KOMENTÁRE: recenzia popredného výskumného pracovníka na tému obezity a závislosti na potravinách.
Zväzok 69, vydanie 4, 24 február 2011, stránky 664 – 679
http://dx.doi.org/10.1016/j.neuron.2011.02.016,
preskúmanie
Paul J. Kenny1, ,
1 Laboratórium správania a molekulárnej neurológie, Katedra molekulárnych terapeutík, Výskumný ústav Scripps, Jupiter, FL 33458, USA
________________________________________
Potraviny sa spotrebúvajú na udržanie energetickej rovnováhy na úrovni homeostatiky. Navyše chutné potraviny sa tiež spotrebúvajú za svoje hedonické vlastnosti nezávislé od energetického stavu. Takáto odmena súvisiaca s odmenou môže vyústiť do prekročenia požiadaviek na kalorický príjem a považuje sa za hlavného vinníka rýchlo sa zvyšujúcej miery obezity v rozvinutých krajinách. V porovnaní s homeostatickými mechanizmami kŕmenia je oveľa menej známe, ako hedonické systémy v mozgu ovplyvňujú príjem potravy. Prekvapujúco môže nadmerná konzumácia chutných potravín spúšťať neuroadaptivne odpovede v obvodoch odmeňovania mozgu podobne ako drogy zneužívania. Okrem toho podobné genetické zraniteľnosti v systéme odmeňovania mozgu môžu zvýšiť predispozíciu k drogovej závislosti a obezite. Tu sa preskúma nedávne pokroky v našom chápaní obvodov mozgu, ktoré regulujú hedonické aspekty kŕmneho správania. Takisto sa uvažuje o objavujúcich sa dôkazoch naznačujúcich, že obezita a drogová závislosť môžu mať spoločné hedonické mechanizmy.
________________________________________
Hlavný text
"Neexistuje žiadna úprimnejšia láska ako láska k jedlu."
George Bernard Shaw
úvod
Obezita, definovaná ako index telesnej hmotnosti (BMI)> 30, je stav, pri ktorom je adipozita abnormálne vysoká a môže byť výsledkom hyperfágie alebo zníženej rýchlosti metabolizmu (O'Rahilly, 2009). Nadmerná adipozita je hlavným rizikovým faktorom kardiovaskulárnych chorôb, rakoviny, cukrovky typu 2 a porúch súvisiacich s náladou, pričom obézni jedinci často trpia sociálnou stigmatizáciou ([Bean et al., 2008], [Centers for Disease Control and Prevention, 2009] a [Luppino et al., 2010]). Podľa Centra pre kontrolu chorôb (CDC) boli náklady na zdravotnú starostlivosť súvisiacu s obezitou v Spojených štátoch v rokoch 1998 až 2000 približne 213 miliárd dolárov. Ďalej možno 300,000 1999 úmrtí v USA ročne pripísať chorobám súvisiacim s nadváhou a obezitou (Allison a kol., 30), pričom obezita je druhou najčastejšou príčinou úmrtia, ktorej sa dá predchádzať v dôsledku užívania tabaku. Napriek tomu sa prevalencia obezity v západných spoločnostiach naďalej dramaticky zvyšuje, pričom súčasné odhady naznačujú, že viac ako 2010% dospelých v USA je obéznych (Flegal et al., XNUMX).
Väčšina konceptualizácií regulácie kŕmenia navrhuje, aby dva paralelné systémy ovplyvňovali príjem potravy ([Hommel et al., 2006], [Lutter a Nestler, 2009] a [Morton et al., 2006]). Homeostatický systém zahŕňa hormonálne regulátory hladiny hladu, sýtosti a adipozity, ako je leptín, ghrelín a inzulín, ktoré pôsobia na hypotalamické a mozgové kmene na stimuláciu alebo inhibíciu kŕmenia, aby sa udržali primerané úrovne energetickej rovnováhy. Dysfunkcia v zložkách homeostatického systému, ako je napríklad vrodený nedostatok leptínu, môže vyústiť do pretrvávajúceho stavu pozitívnej energetickej bilancie a rozvoja obezity ([Campfield et al., 1995], [Halaas et al., 1995] a [Pelleymounter et al., 1995]). Mechanizmy, ktorými hormonálne regulátory hladu a sýtosti pôsobia na hypotalamické a mozgové kmeňové obvody, aby si udržali energetickú homeostázu, boli podrobne opísané inde a čitatelia, ktorí sa zaujímajú o túto tému, sa odvolávajú na mnoho vynikajúcich recenzií na túto tému (napríklad [Abizaid a kol., 2006a] a [Gao a Horvath, 2007]).
Okrem metabolických systémov hrajú dôležitú úlohu v stravovacom správaní aj systémy odmeňovania mozgu ([Lutter a Nestler, 2009] a [Saper et al., 2002]). Spravidla sa jedlá nenápadnej chuti nejedia nadmerne, zatiaľ čo chutné jedlá sa často konzumujú aj po splnení energetických požiadaviek. Ľahký prístup k chutným energeticky výdatným potravinám sa považuje za hlavný environmentálny rizikový faktor obezity (Volkow and Wise, 2005) a nadmerná konzumácia chutných potravín sa považuje za hlavný faktor prispievajúci k nedávnemu nárastu obezity ([Finkelstein et al., 2005], [Hill a kol., 2003] a [Swinburn a kol., 2009]). Získanie príjemných účinkov chutného jedla je skutočne silnou motivačnou silou, ktorá u niektorých jedincov môže potlačiť homeostatické signály ([Shomaker et al., 2010], [Sunday et al., 1983] a [Zheng et al., 2009]) . Ak majú potkany na výber, uprednostňujú predovšetkým konzumáciu roztoku kalórií bez kalórií pred intravenóznymi infúziami kokaínu (Lenoir et al., 2007). Dobre kŕmené potkany sa navyše dobrovoľne vystavia extrémnemu chladu (−15 ° C), škodlivým teplotným bolestiam alebo averznému úderu nohou, aby získali chutné jedlá, ako napríklad koláč, mäsovú paštétu, arašidové maslo, Coca-Colu, cukríky M&M, čokoládu čipy alebo jogurtové kvapky, aj keď sú voľne dostupné menej chutné štandardné jedlá ([Cabanac a Johnson, 1983], [Foo a Mason, 2005] a [Oswald et al., 2010]). Tieto objavy poukazujú na to, ako intenzívne môžu makroživiny v chutných potravinách stimulovať systémy odmeňovania mozgu nezávisle od ich kalorickej hodnoty ([Wang et al., 2004a] a [Wang et al., 2004b]) a aká vysoká môže byť motivácia konzumovať chutné jedlá. aj pri absencii homeostatických energetických požiadaviek. Drogové návyky, ako napríklad kokaín alebo nikotín, môžu podobne spôsobiť vysokú hladinu konzumného správania, aj keď neobsahujú kalorické alebo výživové hodnoty. V skutočnosti sa kvôli mnohým podobnostiam medzi prejedaním pri obezite a nadmerným užívaním drog v závislosti (Volkow a Wise, 2005) tvrdilo, že obezita by sa mala považovať za poruchu mozgu a mala by sa zahrnúť ako diagnostická kategória do nadchádzajúceho piateho vydania. Diagnostického a štatistického manuálu duševných porúch (DSM-V) ([Devlin, 2007] a [Volkow a O'Brien, 2007]). V porovnaní s homeostatickými mechanizmami správania pri kŕmení je oveľa menej známe, ako hedonické systémy ovplyvňujú príjem potravy. Rovnako zostáva nejasný vplyv vnútorných zmien alebo zmien vyvolaných stravou na schopnosť systémov odmeňovania mozgu a na to, ako tieto účinky prispievajú k prejedaniu a obezite. Ďalej sú zhrnuté posledné údaje, ktoré poukazujú na pokrok v našom chápaní hedonických mechanizmov stravovania a stravovacích zmien v činnostiach odmeňovania mozgu, ktoré môžu prispieť k rozvoju obezity.
Aktivácia systémov odmeňovania mozgu v reakcii na potravu, ktorú je možné konzumovať: interakcie s hormonálnymi regulátormi energetickej bilancie
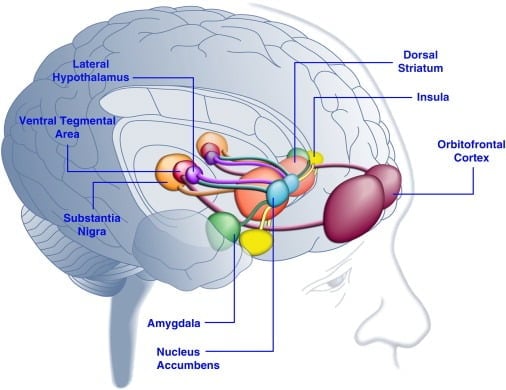
Spotreba chutných potravín môže zlepšiť náladu u ľudí ([Dallman et al., 2003] a [Macht a Mueller, 2007]) a podporiť stanovenie preferencie podmieneného miesta u laboratórnych zvierat ([Imaizumi et al., 2001] a [ Sclafani a kol., 1998]). Tieto účinky pravdepodobne súvisia so stimuláciou systémov odmeňovania mozgu chutnými potravinami (obrázok 1). Štúdie zobrazovania ľudského mozgu ukázali, že vizuálne alebo čuchové podnety súvisiace s potravinami a potravinami môžu aktivovať kortikolimbické a meso accumbens mozgové okruhy zapojené do odmeňovania, najmä orbitofrontálne kortexy (OFC), oblasti ostrovčekov, amygdaly, oblasti hypotalamu, striata a stredného mozgu. vrátane ventrálnej tegmentálnej oblasti (VTA) a substantia nigra (SN) ([Bragulat a kol., 2010], [Pelchat et al., 2004], [Schur et al., 2009] a [Simmons et al., 2005] ). Striatum, insula, anterior cingulate cortex a štruktúry stredného mozgu kódujú subjektívnu hodnotu odmien bez ohľadu na ich typ (napr. Jedlo, sex, peňažné odmeny), čo je v súlade s úlohou tejto neurónovej siete vo všeobecnej hedonickej reprezentácii (Sescousse et al.). , 2010). Na rozdiel od toho OFC zohráva obzvlášť významnú úlohu v reprezentáciách týkajúcich sa hodnoty špecifických typov odmien vrátane chutných potravín ([Man et al., 2009], [Rolls, 2008] a [Sescousse et al., 2010] ). Hlad môže zlepšiť chutnú aktiváciu kortikolimbických a stredných mozgom vyvolanú potravou u ľudí (LaBar et al., 2001). Napríklad intenzita aktivácie ventrálneho striata, amygdaly, insula a OFC v reakcii na vysoko kalorickú chutnú potravu bola ďaleko väčšia, keď boli ľudské subjekty skôr hladné ako dobre kŕmené (Goldstone a kol., 2009). To je v súlade so skutočnosťou, že obdobia hladu a diéty zvyšujú hodnotenie „sily“ chutných potravín a túžby po „lákavých“ potravinách ([Hofmann et al., 2010] a [Rolls et al., 1983]. ). Naopak, prekrmovanie môže redukovať neuronálne reakcie na chutné jedlo, najmä v ostrovnej kortexe a hypotalame (Cornier et al., 2009). Hédonická hodnota potravy je teda ovplyvnená metabolickým stavom, čo naznačuje, že regulátory metabolizmu, ako je leptín a ghrelín, môžu ovplyvňovať aktivitu hedonických systémov v mozgu. V súlade s týmto názorom sú u ľudí liečených leptínom alebo postprandiálnym peptidom YY3-36 (PYY) ([Batterham a kol., 2007] a [Farooqi et al., 2007]) leptínom alebo postprandiálnym faktorom odvodeným od čriev. napodobňovanie požitia jedla (Wang et al., 2008) malo zníženú aktivitu v oblastiach mozgu súvisiacich s odmenou. Naopak, hyperfágni ludskí pacienti s vrodeným deficitom leptínu vykazujú zvýšenú aktivitu v insulóznom kortexe a striate v reakcii na obraz potravy ([Baicy et al., 2007] a [Farooqi et al., 2007]). U týchto jedincov substitučná terapia leptínom zoslabila zvýšenú aktivitu ostrovčekovej a striatálnej aktivity a znížila samozrejmosť potravy ([Baicy et al., 2007] a [Farooqi et al., 2007]). Liečba leptínom tiež blokuje odmeňujúce vlastnosti sacharózy u potkanov s obmedzeným príjmom potravy podobných antagonistovi dopamínového receptora a-flupentixolu (Figlewicz et al., 2001). Okrem toho sú receptory leptínu exprimované na dopamínových neurónoch stredného mozgu v VTA a SN (Figlewicz et al., 2003), čo naznačuje, že leptín môže ovplyvňovať hédonické aspekty správania pri kŕmení prostredníctvom modulácie mesostriatálneho dopamínového prenosu. Potvrdením tejto možnosti infúzie leptínu do VTA inhibovali aktivitu dopamínových neurónov a znížili príjem potravy u potkanov (Hommel et al., 2006; pozri tiež Krügel et al., 2003). Naopak, knockdown leptínových receptorov vo VTA zvyšuje príjem potravy, zvýšenú lokomotorickú aktivitu a zvýšenú preferenciu chutných potravín u potkanov (Hommel et al., 2006). Leptín preto vykazuje inhibičný vplyv na mesoaccumbens dopamínový prenos, neurotransmiterový systém, ktorý bol silne zapojený do odmeňovania a motivácie, ale menej do energetickej homeostázy ([de Araujo et al., 2010] a [Vucetic and Reyes, 2010]). Nedávno sa ukázalo, že hormón ghrelín súvisiaci s hladom ([Kojima et al., 1999] a [Nakazato et al., 2001]) potencuje aktiváciu hedonických systémov v mozgu ako reakciu na podnety na jedlo (Malik et al. , 2008). Špecificky, ghrelín zlepšil aktiváciu OFC, amygdala, insula, striatum, VTA a SN v reakcii na obrazy vysoko chutných potravín u obéznych jedincov (Malik et al., 2008). U potkanov vykazuje ghrelín stimulačný účinok na dopamínové systémy stredného mozgu ([Abizaid et al., 2006b], [Jerlhag et al., 2006] a [Jerlhag et al., 2007]) a zvyšuje odmeňovanie chutných potravín (Perello et al., 2010).
Obrázok 1. Oblasti ľudského mozgu aktivované v reakcii na chutné jedlo alebo na podnety spojené s potravinami. Predpokladá sa, že orbitofrontálna kôra a amygdala kódujú informácie týkajúce sa hodnotovej hodnoty potravy ([Baxter a Murray, 2002], [Holland a Gallagher, 2004], [Kringelbach et al., 2003], [O'Doherty et al. , 2002] a [Rolls, 2010]). Na ostrove sa spracúvajú informácie týkajúce sa chuti jedla a jeho hedonického hodnotenia ([Balleine a Dickinson, 2000] a [Small, 2010]). Nucleus accumbens a dorzálne striatum, ktoré dostávajú dopaminergný vstup z ventrálnej tegmentálnej oblasti a substantia nigra, regulujú motivačné a stimulačné vlastnosti potravy ([Baicy et al., 2007], [Berridge, 1996], [Berridge, 2009], [Farooqi a kol., 2007], [Malik a kol., 2008] a [Söderpalm a Berridge, 2000]). Bočný hypotalamus môže regulovať odmeňujúce sa reakcie na chutné jedlo a riadiť správanie hľadajúce jedlo (Kelley et al., 1996). Tieto mozgové štruktúry pôsobia koordinovane tak, že regulujú poznávanie hedonických vlastností potravín, presúvajú pozornosť a úsilie k získaniu potravinových odmien a regulujú motivačnú hodnotu environmentálnych stimulov, ktoré predpovedajú dostupnosť potravinových odmien (Dagher, 2009). Kvôli prehľadnosti nie sú zobrazené všetky prepojenia medzi týmito štruktúrami.
Podobné mozgové oblasti sú aktivované chutným jedlom v mozgu potkana ako tie, ktoré sú aktivované u ľudí, ako je merané expresiou okamžitých skorých génov (IEG), ako je c-fos, arc alebo zif268. Chutné jedlo skutočne aktivuje dorzálne a ventrálne striatum, VTA, laterálny hypotalamus (LH) a centrálne a bazolaterálne jadrá kortikálnych štruktúr amygdaly a odmien u potkanov ([Angeles-Castellanos et al., 2007], [Park and Carr, 1998] a [Schiltz et al., 2007]). Zaujímavé je, že imunoreaktivita Fos bola v skutočnosti znížená v laterálnom a mediálnom habenule u potkanov po chutnej konzumácii potravín (LHb) (Park a Carr, 1998). U nehumánnych primátov je LHb aktivovaný averzívnymi podnetmi alebo vynechaním očakávaných odmien a inhibovaný dodávkou chutnej šťavy (Matsumoto a Hikosaka, 2007). Okrem toho aktivita LHb inhibuje neuro-neuróny obsahujúce mezoaccumbény, ktoré súvisia s odmenou, prostredníctvom nepriamych dráh zahŕňajúcich rastromediálne tegmentálne jadro (RMTg) (Jhou et al., 2009). Habenulárna aktivita je teda nepriamo úmerná potravinovému hedoniku, čo naznačuje, že habenulárny komplex môže ovplyvniť nehomeostatické stravovanie. Nedávno sa ukázalo, že aktivácia LHb znižuje spotrebu sacharózy u potkanov, zatiaľ čo lézie LHb zvyšujú správanie, pri ktorom dochádza k sacharóze (Friedman et al., 2011). Berúc do úvahy, že habenulárny komplex je malý a náročný na identifikáciu a funkčný obraz u ľudí (Salas a kol., 2010), môže to vysvetliť, prečo neboli zmeny v habenulárnej aktivite hlásené v štúdiách zobrazovania u ľudí v reakcii na chutné jedlo.
Obvody mozgu, ktoré regulujú Hedonic Eating: Midbrain Dopamin Systems
Cesta dopamínu mezoaccumbens sa aktivuje u ľudí a laboratórnych zvierat v reakcii na chutné jedlo alebo chuťové poháriky súvisiace s jedlom a leptín, ghrelín a ďalšie regulátory chuti do jedla ovplyvňujú aktivitu v tomto systéme. To naznačuje, že dopamínové systémy so stredným mozgom hrajú dôležitú úlohu pri chutnej konzumácii potravy. Snáď najjasnejšou indikáciou toho, že prenos dopamínu v strednom mozgu ovplyvňuje chutný príjem potravy u ľudí, je skutočnosť, že pacienti s Parkinsonovou chorobou (PD), u ktorých dochádza k degenerácii neurónov obsahujúcich dopamín v strednom mozgu, majú tendenciu konzumovať menej potravy ako jedinci bez ovplyvnenia (Nirenberg a Waters, 2006). Liečba pacientov s PD agonistami dopamínových receptorov môže navyše vyvolať nutkavú konzumáciu chutného jedla ([Dagher a Robbins, 2009] a [Nirenberg a Waters, 2006]). Agonisty dopamínových receptorov môžu v skutočnosti vyvolať hedonické prejedanie sa aj u jedincov bez PD (Cornelius et al., 2010). U zvierat chutné roztoky sacharózy stimulujú prenos dopamínu v NAc (Hernandez a Hoebel, 1988), čo je účinok v súlade so zobrazovacími štúdiami na ľudskom mozgu (Small et al., 2003). Použitím cyklickej voltametrie s rýchlym skenovaním sa ukázalo, že podnety predpovedajúce dodanie sacharózovej odmeny alebo neočakávané dodanie sacharózy vyvolali prenos dopamínu v NAc ([Roitman et al., 2004] a [Roitman et al., 2008]). Ďalej malo neočakávané dodanie škodlivých chinínových roztokov opačný účinok, pretože znížilo prenos dopamínu naumbumbalu (Roitman et al., 2008). Napokon myši, u ktorých bol deaktivovaný enzým tyrozínhydroxyláza (TH), ktorý spôsobuje ich nedostatok dopamínu, stále preukazujú výraznú preferenciu roztokov sacharózy (alebo sacharínu) v porovnaní s vodou, ale konzumujú menšie celkové množstvo sacharózy ako kontrolné myši ( Cannon a Palmiter, 2003). To naznačuje, že myši s deficitom dopamínu môžu stále detekovať chutnosť sacharózy a uprednostňujú tieto roztoky pred vodou, ale nie sú schopné udržať spotrebu chutných roztokov. Preto sa navrhlo, že prenos dopamínu mezoaccumbens reguluje motivačné aspekty správania pri kŕmení, ktoré sa podieľajú na získavaní potravín, a že ďalšie neurotransmiterové systémy pravdepodobne regulujú hedonické aspekty chutnej spotreby potravy.
Obvody mozgu, ktoré regulujú Hedonic Eating: Striatohypothalamic Systems
Infúzia agonistov μ-opioidných receptorov, ako je [D-Ala2-N-Me-Phe4-gly-ol5] -fenepín (DAMGO) do NAc stimuluje kŕmenie u potkanov s prístupom k potrave ad libitum (tj nehomeostatické podávanie) ( [Peciña a Berridge, 2005] a [Zhang et al., 1998]). Naopak, antagonisty opioidných receptorov infúziou do NAc znižujú spotrebu preferovaných potravín bez ovplyvnenia príjmu menej prijateľných alternatív (Kelley a kol., 1996). Tieto údaje sú v súlade s názorom, že striatálne opioidné systémy regulujú hedonické vlastnosti chutných potravín. Obzvlášť dôležitú úlohu pri nehomeostatickom podávaní hrá oblasť škrupiny NAc a najmä hédonických „horúcich miest“ v rostrodorzálnej oblasti mediálneho obalu ([Peciña a Berridge, 2005] a [Peciña a kol., 2006b]). Pretože aktivácia ó-opioidného receptora vedie k inhibícii aktivity stredného ostnatého neurónu v NAc, navrhlo sa, že NAc shell má tonický inhibičný vplyv na chutnú spotrebu potravy (Kelley et al., 2005). V súlade s týmto názorom, stimulácia inhibičných receptorov GABAA alebo GABAB ([Basso a Kelley, 1999] a [Stratford a Kelley, 1997]) alebo blokáda excitačných ionotropných glutamátových receptorov (Maldonado-Irizarry et al., 1995) v NAc škrupine zvyšuje spotrebu potravín. Podobne excitotoxická lézia NAc škrupiny tiež zvyšuje spotrebu potravy a zvyšuje citlivosť na potravinovú odmenu ([Johnson a kol., 1996] a [Maldonado-Irizarry a Kelley, 1995]). Tieto manipulácie prednostne spúšťajú najmä spotrebu energeticky náročných chutných potravín ([Basso a Kelley, 1999], [Kelley a kol., 2005] a [Zhang et al., 1998]).
Vzhľadom na veľký vplyv akumulovanej signalizácie na hedonické kŕmenie používali Thompson a Swanson (2010) postup sledovania obvodov na presné určenie presných anatomických sietí, prostredníctvom ktorých môže NAc ovplyvniť chutnú spotrebu potravín. V týchto elegantných štúdiách boli potkanom aplikované dve neprekrývajúce sa injekcie anterográdnych / retrográdnych indikátorov (nazývané COIN) do miest NAc škrupiny, ktoré silne ovplyvňujú chutnú spotrebu potravy a identifikovali sa aferentné / eferentné spojenia. Ukázalo sa, že miesta súvisiace s kŕmením v NAc rozširujú inhibičné projekcie prevažne na predný LH a ventrálny pallidum (VP) (Thompson a Swanson, 2010). Na rozdiel od zvyšku NAc, ktorý sa intenzívne premieta do VTA, hédonické horúce miesta súvisiace s potravinami v projekte NAc shell do medzifázového jadra (IFN), štruktúry nachádzajúcej sa v blízkosti VTA, ktorá rozširuje dopaminergné projekcie recipročným spôsobom späť na NAc shell (Thompson a Swanson, 2010). Ďalej, predný LH premieta LHb (Thompson a Swanson, 2010), čo opäť naznačuje, že habenulárny komplex môže hrať úlohu v potravinárskom hedoniku (Friedman et al., 2011).
Vyššie uvedené údaje ukazujú, že LH dostáva prominentný inhibičný vstup z miest v NAc, ktoré majú tonický inhibičný vplyv na chutnú spotrebu potravy. LH má tiež funkčnú konektivitu s inými miestami kortikálneho a limbického mozgu, ktoré sa podieľajú na organizovaní a riadení správania smerom k získaniu chutných potravín (obrázok 1), ako sú OFC, insula a amygdala. Dôležité je, že inaktivácia LH ruší stimulačné účinky NAc manipulácie na príjem potravy ([Maldonado-Irizarry et al., 1995] a [Will et al., 2003]). Okrem toho inaktivácia NAc shell zvyšuje aktivitu LH, najmä neurónov LH, ktoré syntetizujú neuropeptid hypokretín (tiež známy ako orexín), ako sa meria Fos imunoreaktivitou ([Baldo et al., 2004] a [Stratford and Kelley, 1999 ]). Infúzia agonistu μ-opioidného receptora DAMGO do škrupiny NAc aktivuje neuróny obsahujúce hypokretín v hypotalame (Zheng et al., 2007) a narušenie prenosu hypokretínu vo VTA ruší chutný príjem potravy vyvolaný infúziami intra-NAc DAMGO (Zheng a kol., 2007). Hédonické horúce miesta v NAc škrupine teda pôsobia tonicky inhibične na neuróny LH, a najmä neuróny obsahujúce hypokretín (Louis et al., 2010), čím sa obmedzuje spotreba chutných potravín. Narušenie tohto akumulovaného „stop signálu“, napríklad prostredníctvom zvýšenej signalizácie opioidného receptora, vedie k zvýšenej aktivite LH, ktorá riadi nehomeostatickú spotrebu chutných potravín (obrázok 2).
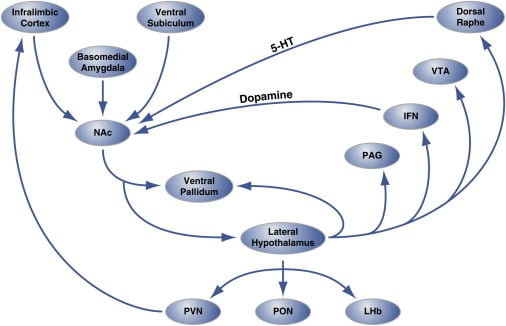
Obrázok 2. Organizácia Hedonic „Hot Spots“ v Nucleus Accumbens Shell, ktorá reguluje Hedonic Eating
Oblasť plášťa nucleus accumbens (NAc) je inervovaná z miest kortikálneho a limbického mozgu a projektuje laterálny hypotalamus a ventrálnu pallidum. Na druhej strane laterálny hypotalamus tiež pôsobí na ventrálnu pallidum a tiež na PAG, IFN, VTA a dorzálne jadro raphe. IFN a dorzálny raphe rozširujú dopaminergné a serotonergné projekcie, v tomto poradí, späť na NAc. Bočný hypotalamus tiež inervuje talamické (PVN a PON) a epithalamické (LHb) štruktúry. Neznázornené sú menšie projekcie z laterálneho hypotalamu do oblastí mozgu septálu. 5-HT, serotonín; IFN, intrakikulárne jadro; LHb, laterálny habenula; PON, preoptické jadro; PVN, paraventrikulárne jadro talamu; VTA, ventrálna tegmentálna oblasť. Obrázok je upravený so súhlasom Thompson a Swanson (2010).
Obvody mozgu, ktoré regulujú Hedonic Stravovanie: Striatopalídové systémy
Okrem LH, NAc shell neuróny tiež projektujú na VP (obrázok 2). V zaujímavej sérii experimentov sa ukázalo, že akumulované projekcie VP a LH môžu regulovať disociovateľné aspekty nehomeostatického jedenia (Smith a Berridge, 2007). Infúzie DAMGO do NAc shell alebo VP zvýšených orofaciálnych reakcií na roztoky sacharózy predpokladali, že budú odzrkadľovať „sympatické“ reakcie u potkanov (tj odpoveď na chuť) a tiež zvýšená spotreba potravy (Smith a Berridge, 2007). Infúzie naloxónu buď do NAc alebo VP znížili reakcie na líčenie tváre na sacharózu, čo naznačuje, že synchronizovaný prenos opioidov v NAc a VP je nevyhnutný na spracovanie chutnosti potravy súvisiacej s informáciami. Avšak naloxón podávaný do NAc, ale nie VP, znižoval nehomeostatické stravovanie (Smith a Berridge, 2007), čo naznačuje, že nehomeostatické stravovanie sa vyskytuje nezávisle od tejto NAc → VP konektivity a namiesto toho sa pravdepodobne spolieha na NAc → LH cestu ([Smith and Berridge, 2007] a [Taha et al., 2009]). V súlade s predstavou, že aspekty nonhomeostatického jedenia môžu byť disociované, záznamy s jednou jednotkou ukázali, že populácia NAc neurónov sa javí, že selektívne kóduje informácie súvisiace s relatívnymi posilňujúcimi vlastnosťami potravy (tj chuť) (Taha a Fields, 2005) , U tých istých potkanov sa ukázalo, že zmeny aktivity druhej populácie NAc neurónov sa zhodujú so začatím kŕmenia (Taha a Fields, 2005).
Obvody mozgu, ktoré regulujú Hedonic Eating: Amygdalar Systems
Ďalej podporuje názor, že aspekty nehomeostatického stravovania sú disociovateľné, infúzie naloxónu do NAc shell alebo VP, ale nie bazolaterálna amygdala (BLA), znížili chutnosť roztokov sacharózy (Wassum et al., 2009). Keď sa však naloxón alebo CTOP podali antagonistom μ-opioidného receptora do BLA, ale nie do NAc shell alebo VP, došlo k výraznému zníženiu zvýšenej motivácie odpovedať na sacharózové roztoky typicky pozorované v hladovom stave ([Wassum et al. , 2011] a [Wassum et al., 2009]). To naznačuje, že stimulačné vlastnosti sacharózy sú regulované amygdalar obvodmi. Vyššie uvedené zistenia celkovo ukazujú, že rôzne aspekty hedonického jedenia, ako je spracovanie informácií týkajúcich sa chutnosti jedla, prístupové správanie a zvýšenie stimulačnej hodnoty chutných potravín u hladných zvierat, sú diferencovane regulované diskrétnymi mikrocirkuláciami v kontexte väčšia kortikolimbicko-striatopalídia-hypotalamicko-talakortikálna štruktúra (obrázok 2).
Do adaptácie v mozgu Hedonic Circuitries Drive kompulzívne jedenie?
Funkčnú relevanciu hedonických hotspotov v NAc škrupine a ich vplyv na širšie obvody súvisiace s výživou v mozgu uvažovali Kelley et al. (2005). Predpokladajú, že dráha NAc shell → LH, spolu s regulačnými oblasťami mozgu (downstream a downstream) (obrázok 2) slúži na „sentinelové“ účely (Kelley et al., 2005). Konkrétne navrhujú, aby aj pri hladných zvieratách, keď je cesta k jedlu silná, bola zachovaná schopnosť zastaviť správanie sa v prípade ohrozenia životného prostredia (Kelley a kol., 2005). Aktivácia neurónov NAc shell a sprievodná inhibícia neurónov LH ako taká môže narušiť pokračujúce podávanie a uľahčiť zmenu správania na vhodnejšie adaptívne reakcie, ako je zmrazenie alebo únik (Kelley et al., 2005). Ak je to skutočne tak, potom bude dôležité preskúmať, či je táto NAc shell → LH kontrolná dráha ohrozená nadmernou konzumáciou chutných potravín alebo genetickými faktormi, ktoré ovplyvňujú zraniteľnosť voči obezite. S týmto vedomím naše laboratórium a iní nedávno uviedli, že nadmerná spotreba chutných kaloricko-hustých potravín je spojená so vznikom nutkavého správania sa kŕmenia u potkanov ([Johnson a Kenny, 2010], [Latagliata et al., 2010] a [Oswald et al., 2010]). Konkrétne sme zistili, že chutná konzumácia potravy u obéznych potkanov bola odolná voči narušeniu averzívnym podmieneným stimulom, ktorý predpovedal negatívny výsledok (elektrický šok) (Johnson a Kenny, 2010). Preto bude dôležité testovať, či deficity v NAc shell → LH kontrolnej dráhe vyvolanej prejedaním prinajmenšom čiastočne prispievajú k výraznému zlyhaniu jedincov s nadváhou a obezitou pri využívaní informácií týkajúcich sa škodlivých dôsledkov ich konzumného správania na zmiernenie ich príjmu potravy. ,
Zmenená aktivita odmeňovania mozgu v obezite: Štúdie zobrazovania ľudského mozgu
Získanie stimulačných účinkov chutných potravín na systémy odmeňovania mozgov sa považuje za dôležitý motivačný faktor prispievajúci k prejedaniu. Dôležitou otázkou teda je, či zmeny funkcie odmeňovania mozgu môžu prispieť k rozvoju obezity. Intuitívna predpoveď je, že zvýšená konštitutívna odozva systémov odmeňovania mozgov na chutné jedlo by viedla k prejedaniu a nárastu telesnej hmotnosti. V súlade s touto hypotézou vykazujú jedinci s vysokou úrovňou citlivosti odmeňovania znakov zvýšenú aktivitu v oblastiach mozgu, ktoré sa podieľajú na odmeňovaní potravín, vrátane NAc, amygdala, OFC a VP, po vystavení chutným potravinám, ako je čokoládový koláč a pizza (Beaver et al. , 2006). Obézni jedinci podobným spôsobom demonštrujú zvýšenú aktiváciu obvodov odmeňovania mozgu v reakcii na chutné potraviny alebo podnety súvisiace s potravou v porovnaní so štíhlymi kontrolami ([Gautier et al., 2000], [Karhunen et al., 1997] a [Rothemund et al., 2007 ]). Vysoká úroveň citlivosti na odmenu bola tiež korelovaná so zvýšenou telesnou hmotnosťou u ľudských subjektov ([Davis et al., 2004] a [Franken a Muris, 2005]). Dôležité však je, že obézna žena (BMI> 30) mala vyššiu hladinu anhedónie (tj zníženú základnú citlivosť na odmenu) ako žena s nadváhou (BMI> 25 <30) (Davis et al., 2004). Podobne, Stice a spolupracovníci (2008b) ukázali, že obézne dospievajúce dievčatá mali zvýšenú aktiváciu oblastí s kĺbmi a inými kortikálnymi oblasťami v reakcii na chutné potraviny alebo podnety súvisiace s potravou v porovnaní s subjektmi s chudobnou kontrolou, ale že aktivácia oblasti caudate v oblasti striatum v reakcii na chutnú potravu nepriamo korelovalo s BMI u obéznych jedincov. Okrem toho ženy, ktoré v priebehu obdobia 6 získali hmotnosť, mali výrazný pokles aktivity striatálnej odozvy na chutnú potravu počas tohto obdobia v porovnaní so ženami, ktoré nezvyšovali hmotnosť (Stice et al., 2010a). Ak vezmeme toto všetko dohromady, zdá sa, že hypersenzitivita obvodov odmeňovania môže predisponovať jedinca k prejedaniu a prírastku hmotnosti (Stice et al., 2010b). Ako sa však zvyšuje prírastok hmotnosti, potom sa môžu začať objavovať deficity v činnosti špecifických zložiek systému odmeňovania mozgu, najmä striatum. Bolo navrhnuté, že vznik tohto stavu odmeny hyposensitivty môže zachovať nadmernú spotrebu chutných potravín s cieľom prekonať tieto deficity odmien ([Stice a kol., 2008a] a [Wang et al., 2002]). Zdá sa teda, že príliš málo alebo príliš veľa potravinovej odmeny zvyšuje zraniteľnosť voči prejedaniu a obezite (Stoeckel, 2010). Atraktívnym koncepčným rámcom na zosúladenie týchto zdanlivo protichodných názorov je to, že kortikolimické oblasti, ktoré sa podieľajú na organizovaní správania pri získavaní odmien za potraviny a pri predpovedaní očakávanej budúcej odmeny v potravinách, sa môžu stať hyperaktívnymi u jednotlivcov s nadváhou a osôb s predispozíciou k obezite. Naopak, striatálne mozgové miesta, ktoré spracúvajú skutočnú skúsenosť radosti zo hedonického jedenia, sa môžu postupne u týchto jedincov postupne znižovať. Očakáva sa preto, že relatívna motivačná hodnota chutných potravín sa bude zvyšovať počas vývoja obezity v tom istom čase, keď sa zníži hédonická hodnota získaná konzumáciou chutných potravín.
Zmenená aktivita odmeňovania mozgu v obezite: Štúdie hlodavcov
Účinky chutnej konzumácie potravín na systémy odmeňovania mozgov boli priamo vyhodnotené na laboratórnych zvieratách s použitím postupu odmeny za mozgovú stimuláciu (BSR). Je dobre známe, že elektrická stimulácia LH, ktorá prijíma tonický inhibičný vstup z akumulovaných hédonických horúcich miest (obrázok 2), je vysoko odmeňovaná a potkany budú tvrdo pracovať na stimulácii tejto oblasti mozgu, napr. (Markou a Frank, 1987 ). Okrem podporovania samostimulačného správania môže elektrická stimulácia LH tiež indukovať intenzívne záchvaty kŕmenia (Margules a Olds, 1962) a navrhlo sa, aby odmeňujúce vlastnosti stimulácie LH mohli súvisieť s vnútornou úlohou tejto stimulácie. miesto mozgu v chutných a stimulačných vlastnostiach potravín (Margules a Olds, 1962). V súlade s týmto názorom hlad a úbytok hmotnosti zvyšujú citlivosť potkanov na samo stimuláciu LH (od firmy Blundell a Herberg, 1968), [Carr a Simon, 1984] a [Margules and Olds, 1962]. ktoré môžu byť blokované intracerebroventrikulárnou infúziou leptínu (Fulton et al., 2000). Naopak, elektrická samo-stimulácia LH je inhibovaná u zvierat na zvieratách (Wilkinson a Peele, 1962). Prekrmovanie potkanov intragastrickou kŕmnou trubicou (Hoebel a Teitelbaum, 1962), žalúdočnou distenziou alebo intravenóznou infúziou glukagónu, ktorá napodobňuje postprandiálnu sýtosť ([Hoebel, 1969], [Hoebel a Balagura, 1967] a [Mount and Hoebel, 1967]) ), všetky zoslabujú reakciu na stimuláciu LH. V skutočnosti, potkany, ktoré predtým rázne reagovali na odmeňovanie stimulácie LH, budú reagovať, ako keby táto stimulácia bola averzívna po príjme potravy alebo vývoji obezity (Hoebel a Thompson, 1969). Z toho vyplýva, že chronické obmedzenie potravy a úbytok hmotnosti zvyšuje, zatiaľ čo prebytok potravy sa znižuje, citlivosť miest súvisiacich s odmenou v LH. Citlivosť LH neurónov na odmeňovanie elektrickej stimulácie môže preto poskytnúť dôležitý pohľad na fungovanie mozgových obvodov, ktoré regulujú hedonické reakcie na potraviny.
Keďže ľahký prístup k energeticky hustým chutným potravinám a následná nadmerná spotreba sa považujú za hlavný environmentálny faktor prispievajúci k obezite (Volkow a Wise, 2005), naše laboratórium nedávno využilo postup BSR na posúdenie aktivity odmeňovania mozgu u potkanov s rozšíreným prístupom k chutným potravinám. , Konkrétne sme zaznamenali reakciu na elektrickú stimuláciu LH u potkanov, ktorí mali ad libitum prístup k nutričnému krmivu samostatne alebo v kombinácii s 18-23 hr denne prístup k chutnej strave. Táto diéta sa skladala z tvarohového koláča, slaniny, klobásy a ďalších chutných potravín (Johnson a Kenny, 2010). Zistili sme, že potkany s rozšíreným prístupom k chutnému jedlu rýchlo získali významné množstvo hmotnosti a preukázali progresívne sa zhoršujúci deficit v mozgovej odmene (odráža sa ako znížená citlivosť na odmeňovanie stimulácie LH) (Johnson a Kenny, 2010; obrázok 3). To naznačuje, že rozvoj obezity vyvolanej diétou je spojený s postupným znižovaním citlivosti miest odmeňovania v LH (Johnson a Kenny, 2010). Deficity v signalizácii odmeňovania boli tiež hlásené u dospelých potkanov, ktoré predtým mali neobmedzený prístup k sacharóze alebo potravinám s vysokým obsahom tuku počas dospievania ([Teegarden et al., 2009], [Vendruscolo et al., 2010a] a [Vendruscolo et al., 2010b]). Tieto účinky u potkanov pripomínajú zníženú aktiváciu striatu v reakcii na potravinovú odmenu opísanú vyššie u ľudských jedincov, keď získali hmotnosť počas periódy 6 (Stice et al., 2010a; pozri obrázok 4). Takýto deficit vyvolaný diétou u potkanov s nadváhou a možno aj u ľudí, ktorí naberajú na váhe, pravdepodobne odráža kontraadaptívnu reakciu v obvodoch potravinovej odmeny, aby sa postavili proti ich nadmernej stimulácii chutnými potravinami (Johnson a Kenny, 2010). Dôležitým aspektom tohto zistenia je, že podobné deficity v odmeňovaní sú zistené aj u potkanov, ktoré nadmerne konzumujú kokaín alebo heroín ([Ahmed a kol., 2002], [Kenny a kol., 2006] a [Markou a Koob, 1991]; Obrázok 3). V skutočnosti sa predpokladalo, že dysfunkcia vyvolaná drogami môže prispieť k prechodu z kontrolovaného na nekontrolované užívanie drog poskytnutím nového zdroja motivácie na konzumáciu lieku s cieľom zmierniť pretrvávajúci stav zníženej odmeny (Ahmed a Koob , 2005] a [Koob a Le Moal, 2008]). Preto je možné, že deficity v citlivosti miest odmeňovania v LH vyvolanom prejedaním môžu zvýšiť dlhodobú perzistenciu chutnej konzumácie potravy u potkanov s nadváhou posunutím preferencií na stravu s vyšším hedonickým dopadom na zmiernenie pretrvávajúceho negatívneho stavu. odmena.
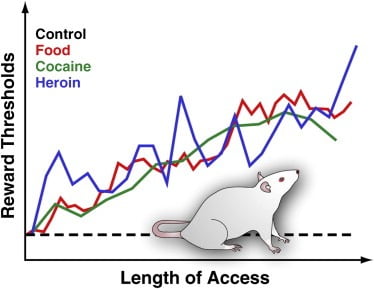
Obrázok 3. Odmeny v potkanoch s predĺženým denným prístupom k potravinám, ktoré sa konzumujú, kokaínu alebo heroínu
Na meranie prahov odmien je stimulačná elektróda chirurgicky implantovaná do laterálneho hypotalamu potkanov, čo je oblasť, v ktorej elektrická stimulácia silne odmeňuje a môže spúšťať intenzívne záchvaty kŕmenia. Po uzdravení sa zvieratám umožňuje samovoľne stimulovať túto oblasť otáčaním kolieska. Po stanovení stabilného samo-stimulačného správania sa stanoví minimálna intenzita stimulácie, ktorá si zachovala vlastné stimulačné správanie (tj prahová hodnota odmeny). Táto hranica odmeňovania poskytuje operačnú mieru činnosti systému odmeňovania. Prahové hodnoty odmien zostávajú stabilné a nezmenené u kontrolných potkanov, ktoré majú prístup k štandardnému laboratórnemu krmivu a ktoré zostávajú naivné. Prahové hodnoty sa však postupne zvyšujú u potkanov s predĺženým denným prístupom k energeticky hustej chutnej strave pozostávajúcej z chutných potravín (napr. Tvarohový koláč, slanina, čokoláda atď.). Podobne, prahy odmeňovania sa postupne zvyšovali u potkanov, ktoré rozšírili denný prístup k intravenóznym infúziám kokaínu alebo heroínu. Zvýšený prah odmeny sa interpretuje tak, aby odrážal zníženú citlivosť systému odmeňovania mozgov. Tieto účinky naznačujú, že nadmerná konzumácia chutných potravín a súvisiaci nárast telesnej hmotnosti môže vyvolať hlboký deficit v odmeňovaní mozgu podobný tým, ktoré sú vyvolané nadmernou konzumáciou návykovej drogy.
Obrázok 4. Striatálna plasticita v ObesityWeight zisku je spojená so zníženou aktiváciou striatálu v reakcii na chutné jedlo, merané pomocou fMRI, a nižšou úrovňou dostupnosti striatálneho dopamínového D2 receptora (D2R) u ľudí (podrobnosti pozri v texte).
Nedostatočný receptor dopamínu D2 v obezite
Niekoľko nedávnych správ odhalilo možné mechanizmy, prostredníctvom ktorých sa môže objaviť deficit v odozve na nadmernú spotrebu chutných potravín počas vývoja obezity. Ako bolo uvedené vyššie, ženy, ktoré v priebehu obdobia 6 získali hmotnosť, mali výrazný pokles striatálnej aktivity v reakcii na chutnú potravu v tomto časovom období v porovnaní so ženami, ktoré nezvyšovali hmotnosť (Stice et al., 2010a; Obrázok 4). Na lačno jednotlivci, ktorí majú možnosť jesť svoje obľúbené jedlo k sýtosti, mali nižšie hladiny väzby dopamínového antagonistu D2 receptora (D2R), raclopridu v striate (Small et al., 2003), čo naznačuje, že signalizácia D2R klesá v reakcii na chutnú spotrebu potravy. Obézni jedinci majú v skutočnosti nižšiu úroveň dostupnosti striatálneho D2R v porovnaní so štíhlymi kontrolami ([Barnard et al., 2009], [Stice a kol., 2008a] a [Wang et al., 2001]; Obrázok 4), zatiaľ čo úbytok hmotnosti u obéznych pacientov je spojená so zvýšenou hustotou striatálneho D2R (Wang et al., 2008). Vzhľadom na to, že prenos striatálneho dopamínu zohráva kľúčovú úlohu pri regulácii hedonického jedenia, adaptívne zníženie signalizácie D2R by mohlo prispieť k zníženiu citlivosti striata na chutné jedlo u obéznych jedincov. Na otestovanie tejto možnosti, Small a spolupracovníci skúmali činnosť v obvodoch odmeňovania mozgu v reakcii na chutný mliečny kokteil u kontrolných jedincov a tých, ktorí nesú TaqIA A1 alelu (Felsted et al., 2010). Polymorfizmus dĺžky restrikčného fragmentu TaqIA je downstream od génu D2R (Neville et al., 2004) a jedinci nesúci alelu A1 polymorfizmu majú medzi 30% –40% menej striatálnych D2R v porovnaní s tými, ktoré niesú alelu ([Jönsson a spol., 1999], [Ritchie a Noble, 2003] a [Stice a kol., 2010b]). Okrem toho majú alely A1 alely tiež znížený metabolizmus glukózy v striatálnych a kortikálnych oblastiach mozgu, ktoré sa podieľajú na hedonických odpovediach na potravu (Jönsson et al., 1999). Jedinci nesúci alelu TaqIA A1 sú nadmerne zastúpení v obéznych populáciách ([Barnard et al., 2009], [Stice a kol., 2008a] a [Wang et al., 2001]). Ďalej, alela A1 tiež zvyšuje zraniteľnosť voči závislosti od alkoholu, opioidov a psychomotorických stimulantov ([Lawford et al., 2000], [Noble et al., 1993] a [Noble et al., 2000]). Bolo zistené, že oblasti stredného mozgu, vrátane VTA a SN, ktoré poskytujú dopaminergný vstup do striata, boli aktivované v reakcii na chutný mliečny koktail u kontrolných jedincov (Felsted et al., 2010). Naopak, aktivita v týchto miestach mozgu bola v skutočnosti znížená v reakcii na potravinovú odmenu u nosičov alel A1 (Felsted et al., 2010). Podobné inverzné reakcie v aktivácii mozgu medzi alelickými nosičmi A1 a ne-nosičmi boli tiež detegované v talamických a kortikálnych miestach mozgu (Felsted et al., 2010). Tieto údaje sú vysoko konzistentné s kľúčovou úlohou D2R pri regulácii mesostriatálnej reakcie na chutné jedlo. Stice a jeho kolegovia (2008a) zistili inverznú koreláciu medzi BMI a aktiváciou striata (caudate a putamen) v reakcii na chutný čokoládový mliečny koks u ľudských pacientov. Navyše tento inverzný vzťah bol najvýraznejší u jedincov nesúcich alelu TaqIA A1 (Stice a kol., 2008a). Budúci prírastok hmotnosti u týchto jedincov, meraný 1 rok po počiatočnom zobrazovaní mozgu, ukázal, že veľkosť striatálnej aktivácie v reakcii na chutnú potravu bola negatívne korelovaná s prírastkom hmotnosti u jedincov s alelou A1 a pozitívne korelovala u subjektov bez alel A1 (alely). Stice a kol., 2008a). V následnej štúdii sa zistilo, že veľkosť aktivácie striatálu v reakcii na predstavu o jedení chutných potravín, na rozdiel od jej skutočnej spotreby, bola nepriamo korelovaná s prírastkom telesnej hmotnosti v nasledujúcom roku u jedincov s alelou A1, ale pozitívne koreluje s non-A1 alelovými subjektmi (Stice et al., 2010b). Tieto zistenia naznačujú, že D2R reguluje striatálnu citlivosť na chutnú potravu a že znížená signalizácia D2R indukovaná prírastkom hmotnosti alebo genetickými faktormi môže zvýšiť zraniteľnosť voči obezite.
Nedostatočná signalizácia D2R prispieva k deficitu odmeňovania v obezite
Podobne ako v prípade downregulovaných hladín striatálneho D2R u obéznych ľudských subjektov, hladiny D2R sú tiež znížené u myší a potkanov kŕmených chutnou stravou (napr. [Colantuoni et al., 2001], [Geiger et al., 2009] a [Johnson and Kenny A 2010]) a u potkanov geneticky predisponovaných k obezite (potkany Zucker) (Thanos et al., 2008). Naše laboratórium priamo skúmalo úlohu narušenia prenosu striatálneho dopamínu vo všeobecnosti a najmä zníženie signalizácie D2R, a to najmä v prípade deficitu závislého od závislosti, ktorý sa objavil u potkanov počas vývoja obezity (pozri obrázok 5). Konkrétne sme testovali účinky zníženia expresie striatálnych D2Rs u potkanov s použitím vírusom sprostredkovanej RNA interferencie, potom stanovením prahových hodnôt BSR, keď potkany mali prístup len k chow alebo chow v kombinácii s 18 – 23 hr denne prístup k chutnému vysokému množstvu energie diéta (tj stravovanie v jedálni) (Johnson a Kenny, 2010). Zistili sme, že reakcia na odmeňovanie stimulácie LH začala klesať takmer okamžite po vystavení stravovacej diéte v potkanoch D2R knockdown (Johnson a Kenny, 2010; obrázok 5). Zníženie hladín striatálneho D2R teda rýchlo zrýchľuje vznik hypofunkcie odmien u potkanov s rozšíreným prístupom k vysoko chutným potravinám, čo je proces, ktorý zvyčajne trvá mnoho týždňov, kým sa u kontrolných potkanov objaví rozšírený prístup k chutnej strave. Avšak knockdown striatálnych D2Rs u potkanov s prístupom k jedlu sa nezmenilo, čo by zodpovedalo za odmeňovanie LH stimulácie, čo naznačuje, že zmenšená striatálna D2R signalizácia interaguje s inými diétami indukovanými adaptívnymi odpoveďami v obvodoch odmeňovania mozgu, aby sa spustila hyposenzitivita odmeny. Okrem znížených hladín D2R sa v mozgoch obéznych potkanov menia aj iné aspekty striatálneho dopaminergného prenosu. Napríklad Sprague-Dawley potkany, ktoré sa živia selektivitou na rýchle zvýšenie hmotnosti na diéte s vysokou energiou (potkany náchylné na obezitu) majú nižšie bazálne a evokované hladiny dopamínu v NAc ako potkany, ktoré sú rezistentné na zvýšenie telesnej hmotnosti (potkany rezistentné voči obezite) (Geiger et al. al., 2008, pozri tiež Rada a kol., 2010). Potkany náchylné na obezitu majú tiež znížené hladiny biosyntetických a skladovacích mechanizmov dopamínu, čo naznačuje, že zlyhanie pri produkcii a uvoľňovaní dopamínu prispieva k deficitu prenosu dopamínu striatum u obéznych potkanov (Geiger et al., 2008). Potkany, u ktorých sa vyvinula obezita prostredníctvom nadmernej konzumácie chutnej vysokoenergetickej diéty, mali tiež nižšie bazálne a evokované hladiny dopamínu v NAc v porovnaní s potkanmi, ktoré mali prístup iba k štandardnému krmivu ([Davis et al., 2008] a [Geiger et al., 2009]). Dôležité je, že jedlo štandardného krmiva postačovalo na zvýšenie hladín dopamínu v NAc kontrolných potkanov, zatiaľ čo iba vysoko chutné potraviny boli dostatočné na vyvolanie uvoľňovania akumulovaného dopamínu u obéznych potkanov, ktoré mali v minulosti rozšírený prístup k chutným potravinám. (Geiger a kol., 2009). Tieto zistenia ukazujú, že vývoj obezity u potkanov je spojený s dysfunkciou pri mesostriatálnom dopamínovom prenose, najvýraznejšie pri striatálnych D2Rs, a že nedostatočná signalizácia D2R prispieva k vzniku deficitu odmien počas vývoja obezity u potkanov. To je v súlade so skutočnosťou, že downregulácia striatálnych D2R je pozoruhodnou neuroadaptívnou odpoveďou na prírastok hmotnosti u ľudí ([Barnard et al., 2009], [Stice et al., 2008a] a [Wang et al., 2001]), a že deficientná striatálna D2R signalizácia môže tupnúť striatálne reakcie na hedonické potraviny u ľudských subjektov, čím predisponuje jedinca k budúcemu prírastku hmotnosti ([Stice a kol., 2008a] a [Wang et al., 2001]).
Obrázok 5. Dopamínové receptory D2, dysfunkcia odmien a kompulzivita pri obezite - Zníženie hladiny dopamínových D2 receptorov (D2R) v striate potkana urýchľuje vznik dysfunkcie odmien a nutkavého stravovania u potkanov s rozšíreným prístupom k chutnej strave..
Nedostatočná signalizácia D2R môže spúšťať kompulzívne stravovanie v obezite
Obezita sa vyznačuje prejedaním, ktoré pretrváva aj napriek vyjadrenej túžbe obmedziť spotrebu a vedomosti o hlbokých negatívnych zdravotných a sociálnych dôsledkoch nepretržitej nadmernej konzumácie (Booth a kol., 2008), [Delin et al., 1997] a [Puhl et al. al., 2008]). Príkladom toho je skutočnosť, že mnohí obézni pacienti podstúpia potenciálne nebezpečnú bariatrickú operáciu (žalúdočného bypassu) na kontrolu svojej hmotnosti (Yurcisin et al., 2009), ale často sa vracajú k prejedaniu, aj keď chirurgický zákrok znižuje subjektívne hodnotenie hladu a znižuje počet pacientov, ktorí sú vystavení riziku. schopnosť konzumovať veľké množstvá potravín ([Kalarchian et al., 2002] a [Saunders, 2001]). Drogová závislosť je podobne definovaná ako strata inhibičnej kontroly nad konzumáciou drog a vytrvalosť v návyku napriek uvedomeniu si potenciálne zničujúcich zdravotných, sociálnych alebo finančných dôsledkov (DSM-IV; American Psychiatric Association, 1994). Obezita a drogová závislosť sa tak vyznačujú charakteristickými znakmi kompulzívnych porúch v tom, že existuje zjavné zlyhanie pri využívaní informácií o budúcich škodlivých následkoch na zmiernenie konzumácie a pretrvávania spotreby napriek dostupnosti menej škodlivého alternatívneho správania.
Kompulzívne užívanie drog bolo operatívne definované u hlodavcov ako spotreba, ktorá je odolná voči potlačeniu trestom alebo environmentálnymi stimulmi predpovedajúcimi trest ([Pelloux a kol., 2007] a [Vanderschuren a Everitt, 2004]). Obdobia rozšíreného prístupu ku kokaínu a iným drogám zneužívania môžu viesť k vzniku kompulzívneho správania užívania drog u potkanov ([Ahmed a Koob, 1998], [Deroche-Gamonet et al., 2004] a [Vanderschuren a Everitt, 2004] ). Potkany s anamnézou rozsiahlej konzumácie kokaínu skutočne vykazujú príjem, ktorý je odolný voči narušeniu averzívnym podmieňovaným stimulom (CS), ktorý predpovedá negatívny výsledok (tzn. Svetlo, ktoré predpovedá dodanie averzívneho footshocku) ([Belin et al., 2008] a [Vanderschuren a Everitt, 2004]). Naopak, ten istý averzívny CS môže hlboko znížiť odpovede na hľadanie liečiv u potkanov s relatívne obmedzeným prístupom k lieku. S ohľadom na podobnosti medzi nutkavým užívaním drog v závislosti a prejedaním sa v obezite sme nedávno skúmali, či obézni potkany konzumujú chutné jedlo kompulzívnym spôsobom a či striatálne D2R hrajú úlohu v tomto procese (Johnson a Kenny, 2010). Zistili sme, že obézne potkany s históriou rozšíreného prístupu k chutnému jedlu pokračovali v konzumácii chutného jedla aj v prítomnosti škodlivého CS (ľahká tágo), ktoré predpovedalo dodanie averzívneho šošovky (Johnson a Kenny, 2010). Na rozdiel od toho, ten istý averzívny CS narušil chutnú spotrebu potravy u chudých potkanov s veľmi obmedzenou expozíciou energeticky hustým chutným jedlám. Konzumatívna konzumácia potravín sa preto môže u obéznych potkanov stať nutkavým, podobne ako konzumácia kokaínu. V súlade s touto interpretáciou údajov, myši, ktoré predtým mali prístup k chutnej diéte s vysokým obsahom tukov, strávili viac času v averzívnom prostredí (jasne osvetlené) na získanie chutnej potravy ako myši, ktoré nemali žiadne predchádzajúce skúsenosti so stravou (Teegarden a Bale). , 2007). Kvôli strachu z predácie sú jasne osvetlené otvorené arény pre myši vysoko averzívne (Suarez a Gallup, 1981). Myši sa preto stávajú rezistentnými voči potenciálne negatívnym dôsledkom ich správania pri potravinách a budú riskovať predáciu, že získajú chutné jedlo, aj keď je k dispozícii menej chutné jedlo v oveľa nižšom nebezpečenstve.
Zaujímavosťou je, že alela A1 polymorfizmu TaqIA, ktorá má za následok zníženú hustotu striatálneho D2R (Noble, 2000) a otupenú striatálnu aktiváciu v reakcii na chutnú potravu (Stice et al., 2008a), je tiež spojená s deficitmi vo vzdelávaní, aby sa vyhlo akciám, ktoré majú negatívne dôsledky (Klein et al., 2007). Práve tento typ zlyhania využívania informácií súvisiacich s budúcimi negatívnymi dôsledkami prejedania sa môže prispieť k rozvoju nutkavého stravovania u obéznych jedincov. Zistili sme, že vznik nutkavého jedenia u potkanov s prístupom k chutnému jedlu sa dramaticky zrýchlil po striatálnom D2R knockdown (Johnson a Kenny, 2010). V skutočnosti, potkany so striatálnym D2R knockdownom, ktoré predtým mali len 14 dni rozšíreného prístupu k energeticky hustým chutným potravinám, vykazovali chutnú spotrebu potravy, ktorá bola odolná voči narušeniu averzívnym CS (Johnson a Kenny, 2010; Obrázok 5). Avšak táto 14 denná doba obmedzenej expozície chutnému jedlu nebola dostatočná na vyvolanie nutkavého jedenia u kontrolných potkanov (Johnson a Kenny, 2010). Tieto zistenia naznačujú, že u obéznych potkanov sa môže objaviť nutkavý príjem chutných potravín. Okrem toho, nedostatočná striatálna D2R signalizácia, ktorá urýchľuje vznik odmeňovanej hyposenzitivity v reakcii na chutnú nadmernú spotrebu potravín, tiež urýchľuje vznik nutkavého jedenia (obrázok 5).
Interakcie medzi D2R a hormonálnymi regulátormi energetickej rovnováhy v obezite
Exogénny leptín podávaný do VTA inhibuje mesoaccumbens dopamínový prenos a stravovacie správanie ([Hommel et al., 2006] a [Krügel et al., 2003]). Okrem svojho akútneho inhibičného účinku na dopamínové systémy stredného mozgu dochádza k hromadeniu dôkazov, že signalizácia tonického leptínu môže byť tiež potrebná na udržanie primeraných hladín mesostriatálnej dopamínovej signalizácie. Flier a kolegovia zistili, že ob / ob myši mali nižšie hladiny tyrozínhydroxylázy v dopamínových neurónoch stredného mozgu, kľúčový enzým v biosyntéze dopamínu (Fulton et al., 2006). Okrem toho, myši ob / ob znížili evolúciu uvoľneného dopamínu do NAc (Fulton et al., 2006) a znížili somatodendritické vezikulárne obchody dopamínu vo VTA a SN (Roseberry et al., 2007). Tieto nedostatky v produkcii dopamínu a mechanizme prenosu signálu u myší ob / ob boli korigované ošetrením leptínom ([Fulton et al., 2006] a [Pfaffly et al., 2010]). Leptín infundovaný len do LH bol v skutočnosti postačujúci na korekciu dysfunkčného prenosu dopamínu u myší ob / ob (Leinninger et al., 2009), čo naznačuje, že bunky exprimujúce leptínový receptor v LH pôsobia na udržanie príslušných úrovní dopamínovej signalizácie. Okrem zníženej produkcie a uvoľňovania dopamínu mali myši ob / ob tiež sklon k nižším úrovniam expresie D2R v striate (Pfaffly et al., 2010). Okrem toho liečba exogénnym leptínom v režime, ktorý vedie k rozvoju necitlivosti na signalizáciu leptínu (tj rezistencia na leptín), výrazne znížila hladiny striatálneho D2R u myší divokého typu (Pfaffly et al., 2010). Obézne potkany vyvinú rezistenciu voči leptínu vo VTA ([Matheny a kol., 2011] a [Scarpace a kol., 2010]) a tiež majú nižšie hladiny TH pri VTA, znížené uvoľňovanie dopamínu v striate a znížené hladiny striatálneho D2R (Geiger et al., 2008). Tieto údaje spoločne ukazujú, že leptín má komplexné účinky na dopamínové systémy stredného mozgu. Na jednej strane akútna aktivácia leptínových receptorov vo VTA vykazuje inhibičný účinok na mesoaccumbens dopamínový prenos a môže inhibovať stravovacie správanie ([Hommel et al., 2006] a [Krügel et al., 2003]). Na druhej strane je signalizácia leptínu v strednom mozgu nevyhnutná na udržanie primeranej produkcie dopamínu a prenosu signálu a genetické deficity v signalizácii leptínu alebo v rozvoji rezistencie leptínu v obezite hlboko narušujú dopamínové systémy mesoaccumbens. Ako taká je zaujímavá možnosť, že vývoj rezistencie leptínu v dopamínergných neurónoch stredného mozgu počas vývoja obezity môže zohrávať hlavnú úlohu v narušení striatálneho D2R signalizácie, ktorá sa javí ako hnacia sila vzniku závislosti od závislostí na odmeňovaní a kompulzívneho prejedania. (Obrázok 5) u obéznych potkanov.
Okrem komplexných účinkov signalizácie leptínu na mezostriatálny dopamínergický prenos existuje dôkaz, že D2Rs môžu následne regulovať signalizáciu leptínu. Agonista bromokriptínu D2R znižuje hladiny cirkulujúceho leptínu ([Doknic et al., 2002], [Kok a kol., 2006] a [Mastronardi et al., 2001]), čo naznačuje, že D2Rs majú inhibičný vplyv na hladiny leptínu. Okrem toho myši s nulovou mutáciou v géne D2R majú zvýšenú signalizáciu leptínu v hypotalame a sú citlivejšie na anorektické účinky leptínu (Kim et al., 2010). Je dobre známe, že hladiny leptínu sa zvyšujú počas vývoja obezity (hyperleptinémia), avšak súčasne dochádza k poklesu citlivosti na signalizáciu leptínu (tj rezistencia na leptín) (Hamilton et al., 1995). Je teda zaujímavou možnosťou, že zníženie striatálnej signalizácie D2R počas vývoja obezity môže predstavovať kompenzačnú reakciu na nadmernú spotrebu chutných potravín a prírastok hmotnosti, ktorý zvyšuje hladiny cirkulujúceho leptínu a zvyšuje jeho signálnu účinnosť na prekonanie vývoja rezistencie na leptín. Takýto účinok môže zvýšiť inhibičné účinky leptínu na striatálne systémy, ktoré regulujú citlivosť na chutnú potravu ([Farooqi et al., 2007], [Fulton et al., 2006] a [Hommel et al., 2006]), čím pôsobia na zmiernenie hedonických reakcií na chutné jedlo. Uvedenie tohto zistenia spolu s regulačnou úlohou leptínu na D2R opísaných vyššie sa zdá, že signalizácia leptínu a D2R môže byť spojená recipročným spôsobom, aby sa regulovali homeostatické a hedonické aspekty stravovacieho správania.
Dysregulované systémy stresu v mozgu v obezite
Leptínová signalizácia v strednom mozgu reguluje mesoaccumbens dopamínový prenos a citlivosť na hedonické potraviny. Avšak neuróny vo VTA, ktoré exprimujú receptory leptínu, prechádzajú riedko na NAc a namiesto toho demonštrujú výraznejšie projekcie centrálneho jadra amygdaly (CeA) (Leshan et al., 2010). V súvislosti s chovaním krmiva je známe, že CeA reguluje inhibičné účinky škodlivých environmentálnych stimulov na spotrebu potravín (Petrovich et al., 2009). Konkrétne, lézie CeA, ale nie bazolaterálna amygdala (BLA), rušia inhibičné účinky podmieneného stimulu spojeného s nohou na kŕmenie, čo naznačuje, že CeA je rozhodujúca pre inhibičnú kontrolu správania pri kŕmení v reakcii na environmentálne podnety predpovedajúce negatívny výsledok ( Petrovich a kol., 2009). Obézne potkany alebo neobézne potkany so striatálnym D2R knockdownom a prístupom k chutnému jedlu pokračujú v konzumácii chutného jedla nutkavým spôsobom v prítomnosti averzívneho CS. Tieto účinky sú nápadne podobné narušeniu reaktivity voči averzívnemu CS u potkanov poškodených CeA (Petrovich et al., 2009). Bude teda dôležité určiť, či zmeny v aktivite CeA, pravdepodobne ako dôsledok down-regulácie striatálneho D2R alebo vývoja rezistencie leptínu v strednom mozgu, prispievajú k vzniku nutkavého stravovania u obéznych potkanov.
Okrem regulácie účinkov škodlivých environmentálnych stimulov na stravovacie správanie môže CeA hrať tiež kľúčovú úlohu v hedonickom stravovaní spojenom so stresom. U ľudí stres silne motivuje výber a konzumáciu energeticky výdatného chutného jedla nezávisle od kalorickej potreby ([Gluck et al., 2004], [Kandiah et al., 2006] a [O'Connor et al., 2008]) a obezita je spojená so zvýšenou sekréciou glukokortikoidov závislou od stresu ([Björntorp a Rosmond, 2000] a [la Fleur, 2006]). Environmentálne a sociálne stresové faktory tiež indukujú hyperfágiu energeticky hustého chutného jedla u hlodavcov a opíc, pričom chutná konzumácia potravy potenciálne zmierňuje nepriaznivé účinky stresu ([Dallman et al., 2003], [Dallman et al., 2006], [Pecoraro et al. al., 2004] a [Wilson et al., 2008]). Ďalej môže liek vyvolávajúci stres yohimbín obnoviť u potkanov predtým zhasnuté chutné reakcie pri hľadaní potravy (stlačenie páky), čo je účinok oslabený antalarmínom, ktorý je antagonistom receptora na kortikotropín-1 (CRF-1) (Ghitza et al., 2006). ). Je dôležité poukázať na to, že presné účinky stresu na konzumáciu potravy u ľudí a laboratórnych zvierat závisia od povahy a rozsahu stresora, druhu potravy dostupnej na konzumáciu (chutné verzus jemné), telesnej hmotnosti a pohlavia (Dallman , 2010).
Myši s rozšíreným prístupom k chutnej diéte s vysokým obsahom tukov mali zníženú expresiu stresového hormónu CRF v CeA (Teegarden a Bale, 2007). Naopak, myši podstupujúce „abstinenčný stav“ z chutnej stravy mali zvýšenú expresiu CRF v CeA (Teegarden a Bale, 2007). Tento posledný účinok je veľmi podobný zvýšenej expresii CRF v CeA zistenej u potkanov, ktorí podstúpili vysadenie zo všetkých hlavných liekov zneužívania (Koob, 2010). Pretože ďalšie užívanie liekov môže normalizovať túto averzívnu neurobiologickú reakciu na abstinenčnú liečbu, predpokladalo sa, že hyperaktívny prenos CRF v CeA a iných limbických štruktúrach môže uľahčiť rozvoj nutkavého užívania drog (Koob a Zorrilla, 2010). V súlade s týmto názorom, myši, ktoré podstúpili vysadenie z chutných potravín s vysokou hustotou energie, ktoré mali zvýšené hladiny CRF v CeA, strávili podstatne dlhšie času v averzívnom (jasne osvetlenom) prostredí na získanie chutných potravín ako myši bez predchádzajúcich skúseností s potravou, dokonca hoci menej chutné, bolo k dispozícii v prostredí, ktoré nie je nebezpečné (tmavo osvetlené) (Teegarden a Bale, 2007). Myši sa preto stávajú rezistentnými voči potenciálne negatívnym dôsledkom ich správania pri potravinách a riskujú predátorstvo, že získajú chutné jedlo, aj keď je k dispozícii menej chutné jedlo v oveľa nižšom nebezpečenstve, čiastočne na zmiernenie prenosu CRF v CeA (Teegarden a Bale, 2007). Niekoľko ďalších dôkazov podporuje úlohu prenosu CRF v nutkavom jedení. Najmä nedávna štúdia hodnotila účinky antagonistu receptora CRF-1 R121919 na spotrebu potravy u potkanov, ktorí podstúpili cyklickú variáciu v strave, v ktorej mali prístup k štandardnému krmivu 5 dní v týždni a prístupu k chutnej cukrovej strave 2 dní v týždni. týždeň (Cottone et al., 2009). Po 7 týždňoch tejto cyklickej variácie v diéte R121919 zoslabil nadmernú konzumáciu vysoko chutnej stravy a zvýšenú spotrebu bow chow (Cottone et al., 2009). Hladiny expresie CRF v CeA sa zvýšili u cyklovaných potkanov počas vysadenia z chutnej stravy, ale po opätovnom vystavení chutnej strave sa vrátili na východiskové hladiny (Cottone et al., 2009). Nakoniec, regulácia CRF GABAergnej transmisie v CeA bola zvýšená u cyklovaných potkanov, ktorí podstúpili vysadenie z chutnej stravy v porovnaní s kontrolnými potkanmi, ktoré predtým mali prístup iba k bow chow, čo sa odráža v skutočnosti, že R121919 narušil evolučný GABAergický inhibičný postsynaptický potenciál v Rezy CeA z cyklovaných potkanov v koncentrácii, ktorá nemenila prenos v rezoch kontrolných potkanov (Cottone et al., 2009). Zaujímavé je, že podobný účinok CRF na GABAergný prenos v CeA bol pozorovaný aj u potkanov, ktorí podstúpili vysadenie z chronickej expozície etanolu (Roberto et al., 2010). Nakoniec, CeA, jadro postele stria terminalis (BNST) a NAc shell tvoria väčšiu súvislú štruktúru nazývanú „rozšírená amygdala“. Infúzia CRF do škrupiny NAc na miestach, ktoré stimulujú hedonické jedenie, zvyšuje motivačnú dôležitosť environmentálneho tá, ktorá bola predtým spárovaná s dostupnosťou chutných potravín (Peciña et al., 2006a). Stres môže preto zvýrazniť význam environmentálnych podnetov spojených s potravinami moduláciou aktivity NAc shell neurónov. Celkovo vzaté, tieto zistenia naznačujú, že nadmerná konzumácia chutných potravín alebo návykových látok môže vyvolať podobné neuroadaptívne odozvy v extrahypothalamických stresových dráhach v mozgu, čo môže prispieť k nutkavému spotrebnému správaniu.
Závery
V poslednej dobe sa dosiahol značný pokrok v identifikácii mozgových systémov zapojených do hedonických účinkov chutného jedla a úprav, ktoré sa vyskytujú v týchto obvodoch v reakcii na nadmernú konzumáciu chutného jedla a prírastok hmotnosti. Je zarážajúce, že podobné mozgové systémy a bežné adaptívne reakcie sa spúšťajú v reakcii na konzumáciu chutných jedál a návykových drog. Najmä nadmerná konzumácia chutných jedál alebo zneužívaných drog spôsobuje podobné deficity v reakcii systémov odmeňovania mozgu. Chutné jedlo a návykové lieky indukujú deficity v striatálnom prenose dopamínu a expresii striatálnych D2R. Chutné jedlo a návykové lieky tiež spúšťajú u hlodavcov vznik nutkavého konzumného správania a vyvolávajú extrahypotalamické stresové reakcie. Nakoniec, bežné genetické chyby v systémoch odmeňovania mozgu môžu predisponovať jednotlivcov k prejedaniu a obezite a tiež k užívaniu drog a závislosti. Na základe týchto a ďalších podobností medzi obezitou a drogovou závislosťou sa v skutočnosti tvrdilo, že obezita by mala byť zahrnutá ako diagnostická kategória do pripravovaného DSM-V ([Devlin, 2007] a [Volkow a O'Brien, 2007]. ). Z tohto hľadiska budú kritické oblasti pre budúci výskum zahŕňať ďalšie skúmanie potenciálnych neurobiologických presahov medzi nutkavým prejedaním a užívaním drog. Bude napríklad dôležité otestovať, či obezita súvisí s vývojom návykového konzumného správania, ktorý je výsledkom plastickosti dorzálneho striata, rovnako ako závislosť od drog môže súvisieť s prestavovaním striata a vznikom návykových drog. správania ([Everitt a Robbins, 2005], [Hollander et al., 2010] a [Kasanetz et al., 2010]). Kortikálne oblasti mozgu zapojené do výkonnej kontroly a rozhodovania (tj. Prefrontálna kôra) a do interoceptívneho spracovania (ostrovná kôra) boli tiež výrazne zapojené do drogovej závislosti ([Everitt et al., 2008], [Fineberg et al., 2010 ], [Koob a Volkow, 2010] a [Naqvi a Bechara, 2009]), o ich presnej úlohe pri kompulzívnom stravovaní a obezite je však zatiaľ známe len málo ([Nair et al., 2011] a [Volkow et al. , 2009]). Spolu preskúmané údaje podporujú názor, že obezita a drogová závislosť môžu vzniknúť z podobných neuroadaptívnych reakcií v obvodoch odmeňovania mozgu, a naznačujú, že známe mechanizmy závislosti môžu poskytnúť heuristický rámec na pochopenie obezity.
Poďakovanie
Autor je podporovaný grantmi z Národného ústavu pre drogovú závislosť (NIDA). Autor je vďačný Paulovi Johnsonovi a Christie Fowlerovej za cenné postrehy a komentáre k rukopisu. Toto je rukopisné číslo 21042 od Výskumného ústavu Scripps.
Referencie
1.
o Abizaid a kol., 2006a
o A. Abizaid, Q. Gao, TL Horváth
o Myšlienky pre potraviny: Mechanizmus mozgu a rovnováha periférnej energie
o Neuron, 51 (2006), s. 691 – 702
o
2.
o Abizaid a kol., 2006b
o A. Abizaid, ZW Liu, ZB Andrews, M. Shanabrough, E. Borok, JD Elsworth, RH Roth, MW Sleeman, MR Picciotto, MH Tschöp et al.
o Ghrelin moduluje aktivitu a synaptickú vstupnú organizáciu dopamínových neurónov stredného mozgu a zároveň podporuje chuť do jedla
o J. Clin. Invest., 116 (2006), str. 3229 – 3239
o
3.
o Ahmed a Koob, 1998
o SH Ahmed, GF Koob
o Prechod zo stredného na nadmerné užívanie drog: zmena v hédonickej hodnote
o Veda, 282 (1998), s. 298 – 300
o
4.
o Ahmed a Koob, 2005
o SH Ahmed, GF Koob
o Prechod na závislosť od drog: negatívny model posilnenia založený na alostatickom poklese funkcie odmeňovania
o Psychofarmakológia (Berl.), 180 (2005), str. 473 – 490
o
5.
o Ahmed a kol., 2002
o SH Ahmed, PJ Kenny, GF Koob, A. Markou
o Neurobiologický dôkaz hedonickej allostázy spojenej s eskalujúcim užívaním kokaínu
o Nat. Neurosci., 5 (2002), pp. 625-626
o
6.
o Allison a kol., 1999
o DB Allison, KR Fontaine, JE Manson, J. Stevens, TB VanItallie
o Ročné úmrtia spôsobené obezitou v Spojených štátoch
o JAMA, 282 (1999), s. 1530 – 1538
o
7.
o Americká psychiatrická asociácia, 1994
o Americká psychiatrická asociácia
o Diagnostický a štatistický manuál duševných porúch
(štvrté vydanie) Americká psychiatrická asociácia, Washington, DC (1994)
o
8.
o Angeles-Castellanos et al., 2007
o M. Angeles-Castellanos, J. Mendoza, C. Escobar
o Obmedzené kŕmenie plánuje denné rytmy fázového posunu c-Fos a imunoreaktivity proteínu Per1 v kortikolimbických oblastiach u potkanov
o Neuroscience, 144 (2007), str. 344 – 355
o
9.
o Baicy a kol., 2007
o K. Baicy, ED Londýn, J. Monterosso, ML Wong, T. Delibasi, A. Sharma, J. Licinio
o Leptínová náhrada mení reakciu mozgu na potravinové podnety u dospelých s nedostatkom geneticky leptínu
o Proc. Natl. Acad. Sci. USA, 104 (2007), s. 18276 – 18279
o
10.
o Baldo a kol., 2004
BA Baldo, L. Gual-Bonilla, K. Sijapati, RA Daniel, CF Landry, AE Kelley
o Aktivácia subpopulácie hypotalamických neurónov obsahujúcich orexín / hypokretín pomocou inhibície jadra accumbens sprostredkovanej receptorom GABAA, ale nie vystavením novému prostrediu
o Eur. J. Neurosci., 19 (2004), str. 376-386
o
11.
o Balleine a Dickinson, 2000
o BW Balleine, A. Dickinson
o Vplyv lézií ostrovného kortexu na inštrumentálne kondicionovanie: dôkaz pre úlohu v stimulačnej pamäti
o J. Neurosci., 20 (2000), str. 8954-8964
o
12.
o Barnard a kol., 2009
o ND Barnard, EP Noble, T. Ritchie, J. Cohen, DJ Jenkins, G. Turner-McGrievy, L. Gloede, AA Green, H. Ferdowsian
o D2 dopamínový receptor Taq1A polymorfizmus, telesná hmotnosť a príjem potravy u diabetikov typu 2
o Výživa, 25 (2009), s. 58 – 65
o
13.
o Basso a Kelley, 1999
o AM Basso, AE Kelley
o Kŕmenie indukované stimuláciou receptora GABA (A) v jadre nucleus accumbens: Regionálne mapovanie a charakterizácia makronutrientov a chuťových preferencií
o Behav. Neurosci., 113 (1999), pp. 324-336
o
14.
o Batterham a kol., 2007
o RL Batterham, DH ffytche, JM Rosenthal, FO Zelaya, GJ Barker, DJ Withers, SC Williams
o PYY modulácia kortikálnych a hypotalamických oblastí mozgu predpovedá stravovacie správanie u ľudí
o Príroda, 450 (2007), s. 106 – 109
o
15.
o Baxter a Murray, 2002
o MG Baxter, EA Murray
o Amygdala a odmena
o Nat. Neurosci., 3 (2002), str. 563-573
o
16.
o Bean a kol., 2008
o MK Bean, K. Stewart, ME Olbrisch
o Obezita v Amerike: dôsledky pre klinických a zdravotných psychológov
o J. Clin. Psychol. Med. Nastavenia, 15 (2008), str. 214 – 224
o
17.
o Beaver a kol., 2006
o JD Beaver, AD Lawrence, J. van Ditzhuijzen, MH Davis, A. Woods, AJ Calder
o Jednotlivé rozdiely v odmeňovaní vedú k predpovedaniu nervových reakcií na obrazy potravín
o J. Neurosci., 26 (2006), str. 5160-5166
o
18.
o Belin a kol., 2008
o D. Belin, AC Mar, JW Dalley, TW Robbins, BJ Everitt
• Vysoká impulzivita predpovedá prechod na kompulzívne užívanie kokaínu
o Veda, 320 (2008), s. 1352 – 1355
o
19.
o Berridge, 1996
o KC Berridge
o Potravinová odmena: Mozgové substráty chcieť a páčiť sa
o Neurosci. Biobehav. Rev., 20 (1996), str. 1 – 25
o
20.
o Berridge, 2009
o KC Berridge
o 'Liking' a 'chcú' potravinové odmeny: Substráty mozgu a úlohy v poruchách príjmu potravy
Fyziol. Behav., 97 (2009), str. 537 – 550
o
21.
o Björntorp a Rosmond, 2000
P. Björntorp, R. Rosmond
o Obezita a kortizol
o Výživa, 16 (2000), s. 924 – 936
o
22.
o Blundell a Herberg, 1968
o JE Blundell, LJ Herberg
o Relatívne účinky nutričného deficitu a obdobia deprivácie na rýchlosť elektrickej sebestačnosti laterálneho hypotalamu
o Príroda, 219 (1968), s. 627 – 628
o
23.
o Booth a kol., 2008
o ML Booth, RL Wilkenfeld, DL Pagnini, SL Booth, LA King
o Vnímanie adolescentov na nadváhu a obezitu: váha štúdie verejnej mienky
o J. Paediatr. Zdravie detí, 44 (2008), s. 248 – 252
o
24.
o Bragulat a kol., 2010
o V. Bragulat, M. Dzemidzic, C. Bruno, CA Cox, T. Talavage, RV Considine, DA Kareken
o Potravinársky príbuzné zápachové sondy Brain Reward Circuits počas hladu: Pilot fMRI Study
o Obezita, Silver Spring, MD (2010)
o
25.
o Cabanac a Johnson, 1983
M. Cabanac, KG Johnson
o Analýza konfliktu medzi chutnosťou a expozíciou za studena u potkanov
Fyziol. Behav., 31 (1983), str. 249 – 253
o
26.
o Campfield a kol., 1995
o LA Campfield, FJ Smith, Y. Guisez, R. Devos, P. Burn
o Rekombinantný myšací OB proteín: dôkaz periférneho signálu spájajúceho adipozitu a centrálne neurónové siete
o Veda, 269 (1995), s. 546 – 549
o
27.
o Cannon a Palmiter, 2003
o CM Cannon, RD Palmiter
o Odmena bez dopamínu
o J. Neurosci., 23 (2003), str. 10827-10831
o
28.
o Carr a Simon, 1984
o KD Carr, EJ Simon
o Sprostredkovanie opioidov je potencovaním odmeňovania hladom
o Brain Res., 297 (1984), str. 369 – 373
o
29.
o Centrá pre kontrolu a prevenciu chorôb, 2009
o Centrá pre kontrolu a prevenciu chorôb (2009). Trendy obezity v USA (Atlanta: Centrá pre kontrolu a prevenciu chorôb).
o
30.
o Colantuoni a kol., 2001
o C. Colantuoni, J. Schwenker, J. McCarthy, P. Rada, B. Ladenheim, JL Cadet, GJ Schwartz, TH Moran, BG Hoebel
o Nadmerný príjem cukru mení väzbu na dopamín a mu-opioidné receptory v mozgu
o Neuroreport, 12 (2001), str. 3549 – 3552
o
31.
o Cornelius a kol., 2010
o JR Cornelius, M. Tippmann-Peikert, NL Slocumb, CF Frerichs, MH Silber
Poruchy kontroly impulzov s použitím dopaminergných látok pri syndróme nepokojných nôh: Štúdia kontroly prípadov
o Spánok, 33 (2010), str. 81 – 87
o
32.
o Cornier a kol., 2009
o MA Cornier, AK Salzberg, DC Endly, DH Bessesen, DC Rojas, JR Tregellas
o Účinky prekrmovania na neuronálnu odozvu na vizuálne potravinové podnety u jedincov so zníženou obezitou
o PLoS ONE, 4 (2009), str. e6310 http://dx.doi.org/10.1371/journal.pone.0006310
o
33.
o Cottone a kol., 2009
o P. Cottone, V. Sabino, M. Roberto, M. Bajo, L. Pockros, JB Frihauf, EM Fekete, L. Steardo, KC Rice, DE Grigoriadis et al.
o Nábor systému CRF sprostredkováva temnú stránku nutkavého jedenia
o Proc. Natl. Acad. Sci. USA, 106 (2009), s. 20016 – 20020
o
34.
o Dagher, 2009
o A. Dagher
o Neurobiológia apetítu: Hlad ako závislosť
o Int. J. Obes. (Lond.), 33 (Suppl 2) (2009), s. S30 – S33
o
35.
o Dagher a Robbins, 2009
o A. Dagher, TW Robbins
o Osobnosť, závislosť, dopamín: Informácie o Parkinsonovej chorobe
o Neuron, 61 (2009), s. 502 – 510
o
36.
o Dallman, 2010
o MF Dallman
o Obezita vyvolaná stresom a emocionálny nervový systém
Trends Endokrinol. Metab., 21 (2010), pp. 159 – 165
o
37.
o Dallman a kol., 2003
o MF Dallman, N. Pecoraro, SF Akana, SE La Fleur, F. Gomez, H. Houshyar, ME Bell, S. Bhatnagar, KD Laugero, S. Manalo
o Chronický stres a obezita: nový pohľad na „komfortné jedlo“
o Proc. Natl. Acad. Sci. USA, 100 (2003), s. 11696 – 11701
o
38.
o Dallman a kol., 2006
o MF Dallman, NC Pecoraro, SE La Fleur, JP Warne, AB Ginsberg, SF Akana, KC Laugero, H. Houshyar, AM Strack, S. Bhatnagar, ME Bell
o Glukokortikoidy, chronický stres a obezita
o Prog. Brain Res., 153 (2006), str. 75 – 105
o
39.
o Davis a kol., 2004
C. Davis, S. Strachan, M. Berkson
o Citlivosť na odmenu: Dôsledky pre prejedanie a nadváhu
o Appetite, 42 (2004), str. 131 – 138
o
40.
o Davis a kol., 2008
o JF Davis, AL Tracy, JD Schurdak, MH Tschöp, JW Lipton, DJ Clegg, SC Benoit
o Expozícia zvýšeným hladinám tuku v strave zmierňuje psychostimulačnú odmenu a mesolimbický obrat dopamínu u potkanov
o Behav. Neurosci., 122 (2008), pp. 1257-1263
o
41.
o de Araujo et al., 2010
o IE de Araujo, X. Ren, JG Ferreira
o Metabolické snímanie v dopamínových systémoch mozgu
o Výsledky Probl. Cell Differ., 52 (2010), pp. 69 – 86
o
42.
o Delin a kol., 1997
o CR Delin, JM Watts, JL Saebel, PG Anderson
o Správanie sa v jedle a skúsenosti s hladom po operácii žalúdočného bypassu pre morbidnú obezitu
o Obes. Surg., 7 (1997), s. 405 – 413
o
43.
o Deroche-Gamonet a kol., 2004
o V. Deroche-Gamonet, D. Belin, PV Piazza
o Dôkaz o návykovom správaní u potkanov
o Veda, 305 (2004), s. 1014 – 1017
o
44.
o Devlin, 2007
o MJ Devlin
o Existuje miesto pre obezitu v DSM-V?
o Int. J. Jedz. Disord., 40 (Suppl) (2007), s. S83 – S88
o
45.
o Doknic et al., 2002
M. Doknic, S. Pekič, M. Zarkovič, M. Medic-Stojanoska, C. Dieguez, F. Casanueva, V. Popovič
o Dopaminergný tón a obezita: pohľad z prolaktinómov liečených bromokriptínom
o Eur. J. Endocrinol., 147 (2002), pp. 77-84
o
46.
o Everitt a Robbins, 2005
o BJ Everitt, TW Robbins
o Neurónové systémy posilňovania drogovej závislosti: Od činov k návykom k donucovaniu
o Nat. Neurosci., 8 (2005), pp. 1481-1489
o
47.
o Everitt a kol., 2008
o BJ Everitt, D. Belin, D. Economidou, Y. Pelloux, JW Dalley, TW Robbins
o Preskúmanie. Neurálne mechanizmy, ktoré sú základom zraniteľnosti pri rozvoji kompulzívnych návykov a návykov pri hľadaní drog
o Philos. Trans. R. Soc. Londa. B Biol. Sci., 363 (2008), str. 3125 – 3135
o
48.
o Farooqi a kol., 2007
o IS Farooqi, E. Bullmore, J. Keogh, J. Gillard, S. O'Rahilly, PC Fletcher
o Leptín reguluje striatálne oblasti a stravovacie správanie ľudí
o Veda, 317 (2007), s. 1355
o
49.
o Felsted a kol., 2010
o JA Felsted, X. Ren, F. Chouinard-Decorte, DM Small
o Geneticky stanovené rozdiely v reakcii mozgu na primárnu potravinovú odmenu
o J. Neurosci., 30 (2010), str. 2428-2432
o
50.
Figlewicz a kol., 2001
o DP Figlewicz, MS Higgins, SB Ng-Evans, PJ Havel
o Leptín zvracia preferenciu miesta so sacharózou, ktorá je podmienená u potkanov s obmedzeným príjmom potravín
Fyziol. Behav., 73 (2001), str. 229 – 234
o
51.
Figlewicz a kol., 2003
o DP Figlewicz, SB Evans, J. Murphy, M. Hoen, DG Baskin
o Expresia receptorov pre inzulín a leptín vo ventrálnej tegmentálnej oblasti / substantia nigra (VTA / SN) potkana
o Brain Res., 964 (2003), str. 107 – 115
o
52.
o Fineberg et al., 2010
NA Fineberg, MN Potenza, SR Chamberlain, HA Berlin, L. Menzies, A. Bechara, BJ Sahakian, TW Robbins, ET Bullmore, E. Hollander
o Sondážne a impulzívne správanie, od zvieracích modelov až po endofenotypy
o Neuropsychofarmakológia, 35 (2010), pp. 591 – 604
o
53.
o Finkelstein et al., 2005
o EA Finkelstein, CJ Ruhm, KM Kosa
o Ekonomické príčiny a následky obezity
o Annu. Verejné zdravie, 26 (2005), s. 239 – 257
o
54.
o Flegal et al., 2010
o KM Flegal, MD Carroll, CL Ogden, LR Curtin
o Prevalencia a trendy obezity u dospelých v USA, 1999-2008
o JAMA, 303 (2010), s. 235 – 241
o
55.
o Foo a Mason, 2005
o H. Foo, P. Mason
o Senzorické potlačenie počas kŕmenia
o Proc. Natl. Acad. Sci. USA, 102 (2005), s. 16865 – 16869
o
56.
o Franken a Muris, 2005
o IH Franken, P. Muris
o Individuálne rozdiely v citlivosti na odmeňovanie súvisia s túžbou po potravinách a relatívnou telesnou hmotnosťou u zdravých žien
o Appetite, 45 (2005), str. 198 – 201
o
57.
o Friedman a kol., 2011
A. Friedman, E. Lax, Y. Dikshtein, L. Abraham, Y. Flaumenhaft, E. Sudai, M. Ben-Tzion, G. Yadid
o Elektrická stimulácia laterálneho habenula vyvoláva inhibičný účinok na samopodávanie sacharózy
o Neurofarmakológia, 60 (2011), pp. 381 – 387
o
58.
o Fulton et al., 2000
S. Fulton, B. Woodside, P. Shizgal
o Modulácia obvodov odmeňovania mozgu leptínom
o Veda, 287 (2000), s. 125 – 128
o
59.
o Fulton et al., 2006
S. Fulton, P. Pissios, RP Manchon, L. Stiles, L. Frank, EN Pothos, E. Maratos-Flier, JS Flier
o Leptínová regulácia dopamínovej cesty mezoaccumbens
o Neuron, 51 (2006), s. 811 – 822
o
60.
o Gao a Horvath, 2007
o Q. Gao, TL Horváth
o Neurobiológia krmenia a výdaj energie
o Annu. Neurosci., 30 (2007), str. 367-398
o
61.
o Gautier et al., 2000
o JF Gautier, K. Chen, AD Salbe, D. Bandy, RE Pratley, M. Heiman, E. Ravussin, EM Reiman, PA Tataranni
o Diferenciálne reakcie mozgu na nasýtenie u obéznych a chudých mužov
o Diabetes, 49 (2000), pp. 838 – 846
o
62.
o Geiger a kol., 2008
o BM Geiger, GG Behr, LE Frank, AD Caldera-Siu, MC Beinfeld, EG Kokkotou, EN Pothos
o Dôkazy o chybnej exolektóze mezolimbického dopamínu u potkanov náchylných na obezitu
o FASEB J., 22 (2008), s. 2740 – 2746
o
63.
o Geiger a kol., 2009
o BM Geiger, M. Haburcak, NM Avena, MC Moyer, BG Hoebel, EN Pothos
o Deficity mesolimbickej dopamínovej neurotransmisie u potkanovej obezity
o Neuroscience, 159 (2009), str. 1193 – 1199
o
64.
o Ghitza a kol., 2006
o UE Ghitza, SM Gray, DH Epstein, KC Rice, Y. Shaham
o Neuropsychofarmakológia
o Anxiogénne liečivo yohimbín obnovuje chutné potravinové hľadisko v modeli relapsu potkanov: Úloha receptorov CRF (1), 33 (2006), pp. 2188 – 2196
o
65.
o Gluck a kol., 2004
o ME Gluck, A. Geliebter, J. Hung, E. Yahav
o Kortizol, hlad a túžba po prejedaní jedla po studenom stresovom teste u obéznych žien s poruchou prejedania sa
o Psychosom. X., 66 (2004), str. 876-881
o
66.
o Goldstone a kol., 2009
o AP Goldstone, CG Prechtl de Hernandez, JD Beaver, K. Muhammed, C. Croese, G. Bell, G. Durighel, E. Hughes, AD Waldman, G. Frost, JD Bell
o Pôstové predsudky systémy odmeňovania mozgov voči vysokokalorickým potravinám
o Eur. J. Neurosci., 30 (2009), str. 1625-1635
o
67.
o Halaas et al., 1995
o JL Halaas, KS Gajiwala, M. Maffei, SL Cohen, BT Chait, D. Rabinowitz, RL Lallone, SK Burley, JM Friedman
o Účinky znižovania hmotnosti plazmatického proteínu kódovaného obéznym génom
o Veda, 269 (1995), s. 543 – 546
o
68.
o Hamilton a kol., 1995
o BS Hamilton, D. Paglia, AY Kwan, M. Deitel
o Zvýšená obézna expresia mRNA v omentálnych tukových bunkách z masívne obéznych ľudí
o Nat. X., 1 (1995), str. 953-956
o
69.
o Hernandez a Hoebel, 1988
o L. Hernandez, BG Hoebel
o Potravinová odmena a kokaín zvyšujú extracelulárny dopamín v nucleus accumbens, merané mikrodialýzou
o Life Sci., 42 (1988), s. 1705 – 1712
o
70.
o Hill et al., 2003
o JO Hill, HR Wyatt, GW Reed, JC Peters
o Obezita a životné prostredie: Odkiaľ ideme?
o Veda, 299 (2003), s. 853 – 855
o
71.
o Hoebel, 1969
o BG Hoebel
o Kŕmenie a samo-stimulácia
o Ann. NY Acad. Sci., 157 (1969), str. 758 – 778
o
72.
o Hoebel a Balagura, 1967
o BG Hoebel, S. Balagura
o Vlastná stimulácia laterálneho hypotalamu modifikovaného inzulínom a glukagónom
Fyziol. Behav., 2 (1967), str. 337 – 340
o
73.
o Hoebel a Teitelbaum, 1962
o BG Hoebel, P. Teitelbaum
o Hypotalamická kontrola kŕmenia a samo-stimulácie
o Veda, 135 (1962), s. 375 – 377
o
74.
o Hoebel a Thompson, 1969
o BG Hoebel, RD Thompson
o Averzia k laterálnej hypotalamickej stimulácii spôsobenej intragastrickým kŕmením alebo obezitou
o J. Comp. Physiol. Psychol., 68 (1969), s. 536 – 543
o
75.
o Hofmann a kol., 2010
o W. Hofmanna, GM van Koningsbruggena, W. Stroebe, S. Ramanathan, H. Aarts
o Ako sa rozvíja radosť: Hedonické reakcie na lákavé jedlo
o Psychol. Sci., 21 (2010), str. 1863 – 1870
o
76.
o Holandsko a Gallagher, 2004
o PC Holland, M. Gallagher
o Amygdala-frontálne interakcie a očakávaná odmena
o Curr. Opin. Neurobiol., 14 (2004), pp. 148-155
o
77.
o Hollander a kol., 2010
o JA Hollander, HI Im, AL Amelio, J. Kocerha, P. Bali, Q. Lu, D. Willoughby, C. Wahlestedt, MD Conkright, PJ Kenny
o Striatálna mikroRNA kontroluje príjem kokaínu prostredníctvom signalizácie CREB
o Príroda, 466 (2010), s. 197 – 202
o
78.
o Hommel et al., 2006
o JD Hommel, R. Trinko, RM Sears, D. Georgescu, ZW Liu, XB Gao, JJ Thurmon, M. Marinelli, RJ DiLeone
Signalizácia receptora leptínu v dopamínových neurónoch stredného mozgu reguluje kŕmenie
o Neuron, 51 (2006), s. 801 – 810
o
79.
Imaizumi a kol., 2001
M. Imaizumi, M. Takeda, A. Suzuki, S. Sawano, T. Fushiki
Preferencia pre potraviny s vysokým obsahom tuku u myší: Zemiaky v porovnaní s varenými zemiakmi
o Appetite, 36 (2001), str. 237 – 238
o
80.
o Jerlhag et al., 2006
o E. Jerlhag, E. Egecioglu, SL Dickson, M. Andersson, L. Svensson, JA Engel
o Ghrelín stimuluje lokomotorickú aktivitu a akumuláciu dopamínu cez centrálne cholinergné systémy u myší: Dôsledky pre jeho zapojenie do odmeňovania mozgu
o Narkoman. Biol., 11 (2006), str. 45 – 54
o
81.
o Jerlhag et al., 2007
o E. Jerlhag, E. Egecioglu, SL Dickson, A. Douhan, L. Svensson, JA Engel
o Podávanie ghrelínu do tegmentálnych oblastí stimuluje pohybovú aktivitu a zvyšuje extracelulárnu koncentráciu dopamínu v nucleus accumbens
o Narkoman. Biol., 12 (2007), str. 6 – 16
o
82.
o Jhou a kol., 2009
o TC Jhou, HL Fields, MG Baxter, CB Saper, PC Holland
o Rastromediálne tegmentálne jadro (RMTg), GABAergný aferentný k dopamínovým neurónom stredného mozgu, kóduje averzívne stimuly a inhibuje motorické reakcie
o Neuron, 61 (2009), s. 786 – 800
o
83.
o Johnson a Kenny, 2010
o PM Johnson, PJ Kenny
o Dopamínové D2 receptory pri dysfunkcii závislej na odmeňovaní a nutkavom jedení u obéznych potkanov
o Nat. Neurosci., 13 (2010), pp. 635-641
o
84.
o Johnson a kol., 1996
o PI Johnson, MA Parente, JR Stellar
o NMDA-indukované lézie jadra accumbens alebo ventrálna palidum zvyšujú odmeňovanie potravy odobratých potkanov
o Brain Res., 722 (1996), str. 109 – 117
o
85.
o Jönsson et al., 1999
o EG Jönsson, MM Nöthen, F. Grünhage, L. Farde, Y. Nakashima, P. Propping, GC Sedvall
o Polymorfizmy v dopamínovom D2 receptorovom géne a ich vzťahy k hustote striatálneho dopamínového receptora zdravých dobrovoľníkov
Mol. Psychiatria, 4 (1999), s. 290 – 296
o
86.
o Kalarchian a kol., 2002
o MA Kalarchian, MD Marcus, GT Wilson, EW Labouvie, RE Brolin, LB LaMarca
o Nadmerné stravovanie u pacientov s žalúdočným bypassom pri dlhodobom sledovaní
o Obes. Surg., 12 (2002), s. 270 – 275
o
87.
o Kandiah a kol., 2006
o J. Kandiah, M. Yake, J. Jones, M. Meyer
o Stres ovplyvňuje chuť k jedlu a pohodlie potravín preferencie v škole žien
o Nutr. Res., 26 (2006), str. 118 – 123
o
88.
o Karhunen a kol., 1997
o LJ Karhunen, RI Lappalainen, EJ Vanninen, JT Kuikka, MI Uusitupa
o Regionálny prietok krvi mozgom počas vystavenia jedlu obéznym a normálnym ženám
o Brain, 120 (1997), str. 1675 – 1684
o
89.
o Kasanetz et al., 2010
F. Kasanetz, V. Deroche-Gamonet, N. Berson, E. Balado, M. Lafourcade, O. Manzoni, PV Piazza
o Prechod na závislosť je spojený s pretrvávajúcim zhoršením synaptickej plasticity
o Veda, 328 (2010), s. 1709 – 1712
o
90.
o Kelley a kol., 1996
o AE Kelley, EP Bless, CJ Swanson
o Skúmanie účinkov antagonistov opiátov infundovaných do nucleus accumbens na kŕmenie a pitie sacharózy u potkanov
J. Pharmacol. Exp. Ther., 278 (1996), s. 1499 – 1507
o
91.
o Kelley a kol., 2005
o AE Kelley, BA Baldo, WE Pratt, MJ Will
o Kortikostriatálne-hypotalamické obvody a potravinová motivácia: integrácia energie, pôsobenia a odmeny
Fyziol. Behav., 86 (2005), str. 773 – 795
o
92.
o Kenny a kol., 2006
o PJ Kenny, SA Chen, O. Kitamura, A. Markou, GF Koob
o Kondicionovaný odber poháňa spotrebu heroínu a znižuje citlivosť na odmenu
o J. Neurosci., 26 (2006), str. 5894-5900
o
93.
Kim a kol., 2010
o KS Kim, YR Yoon, HJ Lee, S. Yoon, SY Kim, SW Shin, JJ An, MS Kim, SY Choi, W. Sun, JH Baik
o Zvýšená signalizácia hypotalamického leptínu u myší bez receptorov dopamínu D2
J. Biol. Chem., 285 (2010), str. 8905-8917
o
94.
o Klein et al., 2007
o TA Klein, J. Neumann, M. Reuter, J. Hennig, DY von Cramon, M. Ullsperger
o Geneticky stanovené rozdiely v učení sa z chýb
o Veda, 318 (2007), s. 1642 – 1645
o
95.
o Kojima a kol., 1999
M. Kojima, H. Hosoda, Y. Date, M. Nakazato, H. Matsuo, K. Kangawa
o Ghrelín je acylovaný peptid uvoľňujúci rastový hormón zo žalúdka
o Príroda, 402 (1999), s. 656 – 660
o
96.
Kok a kol., 2006
o P. Kok, F. Roelfsema, M. Frölich, J. van Pelt, AE Meinders, H. Pijl
o Aktivácia dopamínových D2 receptorov znižuje cirkadiánne koncentrácie leptínu u obéznych žien
o J. Clin. Endocrinol. Metab., 91 (2006), pp. 3236 – 3240
o
97.
o Koob, 2010
o GF Koob
o Úloha CRF a CRF-príbuzných peptidov v temnej časti závislosti
o Brain Res., 1314 (2010), str. 3 – 14
o
98.
o Koob a Le Moal, 2008
o GF Koob, M. Le Moal
o Závislosť a protivzdušný systém mozgu
o Annu. Psychol., 59 (2008), str. 29 – 53
o
99.
o Koob a Volkow, 2010
o GF Koob, ND Volkow
o Neurocircuitry závislosti
o Neuropsychofarmakológia, 35 (2010), pp. 217 – 238
o
100.
o Koob a Zorrilla, 2010
o GF Koob, EP Zorrilla
o Neurobiologické mechanizmy závislosti: zameranie na faktor uvoľňujúci kortikotropín
o Curr. Opin. Investig. Drugs, 11 (2010), s. 63 – 71
o
101.
o Kringelbach a kol., 2003
o ML Kringelbach, J. O'Doherty, ET Rolls, C. Andrews
o Aktivácia ľudského orbitofrontálneho kortexu na kvapalný potravinový stimul koreluje s jeho subjektívnou príjemnosťou
o Cereb. Cortex, 13 (2003), str. 1064 – 1071
o
102.
o Krügel et al., 2003
U. Krügel, T. Schraft, H. Kittner, W. Kiess, P. Illes
o Bazálne a kŕmenie vyvolané uvoľňovanie dopamínu v krysom jadre accumbens je deprimované leptínom
o Eur. J. Pharmacol., 482 (2003), str. 185-187
o
103.
o la Fleur, 2006
o SE la Fleur
Účinky glukokortikoidov na kŕmenie u potkanov
Fyziol. Behav., 89 (2006), str. 110 – 114
o
104.
o LaBar et al., 2001
o KS LaBar, DR Gitelman, TB Parrish, YH Kim, AC Nobre, MM Mesulam
o Hlad selektívne moduluje kortikolimickú aktiváciu na potravinové stimuly u ľudí
o Behav. Neurosci., 115 (2001), pp. 493-500
o
105.
o Latagliata a kol., 2010
o EC Latagliata, E. Patrono, S. Puglisi-Allegra, R. Ventura
o Hľadanie potravy napriek škodlivým následkom je pod kontrolou prefrontálnej kortikálnej noradrenergnej kontroly
o BMC Neurosci., 11 (2010), str. 15
o
106.
o Lawford et al., 2000
o BR Lawford, RM Young, EP Noble, J. Sargent, J. Rowell, S. Shadforth, X. Zhang, T. Ritchie
o Alela D (2) dopamínového receptora A (1) a závislosti od opioidov: asociácia s užívaním heroínu a odpoveď na liečbu metadónom
o Am. J. Med. Genet., 96 (2000), str. 592 – 598
o
107.
o Leinninger a kol., 2009
o GM Leinninger, YH Jo, RL Leshan, GW Louis, H. Yang, JG Barrera, H. Wilson, DM Opland, MA Faouzi, Y. Gong a kol.
o Leptín pôsobí prostredníctvom laterálnych hypotalamických neurónov exprimujúcich leptínový receptor na moduláciu mezolimbického dopamínového systému a potláča kŕmenie.
o Cell Metab., 10 (2009), pp. 89-98
o
108.
o Lenoir a kol., 2007
M. Lenoir, F. Serre, L. Cantin, SH Ahmed
o Intenzívna sladkosť prevyšuje odmenu kokaínu
o PLoS ONE, 2 (2007), str. e698 http://dx.doi.org/10.1371/journal.pone.0000698
o
109.
o Leshan a kol., 2010
o RL Leshan, DM Opland, GW Louis, GM Leinninger, CM Patterson, CJ Rhodes, H. Münzberg, MG Myers Jr.
o neuróny receptorov ventrálnej tegmentálnej oblasti špecificky projektujú a regulujú transkriptové neuróny regulované kokaínom a amfetamínom v rozšírenej centrálnej amygdale
o J. Neurosci., 30 (2010), str. 5713-5723
o
110.
o Louis a kol., 2010
o GW Louis, GM Leinninger, CJ Rhodes, MG Myers Jr.
Priama inervácia a modulácia neurónov orexínu laterálnymi hypotalamickými neurónmi LepRb
o J. Neurosci., 30 (2010), str. 11278-11287
o
111.
o Luppino et al., 2010
o FS Luppino, LM de Wit, PF Bouvy, T. Stijnen, P. Cuijpers, BW Penninx, FG Zitman
o Nadváha, obezita a depresia: systematický prehľad a metaanalýza dlhodobých štúdií
o Arch. Psychiatria, 67 (2010), str. 220 – 229
o
112.
o Lutter a Nestler, 2009
M. Lutter, EJ Nestler
o Homeostatické a hedonické signály ovplyvňujú reguláciu príjmu potravy
J. Nutr., 139 (2009), str. 629 – 632
o
113.
o Macht a Mueller, 2007
M. Macht, J. Mueller
o Okamžité účinky čokolády na experimentálne vyvolané stavy nálady
o Appetite, 49 (2007), str. 667 – 674
o
114.
o Maldonado-Irizarry a Kelley, 1995
o CS Maldonado-Irizarry, AE Kelley
o Excitotoxické lézie jadra a shell subregiónov nucleus accumbens odlišne narušujú reguláciu telesnej hmotnosti a motorickú aktivitu u potkanov
o Brain Res. Bull., 38 (1995), s. 551 – 559
o
115.
o Maldonado-Irizarry a kol., 1995
o CS Maldonado-Irizarry, CJ Swanson, AE Kelley
o Glutamátové receptory v jadre accumbens kontrolujú kŕmenie prostredníctvom laterálneho hypotalamu
o J. Neurosci., 15 (1995), str. 6779-6788
o
116.
o Malik a kol., 2008
S. Malik, F. McGlone, D. Bedrossian, A. Dagher
o Ghrelin moduluje mozgovú aktivitu v oblastiach, ktoré riadia chuťové správanie
o Cell Metab., 7 (2008), pp. 400-409
o
117.
o Man a kol., 2009
o MS Man, HF Clarke, AC Roberts
o Úloha orbitofrontálneho kortexu a mediálneho striata pri regulácii prepotentných reakcií na potravinové odmeny
o Cereb. Cortex, 19 (2009), str. 899 – 906
o
118.
o Margules a Olds, 1962
o DL Margules, J. Olds
o Identické systémy „kŕmenia“ a „odmeňovania“ v laterálnom hypotalame potkanov
o Veda, 135 (1962), s. 374 – 375
o
119.
o Markou a Frank, 1987
o A. Markou, RA Frank
o Účinok umiestnenia operátora a elektródy na funkcie odozvy vlaku s vlastnou stimuláciou
Fyziol. Behav., 41 (1987), str. 303 – 308
o
120.
o Markou a Koob, 1991
o A. Markou, GF Koob
o Postkokaínová anhedónia. Zvierací model stiahnutia kokaínu
o Neuropsychofarmakológia, 4 (1991), pp. 17 – 26
o
121.
o Mastronardi et al., 2001
o CA Mastronardi, WH Yu, VK Srivastava, WL Dees, SM McCann
Uvoľňovanie leptínu indukované lipopolysacharidom je nervovo kontrolované
o Proc. Natl. Acad. Sci. USA, 98 (2001), s. 14720 – 14725
o
122.
o Matheny a kol., 2011
M. Matheny, A. Shapiro, N. Tumer, PJ Scarpace
o Regionálne špecifická diéta indukovaná a leptínom indukovaná rezistencia na leptín v bunkách zahŕňa ventrálnu tegmentálnu oblasť u potkanov
o Neurofarmakológia (2011) http://dx.doi.org/10.1016/j.neuropharm.2010.11.002 v tlači. Publikované online November 5, 2010
o
123.
o Matsumoto a Hikosaka, 2007
M. Matsumoto, O. Hikosaka
o Bočný habenula ako zdroj negatívnych signálov odmeny v dopamínových neurónoch
o Príroda, 447 (2007), s. 1111 – 1115
o
124.
o Morton et al., 2006
o GJ Morton, DE Cummings, DG Baskin, GS Barsh, MW Schwartz
Kontrola centrálneho nervového systému príjmu potravy a telesnej hmotnosti
o Príroda, 443 (2006), s. 289 – 295
o
125.
o Mount a Hoebel, 1967
o G. Mount, BG Hoebel
o Samostatná bočná hypotalamická stimulácia: Samostatná prahová hodnota zvýšená príjmom potravy
o Psychon. Sci., 9 (1967), str. 265 – 266
o
126.
o Nair et al., 2011
o SG Nair, BM Navarre, C. Cifani, CL Pickens, JM Bossert, Y. Shaham
o Úloha dorzálnych mediálnych prefrontálnych kortexov dopamínových receptorov rodiny D1 pri relapsu k potravinám s vysokým obsahom tukov, ktoré sú vyvolané anxiogénnym liečivom yohimbín
o Neuropsychofarmakológia, 36 (2011), pp. 497 – 510
o
127.
Nakazato et al., 2001
M. Nakazato, N. Murakami, Y. Date, M. Kojima, H. Matsuo, K. Kangawa, S. Matsukura
o Úloha ghrelínu v centrálnej regulácii kŕmenia
o Príroda, 409 (2001), s. 194 – 198
o
128.
o Naqvi a Bechara, 2009
o NH Naqvi, A. Bechara
Skrytý ostrov závislosti: Ostrov
o Trends Neurosci., 32 (2009), str. 56 – 67
o
129.
o Neville a kol., 2004
o MJ Neville, EC Johnstone, RT Walton
o Identifikácia a charakterizácia ANKK1: Nový gén kinázy úzko spojený s DRD2 na chromozómovom pásme 11q23.1
o Hum. Mutat., 23 (2004), pp. 540 – 545
o
130.
o Nirenberg a Waters, 2006
o MJ Nirenberg, C. Waters
o Kompulzívne stravovanie a prírastok hmotnosti súvisiaci s použitím agonistu dopamínu
o Mov. Disord., 21 (2006), str. 524 – 529
o
131.
o Noble, 2000
o EP Noble
o Závislosť a jej proces odmeňovania prostredníctvom polymorfizmov génu dopamínového receptora D2: Prehľad
o Eur. Psychiatria, 15 (2000), s. 79 – 89
o
132.
o Noble a kol., 1993
o EP Noble, K. Blum, ME Khalsa, T. Ritchie, A. Montgomery, RC Wood, RJ Fitch, T. Ozkaragoz, PJ Sheridan, MUDr.
o Alelická asociácia D2 dopamínového receptorového génu so závislosťou od kokaínu
o závislosť na alkohole, 33 (1993), s. 271 – 285
o
133.
o Noble a kol., 2000
o EP Noble, X. Zhang, TL Ritchie, RS Sparkes
o Haplotypy na mieste DRD2 a ťažký alkoholizmus
o Am. J. Med. Genet., 96 (2000), str. 622 – 631
o
134.
o O'Connor a kol., 2008
o DB O'Connor, F. Jones, M. Conner, B. McMillan, E. Ferguson
Účinky denných ťažkostí a stravovacieho štýlu na stravovacie návyky
o Zdravie Psychol., 27 (1, Suppl) (2008), s. S20 – S31
o
135.
o O'Doherty a kol., 2002
o JP O'Doherty, R. Deichmann, HD Critchley, RJ Dolan
o Neurónové reakcie počas predvídania primárnej chuťovej odmeny
o Neuron, 33 (2002), s. 815 – 826
o
136.
o O'Rahilly, 2009
o S. O'Rahilly
o Ľudská genetika osvetľuje cesty metabolického ochorenia
o Príroda, 462 (2009), s. 307 – 314
o
137.
o Oswald a kol., 2010
o KD Oswald, DL Murdaugh, VL King, MM Boggiano
o Motivácia k chutnému jedlu napriek dôsledkom na zvieracom modeli nadmerného jedenia
o Int. J. Jedz. Disord. (2010) http://dx.doi.org/10.1002/eat.20808 v tlači. Publikované online Február 22, 2010
o
138.
o Park a Carr, 1998
o TH Park, KD Carr
o Neuroanatomické vzorce imunoreaktivity podobné fosfo indukovanej chutným jedlom a prostredím spárovaným s jedlom u potkanov liečených fyziologickým roztokom a naltrexonom
o Brain Res., 805 (1998), str. 169 – 180
o
139.
o Peciña a Berridge, 2005
o S. Peciña, KC Berridge
o Hedonické horúce miesto v jadre nucleus accumbens: kde mu-opioidy spôsobujú zvýšený hedonický vplyv sladkosti?
o J. Neurosci., 25 (2005), str. 11777-11786
o
140.
o Peciña a kol., 2006a
S. Peciña, J. Schulkin, KC Berridge
o Nucleus accumbens faktor uvoľňujúci kortikotropín zvyšuje motiváciu stimulovať stimuly pre sacharózu: Paradoxné pozitívne stimulačné účinky v strese?
o BMC Biol., 4 (2006), str. 8
o
141.
o Peciña a kol., 2006b
S. Peciña, KS Smith, KC Berridge
o Hedonické horúce miesta v mozgu
o Neuroscientist, 12 (2006), s. 500 – 511
o
142.
o Pecoraro et al., 2004
o N. Pecoraro, F. Reyes, F. Gomez, A. Bhargava, MF Dallman
o Chronický stres podporuje chutné kŕmenie, ktoré znižuje príznaky stresu: Forward a spätné účinky chronického stresu
o Endokrinológia, 145 (2004), s. 3754 – 3762
o
143.
o Pelchat et al., 2004
o ML Pelchat, A. Johnson, R. Chan, J. Valdez, JD Ragland
o Obrazy túžby: aktivácia túžby po potravinách počas fMRI
o Neuroimage, 23 (2004), s. 1486 – 1493
o
144.
o Pelleymounter a kol., 1995
o MA Pelleymounter, MJ Cullen, MB Baker, R. Hecht, D. Winters, T. Boone, F. Collins
Účinky obézneho génového produktu na reguláciu telesnej hmotnosti u ob / ob myší
o Veda, 269 (1995), s. 540 – 543
o
145.
o Pelloux a kol., 2007
o Y. Pelloux, BJ Everitt, A. Dickinson
o Kompulzívne hľadanie liekov u potkanov pod trestom: Vplyv histórie užívania drog
o Psychofarmakológia (Berl.), 194 (2007), str. 127 – 137
o
146.
o Perello et al., 2010
M. Perello, I. Sakata, S. Birnbaum, JC Chuang, S. Osborne-Lawrence, SA Rovinsky, J. Woloszyn, M. Yanagisawa, M. Lutter, JM Zigman
o Ghrelín zvyšuje odmeňovanie potravy s vysokým obsahom tuku v závislosti od orexínu
o Biol. Psychiatria, 67 (2010), s. 880 – 886
o
147.
o Petrovich et al., 2009
o GD Petrovich, CA Ross, P. Mody, PC Holland, M. Gallagher
o Centrálna, ale nie bazolaterálna, amygdala je rozhodujúca pre kontrolu kŕmenia pomocou averzívnych naučených podnetov
o J. Neurosci., 29 (2009), str. 15205-15212
o
148.
o Pfaffly a kol., 2010
o J. Pfaffly, M. Michaelides, GJ Wang, JE Pessin, ND Volkow, PK Thanos
o Leptín zvyšuje väzbu striatálneho dopamínového D2 receptora u obéznych (ob / ob) myší s deficitom leptínu
o Synapse, 64 (2010), str. 503 – 510
o
149.
o Puhl a kol., 2008
o RM Puhl, CA Moss-Racusin, MB Schwartz, KD Brownell
o Stigmatizácia hmotnosti a redukcia zaujatosti: Perspektívy dospelých s nadváhou a obezitou
o Zdravie Educ. Res., 23 (2008), str. 347 – 358
o
150.
o Rada a kol., 2010
o P. Rada, ME Bocarsly, JR Barson, BG Hoebel, SF Leibowitz
o Redukovaný akumulovaný dopamín u potkanov Sprague-Dawley náchylných na prejedanie sa diétou bohatou na tuky
Fyziol. Behav., 101 (2010), str. 394 – 400
o
151.
o Ritchie a Noble, 2003
o T. Ritchie, EP Noble
o Asociácia siedmich polymorfizmov génu dopamínového receptora D2 s charakteristikami viazania na mozgový receptor
o Neurochem. Res., 28 (2003), str. 73 – 82
o
152.
o Roberto a kol., 2010
o M. Roberto, MT Cruz, NW Gilpin, V. Sabino, P. Schweitzer, M. Bajo, P. Cottone, SG Madamba, DG Stouffer, EP Zorrilla a kol.
Uvoľňovanie kortikotropínu uvoľňujúceho faktoru amygdala gama-aminomaslovej kyseliny hrá kľúčovú úlohu v závislosti od alkoholu
o Biol. Psychiatria, 67 (2010), s. 831 – 839
o
153.
o Roitman a kol., 2004
o MF Roitman, GD Stuber, PE Phillips, RM Wightman, RM Carelli
o Dopamín funguje ako subsekundový modulátor vyhľadávania potravín
o J. Neurosci., 24 (2004), str. 1265-1271
o
154.
o Roitman a kol., 2008
o MF Roitman, RA Wheeler, RM Wightman, RM Carelli
o Chemické reakcie v reálnom čase v nucleus accumbens rozlišujú odmeňovanie a averzívne podnety
o Nat. Neurosci., 11 (2008), pp. 1376-1377
o
155.
o Rolls, 2008
o ET Rolls
o Funkcie orbitofrontálneho a pregenualného cingulárneho kortexu v chuti, čuchu, chuti do jedla a emóciách
o Acta Physiol. Hung., 95 (2008), s. 131 – 164
o
156.
o Rolls, 2010
o ET Rolls
o Chuť, čuchové a potravinárske spracovanie textúry spracovanie v mozgu a obezite
o Int. J. Obes. (Lond.) (2010) http://dx.doi.org/10.1038/ijo.2010.155 v tlači. Publikované online August 3, 2010
o
157.
o Rolls a kol., 1983
o ET Rolls, BJ Rolls, EA Rowe
o Senzoricky špecifická a motivačne špecifická sýtosť pre zrak a chuť jedla a vody u človeka
Fyziol. Behav., 30 (1983), str. 185 – 192
o
158.
o Roseberry a kol., 2007
o AG Roseberry, T. Painter, GP Mark, JT Williams
o Znížené vezikulárne somatodendritické zásoby dopamínu u myší s deficitom leptínu
o J. Neurosci., 27 (2007), str. 7021-7027
o
159.
o Rothemund a kol., 2007
o Y. Rothemund, C. Preuschhof, G. Bohner, HC Bauknecht, R. Klingebiel, H. Flor, BF Klapp
o Diferenciálna aktivácia dorzálneho striata pomocou vysokokalorických vizuálnych potravinových stimulov u obéznych jedincov
o Neuroimage, 37 (2007), s. 410 – 421
o
160.
o Salas a kol., 2010
o R. Salas, P. Baldwin, M. de Biasi, PR Montague
o BOLD odpovede na negatívne chyby predikcie predpovede v ľudskom habenula
o Predné. Hum. Neurosci., 4 (2010), str. 36
o
161.
o Saper a kol., 2002
o CB Saper, TC Chou, JK Elmquist
o Potreba kŕmiť: homeostatické a hedonické riadenie jedenia
o Neuron, 36 (2002), s. 199 – 211
o
162.
o Saunders, 2001
o R. Saunders
o Kompulzívna konzumácia jedla a žalúdočného bypassu: Čo s tým má hlad?
o Obes. Surg., 11 (2001), s. 757 – 761
o
163.
o Scarpace a kol., 2010
o PJ Šatka, M. Matheny, Y. Zhang
o Beh kolies eliminuje preferencie s vysokým obsahom tuku a zvyšuje signalizáciu leptínu vo ventrálnej tegmentálnej oblasti
Fyziol. Behav., 100 (2010), str. 173 – 179
o
164.
o Schiltz a kol., 2007
o CA Schiltz, QZ Bremer, CF Landry, AE Kelley
o Podnety súvisiace s jedlom menia funkčnú konektivitu predného mozgu, ako sa hodnotí s okamžitou expresiou skorého génu a proenkefalínu
o BMC Biol., 5 (2007), str. 16
o
165.
o Schur et al., 2009
o EA Schur, NM Kleinhans, J. Goldberg, D. Buchwald, MW Schwartz, K. Maravilla
o Aktivácia v regulácii energie mozgu a centier odmeňovania pomocou potravinových podnetov sa líši podľa výberu vizuálneho stimulu
o Int. J. Obes. (Lond.), 33 (2009), s. 653 – 661
o
166.
o Sclafani et al., 1998
A. Sclafani, RJ Bodnar, AR Delamater
o Farmakológia preferencií potravín
o Appetite, 31 (1998), str. 406
o
167.
o Sescousse a kol., 2010
o G. Sescousse, J. Redouté, JC Dreher
o Architektúra odmeňovania hodnoty kódovanie v ľudskej orbitofrontal kortex
o J. Neurosci., 30 (2010), str. 13095-13104
o
168.
o Shomaker a kol., 2010
o LB Shomaker, M. Tanofsky-Kraff, JM Zocca, A. Courville, M. Kozlosky, KM Columbo, LE Wolkoff, SM Brady, MK Crocker, AH Ali a kol.
o Stravovanie v neprítomnosti hladu u adolescentov: Príjem po jedle veľkého poľa v porovnaní s jedlom po štandardizovanom jedle
o Am. J. Clin. Nutr., 92 (2010), str. 697 – 703
o
169.
o Simmons a kol., 2005
o WK Simmons, A. Martin, LW Barsalou
o Obrázky chutných potravín aktivujú chuťové kôry pre chuť a odmenu
o Cereb. Cortex, 15 (2005), str. 1602 – 1608
o
170.
o Malé, 2010
o DM Small
o Zastúpenie chuti v ľudskej izolácii
o Štruktúra mozgu. Funkcia, 214 (2010), s. 551 – 561
o
171.
o Small et al., 2003
o DM Small, M. Jones-Gotman, A. Dagher
o Uvoľňovanie dopamínu vyvolané kŕmením v dorzálnom striatu koreluje s hodnotami príjemnosti jedla u zdravých dobrovoľníkov
o Neuroimage, 19 (2003), s. 1709 – 1715
o
172.
o Smith a Berridge, 2007
o KS Smith, KC Berridge
o Opioidný limbický okruh pre odmeňovanie: interakcia medzi hedonickými hotspotmi nucleus accumbens a ventral pallidum
o J. Neurosci., 27 (2007), str. 1594-1605
o
173.
o Söderpalm a Berridge, 2000
o AH Söderpalm, KC Berridge
Príjem potravy po diazepame, morfíne alebo muscimole: Mikroinjekcia V jadre nucleus accumbens
o Pharmacol. Biochem. Behav., 66 (2000), str. 429 – 434
o
174.
o Stice a kol., 2008a
E. Stice, S. Spoor, C. Bohon, DM Small
o Vzťah medzi obezitou a tupou striatálnou odpoveďou na potravu moderuje alela TaqIA A1
o Veda, 322 (2008), s. 449 – 452
o
175.
o Stice a kol., 2008b
o E. Stice, S. Spoor, C. Bohon, MG Veldhuizen, DM Small
Vzťah odmeňovania od príjmu potravy a očakávaného príjmu potravy k obezite: Funkčná štúdia magnetickej rezonancie
o J. Abnorm. Psychol., 117 (2008), s. 924 – 935
o
176.
o Stice a kol., 2010a
o E. Stice, S. Yokum, K. Blum, C. Bohon
o Prírastok hmotnosti je spojený so zníženou striatálnou odpoveďou na chutné jedlo
o J. Neurosci., 30 (2010), str. 13105-13109
o
177.
o Stice a kol., 2010b
o E. Stice, S. Yokum, C. Bohon, N. Marti, A. Smolen
o Odpoveď na odmeňovanie obvodov na potraviny predpovedá budúce zvýšenie telesnej hmotnosti: Moderujúce účinky DRD2 a DRD4
o Neuroimage, 50 (2010), s. 1618 – 1625
o
178.
o Stoeckel, 2010
o Stoeckel, LE (2010). Princíp obezity Zlatovláska. Vedecký Američan. Jún 8, 2010. http://www.scientificamerican.com/article.cfm?id=the-goldilocks-principle-obesity.
o
179.
Stratford a Kelley, 1997
TR Stratford, AE Kelley
o GABA v jadre nucleus accumbens sa zúčastňuje centrálnej regulácie kŕmenia
o J. Neurosci., 17 (1997), str. 4434-4440
o
180.
Stratford a Kelley, 1999
TR Stratford, AE Kelley
o Dôkaz funkčného vzťahu medzi jadrom nucleus accumbens a laterálnym hypotalamom pod kontrolou kontroly správania sa pri kŕmení
o J. Neurosci., 19 (1999), str. 11040-11048
o
181.
o Suarez a Gallup, 1981
o SD Suarez, GGJ Gallup
o Etologická analýza správania otvorených polí u potkanov a myší
o Naučte sa. Motiv., 12 (1981), s. 342 – 363
o
182.
o Sunday et al., 1983
o SR Nedeľa, SA Sanders, G. Collier
o Palatability a jedlo vzory
Fyziol. Behav., 30 (1983), str. 915 – 918
o
183.
o Swinburn a kol., 2009
B. Swinburn, G. Sacks, E. Ravussin
o Zvýšené dodávky energie z potravín sú viac než dostatočné na vysvetlenie epidémie obezity v USA
o Am. J. Clin. Nutr., 90 (2009), str. 1453 – 1456
o
184.
o Taha a Fields, 2005
o SA Taha, HL Fields
o Kódovanie chutnosti a chutného správania odlišnými neuronálnymi populáciami v nucleus accumbens
o J. Neurosci., 25 (2005), str. 1193-1202
o
185.
o Taha a kol., 2009
o SA Taha, Y. Katsuura, D. Noorvash, A. Seroussi, HL Fields
o Konvergentné, nie sériové, striatálne a palidálne okruhy regulujú príjem potravy vyvolanej opioidmi
o Neuroscience, 161 (2009), str. 718 – 733
o
186.
o Teegarden a Bale, 2007
o SL Teegarden, TL Bale
o Zníženie preferencií v strave spôsobuje zvýšenú emocionalitu a riziko relapsu diét
o Biol. Psychiatria, 61 (2007), s. 1021 – 1029
o
187.
o Teegarden a kol., 2009
o SL Teegarden, AN Scott, TL Bale
o Včasné vystavenie dietám s vysokým obsahom tuku podporuje dlhodobé zmeny v stravovacích preferenciách a centrálnu signalizáciu odmeňovania
o Neuroscience, 162 (2009), str. 924 – 932
o
188.
o Thanos a kol., 2008
o PK Thanos, M. Michaelides, YK Piyis, GJ Wang, ND Volkow
o Obmedzenie potravín výrazne zvyšuje dopamínový receptor D2 (D2R) na potkaních modeloch obezity, ako sa hodnotí in-vivo zobrazovaním muPET ([11C] racloprid) a in vitro ([3H] spiperón) autorádiografiou
o Synapse, 62 (2008), str. 50 – 61
o
189.
o Thompson a Swanson, 2010
RH Thompson, LW Swanson
o Analýza štruktúrneho prepojenia riadená hypotézou podporuje sieť nad hierarchickým modelom architektúry mozgu
o Proc. Natl. Acad. Sci. USA, 107 (2010), s. 15235 – 15239
o
190.
o Vanderschuren a Everitt, 2004
o LJ Vanderschuren, BJ Everitt
o Hľadanie lieku sa stáva nutkavým po dlhšom užívaní kokaínu
o Veda, 305 (2004), s. 1017 – 1019
o
191.
o Vendruscolo et al., 2010a
o LF Vendruscolo, AB Gueye, M. Darnaudéry, SH Ahmed, M. Cador
• Nadmerná spotreba cukru počas dospievania selektívne mení motiváciu a odmeňovanie u dospelých potkanov
o PLoS ONE, 5 (2010), str. e9296 http://dx.doi.org/10.1371/journal.pone.0009296
o
192.
o Vendruscolo et al., 2010b
o LF Vendruscolo, AB Gueye, JC Vendruscolo, KJ Clemens, P. Mormède, M. Darnaudéry, M. Cador
o Redukované pitie alkoholu u dospelých potkanov vystavených sacharóze počas dospievania
o Neurofarmakológia, 59 (2010), pp. 388 – 394
o
193.
o Volkow a O'Brien, 2007
o ND Volkow, CP O'Brien
o Problémy pre DSM-V: Mala by byť obezita zahrnutá ako porucha mozgu?
o Am. J. Psychiatry, 164 (2007), str. 708 – 710
o
194.
o Volkow a Wise, 2005
o ND Volkow, RA Wise
o Ako nám drogová závislosť pomôže pochopiť obezitu?
o Nat. Neurosci., 8 (2005), pp. 555-560
o
195.
o Volkow a kol., 2009
o ND Volkow, GJ Wang, F. Telang, JS Fowler, RZ Goldstein, N. Alia-Klein, J. Logan, C. Wong, PK Thanos, Y. Ma, K. Pradhan
o Inverzná súvislosť medzi BMI a prefrontálnou metabolickou aktivitou u zdravých dospelých jedincov
o Obezita (Silver Spring), 17 (2009), pp. 60 – 65
o
196.
o Vucetic a Reyes, 2010
o Z. Vucetic, TM Reyes
o Centrálne dopaminergné obvody riadiace príjem a odmenu za potravu: dôsledky pre reguláciu obezity
o Wiley Interdiscip. Syst. Biol. X., 2 (2010), str. 577-593
o
197.
o Wang a kol., 2001
o GJ Wang, ND Volkow, J. Logan, NR Pappas, CT Wong, W. Zhu, N. Netusil, JS Fowler
o dopamín mozgu a obezita
o Lancet, 357 (2001), s. 354 – 357
o
198.
o Wang a kol., 2002
o GJ Wang, ND Volkow, JS Fowler
o Úloha dopamínu v motivácii pre potraviny u ľudí: dôsledky pre obezitu
o Expert Opin. Ther. Ciele, 6 (2002), str. 601 – 609
o
199.
o Wang a kol., 2004a
o GJ Wang, ND Volkow, F. Telang, M. Jayne, J. Ma, M. Rao, W. Zhu, CT Wong, NR Pappas, A. Geliebter, JS Fowler
o Expozícia chutným potravinovým stimulom výrazne aktivuje ľudský mozog
o Neuroimage, 21 (2004), s. 1790 – 1797
o
200.
o Wang a kol., 2004b
o GJ Wang, ND Volkow, PK Thanos, JS Fowler
o Podobnosť medzi obezitou a drogovou závislosťou podľa neurofunkčného zobrazovania: prehľad koncepcie
o J. Addict. Dis., 23 (2004), str. 39 – 53
o
201.
o Wang a kol., 2008
o GJ Wang, D. Tomasi, W. Backus, R. Wang, F. Telang, A. Geliebter, J. Korner, A. Bauman, JS Fowler, PK Thanos, ND Volkow
o Gastrická distenzia aktivuje obvody sýtosti v ľudskom mozgu
o Neuroimage, 39 (2008), s. 1824 – 1831
o
202.
o Wassum et al., 2009
o KM Wassum, SB Ostlund, NT Maidment, BW Balleine
o Zreteľné okruhy opioidov určujú chutnosť a vhodnosť odmeňovania udalostí
o Proc. Natl. Acad. Sci. USA, 106 (2009), s. 12512 – 12517
o
203.
o Wassum et al., 2011
o KM Wassum, IC Cely, BW Balleine, NT Maidment
o Aktivácia μ-opioidného receptora v bazolaterálnej amygdale sprostredkováva učenie zvýšenia, ale nie zníženie stimulačnej hodnoty potravinovej odmeny
o J. Neurosci., 31 (2011), str. 1591-1599
o
204.
o Wilkinson a Peele, 1962
o HA Wilkinson, TL Peele
o Modifikácia intrakraniálnej samostimulácie hladu
o Am. J. Physiol., 203 (1962), str. 537-540
o
205.
o Will et al., 2003
o MJ Will, EB Franzblau, AE Kelley
o Nucleus accumbens mu-opioidy regulujú príjem stravy s vysokým obsahom tuku prostredníctvom aktivácie distribuovanej siete mozgu
o J. Neurosci., 23 (2003), str. 2882-2888
o
206.
o Wilson a kol., 2008
o ME Wilson, J. Fisher, A. Fischer, V. Lee, RB Harris, TJ Bartness
o Kvantifikácia príjmu potravy v sociálne umiestnených opiciach: účinky na sociálny stav na spotrebu kalórií
Fyziol. Behav., 94 (2008), str. 586 – 594
o článok PDF (553 K) |
Zobraziť záznam v Scopus
| Citované v Scopus (22)
207.
o Yurcisin et al., 2009
o BM Yurcisin, MM Gaddor, EJ DeMaria
o Obezita a bariatrická chirurgia
o Clin. Chest Med., 30 (2009), str. 539 – 553, ix
o
208.
o Zhang a kol., 1998
o M. Zhang, BA Gosnell, AE Kelley
o Príjem potravy s vysokým obsahom tuku je selektívne zvýšený stimuláciou mu opioidného receptora v jadre nucleus accumbens
J. Pharmacol. Exp. Ther., 285 (1998), s. 908 – 914
o Zobraziť záznam v Scopus
| Citované v Scopus (148)
209.
o Zheng a kol., 2007
o H. Zheng, LM Patterson, HR Berthoud
o Signalizácia orexínu vo ventrálnej tegmentálnej oblasti je potrebná pre apetít s vysokým obsahom tuku vyvolaný opioidnou stimuláciou nucleus accumbens
o J. Neurosci., 27 (2007), str. 11075-11082
o Zobraziť záznam v Scopus
| Citované v Scopus (64)
210.
o Zheng a kol., 2009
o H. Zheng, NR Lenard, AC Shin, HR Berthoud
• Regulácia chuti k jedlu a regulácia energetickej bilancie v modernom svete: Rehabilitovaný mozog má prednosť pred signálmi o opätovnom naplnení
o Int. J. Obes. (Lond.), 33 (Suppl 2) (2009), s. S8 – S13
o Zobraziť záznam v Scopus
| Citované v Scopus (34)