Front Behav Neurosci. 2015 môže 19; 9: 128. doi: 10.3389 / fnbeh.2015.00128. eCollection 2015.
Andreatta M1, Pauli P1.
abstraktné
Pri klasickom podmieňovaní sa pôvodne neutrálny stimul (kondicionovaný stimul, CS) spája s biologicky významnou udalosťou (nepodmienený stimul, USA), ktorou môže byť bolesť (averzívne podmieňovanie) alebo jedlo (apetitívne podmieňovanie). Po niekoľkých asociáciách je CS schopný iniciovať obranné alebo dohadovacie reakcie. Na rozdiel od averzného podmieňovania sa apetitívny podmieňok u ľudí skúma zriedka, hoci jeho význam pre normálne a patologické správanie (napr. Obezita, závislosť) je nepopierateľný. Zámerom tejto štúdie je previesť nálezy na zvieratách týkajúce sa apetitívnych úprav na ľudí, ktorí používajú jedlo v USA. Tridsaťtri účastníkov bolo vyšetrovaných medzi 8. a 10. hodinou bez raňajok, aby sa ubezpečil, že pociťujú hlad. Počas dvoch fáz akvizície jeden geometrický tvar (avCS +) predpovedal averzívny USA (bolestivý elektrický šok), ďalší tvar (appCS +) predpovedal apetitívny USA (čokoláda alebo slaný praclík podľa preferencie účastníkov) a tretí tvar (CS– ) nepredpovedali ani USA. Vo fáze zániku boli tieto tri tvary plus nový tvar (NOVÝ) predstavené opäť bez doručenia do USA. Hodnoty valencie a vzrušenia, ako aj reakcie šoku a vodivosti kože (SCR) sa zbierali ako indexy učenia. Našli sme úspešnú averzívnu a apetitívnu úpravu. Na jednej strane bol avCS + hodnotený ako negatívnejší a vzrušujúcejší ako CS– a indukoval potenciu potláčania a zlepšenú SCR. Na druhej strane bol appCS + hodnotený pozitívnejšie ako CS– a indukoval útlm úľavy a väčšiu SCR. V súhrne sme úspešne potvrdili nálezy na zvieratách u (hladných) ľudí demonštráciou apetitívneho učenia a normálneho averzného učenia.
úvod
Predpovedanie hrozby a jedla má zásadný význam pre prežitie každého organizmu. V klasickom kondicionovaní (Pavlov, 1927), narážka niekoľkokrát predchádza averzívnej udalosti, ako je mierny bolestivý elektrický šok (averzívny nepodmienený stimul, USA) alebo apetitívna udalosť, ako napríklad potravinová peleta (appetitive US). Potom je tento samotný stimul (teraz označený ako CS, CS +) schopný vyvolať buď obranné, alebo konzumné reakcie. Prvý typ asociatívneho učenia sa nazýva averzívne kondicionovanie, zatiaľ čo druhé sa nazýva apetitívne kondicionovanie.
Aj keď predpovede príhodných príhod sú pre prežitie rovnako dôležité ako predpovede averzívnych príhod, na zvieratách sa pozoruhodne menej skúma podmienená chuť k jedlu (Bouton a Peck, 1989; Koch a spol., 1996; McDannald a kol., 2011, pre prehľad pozri Martin-Soelch a kol., 2007) ako aj u ľudí (Klucken a kol., 2009, 2013; Austin a Duka, 2010; Delgado a kol., 2011; Levy a Glimcher, 2011). Tento nedostatok výskumu by mohol byť pravdepodobne spôsobený zložitosťou chutnej paradigmy v porovnaní s averzívnym. Napríklad, jedlo ako primárny zosilňovač sa musí dodávať, keď je organizmus hladný, aby bol prospešný (posledný prehľad pozri Dickinson a Balleine, 1994; Clark a kol., 2012). V ľudskom výskume sa tento problém prekonal pomocou peňazí (Austin a Duka, 2010; Delgado a kol., 2011; Levy a Glimcher, 2011) alebo erotické obrázky (Klucken a kol., 2009, 2013). Štúdie neuro-zobrazovania však poukázali na to, že primárne (tj občerstvenie alebo nápoje) a sekundárne (tj peniaze) zosilňovače aktivujú niektoré bežné oblasti mozgu (napr. Striatum), ale tiež znamenajú odlišné vzorce aktivácie (Delgado a kol., 2011; Levy a Glimcher, 2011).
Pokiaľ je nám známe, iba niekoľko štúdií kondicionovania u ľudí skúmalo účinky primárnych zosilňovačov chuti, ako je zápach (Gottfried a kol., 2002), voda (Kumar a kol., 2008) alebo jedlo (Prévost a kol., 2012). V prvej štúdii Gottfried a kol. (2002) spojené neutrálne tváre (CS) s nepríjemným, príjemným alebo neutrálnym zápachom. Zaujímavé je, že našli väčšiu aktiváciu v orbitofrontálnej kôre (OFC) a vo ventrálnom striate v reakcii na chutný CS + v porovnaní s averzívnym CS +. Dospeli k záveru, že OFC spracúva hodnotu zápachu a podieľa sa na prenose afektívnej hodnoty z čuchovej (US) do vizuálnej (CS) sústavy. Aktivácia (ventro) striatalu sa interpretovala tak, že odzrkadľuje chutný CR vyvolaný chutný CS +. V druhej štúdii Kumar a kol. (2008) pozvali svojich účastníkov (zdravé kontroly a pacientov s výraznou depresiou) do laboratória skoro ráno a požiadali ich, aby sa zdržali pitia počas noci, aby sa pred skenovaním ubezpečili, že majú smäd. Fraktálne obrázky boli CS a 0.1 ml vody v USA. Je zaujímavé, že okrem iných aktivácií zdraví účastníci (ale nie depresívni pacienti) vykazovali väčšiu aktiváciu vo ventrálnom striatume na chutný CS +, čo naznačuje, že tento stimul bol spracovaný ako prospešný. V tretej štúdii Prévost a kol. (2012) tiež prezentoval fraktálne obrázky ako CS a podľa preferencie účastníkov buď sladké alebo slané občerstvenie ako USA. CS bol prezentovaný po dobu 6 s a počas poslednej sekundy bol dodatočne predstavený potravinový obraz. Zakaždým, keď sa objavil obrázok jedla, experimentátor vložil kúsok jedla do rúk účastníkov, ktorí mali dovolené občerstvenie okamžite skonzumovať. Autori bohužiaľ nehlásili aktiváciu mozgu apetitívnemu CS + počas fázy klasickej úpravy; pozorovali však menšie spomalenie srdca na CS + s odmenou v porovnaní s neodmenenými CS–, čo naznačuje rozdielne kondičné účinky.
Nedostatok štúdií, ktoré využívajú prekvapujúcu reakciu ako index apetitívneho kondicionovania, je prekvapujúci, najmä vzhľadom na jeho široké využitie pri averzívnom kondicionovaní. Šokujúca reakcia je predková a automatická obranná reakcia na náhle, neočakávané a silné averzívne udalosti (Koch, 1999). Táto obranná reakcia je sprostredkovaná relatívne jednoduchou neuronálnou cestou zahŕňajúcou kochleárne koreňové neuróny, jadro kaudálneho pontínu retikulárnej formácie (PnC) a spinálne motoneuróny (Fendt a Fanselow, 1999; Koch, 1999). Štúdie na zvieratách odhalili, že desivé potencovanie závisí od projekcií amygdaly k PnC (Fendt a Fanselow, 1999; Koch, 1999), zatiaľ čo úľava od prekvapenia závisí od neporušeného jadra accumbens (NAcc, Koch, 1999). Takáto modulácia prekvapujúcej reakcie je užitočným implicitným meradlom valencie prezentovaných stimulov v popredí. Potenciacia teda naznačuje negatívnu valenciu, zatiaľ čo útlm naznačuje pozitívnu valenciu, pričom obidve tieto procesy nie sú výrazne ovplyvnené kognitívnymi procesmi (Hamm a Weike, 2005; Andreatta a kol., 2010). Pokiaľ je nám známe, iba jedna štúdia na zvieratách skúmala podmienenú chuť do jedla pomocou merania prekvapujúcich reakcií ako závislého opatrenia. Je zaujímavé, že skúmané potkany vykazovali prekvapujúci útlm na CS + (tj. Svetlo) spojené s dodávaním roztoku sacharózy. Takéto utlmenie bolo narušené najmä u zvierat s léziami NAcc, ale nie u zvierat s léziami na amygdale, čo naznačuje, že NAcc hrá špecifickú úlohu pri vyvolaní apetitívnych CR a pri tlmení prekvapujúcich reakcií (Koch a spol., 1996).
Súčasná štúdia je podľa našich vedomostí prvá, ktorá preložila túto apetitívnu podmienku kondicionovania na ľudí pomocou primárnych zosilňovačov ako USA, tj sladkých (čokoládové smarties)®) alebo slané (slané praclíky) a prekvapujúca modulácia ako miera CR. Očakávali sme, že appCS + bude spúšťať silné apetitívne CR, čo sa odráža v prekvapivom útlme, zvýšenej SCR a pozitívnom valenčnom hodnotení v porovnaní s ostatnými stimulmi, tj avCS + a CS–.
Materiály a metódy
účastníci
Štyridsaťdva dobrovoľníkov prijalo účasť na štúdii a získalo kredity za kurz. Deväť účastníkov bolo z analýzy vylúčených, dvaja z dôvodu technických problémov, traja preto, lebo boli kódovaní ako nereagujúci (stredná amplitúda úľaku <5 μV), a štyria z dôvodu, že nevykazovali dostatok úľakových reakcií na jednu podmienku (minimálne = 2; podrobnosti pozri Materiály a metóda). Nakoniec bolo do analýzy zahrnutých 33 účastníkov (16 mužov; priemerný vek: 22.09 rokov, SD: 2.84; rozsah: 18 – 29 rokov). Štyria účastníci neboli rodení Nemci a šesť bolo ľavákov. Jeden účastník si počas experimentu nebol vedomý asociácií CSs-USA (pozri postup), rozhodli sme sa však tohto účastníka nevylúčiť, pretože jeho odpovede boli normálne a neovplyvnili výsledky.
materiály
Nepodmienené Stimuli (USA)
Boli použité dva druhy USA. Ako averzívny USA sme na dominantné predlaktie účastníkov pôsobili mierne bolestivým elektrickým prúdom. Elektrický šok bol dodávaný pomocou dvoch elektród s priemerom 9 mm a rozstupom 30 cm. Elektrický stimul pozostával z impulzného stimulu s frekvenciou 50 Hz a trvania 200 ms, generovaného prúdovým stimulátorom (Digitimer DS7A, Digitimer Ltd, Welwyn Garden City, UK, 400 V, maximum 9.99 mA). Intenzita elektrického šoku bola stanovená individuálne pomocou postupu opísaného v predchádzajúcom texte (Andreatta a kol., 2010). Stručne povedané, účastníci podstúpili dve série stúpajúcich a klesajúcich intenzít v intervaloch 0.5 mA. Museli hodnotiť každý podnet na vizuálnej stupnici od 0 (vôbec necítiac) do 10 (skutočne intenzívna bolesť), pričom 4 bol kotvou prahu (len viditeľná bolesť). Priemerná intenzita elektrického stimulu bola 2.12 mA (SD = 0.56) a bolo hodnotené ako bolestivé (M = 6.45, SD = 1.73). Chutný USA pozostával buď z čokolády (Smarties®) alebo malý slaný praclík. Výber apetitívneho USA závisel od individuálnych preferencií účastníka, ako sa uviedli počas predbežného rozhovoru. Účastníci museli menovite uviesť, či počas raňajok bežne jedia slané alebo sladké jedlá. Tiež si mohli slobodne zvoliť, či majú pri samotnom experimente radšej čokoládu alebo slaný praclík. Nakoniec si 22 účastníkov vybralo čokoládu a 11 malý slaný praclík.
Podmienené stimuly (CS)
Geometrické tvary (modrý štvorec, žltý kruh, zelený trojuholník, červený šesťuholník) s uhlopriečkou 8 cm boli prezentované ako CS. Tvary boli prezentované v strede čiernej obrazovky počítača pre 8 s. Jeden tvar (avCS +) bol vždy spojený s averzívnym USA (bolestivý elektrický šok), jeden tvar (appCS +) bol vždy spojený s apetítom USA (čokoláda alebo slaný preclík), jeden tvar (CS -) nebol nikdy spojený s ani averzívnou alebo apetitívne US a štvrtý tvar (NEW) bol prezentovaný vo fáze vyhynutia, ale nie počas akvizičnej fázy, aby sa zabezpečila jeho neutralita.
Štartovacia sonda
Ako sonda na prekvapenie sa použil biely šum 103 dB s trvaním 50 ms. Akustické podnety boli prezentované binaurálne cez slúchadlá a vyskytli sa náhodne 4–6 s po nástupe tvaru.
dotazníky
Pred a po experimente museli účastníci vyplniť nemecké verzie Inventarizácie stavu úzkosti (STAI). Laux a kol., 1981) a plán pozitívnych negatívnych vplyvov (PANAS, Krohne a kol., 1996). STAI je zoznam na hodnotenie znakov a / alebo stavov úzkosti účastníkov a skladá sa z 20 položiek pre danú vlastnosť aj pre štátnu verziu. Úroveň úzkosti účastníkov predtým (M = 37.06, SD = 7.80) a po ňom (M = 39.33, SD = 9.16) experiment sa významne nezmenil [t(32) = 1.61, p = 0.117]. Skóre skóre úzkosti v aktuálnej vzorke sa pohybovalo medzi 20 a 58 (M = 36.6, SD = 8.98), čo je porovnateľné s publikovaným normálnym rozsahom dospelých (Laux a kol., 1981). PANAS (Krohne a kol., 1996) je index pozitívnej a negatívnej nálady. Jedinci s vysokým skóre na škále pozitívnych vplyvov (PAS) sú náchylní na emócie, ako je nadšenie, zatiaľ čo jedinci s vysokým skóre na škále negatívnych vplyvov (NAS), sú náchylní k emóciám, ako je strach. Každá položka sa skladá z prídavného mena a účastníci naznačujú na škále od 1 (veľmi mierne) do 5 (extrémne), do akej miery prídavné meno odráža ich pocity v konkrétnom okamihu. V dôsledku experimentu sa nezistili významné rozdiely v negatívnom vplyve účastníkov [začať: M = 11.67, SD = 2.29; koniec: M = 12.88, SD = 4.69; t(32) = 1.55, p = 0.130]. Účastníci od začiatku výrazne zmenili svoju pozitívnu náladu od začiatku (M = 26.72, SD = 4.70) až do konca (M = 23.66, SD = 7.02) experimentu [t(31) = 3.11, p = 0.004]. Toto zníženie pozitívnej nálady účastníkov mohlo súvisieť s nepríjemnosťou paradigmy (boli predstavené bolestivé elektrické šoky, ako aj averzívny biely šum).
Postup
Po príchode do laboratória si účastníci prečítali a podpísali formulár informovaného súhlasu schválený etickou komisiou Katedry psychológie Univerzity vo Würzburgu. Neboli informovaní o podmienenosti medzi CS a USA. Po vyplnení dotazníkov boli elektródy pripojené a procedúra prahu bolesti bola vykonaná tak, ako je opísané vyššie.
Pri fáza návyku, štyri geometrické tvary boli prezentované dvakrát s inter-trial intervalom (ITI) medzi 18 a 25 s (priemer: 21.5 s). Počas tejto fázy neboli dodané žiadne sondy v USA alebo prekvapujúce.
Pred akvizičnou fázou bolo každému 7-15u dodaných sedem výbojov bieleho šumu, aby sa znížila počiatočná reaktivita.
Nasledujúce dva fázach akvizície boli identické (obrázok č. \ t 1). Každá akvizičná fáza pozostávala z testov 24: štúdie 8 CS -, štúdie 8 avCS + a štúdie 8 appCS +. Sekvencia CS bola pseudonáhodná s jediným obmedzením, že rovnaký stimul nemohol byť prezentovaný viac ako dvakrát za sebou. Pozoruhodne, avCS + bol prezentovaný spolu s bleskom ako symbol pre elektrický šok a bolestivé USA boli dodané na ofset. Aplikácia appCS + bola prezentovaná v zlúčenine s obrazom Smarties alebo slaného preclíka a účastník si mohol vybrať pohárik alebo preclík z pohára. CS - bol predložený spolu so symbolom zákazu a žiadne USA neboli dodané. Počas troch z 8 CS prezentácií každého typu bola medzi 4 a 6 s po nástupe stimulačnej sondy dodaná šoková sonda. Počas ITI boli prezentované tri ďalšie senzačné sondy, aby sa zabezpečila ich nepredvídateľnosť a znížila sa návyková situácia. ITI, skladajúci sa z čiernej obrazovky, sa líšil medzi 18 a 25 s priemerom 21.5 s.
OBRÁZOK 1
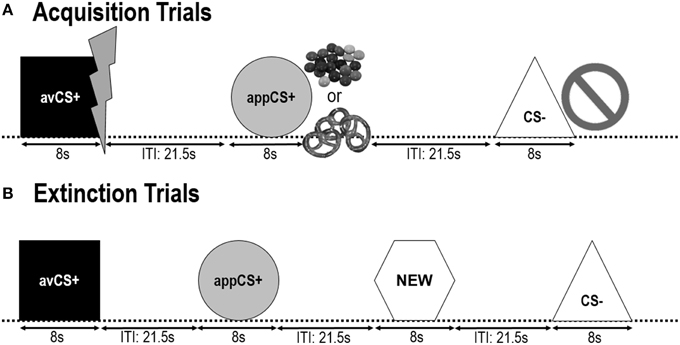
Obrázok 1. Skúšky počas dvoch akvizičných fáz (A) a fázy zániku (B), Účastníci sa dozvedeli, že jeden tvar (avCS +) predpovedal mierny bolestivý elektrický šok, jeden tvar (appCS +) predpovedal buď kúsok čokolády alebo malý slaný preclík (podľa ich preferencie) a tretí tvar (CS–) nepredpovedal žiadne biologicky najvýraznejšia udalosť. Každý tvar bol prezentovaný v spojení s obrázkom zobrazujúcim elektrický šok, šialenstvo / slaný preclík, alebo nič v závislosti od asociácie s USA. Počas fázy zániku boli opäť prezentované tri geometrické tvary, ale neboli dodané žiadne USA. Okrem toho, štvrtý geometrický tvar (NOVÝ) bol prezentovaný ako neutrálna kontrola.
Pri fáza zánikuúčastníci opäť videli tri geometrické tvary (napr. avCS +, appCS + a CS–) plus nový neutrálny tvar (NOVÝ). USA neboli dodané a tvary neboli prezentované v spojení s fotografiami zábleskov, čokolády / slaných praclíkov alebo zákazov. Každý stimul bol prezentovaný osemkrát v pseudonáhodnom poradí (tj rovnaký stimul nebol prezentovaný viac ako dvakrát v rade), čím sa vytvorili 32 štúdie. Stimuly štartovacej sondy boli prezentované počas 4 z 8 stimulačných prezentácií každého CS typu. Rovnako ako vo fáze akvizície a návyku, ITI sa líšili medzi 18 a 25 s a 4 ďalšie prekvapujúce sondy boli počas ITI nepredvídateľne dodané.
Po každej fáze účastníci hodnotili valenciu (príjemnosť) a vzrušenie (excitáciu) CS s vizuálnymi analógovými váhami (VAS) od 1 po 9. Stupnica stupnice sa pohybovala od „1“, čo označuje „veľmi nepríjemné“ až „9“, čo znamená „veľmi príjemné“, stupnica vzrušenia sa pohybovala od „1“, čo znamená „pokoj“ až „9“, čo znamená „vzrušujúce“. vyhodnotené po dvoch fázach akvizície a fáze zániku. Účastníci videli geometrický tvar pre 1 s a potom museli ukázať, či tento tvar bol spojený s elektrickým šokom, s čokoládou / slaným preclíkom, s ničím, alebo či neboli schopní urobiť žiadne spojenie. Všetci účastníci (okrem jedného) si boli vedomí nepredvídateľnosti po akvizícii 2.
Redukcia dát
Fyziologické reakcie boli zaznamenané pomocou zosilňovača V-Amp 16 a Vision Amp Edition V-Amp Edition (verzia 1.03.0004, BrainProducts Inc., Mníchov, Nemecko). Použila sa vzorkovacia frekvencia 1000 Hz a vrubový filter 50 Hz. Analýzy offline sa uskutočnili s analyzátorom Brain Vision (verzia 2.0; BrainProducts Inc., Mníchov, Nemecko).
Odozva odštartovania
Odozva na vyľakanie bola meraná pomocou elektromyografie (EMG) vľavo orbicularis oculi svalov s dvoma elektródami 5 mm Ag / AgCl. V súlade s pokynmi (Blumenthal a kol., 2005), jedna elektróda bola umiestnená pod zornicu a druhá 1 cm bočne. Uzemnené a referenčné elektródy boli umiestnené vpravo a vľavo. Pred pripevnením elektród sa koža ľahko obrúsila a vyčistila alkoholom, aby sa udržala impedancia nižšia ako 10 kΩ. Elektromyografický signál bol offline filtrovaný s 28 Hz low cutoff filtrom a 500 Hz high cutoff filter. Potom sa usmernil EMG signál a aplikoval sa kĺzavý priemer 50 ms. Použili sme 50 ms pred začiatkom sondy ako východiskovú hodnotu (Grillon a kol., 2006). Odpovede na vyľakané sondy boli vyhodnotené manuálne a pokusy s nadmernými základnými posunmi (± 5 μV) alebo pohybovými artefaktmi boli z ďalšej analýzy vylúčené. Odozvy na počiatočné hodnoty nižšie ako 5 μV boli kódované ako nulové a zohľadnili sa pri výpočte veľkosti vyľakania (Blumenthal a kol., 2005). Spolu, 10.4% testov bolo zamietnutých, a minimálne 2 z 3 vyľakaných reakcií v akvizičných fázach a 4 z 8 odozvy odozvy v extinkčnej fáze pre každú podmienku boli vyžadované na udržanie účastníka v skupine analýz. Z tohto dôvodu boli vylúčení štyria účastníci. Maximálna amplitúda bola definovaná ako maximálny vrchol vo vzťahu k základnej čiare počas časového okna 20-120 ms po nástupe sondy. Surové dáta sa potom normalizovali v rámci subjektov s použitím z-scores a potom T-skóre s cieľom znížiť vplyv individuálnej variability a lepšie odhaliť psychologické procesy. T-skóre bolo spriemerované pre každú podmienku (avCS +, appCS +, CS-, NEW a ITI). Na zistenie potencovania úľavy alebo útlmu úľavy boli odčítané hodnoty pre reakcie ITI vyľakania od vyľakaných reakcií pre každý stav.
Reakcia vodivosti kože (SCR)
Reakcia vodivosti kože (SCR) bola zaznamenávaná s použitím dvoch elektród 5 mm Ag / AgCl umiestnených na dlani nedominantnej ruky. Galvanická odozva bola filtrovaná offline pomocou filtra 1 Hz. SCR bola definovaná ako rozdiel (v μS) medzi nástupom odpovede (1 – 3 s po nástupe stimulu) a píkom odpovede (Tranel a Damasio, 1994; Delgado a kol., 2011). Pri analýze SCR sa nebrali do úvahy pokusy obsahujúce sondy. Reakcie pod 0.02 μS boli kódované ako nulové. Päť ďalších účastníkov bolo vylúčených z SCR analýzy, pretože mali priemernú SCR nižšiu ako 0.02 μS. Údaje o surovej vodivosti kože boli odmocninové transformované, aby sa normalizovala distribúcia a skóre boli spriemerované pre každú podmienku samostatne pre dve akvizičné fázy (avCS +, appCS +, CS–) a fázu extinkcie (avCS +, appCS +, CS– a NEW). ).
Štatistická analýza
Všetky údaje boli analyzované pomocou SPSS pre Windows (verzia 20.0, SPSS Inc.). Pre fyziologické reakcie sa vypočítali oddelené multivariačné analýzy rozptylu (ANOVA) pre dve fázy akvizície a fázu extinácie. ANOVA pre fázy akvizície mala stimuly (avCS +, appCS +, CS–) a fázu (Acquisition 1, Acquisition 2) ako faktory v rámci subjektov. ANOVA pre fázu extinkcie mala iba stimulačný faktor (avCS +, appCS +, CS–, NEW) ako faktor v rámci subjektu. Valenčné, vzrušujúce a kontingenčné hodnotenia boli analyzované s oddelenými ANOVA, ktoré obsahujú stimuly interných subjektov (avCS +, appCS +, CS– a NEW) a fázu. Tento faktor mal štyri úrovne pre hodnoty valencie a vzrušenia (T1: po návykovej fáze, T2: po prvej akvizičnej fáze, T3: po druhej akvizičnej fáze, T4: po fáze extinkcie), ale tri úrovne pre kontingenčné hodnotenia (T1: po prvá akvizičná fáza, T2: po druhej akvizičnej fáze, T3: po fáze extinácie).
Úroveň alfa (α) bola nastavená na 0.05 pre všetky analýzy. Veľkosť účinku sa uvádza ako čiastočná η2.
výsledky
Hodnoty valencie a vzrušenia pre každú fázu sú znázornené na obrázku č 2; prekvapujúce reakcie a SCR sú znázornené na obr 3.
OBRÁZOK 2
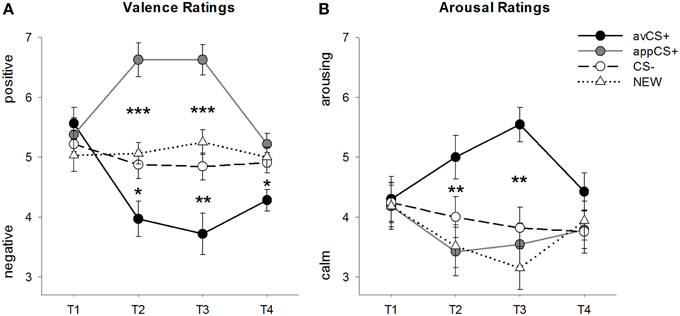
Obrázok 2. Hodnotenia pre valenciu (A) a vzrušenie (B), Čiary (so štandardnými chybami) zobrazujú hodnotenia po fáze návyku (T1), akvizícia 1 (T2), akvizícia 2 (T3) a extinkčná fáza (T4). Averzívna CS + (čierna plná čiara) získala negatívnu valenciu a vysoké vzrušenie po dvoch fázach získavania v porovnaní s CS– (čierna prerušovaná čiara) a NOVÁ (čierna bodkovaná čiara). Dôležité je, že chutný CS + (šedá plná čiara) získal v porovnaní s CS– a NOVÉ pozitívnu valenciu. *p <0.05, **p > 0.01, ***p <0.001.
OBRÁZOK 3
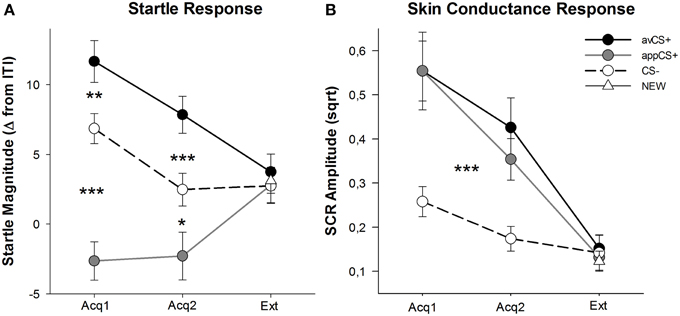
Obrázok 3. Úžasné reakcie (A) a vodivosť kože (B) (so štandardnými chybami) počas prvej akvizičnej fázy (Acq1), druhej akvizičnej fázy (Acq2) a extinkčnej fázy (Ext), Úžasné reakcie boli významne zosilnené na averzívnu CS + (čierna plná čiara) a významne oslabené na chutný CS + (šedá plná čiara) v porovnaní s CS– (čierna prerušovaná čiara) počas akvizičných fáz. SCR bola významne vyššia ako avCS + a appCS + v porovnaní s CS–. Počas zánikovej fázy neboli zistené žiadne rozdiely. *p <0.05, **p > 0.01, ***p <0.001.
Hodnotenie
ANOVA pre valenčné ratingy počas akvizície odhalili významné hlavné účinky stimulu [F(3, 93) = 17.26, GG-ε = 0.801, p <0.001, čiastočné η2 = 0.358] a fáza [F(3, 93) = 3.30, GG-ε = 0.731, p = 0.039, čiastočné η2 = 0.096], ako aj významnú interakciu medzi stimulom a fázou [F(9, 279) = 9.54, GG-ε = 0.463, p <0.001, čiastočné η2 = 0.235]. Nasleduj t- testy naznačujú, že valencie štyroch geometrických tvarov boli na začiatku experimentu rovnaké (ps> 0.19), zatiaľ čo po akvizícii 1 aj akvizícii 2 bol avCS + hodnotený ako obzvlášť negatívny a appCS + ako obzvlášť pozitívny. Konkrétne bol avCS + hodnotený ako viac negatívne hodnotený v porovnaní s CS– [Acq1: t(31) = 2.34, p = 0.026; Acq2: t(31) = 3.07, p = 0.004], NOVÝ [Acq1: t(31) = 2.70, p = 0.011; Acq2: t(31) = 3.89, p <0.001] a appCS + [Acq1: t(31) = 5.41, p <0.001; Acq2: t(31) = 6.11, p <0.001]. AppCS + bol hodnotený ako výrazne pozitívnejší ako CS– [Acq1: t(31) = 4.99, p <0.001; Acq2: t(31) = 5.31, p <0.001] a NOVÉ [Acq1: t(31) = 4.92, p <0.001; Acq2: t(31) = 4.14, p <0.001]. Rozdiely medzi CS– a NOVÝM neboli nikdy významné (ps> 0.18).
Dôležité je, že pri porovnaní valenčných hodnotení aplikácie appCS + spojených s čokoládou verzus soľným preclíkom neboli nájdené žiadne významné rozdiely [Acq1: t(30) = 0.03, p = 0.477; Acq2: t(30) = 0.29, p = 0.775].
Po vyhynutej fáze bol avCS + stále hodnotený ako negatívnejší v porovnaní s CS– [t(31) = 2.40, p = 0.023], NOVÝ [t(31) = 3.40, p = 0.002] a appCS + [t(31) = 3.35, p = 0.002]. Naopak, valencia appCS + sa už nelíšila od CS– [t(31) = 1.77, p = 0.086] alebo NEW [t(31) = 1.07, p = 0.293].
ANOVA pre hodnotenia vzrušenia počas akvizície odhalil významný hlavný efekt stimulu [F(3, 96) = 7.07, GG-ε = 0.737, p = 0.001, čiastočné η2 = 0.181], ale nie z fázy [F(3, 96) = 1.27, GG-ε = 0.805, p = 0.289, čiastočné η2 = 0.038] a významná interakcia medzi stimulom a fázou [F(9, 288) = 4.53, GG-ε = 0.582, p = 0.001, čiastočné η2 = 0.124]. Nasleduj t- testy neukázali žiadne významné rozdiely medzi stimulmi, pokiaľ ide o ich počiatočné vzrušenie (ps> 0.74). Avšak po dvoch akvizičných fázach bol avCS + hodnotený vzrušujúcejšie ako NOVINKA [Acq1: t(32) = 2.99, p = 0.005; Acq2: t(32) = 5.97, p <0.001] a appCS + [Acq1: t(32) = 2.62, p = 0.013; Acq2: t(32) = 4.42, p <0.001] a o niečo vzrušujúcejšie ako CS– po prvej fáze akvizície [t(32) = 1.96, p = 0.058], ale po druhej fáze akvizície je to výrazne vzrušujúce [t(32) = 3.65, p = 0.001]. Na rozdiel od valenčných hodnotení sa vzrušenie appCS + nelíšilo od vzruchu CS - a NOVÉ (ps > 0.13) po fázach akvizície.
Podobne ako v prípade valenčných ratingov sa CS– a NEW nelíšili v ich vzrušení (ps> 0.07).
Rovnako ako v prípade valencie sa nezistili žiadne rozdiely pre appCS + asociovaný s čokoládou a appCS + asociovaný so slaným preclíkom [Acq1: t(31) = 0.26, p = 0.797; Acq2: t(31) = 0.33, p = 0.724].
Po vyhynutej fáze sa nezistili žiadne významné rozdiely v hodnotení vzrušenia (ps> 0.08).
Za ďalšie post-hoc t- testy porovnávajúce hodnotenia po habituačnej fáze, prvej a druhej akvizičnej fáze a extinkčnej fáze, pozri doplnkové materiály.
Odozva odštartovania
ANOVA pre akvizičné fázy vrátila hlavný efekt stimulu [F(2, 64) = 49.92, GG-ε = 0.964, p <0.001, čiastočné η2 = 0.609], ale nie vo fáze [F(1, 32) = 3.16, p = 0.085, čiastočné η2 = 0.090] a významná interakcia Stimulus × Fáza [F(2, 64) = 3.37, GG-ε = 0.875, p = 0.048, čiastočné η2 = 0.095]. Nasleduj t-testy odhalili výraznú potencovanú úľavu na avCS + v porovnaní s CS- počas prvého [t(32) = 3.27, p = 0.003] a druhý [t(32) = 4.00, p <0.001] fázy akvizície. Vyľakané reakcie na avCS + boli tiež významne potencované v porovnaní s odpoveďami na appCS +, opäť po oboch akvizíciách 1 [t(32) = 8.20, p <0.001] a akvizícia 2 [t(32) = 5.74, p <0.001]. Dôležité je, že veľkosť zľaknutia voči appCS + bola významne zoslabená v porovnaní s CS– počas oboch akvizícií 1 [t(32) = 6.34, p <0.001] a akvizícia 2 [t(32) = 2.91, p = 0.007]. Opäť a v súlade s hodnotením neboli zistené žiadne diferenčné odozvové reakcie pre appCS +, keď sú spojené s čokoládou alebo slanou preclíkou [Acq1: t(31) = 1.04, p = 0.309; Acq2: t(31) = 0.07, p = 0.947]. Počas fázy extinkcie sa nezistil žiadny významný účinok [F(3, 96) = 0.26, GG-ε = 0.906, p = 0.833, čiastočné η2 = 0.008].
Reakcia vodivosti kože (SCR)
Z ANOVA pre SCR počas dvoch fáz akvizície, stimuly hlavných účinkov [F(2, 54) = 18.04, GG-ε = 0.908, p <0.001, čiastočné η2 = 0.401] a fáza [F(1, 27) = 20.91, p <0.001, čiastočné η2 = 0.436] sa ukázalo ako významné, ale nie ich interakcia [F(2, 54) = 0.68, GG-ε = 0.637, p = 0.451, čiastočné η2 = 0.024]. Post-hoc t-testy ukázali výrazne vyššiu SCR na avCS + [t(27) = 6.46, p <0.001] a do aplikácie CS + [t(27) = 4.84, p <0.001] v porovnaní s CS–, zatiaľ čo účastníci vykazovali porovnateľnú SCR s avCS + a appCS + [t(27) = 0.64, p = 0.527]. Najmä sa nezistili žiadne rozdiely v hodnotách SCR voči čokoládovému appCS + a slanému preclíku appCS + [Acq1: t(26) = 2.55, p = 0.120; Acq2: t(26) = 1.29, p = 0.210]. Ako to bolo v prípade hodnotení a prekvapivej reakcie, nezistili sa žiadne významné účinky vo fáze vyhynutia [F(3, 81) = 0.28, GG-ε = 0.634, p = 0.743, čiastočné η2 = 0.010].
Diskusia
Cieľom tejto štúdie bolo previesť zistenia zvierat na ľudí pomocou klasickej apetitívnej paradigmy s primárnym zosilňovačom ako nepodmieneného stimulu (USA). Aby tak urobili, účastníci prišli do laboratória skoro ráno bez raňajok, aby sa ubezpečili, že majú hlad, a podľa ich preferencií sa kúsky čokolády alebo slané praclíky používali ako chutné USA. Počas akvizičnej fázy sa jeden geometrický tvar (avCS +) spojil s mierne bolestivým elektrickým šokom (averzívnym USA), iným tvarom (appCS +) s apetítom v USA a tretím tvarom (CS-), ani s apetítom USA, ani s USA. averzívne USA. Výsledky naznačujú úspešnú averzívnu a apetitívnu kondíciu na explicitnej verbálnej úrovni (tj hodnotenie), na implicitnej úrovni správania (tj úľaková odozva) a na fyziologickej úrovni (tj SCR). Konkrétne, avCS + v porovnaní s CS- vyvolalo viac negatívnych valenčných hodnotení, vyšších hodnotách vzrušenia, vyľakania úľavy a väčšej SCR. Najdôležitejšie je, že aplikácia appCS + v porovnaní s CS - spustila viac pozitívnych valenčných hodnotení, útlmu útlmu a väčšej SCR. Naše zistenia o averzívnom kondicionovaní boli v súlade s očakávaniami, keďže predchádzajúce štúdie zistili, že hrozba stimulovania stimulov (avCS +) je hodnotená ako averzívna, vyvoláva väčšie reakcie na strach a zvyšuje fyziologické vzrušenie (Fendt a Fanselow, 1999; Hamm a Weike, 2005; Andreatta a kol., 2010, 2013). Naše výsledky v oblasti apetítnej kondicionácie boli tiež v súlade s predchádzajúcimi štúdiami na ľuďoch a zvieratách, ktoré ukázali, že stimulačná predpovedná odmena (appCS +) je hodnotená ako pozitívna, inhibuje reakcie na strach a zvyšuje fyziologické vzrušenie (Koch a spol., 1996; Gottfried a kol., 2002; Kumar a kol., 2008; Klucken a kol., 2009, 2013; Austin a Duka, 2010; Prévost a kol., 2012). Podľa našich vedomostí je to prvá štúdia, ktorá demonštruje podmienený útlmový útlm u ľudí na podnet predpovedajúci primárnu odmenu. Dôležité je, že sme dokázali preniesť a potvrdiť výsledky štúdie na zvieratách (Koch a spol., 1996). Táto štúdia na zvieratách ukázala, že útlmový útlm u potkanov závisí od projekcie od NAcc (časť ventrálneho striata) k PnC. Preto útlm útlmu v našej štúdii by mohol znamenať NAcc aktivitu, ktorá je tiež v súlade s nálezmi fMRI (Gottfried a kol., 2002; Kumar a kol., 2008; Klucken a kol., 2009, 2013; Delgado a kol., 2011; Levy a Glimcher, 2011). Preto sme dospeli k záveru, že naša apetitívna paradigma kondicionovania bola úspešná, ako sa ukázalo ako explicitná (rating), tak aj implicitná (útlmová útlm) pozitívna valencia.
Okrem tohto nového, ale skôr predvídateľného zistenia si zaslúžia zmienku dva ďalšie zaujímavé výsledky. Po prvé, verbálne a fyziologické reakcie na appCS + sa oddelili. Po druhé, zistili sme rýchlejší zánik chutných CR v porovnaní s averzívnymi CR.
SCR indikuje sympatickú aktiváciu, ktorá sa zvyšuje na stimuly spojené s odmenou (appCS +) a stimulom asociovaným s hrozbou (avCS +). Tento výsledok je v súlade s predchádzajúcou štúdiou kondicionovania, v ktorej boli erotické obrázky použité ako apetitívne US (Klucken a kol., 2013) a štúdiu, ktorá odhaľuje porovnateľnú SCR s tágom, ktorý predpovedá peniaze a tágo, ktoré predpovedá averzívny hluk (Austin a Duka, 2010). SCR je orientačná reakcia súvisiaca s aktiváciou sympatického systému. Táto reakcia bola navrhnutá tak, aby odrážala prípravu behaviorálnej reakcie na motivačné hlavné udalosti (Bradley, 2009). Na základe toho si myslíme, že stimuly spojené s hrozbou a odmenou vyvolali prípravnú reakciu na následné reakcie správania. Inými slovami, potravinové a bolestivé signály iniciovali prípravu na prístupové a vyhýbavé správanie. Na rozdiel od vysokého fyziologického vzrušenia verbálne reakcie indikovali nízke vzrušenie stimulu spojeného s odmenou. Je možné, že verbálne vzrušenie môže byť skôr ovplyvnené vzrušujúcou povahou USA ako fyziologickou aktiváciou sama o sebe, V skutočnosti si myslíme, že appCS + vs. avCS + boli hodnotené ako nízko vs. vysoko vzrušujúce, pretože boli prepojené s nenápadnými a vysoko vzrušujúcimi USA. Bohužiaľ, nezískavali sme vzrušujúce ratingy pre USA, a preto bude na budúcich štúdiách, aby túto hypotézu otestovali explicitne.
Počas fázy zániku neboli dodané žiadne USA. To mohlo vyvolať nové inhibičné učenie nazývané zánik (pozri prehľad Milad a Quirk, 2012), a to ako s averzívnou, tak aj s apetitívnou CR. Je zrejmé, že na úrovni správania (tj úľaková odozva) a na fyziologických (tj SCR) úrovniach už neboli zistené žiadne diferenciálne reakcie na avCS +, appCS + a CS -. Podobne, explicitné vzrušenie podmieňovaných podnetov hodnotené po tom, ako sa fáza extinkcie vyrovnala na nízkej úrovni, čo naznačuje úspešné učenie sa zániku. AvCS + však bol stále hodnotený výrazne negatívnejšie ako CS-, zatiaľ čo appCS + valencia sa už nelíši od CS-valencie. Pomalší zánik averzívnej explicitnej odozvy by mohol byť spôsobený evolučným konzervativizmom, čo znamená, že signály hrozieb je veľmi ťažké zabudnúť, pretože neodpovedanie na hrozbu môže byť život ohrozujúce.
Zostáva ešte jedna otázka: prečo potom vyľakávacia reakcia (tj implicitná valencia) úplne zanikla počas fázy zániku? Po prvé, tieto reakcie boli vypočítané počas celého priebehu fázy. Preto je možné, že by boli diskriminačné CRs stále detegovateľné počas prvých pokusov fázy zániku. Výskumným spôsobom sme sledovali túto hypotézu a pozreli sme sa na odozvy aj na SCR počas fázy zániku (pozri Dodatočný materiál). Hoci sme nenašli významné rozdiely, pozorovali sme mierne vyššiu vyľakávaciu veľkosť pre avCS + v porovnaní s CS- a novým kontrolným stimulom. Tiež sme pozorovali o niečo viac prekvapujúci útlm k appCS + v porovnaní s CS- a novým kontrolným stimulom na samom začiatku fázy vyhynutia, ktorá však v niekoľkých pokusoch zanikla. SCR na appCS + klesla už po 2nd skúške zániku, zatiaľ čo SCR na avCS + zostali vyššie pre takmer všetky zániku v porovnaní s SCR na NOVÉ. Hoci by sme mali tieto výsledky interpretovať s veľkou opatrnosťou, zdalo sa, že prekvapujúce reakcie sú paralelné s hodnotami valencie. Okrem toho, aj prekvapujúce reakcie a SCR ďalej podporujú myšlienku evolučného konzervativizmu voči hrozbám.
Nakoniec musíme uznať niektoré obmedzenia tejto štúdie. Po prvé, kvôli technickému problému sme nemohli nahlásiť hodnotenie príjemnosti (a vzrušenia) pre čokoládu a slaný preclík. Avšak, chutné CR naznačujú, že účastníci skutočne zažili dva USA ako apetít. Po druhé, trvanie averzívne a chutné USA veľmi líšili. Bolestivý úraz elektrickým prúdom bol teda dosiahnutý pri vykompenzovaní avCS + a trval presne 200 ms, zatiaľ čo čokoláda a slaný preclík boli predstavené účastníkom okolo 2 s po nástupe appCS + a trvanie bolo nedefinovateľné, pretože záviselo na tom, ako rýchlo jednotlivec ich jedol. Voľba dodať chutné USA týmto spôsobom bola založená na predchádzajúcej štúdii u ľudí (Prévost a kol., 2012). Bolo by však metodicky elegantnejšie dodať chutné USA, ktoré by boli porovnateľnejšie s averzívnym USA, ako je napríklad popíjanie džúsu alebo zázvorového piva pri aplikácii appCS + offset. S cieľom kompenzovať tento veľký percepčný rozdiel sme prezentovali geometrické tvary v spojení s vizuálnym podnetom symbolizujúcim USA. Po tretie, nemôžeme jednoznačne vylúčiť možnosť, že rýchlo zaniknuté fyziologické reakcie vo fáze zániku sú spôsobené metodologickým aspektom. V skutočnosti neboli vizuálne podnety v tejto fáze prezentované v spojení so symbolom USA, ako to bolo v prípade akvizičných fáz. Je možné, že samotná prezentácia CS mohla ovplyvniť CR a z tohto dôvodu neboli počas prvých skúšok zániku viditeľné žiadne významné rozdiely. Naše rýchle hasiace reakcie sú však v súlade s vyhasnutými odpoveďami v predchádzajúcej štúdii, v ktorej bola CS + (tvár) počas akvizície prezentovaná v zlúčenine s USA (scream), ale nie počas extinkcie (Lissek a kol., 2008).
Na záver sme zistili úspešné averzívne a chutné podmienené reakcie na podnet spojený s hrozbou a podnetom spojeným s odmenou, resp. Zaujímavé je, že explicitné (hodnotenia) a implicitná (odrazová) úroveň odpovedí pracovali synergickým spôsobom v tom, že avCS + bola hlásená ako negatívna a indukovaná potenciácia úľavy a appCS + bol zaznamenaný ako pozitívny a indukovaný útlm. Okrem toho, explicitné (hodnotenia) a fyziologické (SCR) vzrušenie appCS + disociované, odrážajúce dva odlišné procesy.
Vyhlásenie o konflikte záujmov
Autori vyhlasujú, že výskum bol vykonaný bez obchodných alebo finančných vzťahov, ktoré by mohli byť interpretované ako potenciálny konflikt záujmov.
Poďakovanie
Táto práca bola podporená Centrom kolaboratívneho výskumu „Strach, úzkosť a úzkostné poruchy“, SFB-TRR 58, projekt B1.
Doplnkový materiál
Doplnkový materiál tohto článku sa nachádza na adrese: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
Referencie
Andreatta, M., Mühlberger, A., Glotzbach-Schoon, E. a Pauli, P. (2013). Predpovedateľnosť bolesti mení valenčné hodnotenia stimulu súvisiaceho s úľavou. Predná. Syst. Neurosci, 7: 53. doi: 10.3389 / fnsys.2013.00053
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Andreatta, M., Mühlberger, A., Yarali, A., Gerber, B. a Pauli, P. (2010). Rozpor medzi implicitnou a explicitnou podmienenou valenciou po učení úľavy od bolesti u ľudí. Proc. Biol. sci, 277, 2411 – 2416. doi: 10.1098 / rspb.2010.0103
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Austin, AJ, a Duka, T. (2010). Mechanizmy pozornosti pre apetitívne a averzívne výsledky v Pavlovianskej úprave. Behave. Brain Res, 213, 19 – 26. doi: 10.1016 / j.bbr.2010.04.019
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Blumenthal, TD, Cuthbert, BN, Filion, DL, Hackley, S., Lipp, OV a van Boxtel, A. (2005). Správa výboru: Usmernenia pre elektromyografické štúdie zamerané na ľudské prekvapenie. psychofyziológia 42, 1-15. dva: 10.1111 / j.1469-8986.2005.00271.x
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Bouton, ME, a Peck, CA (1989). Kontextové vplyvy na kondicionovanie, zánik a obnovenie v chutnej kondičnej príprave. Anim. Učiť. behave, 17, 188 – 198. doi: 10.3758 / BF03207634
CrossRef Plný text | Študovňa Google
Bradley, MM (2009). Prírodná selektívna pozornosť: orientácia a emócie. psychofyziológia 46, 1-11. dva: 10.1111 / j.1469-8986.2008.00702.x
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Clark, JJ, Hollon, NG a Phillips, PEM (2012). Pavlovovské systémy oceňovania vo vzdelávaní a rozhodovaní. Akt. Opin. Neurobiol, 22, 1054 – 1061. doi: 10.1016 / j.conb.2012.06.004
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Delgado, MR, Jou, RL a Phelps, EA (2011). Neurónové systémy, ktoré sú základom averzívnej kondície u ľudí s primárnymi a sekundárnymi zosilňovačmi. Predná. Neurosci, 5: 71. doi: 10.3389 / fnins.2011.00071
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Dickinson, A. a Balleine, B. (1994). Motivačná kontrola cielenej akcie. Anim. Učiť. behave, 22, 1 – 18. doi: 10.3758 / BF03199951
CrossRef Plný text | Študovňa Google
Fendt, M., a Fanselow, MS (1999). Neuroanatomický a neurochemický základ podmieneného strachu. Neurosci. Biobehav. otáčka, 23, 743 – 760. doi: 10.1016 / j.ijpsycho.2012.09.006
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Gottfried, JA, O'Doherty, J. a Dolan, RJ (2002). Chuťové averzné čuchové učenie u ľudí sa študovalo pomocou funkčnej magnetickej rezonancie súvisiacej s udalosťami. J. Neurosci, 22, 10829 – 10837. K dispozícii online na adrese: http://www.jneurosci.org/content/22/24/10829.full
Grillon, C., Baas, JM, Cornwell, B. a Johnson, L. (2006). Kontextové podmieňovanie a vyhýbanie sa správaniu v prostredí virtuálnej reality: účinok predvídateľnosti. Biol. psychiatrie 60, 752-759. dva: 10.1016 / j.biopsych.2006.03.072
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Hamm, AO a Weike, AI (2005). Neuropsychológia učenia strachu a regulácia strachu. Int. J. Psychophysiol, 57, 5 – 14. doi: 10.1016 / j.ijpsycho.2005.01.006
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Klucken, T., Schweckendiek, J., Merz, CJ, Tabbert, K., Walter, B., Kagerer, S., et al. (2009). Nervové aktivácie získavania podmieneného sexuálneho vzrušenia: účinky podmieneného uvedomenia a pohlavia. J. Sex. med, 6, 3071-3085. dva: 10.1111 / j.1743-6109.2009.01405.x
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Klucken, T., Wehrum, S., Schweckendiek, J., Merz, CJ, Hennig, J., Vaitl, D. a kol. (2013). Polymorfizmus 5-HTTLPR je spojený so zmenenými hemodynamickými reakciami počas predvídateľnej kondicionácie. Hum. Brain Mapp, 34, 2549 – 2560. doi: 10.1002 / hbm.22085
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Koch, M. (1999). Neurobiológia vyľakania. Prog. Neurobiol. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Koch, M., Schmid, A., a Schnitzler, H.-U. (1996). Potešenie-útlm vyľakania je narušený léziami nucleus accumbens. Neuroreport 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Krohne, HW, Egloff, B., Kohmann, C.-W., a Tausch, A. (1996). Untersuchungen mit einer deutschen version of „Pozitívny a negatívny vplyv“ (PANAS). Diagnostica 42, 139-156.
Kumar, P., Waiter, G., Ahearn, T., Milders, M., Reid, I. a Steele, JD (2008). Abnormálne časové rozdiely odmeňovanie-učenie signály pri veľkej depresii. Mozog 131, 2084 – 2093. doi: 10.1093 / brain / awn136
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Laux, L., Glanzmann, P., Schaffner, P. a Spielberger, CD (1981). Das State-Trait Angstinventar, Weinheim: Beltz test.
Levy, DJ a Glimcher, PW (2011). Porovnávanie jabĺk a pomarančov: použitie reprezentácie reprezentatívnej hodnoty, ktorá je špecifická pre odmenu a odmenu v mozgu. J. Neurosci, 31, 14693 – 14707. doi: 10.1523 / JNEUROSCI.2218-11.2011
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Lissek, S., Levenson, J., Biggs, AL, Johnson, LL, Ameli, R., Pine, DS, et al. (2008). Zvýšené podmieňovanie strachu na spoločensky relevantné nepodmienené stimuly v sociálnej úzkostnej poruche. Am. J. Psychiatry 165, 124-132. dva: 10.1176 / appi.ajp.2007.06091513
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Martin-Soelch, C., Linthicum, J., a Ernst, M. (2007). Chutné kondicionovanie: neurónové bázy a implikácie pre psychopatológiu. Neurosci. Biobehav. otáčka, 31, 426 – 440. doi: 10.1016 / j.neubiorev.2006.11.002
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
McDannald, MA, Lucantonio, F., Burke, KA, Niv, Y. a Schoenbaum, G. (2011). Ventrálna striatum a orbitofrontálna kôra sú potrebné na modelovanie, ale nie na modeli, posilňujúce učenie. J. Neurosci, 31, 2700 – 2705. doi: 10.1523 / JNEUROSCI.5499-10.2011
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Milad, MR a Quirk, G. (2012). Strach vyhynutia ako model pre translačné neurovedy: desať rokov pokroku. Annu. Psychol, 63, 129 – 111. doi: 10.1146 / annurev.psych.121208.131631
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Pavlov, IP (1927). Kondicionované reflexy: skúmanie fyziologickej aktivity mozgovej kôry, London: Oxford University Press.
Prévost, C., Liljeholm, M., Tyszka, JM a O'Doherty, JP (2012). Neurálne koreláty špecifického a všeobecného prenosu pavloviánov do inštrumentálov v rámci subregiónov ľudských amygdalarov: štúdia fMRI s vysokým rozlíšením. J. Neurosci, 32, 8383 – 8390. doi: 10.1523 / jneurosci.6237-11.2012
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Tranel, D. a Damasio, H. (1994). Neuroanatomické korelácie elektrodermálnych reakcií vodivosti kože. psychofyziológia 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
PubMed Abstrakt | CrossRef Plný text | Študovňa Google
Kľúčové slová: klasické podmieňovanie, odmena, trest, úľavový reflex, reakcia vodivosti kože
Citácia: Andreatta M a Pauli P (2015) Chutné vs. Aversive klimatizácia u ľudí. Predná. Behave. Neurosci. 9: 128. dva: 10.3389 / fnbeh.2015.00128
Prijaté: 11 február 2015; Prijaté: 05 May 2015;
Pridané: 19 May 2015.
strih:
Niels Birbaumer, University of Tuebingen, Nemecko
Hodnotené:
Tim KluckenUniverzita Justus Liebig Giessen, Nemecko
Alessandro Angrilli, Univerzita Padova, Taliansko
Copyright © 2015 Andreatta a Pauli. Toto je článok s otvoreným prístupom distribuovaný podľa podmienok Creative Commons Attribution License (CC BY), Používanie, distribúcia alebo reprodukcia na iných fórach je povolené za predpokladu, že pôvodný autor (autorov) alebo poskytovateľ licencie je pripísaný a že pôvodná publikácia v tomto časopise je citovaná v súlade s prijatou akademickou praxou. Nepoužíva sa žiadna distribúcia alebo reprodukcia, ktorá nie je v súlade s týmito podmienkami.
* Korešpondencia: Marta Andreatta, Katedra psychológie (biologická psychológia, klinická psychológia a psychoterapia), Univerzita vo Würzburgu, Marcusstraße 9-11, D-97070 Würzburg, Nemecko, [chránené e-mailom]
