eller tidlig ungdom til seksuell atferd og seksuell tilpasning i ung voksen alder. Arch Sex Behav. 1993, 22: 87 – 98. 10.1007 / BF01542359
- Fetterman AK, Kruger NN, Robinson MD: Kjønnsmessige parringsstrategier avviker med en manipulering av kjønnssaliitet. Motiv Emot. 2015, 39: 99 – 103. 10.1007/s11031-014-9420-7
- Basson R: Ved å bruke en annen modell for kvinnelig seksuell respons for å adressere kvinners problematiske lave seksuelle lyst. J Sex Marital Ther. 2001, 27: 395–403. 10.1080/713846827
- Andersen BL, Cyranowski JM, Aarestad S: Utover kunstige, kjønnsbundne distinksjoner for å konseptualisere kvinnelig seksualitet: kommenter Baumeister. Psychol Bull. 2000, 126: 380 – 389. 10.1037 // 0033-2909.126.3.380
- Knight RA, Sims-Knight JE: De utviklingsmessige forfølgene ved seksuell tvang mot kvinner: testing av alternative hypoteser med strukturell ligningsmodellering. Ann NY Acad Sci. 2003, 989: 72 – 85. 10.1111 / j.1749-6632.2003.tb07294.x
- Blum K, Werner T, Carnes S, Carnes P, Bowirrat A, Giordano J, Oscar-Berman M, Gold M: Sex, narkotika og rock 'n' roll: hypoteser om vanlig mesolimbisk aktivering som en funksjon av belønningsgenpolymorfier. J Psykoaktive medikamenter. 2012, 44: 38 – 55. 10.1080/02791072.2012.662112
- Carnes PJ, Green BA, Merlo LJ, Polles A, Carnes S, Gold MS: PATHOS: en kort screeningapplikasjon for vurdering av seksuell avhengighet. J Addict Med. 2012, 6: 29 – 34. 10.1097/ADM.0b013e3182251a28
- Hanson RK: Forutsier Statisk-99 tilbakefall blant eldre seksuelle krenkere?. Sexmisbruk. 2006, 18: 343–355. 10.1007 / s11194-006-9027-y
- Weinstein A, Katz L, Eberhardt H, Cohen K, Lejoyeux M: Seksuell tvang - Forhold til sex, tilknytning og seksuell legning. J Behav Addict. 2015, 4: 22 – 26. 10.1556 / JBA.4.2015.1.6
- Chung WS, Lim SM, Yoo JH, Yoon H: Kjønnsforskjell i hjerneaktivering til audiovisuell seksuell stimulering; opplever kvinner og menn det samme opphisselsesnivået som svar på det samme videoklippet?. Int J Impot Res. 2013, 25: 138-142. 10.1038 / ijir.2012.47
- Rupp HA, Wallen K: Kjønnsforskjeller som svar på visuell seksuell stimuli: en anmeldelse. Arch Sex Behav. 2008, 37: 206 – 218. 10.1007/s10508-007-9217-9
- Hanson RK, Harris AJ, Helmus L, Thornton D: Fornærmede sexforbrytere med høy risiko er kanskje ikke høyrisiko for alltid. J Interpers Vold. 2014, 29: 2792-813. 10.1177/0886260514526062
- Reid RC, Cyders MA, Moghaddam JF Fong TW: Psykometriske egenskaper ved Barratt Impulsivity Scale hos pasienter med spillelidelser, hyperseksualitet og metamfetaminavhengighet. Addict Behav. 2014, 39: 1640 – 1645. 10.1016 / j.addbeh.2013.11.008
- Reid RC, Carpenter BN, Hook JN, Garos S, Manning JC, Gilliland R, Cooper EB, McKittrick H, Davtian M, Fong T: Rapport om funn i en DSM-5 feltstudie for hyperseksuell lidelse. J Sex Med. 2012, 9: 2868 – 2877. 10.1111 / j.1743-6109.2012.02936.x
- Garcia FD, Thibaut F: Seksuell avhengighet. Am J Narkotikamisbruk. 2010, 36: 254-260. 10.3109/00952990.2010.503823
- Reid RC, Bramen JE, Anderson A, Cohen MS: Mindfulness, emosjonell dysregulering, impulsivitet og stressfølelse blant hyperseksuelle pasienter. J Clin Psychol. 2014, 70: 313 – 321. 10.1002 / jclp.22027
- Innehaver MK, Hadjimarkou MM, Zup SL, Blutstein T, Benham RS, McCarthy MM, Mong JA: Metamfetamin letter seksuell atferd hos kvinner og forbedrer neuronal aktivering i den mediale amygdala og ventromedialkjernen i hypothalamus. Psychoneuroendocrinology. 2010, 35: 197 – 208. 10.1016 / j.psyneuen.2009.06.005
- Spauwen LW, Niekamp AM, Hoebe CJ, Dukers-Muijrers NH: Medikamentbruk, seksuell risikoatferd og seksuelt overførbare infeksjoner blant swingers: en tverrsnittsstudie i Nederland. Sextransmisjon infisere. 2015, 91: 31 – 36. 10.1136 / sextrans-2014-051626
- Castelo-Branco C, Parera N, Mendoza N, Pérez-Campos E, Lete I, CEA-gruppe: Alkoholmisbruk og risikofylt seksuell atferd hos unge voksne kvinner. Gynekol Endokrinol. 2014, 30: 581 – 586. 10.3109/09513590.2014.910190
- Jia ZJ, Yan SY, Bao YP, Lian Z, Zhang HR, Liu ZM: Seksuelle atferdsforskjeller mellom brukere av stimulerende stoffer av amfetamin og heroinbrukere. J Addict Med. 2013, 7: 422 – 427. 10.1097/ADM.0b013e3182a952b2
- Bruijnzeel AW: kappa-opioid reseptorsignalering og hjernebelønningsfunksjon. Brain Res Rev. 2009, 62: 127 – 146. 10.1016 / j.brainresrev.2009.09.008
- Orsini CA, Ginton G, Shimp KG, Avena NM, Gold MS, Setlow B: Matforbruk og vektøkning etter opphør av kronisk amfetaminadministrasjon. Appetitt. 2014, 78: 76 – 80. 10.1016 / j.appet.2014.03.013
- Ibrahim C, Reynaert C: Hyperseksualitet ved nevrokognitive lidelser hos eldre mennesker - en omfattende gjennomgang av litteraturen og casestudien. Psykiater Danub. 2014, 26: 36 – 40.
- Vallejo-Medina P, Sierra JC: Effekt av medikamentbruk og påvirkning av avholdenhet på seksuell fungering i en spansk mannlig medikamentavhengig prøve: en flerstedsstudie. J Sex Med. 2013, 10: 333 – 341. 10.1111 / j.1743-6109.2012.02977.x
- Baskerville TA, Douglas AJ: Dopamin og oksytocin-interaksjoner som ligger til grunn for atferd: potensielle bidrag til atferdsforstyrrelser. CNS Neurosci Ther. 2010, 16: e92-123. 10.1111 / j.1755-5949.2010.00154.x
- Riebe CJ, Lee TT, Hill MN, Gorzalka BBB: Presipitert abstinens teller de negative virkningene av subkronisk cannabinoid-administrasjon på seksuell atferd hos hannrotter. Neurosci Lett. 2010, 472: 171 – 174. 10.1016 / j.neulet.2010.01.079
- Korda JB, Pfaus JG, Kellner CH, Goldstein I: Vedvarende kjønnsopphisselsessykdom (PGAD): caserapport om langvarig symptomatisk behandling med elektrokonvulsiv terapi. J Sex Med. 2009, 6: 2901 – 2909. 10.1111 / j.1743-6109.2009.01421.x
- Huang CJ, Liao HT, Yeh GC, Hung KL: Distribusjon av HLA-DQB1-alleler hos pasienter med Kleine-Levin-syndrom. J Clin Neurosci. 2012, 19: 628 – 630. 10.1016 / j.jocn.2011.08.020
- Blum K, Noble EP, Sheridan PJ, Montgomery A, Ritchie T, Jagadeeswaran P, Nogami H, Briggs AH, Cohn JB: Allelisk forening av humant dopamin D2-reseptorgen i alkoholisme. JAMA. 1990, 263: 2055 – 2060. 10.1001 / jama.263.15.2055
- Blum K, Sheridan PJ, Wood RC, Braverman ER, Chen TJ, Cull JG, Comings DE: D2 dopaminreseptorgenet som en determinant av belønningsbristsyndrom. JR Soc Med. 1996, 89: 396-400. 10.1177/014107689608900711
- Miller WB, Pasta DJ, MacMurray J, Chiu C, Wu H, Comings DE: Dopaminreseptorgener er assosiert med alder ved første samleie. J Biosoc Sci. 1999, 31: 43 – 54. 10.1017 / S0021932099000437
- Hamann S, Herman RA, Nolan CL, Wallen K: Menn og kvinner er forskjellige i amygdala-respons på visuelle seksuelle stimuli. Nat Neurosci. 2004, 7: 411 – 416. 10.1038 / nn1208
- Quiñones-Jenab V: Hvorfor er kvinner fra Venus og menn fra Mars når de misbruker kokain?. Brain Res. 2006, 1126: 200 – 203. 10.1016 / j.brainres.2006.08.109
- Noble EP, Blum K, Khalsa ME, Ritchie T, Montgomery, A Wood RC, Fitch RJ, Ozkaragoz T, Sheridan PJ, Anglin MD, Paredes A, Treiman LJ, Sparkes RS: Allelisk assosiasjon av D2-dopaminreseptorgenet med kokainavhengighet. Narkotika-alkoholavhengighet. 1993, 33: 271 – 285. 10.1016/0376-8716(93)90113-5
- Harrison PJ, Tunbridge EM: Catechol-O-methyltransferase (COMT): et gen som bidrar til kjønnsforskjeller i hjernens funksjon, og til seksuell dimorfisme i predisposisjon for psykiatriske lidelser. Neuropsychopharmacology. 2008, 33: 3037 – 3045. 10.1038 / sj.npp.1301543
- Guo G, Tong Y: Alder ved første samleie, gener og sosial kontekst: bevis fra tvillinger og Dopamin D4 reseptorgen. Demografi. 2006, 43: 747 – 769. 10.1353 / dem.2006.0029
- Bradley KC, Boulware MB, Jiang H, Doerge RW, Meisel RL, Mermelstein PG: Endringer i genuttrykk i nucleus accumbens og striatum etter seksuell erfaring. Gener Brain Behav. 2005, 4: 31 – 44. 10.1111 / j.1601-183X.2004.00093.x
- Rao BS, Raju TR, Meti BL: Økt numerisk tetthet av synapser i CA3-regionen i hippocampus og molekylært lag av motorisk cortex etter selvstimulerende givende opplevelse. Neuroscience. 1999, 91: 799 – 803. 10.1016/S0306-4522(99)00083-4
- Heshmati M: Kokainindusert LTP i det ventrale tegmentale området: ny innsikt i mekanisme og tidsforløp belyser cellens underlag. J Neurophysiol. 2009, 101: 2735 – 2737. 10.1152 / jn.00127.2009
- Beaver KM, Wright JP, Walsh A: En genbasert evolusjonær forklaring på sammenhengen mellom kriminelt engasjement og antall sexpartnere. Biodemografi Soc Biol. 2008, 54: 47 – 55. 10.1080/19485565.2008.9989131
- Santtila P, Jern P, Westberg L, Walum H, Pedersen CT, Eriksson E, Sandnabba N: Dopamintransportørgenet (DAT1) polymorfisme er assosiert med for tidlig utløsning. J Sex Med. 2010, 7: 1538 – 1546. 10.1111 / j.1743-6109.2009.01696.x
- Chen TJH, Blum K, Mathews D, Fisher L, Schnautz N, Braverman Er, Schoolfield J, Downs W, Blum SH, Mengucci J, Meshkin B, Arcuri V, Bajaj A, Waite RL, Comings DE: Foreløpig assosiasjon av både Dopamine D2-reseptoren (DRD2) [Taq1 A1 Allele] og Dopamine Transporter (DAT1) [480 bp Allele] -gener med patologisk aggressiv atferd, en klinisk subtype av belønningssviktssyndrom (RDS) hos ungdom.. Gene Ther Mol Biol. 2007, 1: 93-112. Tilgang: juli 23, 2015: http://gtmb.org/pages/Vol11A/HTML/11._Chen_et_al,_93-102.htm.
- Burt SA, Mikolajewski AJ: Foreløpig bevis for at spesifikke kandidatgener er assosiert med antisosial atferd hos ungdom. Aggress Behav. 2008, 34: 437 – 445. 10.1002 / ab.20251
- Jóźków P, Słowińska-Lisowska M, Łaczmański Ł, Mędraś M: Polymorfe varianter av nevrotransmitterreseptorgener kan påvirke seksuell funksjon hos aldrende menn: data fra HALS-studien. Neuroendocrinology. 2013, 98: 51 – 59. 10.1159/000350324
- Salg JM, DiClemente RJ, Brody GH, Philibert RA, Rose E: Interaksjon mellom 5-HTTLPR polymorfisme og misbrukshistorie på ungdoms afroamerikanske kvinnes atferd for bruk av kondom etter deltakelse i en HIV-forebyggende intervensjon. Prev Sci. 2014, 15: 257 – 267. 10.1007/s11121-013-0378-6
- Barrett CE, Keebaugh AC, Ahern TH, Bass CE, Terwilliger EF, Young LJ: Variasjon i vasopressinreseptor (Avpr1a) uttrykk skaper mangfoldighet i atferd relatert til monogami i prærievolum. Horm Behav. 2013, 63: 518 – 526. 10.1016 / j.yhbeh.2013.01.005
- Garcia JR, MacKillop J, Aller EL, Merriwether AM, Wilson DS, Lum JK: Forbindelser mellom dopamin D4 reseptorgenvariasjon med både utroskap og seksuell promiskuitet. PLoS One. 2010, 5: e14162. 10.1371 / journal.pone.0014162
- Daw J, Guo G: Påvirkningen av tre gener på om ungdommer bruker prevensjon, USA 1994-2002. Popul Stud (Camb). 2011, 65: 253 – 271. 10.1080/00324728.2011.598942
- Emanuele E, Brondino N, Pesenti S, Re S, Geroldi D: Genetisk belastning på menneskelige kjærlige stiler. Neuro Endocrinol Lett. 2007, 28: 815 – 821.
- Matsuda KI: Epigenetiske endringer i østrogenreseptor α-genpromotor: implikasjoner i sosiosexual atferd. Neurosci foran. 2014, 8: 344. 10.3389 / fnins.2014.00344
- Rice WR, Friberg U, Gavrilets S: Homoseksualitet via kanalisert seksuell utvikling: en testprotokoll for en ny epigenetisk modell. Bioessays. 2013, 35: 764 – 770. 10.1002 / bies.201300033
- Gundersen B: Parbinding gjennom epigenetikk. Nat Neurosci. 2013, 16: 779. 10.1038 / nn0713-779
- Wang H, Duclot F, Liu Y, Wang Z, Kabbaj M: Histon deacetylase-hemmere letter partnerpreferansedannelse i kvinnelige prærievolum. Nat Neurosci. 2013, 16: 919 – 924. 10.1038 / nn.3420
- Zeh JA, Zeh DW: Mors arv, epigenetikk og evolusjonen av polyandri. Genetica. 2008, 134: 45 – 54. 10.1007 / s10709-007-9192-z
- Addad M, Lesiau A: Ekstraversjon, nevrotisisme, umoralsk dømmekraft og kriminell oppførsel. Med Law. 1989, 8: 611 – 622.
- Brodie R: Virus of the Mind: The New Science of the Meme. Hay House, Inc, New York, NY; 1996, s. 66.
- Hernnstein R, Murray C: Klokkekurven: intelligens og klassestruktur i amerikansk liv. The Free Press, New York, NY; 1994.
- Smillie LD, Cooper AJ, Proitsi P, Powell JF, Pickering AD: Variasjon i DRD2 dopamingen forutsier ekstravert personlighet. Neurosci Lett. 2010, 468: 234 – 327. 10.1016 / j.neulet.2009.10.095
- Kommer DE: Genebomben. Akselererer høyere utdanning og avansert teknologi valg av gener for læringsforstyrrelser, ADHD, vanedannende og forstyrrende atferd?. Hope Press, Duarte CA; 1996.
- Kommer DE: Problemet for adferdssyndrom. Genebomben. Akselererer høyere utdanning og avansert teknologi valg av gener for læringsforstyrrelser, ADHD, vanedannende og forstyrrende atferd? Hope Press, Duarte CA; 1996. s. 91-94.
- Kommer DE: Genvalg. Genebomben. Akselererer høyere utdanning og avansert teknologi valg av gener for læringsforstyrrelser, ADHD, vanedannende og forstyrrende atferd? Hope Press, Duarte CA; 1996. s. 89-90.
- Kommer DE: The Berkeley Study. Genebomben. Akselererer høyere utdanning og avansert teknologi valg av gener for læringsforstyrrelser, ADHD, vanedannende og forstyrrende atferd? Hope Press, Duarte California; 1996. s. 105 – 210.
- Borsten J: Malibu Beach Recovery Diet Cookbook. Vidov Publishing Inc, Malibu, CA; 2015.
- Derbyshire KL, Grant JE: Tvangsmessig seksuell atferd: En gjennomgang av litteraturen. J Behav Addict. 2015, 4: 37 – 43. 10.1556/2006.4.2015.003
- Nirenberg MJ: Dopamin agonist abstinenssyndrom: implikasjoner for pasientbehandling. Legemidler aldring. 2013, 30: 587 – 592. 10.1007 / s40266-013-0090-z
- Grant JE, Brewer JA, Potenza MN: Neurobiologi av substans og atferdsmessig avhengighet. CNS Spectr. 2006, 11: 924 – 930.
- Sakata K, Duke SM: Mangel på BDNF-ekspresjon gjennom promotor IV forstyrrer ekspresjon av monoamingener i frontal cortex og hippocampus. Neuroscience. 2014, 260: 265 – 75. 10.1016 / j.neuroscience.2013.12.013
- Blum K, Liu Y, Wang W, Wang Y, Zhang Y, Oscar-Berman M, Smolen A, Febo M, Han D, Simpatico T, Cronjé FJ, Demetrovics Z, Gold MS: rsfMRI-effekter av KB220Z ™ på nevrale traséer i belønningskretsløp for avholdende genotypede heroinavhengige. Postgrad Med. 2015, 127: 232 – 241.
- McLaughlin T, Blum K, Oscar-Berman M, Febo M, Demetrovics Z, Agan G, Fratantonio J, Gold MS: Bruke Neuroadaptagen KB200z ™ for å forbedre skremmende, klarsomme mareritt hos RDS-pasienter: rollen som forbedret, hjernebelønning, funksjonell tilkobling og dopaminerg homeostase. J Reward Defic Syndr. 2015, 1: 24-35. 10.17756 / jrds.2015-006
- Blum K, Thanos PK, Badgaiyan RD, Febo M, Oscar-Berman M, Fratantonio J, Demotrovics Z, Gold MS: Neurogenetikk og genterapi for belønningsmangel syndrom: skal vi til det lovede land?. Ekspert Opin Biol Ther. 2015, 5: 973 – 985. 10.1517/14712598.2015.1045871
- Joranby L, Pineda-Frost KY, Gold MS: Avhengighet til belønningssystemer for mat og hjerne. Seksuell avhengighet og kompulsivitet. 2005, 12: 201–217. 10.1080/10720160500203765
- Edge PJ, Gold MS: Medikamentuttak og hyperfagi: leksjoner fra tobakk og andre medikamenter. Curr Pharm Des. 2011, 17: 1173 – 1179. 10.2174/138161211795656738
- Moholy M, Prause N, Proudfit, GH, Rahman A, Fong T: Seksuell lyst, ikke hypersexualitet, forutsier selvregulering av seksuell opphisselse. Cogn Emot. 2015, 6: 1012.
- Prause N, Steele, VR, Staley C, Sabatinelli, D, Hajcak G: Modulering av sent positive potensialer ved seksuelle bilder i problembrukerne og kontrollerer inkonsekvent med "pornoavhengighet". Biol Psychol. 2015, 109: 192-199. 10.1016 / j.biopsycho.2015.06.005
- Mechelmans DJ, Irvine M, Banca P, et al: Forbedret oppmerksomhet mot seksuelt eksplisitte tegn på individer med og uten tvangsmessig seksuell adferd. PLoS One. 2014, 25, 9 (8): e105476. 10.1371 / journal.pone.0105476
- Blum K, Gardner E, Oscar-Berman M, Gold M: "Liking" og "wanting" knyttet til Reward Deficiency Syndrome (RDS): hypoteser om differensiell respons i hjernebelønningskretser. Curr Pharm Des. 2012, 18 (1): 113-118.
- Hyman SE: Diagnostisering av DSM: Diagnostisk klassifisering trenger grunnleggende reform. Cerebrum. 2011, 2011: 6. Tilgang: 2011 april 26: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3574782/.
- Casey BJ, Craddock N, Cuthbert BN, Hyman SE, Lee FS, Ressler KJ: DSM-5 og RDoC: fremgang i psykiatrisk forskning?. Nat Rev Neurosci. 2013, 14: 810-14. 10.1038 / nrn3621
- Karila L, Wéry A, Weinstein A, Cottencin O, Petit A, Reynaud M, Billieux J: Seksuell avhengighet eller hyperseksuell lidelse: forskjellige vilkår for samme problem? En gjennomgang av litteraturen. Curr Pharm Des. 2014, 20: 4012-20. 10.2174/13816128113199990619
- Carvalho J, Štulhofer A, Vieira AL, Jurin T: Hyperseksualitet og høy seksuell lyst: å utforske strukturen til problematisk seksualitet. J Sex Med. 2015, 12: 1356-67. Tilgang: 2015 Mar 23: http://onlinelibrary.wiley.com/doi/10.1111/jsm.12865/abstract;jsessionid=6F416CCBB66B7F0EA28E428D4993EBD5.f01t04. 10.1111 / jsm.12865
- Rettenberger M, Klein V, Briken P: Forholdet mellom hyperseksuell atferd, seksuell eksitasjon, seksuell hemning og personlighetstrekk. Arch Sex Behav. 2015, Jan 6: (Epub foran trykk). Tilgang: 2015 Jan 6: 10.1007/s10508-014-0399-7
- Walters GD, Knight RA, Långström N: Er hyperseksualitet dimensjonalt? Bevis for DSM-5 fra generell populasjon og kliniske prøver. Arch Sex Behav. 2011, 40: 1309-21. Tilgang: 2011 februar 3: 10.1007/s10508-010-9719-8