 Stephanie E. Tedford
Stephanie E. Tedford- 1Institutionen för farmakologi, Centrum för kompulsiv beteende och missbruk, Rush University Medical Center, Chicago, IL, USA
- 2Institutionen för farmakologi, Rush University Medical Center, Chicago, IL, USA
- 3Institutionen för psykiatri, Rush University Medical Center, Chicago, IL, USA
Patologiskt spel är en manifestation av impulskontrollsjukdomar. De biologiska grunden för dessa sjukdomar förblir svimmade och behandlingen är långt ifrån idealisk. Djurmodeller av impulskontrollsjukdomar är ett kritiskt forskningsverktyg för att förstå detta tillstånd och för medicinutveckling. Modellering av sådant komplicerat beteende är skrämmande, men genom dess dekonstruktion har forskare rekapitulerat i djurens kritiska aspekter av spelande. En aspekt av spelande är kostnads / nytta beslutsfattande där man väger de förväntade kostnaderna och de förväntade fördelarna med en handlingsplan. Risk / belöning, fördröjningsbaserat och ansträngningsbaserat beslutsfattande utgör alla kostnader / nytta val. Dessa egenskaper studeras hos människor och har översatts till djurprotokoll för att mäta beslutsprocesser. Traditionellt är den positiva förstärkaren som används i djurstudier mat. Här beskriver vi hur intrakraniell självstimulering kan användas för kostnad / nytta beslutsfattande uppgifter och överblick våra senaste studier som visar hur farmakologiska terapier förändrar dessa beteenden i laboratorie råttor. Vi föreslår att dessa modeller kan ha värde vid screening av nya föreningar för förmågan att främja och förebygga aspekter av spelbeteende.
Beskrivning
Problem eller missbrukande spel, inklusive det extrema tillståndet betecknat patologiskt spelande, kännetecknas av beteenden som ofta kvarstår under längre perioder. Problemspel kan få en väsentlig negativ inverkan på personligt, professionellt och ekonomiskt välbefinnande. Under de senaste två decennierna har spelmöjligheterna ökat genom förändringar i lagstiftningen och införandet av nya arenor (t.ex. internetspel). Följaktligen har förekomsten av problemgambling ökat. Det finns inga FDA-godkända behandlingar för denna sjukdom, och det är därför kritiskt att bättre förstå dessa beteenden för att utveckla effektiva terapier.
Problemspel är ett komplicerat fenomen, vilket inkluderar ökade impulsiva beslutsfattande (Alessi och Petry, 2003; Dixon et al., 2003; Holt et al., 2003; Kraplin et al., 2014) som härrör från ofördelaktiga utvärderingar av kostnader / förmåner. Kliniska bedömningar av beslutsfattande, som ofta använder undersökning och interaktiva datorbaserade verktyg, har bidragit till att bestämma suboptima beslutsprofiler i olika patologier, inklusive patologiska spelare (Ledgerwood et al., 2009; Madden et al., 2009; Michalczuk et al., 2011; Petry, 2011; Miedl et al., 2012). Kliniska bedömningar görs ofta på grundval av tre olika, om än överlappande aspekter av kostnads / nytta beslutsfattande, inklusive följande: (i) risken för att få en belöning (risk / belöning beslutsfattande), ii) en Fördröjning upplevd före belöningsleverans (fördröjningsbaserat beslutsfattande), och (iii) Mängden ansträngning som krävs för att erhålla belöning (ansträngningsbaserat beslutsfattande). Flera uppgifter har utvecklats för att mäta dessa kritiska egenskaper av suboptimal beslutsfattande för att ytterligare förstå processer som innefattar problemspel. I dessa uppgifter väljer ämnet mellan en liten och stor belöning, som varje förknippas med specifika responsförhållanden. I risk / belöning beslutsfattande väljer man ämnen mellan en liten belöning som levereras konsekvent vid höga sannolikheter (t.ex. 100% sannolikhet för att ta emot $ 10) och en stor belöning som levereras med varierande sannolikheter (t.ex. 10-80 % sannolikhet för att ta emot $ 100). I kliniska och prekliniska studier är frånvaron av en förväntad belöning en aversiv händelse som framkallar motsvarande fysiologiska svar (Douglas och Parry, 1994; Papini och Dudley, 1997). Preference för det större alternativet "riskabelt" över det lilla, vissa alternativet anses anses återspegla suboptimal risk / belöning beslutsfattande och har rapporterats för flera mänskliga patologier som uppvisar förbättrad impulsivitet (Reynolds et al., 2004; Rasmussen et al., 2010; Dai et al., 2013). I fördröjningsbaserat beslutsfattande (dvs. fördröjning av diskontering, ett mått på impulsivt val) levereras den lilla belöningen snart efter att valet väljs, medan den stora belöningen levereras efter en variabel fördröjning (t.ex. $ 10 nu eller $ 100 i 2 veckor). Individer som uppvisar hög impulsivitet visar preferens för omedelbart tillgängliga belöningar (även om de är mindre), över försenade belöningar (även om de är större), även om det senare alternativet kan vara mer fördelaktigt för individen (Crean et al., 2000; Reynolds et al., 2004; Bickel et al., 2012). I ansträngningsbaserat beslutsfattande väljer ämnet mellan en liten belöning som levereras efter små mängder ansträngning eller ett stort belöning som levereras efter att större ansträngningar har utövats. I den här uppgiften bestäms individuell preferens för det höga insatsen / stora belöningsalternativet och den "punkt" där personen växlar till alternativet låg insats / litet belöning. Studier av ansträngningsbaserat beslutsfattande hos människor som spelats har ännu inte genomförts, men skulle vara av stort intresse för att bedöma kognitiv funktion i denna population.
Beslutsfattande protokoll som används vid kliniska bedömningar kan modifieras för att studera beslutsfattande i laboratorieråttor, och dessa modeller är kritiska för att utforska beteende- och neurofarmakologiska aspekterna av patologiskt spelande. Vid råttor kan beslutsfattandet utvärderas genom att djuret placeras i en operant konditioneringskammare och tillåter djuret att välja mellan två hävstångar (eller två nospinnar) som görs tillgängliga samtidigt. Den etablerade belöningsmodaliteten för den positiva förstärkaren i dessa gnagareuppgifter är mat (Stopper och Floresco, 2011; Eubig et al., 2014). Vi diskuterar här en ny metod som används i vårt laboratorium, som använder direkt elektrisk stimulering av hjärnbelöningsbanor (intrakranial självstimulering, ICSS) för att bedöma kostnads / nytta beslutsfattande hos råttor och monoaminerga neurotransmittors bidrag i beslutsfattandet (Rokosik och Napier, 2011, 2012; Tedford et al., 2012; Persons et al., 2013).
Intrakraniell självstimulering
En operant förstärkare är en stimulans, som när den är beroende av en viss handling ökar sannolikheten för återkomsten av den åtgärden. Intrakranial självstimulering (ICSS) är ett operant beteende där djur själv administrerar elektrisk stimulering till hjärnregioner som är kända för att vara involverade i positiv förstärkning. ICSS studerades först i 1950s när James Olds och Peter Milner (Olds och Milner, 1954) bestämde att råttor återkommande skulle återvända till en plats i en låda där de fick elektrisk stimulering till belöningsrelaterade regioner i hjärnan. De gjorde det möjligt för råttor att arbeta för denna elektriska hjärnstimulering (EBS) genom att reagera på en operant manipulandum (t ex tryck på en spak, snurra ett hjul) (Olds och Milner, 1954). Upptäckten av denna teknik har bidragit till kartläggning av belöningsbanor i hela hjärnan, och medan det finns många regioner i hjärnan som kan användas för att stödja ICSS (Olds och Milner, 1954; Wise och Bozarth, 1981; Klokt, 1996), är det väl dokumenterat att stimulering av medial forebrain bundle (MFB) främjar djupa och tillförlitliga beteendeutgångar (Corbett och Wise, 1980; Pirch et al., 1981; McCown et al., 1986; Tehovnik och Sommer, 1997). Stimuleringsströmparametrar kan manipuleras för att påverka förstärkningsvärdet för EBS och därför förändra ICSS-beteende. Dessa parametrar inkluderar intensiteten (dvs. ampere) av den elektriska strömmen och den aktuella frekvensen (dvs hertz). Höjningar i båda parametrarna resulterar vanligen i ökad excitation av de belöningsrelevanta neuronerna som stimuleras, antingen genom att öka antalet neuroner involverade av stimulansen (amperes) (Keesey, 1962; Wise et al., 1992) eller genom att öka frekvensen i vilken en bestämd population av neuronbrand (hertz) (Wise and Rompre, 1989; Klokt, 2005). Manipuleringar av nuvarande intensitet förändrar antalet aktiverade neuroner, dvs större strömintensiteter påverkar en större population av neuroner än mindre strömningar. Således, när denna parameter hålls konstant, är populationen av neuroner som upphetsas av EBS relativt lika oberoende av aktuell frekvens. Stimuleringsparametervariabeln som valts för dessa protokoll är aktuell frekvens, eftersom det här valet tillåter oss att manipulera avfyrningshastigheten för samma grupp av neuroner med minimal effekt på tiden eller rymden för stimuleringsintegration. Genom att manipulera dessa EBS-parametrar har vi utvecklat sofistikerade modeller av kostnads / nytta beslutsfattande som använder ICSS (Rokosik och Napier, 2011, 2012; Tedford et al., 2012; Persons et al., 2013). Denna ansökan representerar en radikal avvikelse från den traditionellt använda förstärkande stimulansen (dvs. mat) i uppgifter som bedömer beslutsfattandet hos gnagare. ICSS kan ge flera experimentella fördelar jämfört med traditionella förstärkningsmetoder. För att underlätta operant att svara på mat är det dagliga intaget ofta begränsat (Feja och Koch, 2014; Hosking et al., 2014; Mejia-Toiber et al., 2014). Denna praxis kan konfrontera utfallsåtgärder, eftersom det finns väsentliga överlappningar i de neurobiologiska systemen som förändras under kronisk matrestriktion och de som medger impulsiv beslutsfattande (Schuck-Paim et al., 2004; Minamimoto et al., 2009). Dessutom förstärks djuren förstärkta med mat alltmer under en session, vilket minskar värdet av matförstärkning (Bizo et al., 1998), även om denna effekt kan vara beroende av förstärkare storlek (Roll et al., 1995). I motsats till matförstärkning förblir förstärkningsvärdet för EBS stabilt under en session, vilket möjliggör mer omfattande och konsekvent beteendebedömning (Trowill et al., 1969). Denna funktion möjliggör att test sessioner uppträder upprepade gånger under en dag, vilket kan vara fördelaktigt när man studerar effekterna av farmakologiska terapier, särskilt kronisk läkemedelsbehandling. Våra publicerade riskbedömningsstudier (diskuteras nedan) genomfördes flera gånger om dagen under behandling med kronisk dopaminagonist (pramipexol). Vi föreslår att denna processförmån är mer tillämplig på det mänskliga tillståndet och därigenom ger förbättrade translationella resultat. Hittills har liknande studier av dopaminagonister på impulsiv beslutsfattande med hjälp av livsmedelsbelöning endast utvärderat akuta läkemedelsbehandlingar (St Onge och Floresco, 2009; Zeeb et al., 2009; Madden et al., 2010; Johnson et al., 2011; Koffarnus et al., 2011) och det kommer att vara av stort intresse att jämföra beteendemässiga resultat efter både akut och kronisk läkemedelsbehandling mellan dessa olika förstärkare. Medan ICSS ger flera fördelar jämfört med matförstärkning, presenterar ICSS också flera nackdelar. ICSS kräver till exempel invasiv hjärnkirurgi och återhämtning, och ohämmade huvudsteg kan leda till förlust av ämnen i hela beteendeparadigmet. Trots dessa nackdelar hävdar vi att ICSS är ett livskraftigt alternativ till matförstärkning och ger stora fördelar med matförstärkning i dessa beteendeuppgifter.
Kostnads / nytta beslutsfattande uppgifter kräver att man väljer mellan alternativ som är förknippade med varierande belöningsnivåer. Följaktligen bör förstärkare som används i dessa uppgifter visa förmågan att framställa sådana förändringar i belöningsstyrkan och därefter måste råttor kunna diskriminera mellan alternativet litet förstärkare (SR) och stor förstärkare (LR). I förfaranden som använder matförstärkning uppnås detta genom att ändra antalet matpellets erhållna efter ett svar. I ICSS kan EBS varieras genom att ändra stimulans aktuell intensitet eller aktuell frekvens. Figur 1 illustrerar hävstångsreaktion erhållen när strömstyrkan varieras (dvs. den aktuella frekvensen hölls konstant, Figur 1A) eller när den aktuella frekvensen varieras (dvs den aktuella intensiteten hölls konstant; Figur 1B). När någon av parametrarna förändras uppvisar råttor måttlig häftning för små EBS-värden och visar ökade hävstryckningshastigheter för stora EBS-värden, vilket tyder på att förstärkningsvärdet för den större stimulansen är större (oberoende av huruvida aktuell intensitet eller frekvens manipuleras). EBS kan därför skräddarsys för den lilla och stora förstärkaren som är nödvändig för kostnads / nytta beslutsfattande protokoll. Dessa förstärkningsvärden kan bestämmas i enskilda råttor genom att generera stabila svänghastighetsresponskurvor för varje djur (Rokosik och Napier, 2011, 2012). Alternativt kan en populationskurva genereras från en grupp av råttor från vilka ett standardiserat SR- och LR-värde kan bestämmas (Tedford et al., 2012; Persons et al., 2013). Detta senare tillvägagångssätt ger ett mer tidseffektivt och ändå tillförlitligt sätt att härleda SR och LR. I en andra serie studier använde vi antingen manipuleringar med aktuell intensitet eller frekvens för att fastställa SR / LR-värden i en sannolikhetsdiskonteringsuppgift (dvs. risk / belöning beslutsfattande). Förändringar i nuvarande intensitetsförstärkningsvärden (dvs. den aktuella frekvensen hölls konstant) och nuvarande frekvensvärden (dvs den aktuella intensiteten hölls konstant) båda ger betydande diskonteringsbeteende hos råttor (figurer 1C, D). Baserat på del av diskonteringskurvens branthet bestämdes nuvarande frekvens att den var lämplig parameter för att manipulera armeringsvärden. När det väl har fastställts att råttor kan skilja mellan de standardiserade strömfrekvenser som används för SR och LR, kan de testas i något av våra ICSS-medierade beslutsfattande paradigmer: (i) risk / belöning beslutsfattande (Rokosik och Napier, 2011, 2012), (ii) fördröjningsbaserat beslutsfattande (Tedford et al., 2012), eller (iii) ansträngningsbaserat beslutsfattande (Persons et al., 2013).
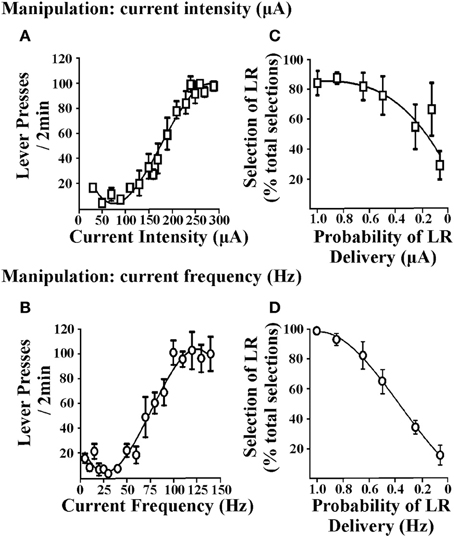
Figur 1. Effekter av hjärnstimuleringsparametrar på tryckpressrespons och sannolikhetsdiskontering. De två EBS-parametrarna som testades var nuvarande intensitet och strömfrekvens. Rattarmen pressad för EBS (i ett fast förhållande-1-schema för förstärkning) där varje 2-min, en parameter för EBS manipulerades och den andra parametern hölls konstant. (A) Manipulering av nuvarande intensitet. Nuvarande intensiteter som sträcker sig från 10 till 350 μA presenterades i slumpmässig ordning (n = 6); aktuell frekvens hölls vid 100 Hz. (B) Manipulering av aktuell frekvens. Aktuella frekvenser från 5 till 140 Hz presenterades i slumpmässig ordning (n = 3); Nuvarande intensitet hölls konstant på en nivå som var individualiserad och bestämd i tidigare träningssessioner. Manipulering av strömintensitet eller strömfrekvens resulterade i liknande mönster av hävstångsreaktion. Data visas som medelvärde ± som för de senaste tre på varandra följande sessionerna. Råttor utbildades därefter i sannolikhetsdiskonteringsuppgiften och värdena för de små och stora förstärkarna bestämdes individuellt för varje djur genom att beräkna de effektiva stimulansströmsintensiteterna och strömfrekvenserna erhållna från EBS vs handtryck-svarkurvan som framkallade 60 och 90% av maximal tryckpressresponshastighet, respektive. Varierar storleken på aktuell intensitet (C) eller aktuell frekvens (D) resulterade i diskontering av den stora förstärkaren (LR), eftersom sannolikheten för leverans minskade (dvs. minskning i procent av valet som var associerat med LR över totala val). Data visas som medelvärde ± som för dag 1 av diskontering med aktuell intensitet och 2-dagar med diskontering med användning av aktuell frekvens. Figur modifierad från Rokosik och Napier (2011) och reprinted med tillstånd från utgivaren.
Validera användningen av ICSS för att utvärdera impulsivitetsåtgärder och beslutsfattande
Utvecklingen av nya djurmodeller kräver noggrant överväganden om validitet. Vid utformningen av dessa ICSS-medierade beslutsuppgifter har vi alltså strävat efter att verifiera ansiktet och konstruera giltigheten och för att fastställa sannolikheten för förutsägbar validitet.
Ansiktsgiltighet avser den grad i vilken ett test subjektivt förefaller mäta det avsedda fenomenet. Utformningen av varje ICSS-medierad beslutsfattande uppgift baserades på nuvarande protokoll som användes hos människor för fördröjning och sannolikhetsdiskontering (Rasmussen et al., 2010; Leroi et al., 2013) och andra insatsbaserade beslutsfattande uppgifter (Treadway et al., 2009; Buckholtz et al., 2010; Wardle et al., 2011). På människor är åtgärderna för kostnads / nytta beslutsfattande härledda från att be enskilda att välja mellan flera alternativ som finns tillgängliga med specifika händelser som ställs på varje urval (dvs. risk, fördröjning eller ansträngning). Vi efterliknar detta scenario genom att presentera råttor med två samtidigt utvidgade hävstångar, varvid ett urval av antingen hävarmen är förknippat med små eller större belöningar som också levereras under speciella parametrar av beredskap. Således visar alla våra ICSS-medierade beslutsuppgifter uppgifter om ansikte.
Konstruktionsgiltighet avser paradigmets förmåga att noggrant bedöma vad den föreslår att mäta. I risk / belöning och fördröjningsbaserat beslutsfattande minskar preferensen för den stora belöningen som sannolikheten för leverans sänks, eller förseningen mot belöningsleverans ökar respektive. I ansträngningsbaserat beslutsfattande visar individer första preferensen för alternativet hög insats / stort belöning när ansträngningen för den stora belöningen anses rimlig. Ett skifte i stället för den låga ansträngningen / lilla belöningen observeras när den höga ansträngningen inte längre är värda energiförbrukningen. Det är väl dokumenterat att gnagare uppvisar liknande mönster av risk / belöning, fördröjningsbaserad och ansträngningsbaserad beslutsfattande jämfört med människor (Rachlin et al., 1991; Buelow och Suhr, 2009; Jimura et al., 2009), och vi har observerat dessa profiler i alla våra uppgifter (Rokosik och Napier, 2011, 2012; Tedford et al., 2012; Persons et al., 2013) (se till exempel Figur 2).
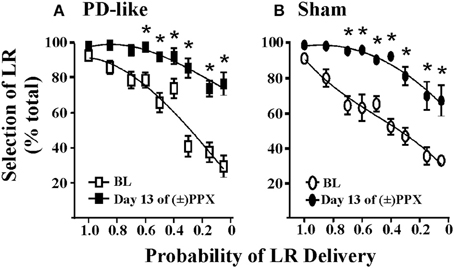
Figur 2. Effekter av pramipexol på risk / belöning beslutsfattande med hjälp av en sannolikhetsdiskonteringsuppgift. Kronisk (±) PPX minskar diskontering i PD-liknande (A) och sham kontroll (B) råttor. Kortfattat beskriver uppgiften PD-liknande (n = 11) och sham kontroll (n = 10) råttor utbildades i sannolikhetsdiskonteringsuppgiften med hjälp av ICSS. Sannolikheter i samband med leverans av den stora förstärkaren (LR) presenterades i en pseudo-randomiserad ordning. När ett stabilt beteende observerades behandlades råttor kroniskt med två gånger dagliga injektioner av 2 mg / kg (±) PPX för 13 dagar. Data som visades samlades in från den tidpunkt då vi observerade toppeffekten på den sista behandlingsdagen (dvs 6 h efter injektionen) och jämförs med förbehandlingsbaslinjen (BL). Visad är det procentuella valet av LR (dvs. frivilligt förhållande) jämfört med sannolikheten för att LR levererades. En tvåvägs rmANOVA med post hoc Newman-Keuls avslöjade signifikanta ökningar i% urval av osäker, LR efter kronisk PPX-behandling (*p <0.05) för både PD-liknande och sham-råttgrupper. Även om gruppgenomsnittet indikerar en PPX-inducerad ökning av suboptimal risk / belöningsbeslut, visade två råttor i varje grupp mindre än 20% ökning från baslinjen vid den lägsta testade sannolikheten; därför verkade vissa råttor vara okänsliga för läkemedlets förmåga att modifiera sannolikhetsdiskontering. Figur modifierad från Rokosik och Napier (2012) och reprinted med tillstånd från utgivaren.
Prediktiv giltighet avser modellernas förmåga att förutse framtida relationer, och vi föreslår att våra modeller kan användas för att förutsäga kapaciteten hos nya farmakologiska behandlingar för att förändra beslutsfattandet mellan kostnads och nytta. Genom att visa bevis på koncept genom att replikera effekterna av farmakologiska medel på beslutsbeteenden som redan har fastställts hos människor föreslår vi att våra modeller kan vara effektiva för att förutsäga hur andra läkemedel kan förmedla dessa beteenden på kliniken . Till exempel visar en delmängd av patienter med Parkinsons sjukdom (PD) som behandlas med dopaminagonistterapier en ökad förekomst av spelbeteende (Weintraub et al., 2010) och ökad diskontering i fördröjningsbaserat beslutsfattande (Housden et al., 2010; Milenkova et al., 2011; Voon et al., 2011; Leroi et al., 2013; Szamosi et al., 2013). Således har vårt laboratorium utgått för att modifiera PD hos råttor och studera effekterna av pramipexol, en vanlig dopaminagonist associerad med spelbeteenden (Weintraub et al., 2010) om kostnads / nytta beslutsfattande i råttan med hjälp av sannolikhetsdiskonteringsuppgiften (risk / belöning beslutsfattande) (Rokosik och Napier, 2012). För att göra detta gjordes råttor "PD-liknande" genom selektiv lesion av dopaminerga terminaler inom dorsolateral striatum via bilaterala infusioner av 6-OHDA, medan kontrollråttor fick infusioner av 6-OHDA-vehikeln (Rokosik och Napier, 2012). Neuroner i den dorsolaterala striatumen av endast de 6-OHDA-behandlade råttorna visar en minskning av tyrosinhydroxylas (Rokosik och Napier, 2012), en markör för dopamin. PD-liknande råttor uppvisar motorstörningar som liknar människor med tidigt stadium PD, som kan reverseras dosberoende med pramipexolbehandling. Dosen av pramipexol som vi administrerade för att studera risker / belöning beslutsfattande lindrar motoriska underskott och är således terapeutiskt relevant (Rokosik och Napier, 2012). Medan vi inte finner någon skillnad i "riskabelt" beteende mellan kontrollrotter och PD-liknande råttor ökar kronisk pramipexolbehandling valet av riskabelt LR i båda grupperna av råttor när sannolikheten för leverans var liten (Figurer 2A, B), vilket indikerar att pramipexol inducerar suboptimal risk / belöning beslutsfattande. Dessa data överensstämmer med studier som har utvärderat effekterna av pramipexol hos människor (Spengos et al., 2006; Pizzagalli et al., 2008; Riba et al., 2008). Ändå slår vi fram våra prediktiva giltighetsmodeller för att indikera andra farmakologiska medel som kan medföra kostnad / nytta beslutsfattande hos människor.
Vi har också testat mirtazapin, en atypisk anti-depressant, i den ansträngningsbaserade beslutsfattandet. Behavioral missbruk och drogmissbruk delar många överlappande egenskaper, inklusive suboptimal beslutsfattande, och nya studier på människor och icke-mänskliga djur illustrerar att mirtazapin är effektivt för att minska beteenden motiverade av missbrukade droger (t.ex. opiater och psykostimulanter) även de som är associerade med återfall under perioder av abstinens (för granskning, se Graves et al., 2012). Data som samlats in från vår ICSS-medierade insatsbaserade beslutsfattande uppgift indikerar att mirtazapin effektivt reducerade preferensen för en hög ansträngning / LR, övergång till låg insats / SR, vilket tyder på att den mängd ansträngning som krävs för LR inte längre var värd det "eller att belöningsvärdet för LR minskade (Persons et al., 2013). Dessa resultat tyder på att det kan vara av intresse att studera effekterna av mirtazapin på suboptimal beslutsfattande hos problemspelare i kliniken.
Slutsats
Sammanfattningsvis har vi använt ICSS som en positiv förstärkare i flera nya uppgifter som syftar till att mäta separata men ändå överlappande aspekter av kostnad / nytta beslutsfattande uppvisade i problemgambling. Dessa åtgärder kan användas för att ytterligare undersöka bidrag från olika neuroanatomiska substrat och neurotransmittorsystem i problemgambling. ICSS-medierade uppgifter ger ett livskraftigt alternativ till matförstärkning i dessa komplexa operantparadigmer. Vi tror att giltigheten av dessa uppgifter tyder på att de kan hjälpa till med att screena droger för att de kan inducera impulskontrollsjukdomar, som problemgambling och att hjälpa till att identifiera droger som minskar dessa störningar.
Intresseanmälan
Dr. Napier har fått forskningsstöd från National Institutes of Health, Michael J. Fox Foundation och National Center for Responsible Gaming. Dr Napier har fått ersättning för följande: Rådgivning för ett icke-vinstdrivande hälsoutbildningscenter och för advokatbyråer i frågor som relaterar till missbruk och impulskontrollsjukdomar; talar om missbruk vid möteshallens möten, offentliga högskolor, samhällsbaserade icke-vinstdrivande och professionella möten för drogdomstolar; tillhandahålla bidragsrevisioner för National Institutes of Health och andra organ och akademiska föreläsningar och stora rundor. Dr. Napier är medlem i Illinois Alliance on Problem Gambling, och hon tillhandahåller expertråd om medicinutveckling till Cures Within Research Foundation. Dr. Holtz, Dr. Persons och Ms Tedford förklara att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
Erkännanden
Detta arbete stöddes av National Center for Responsible Gaming, Michael J. Fox Foundation, Daniel F. och Ada L. Rice Foundation, och USPHSGs NS074014 till T. Celeste Napier och DA033121 till Stephanie E. Tedford och T. Celeste Napier .
Referensprojekt
Alessi, SM och Petry, NM (2003). Patologisk spelighetsgrad är associerad med impulsivitet vid fördröjning av diskonteringsförfarandet. Behav. processer 64, 345–354. doi: 10.1016/S0376-6357(03)00150-5
Bickel, WK, Jarmolowicz, DP, Mueller, ET, Koffarnus, MN och Gatchalian, KM (2012). Överdriven diskontering av fördröjda förstärkare som en trans-sjukdomsprocess som bidrar till missbruk och andra sjukdomsrelaterade sårbarheter: framväxande bevis. Pharmacol Ther. 134, 287-297. doi: 10.1016 / j.pharmthera.2012.02.004
Bizo, LA, Bogdanov, SV och Killeen, PR (1998). Satiation orsakar in-session minskningar i instrumentalt svar. J. Exp. Psychol. Anim. Behav. Bearbeta. 24, 439-452. doi: 10.1037 / 0097-7403.24.4.439
Buckholtz, JW, Treadway, MT, Cowan, RL, Woodward, ND, Li, R., Ansari, MS, et al. (2010). Dopaminerga nätverksskillnader i humant impulsivitet. Vetenskap 329, 532. doi: 10.1126 / science.1185778
Buelow, MT och Suhr, JA (2009). Konstruera giltigheten av iowa-speluppgiften. Neuropsychol. Varv. 19, 102-114. doi: 10.1007 / s11065-009-9083-4
Corbett, D. och Wise, RA (1980). Intrakraniell självstimulering i förhållande till de uppåtgående dopaminerga systemen i mitten: En rörlig elektrodkartläggningsstudie. Brain Res. 185, 1–15. doi: 10.1016/0006-8993(80)90666-6
Crean, JP, de, WH och Richards, JB (2000). Belöningsrabattning som ett mått på impulsivt beteende hos en psykiatrisk öppenvårdspopulation. Exp. Clin. Psychopharmacol. 8, 155-162. doi: 10.1037 / 1064-1297.8.2.155
Dai, Z., Harrow, SE, Song, X., Rucklidge, J., och Grace, R. (2013). Spel, fördröjning och sannolikhet diskontering hos vuxna med och utan ADHD. J. Atten. Disord. doi: 10.1177 / 1087054713496461. [Epub före utskrift].
Dixon, MR, Marley, J. och Jacobs, EA (2003). Fördröjning av diskontering av patologiska spelare. J. Appl. Behav. Anal. 36, 449-458. doi: 10.1901 / jaba.2003.36-449
Douglas, VI och Parry, PA (1994). Effekter av belöning och nonreward på frustration och uppmärksamhet i uppmärksamhet underskott störning. J. Abnorm. Barnpsykol. 22, 281-302. doi: 10.1007 / BF02168075
Eubig, PA, Noe, TE, Floresco, SB, Sable, JJ och Schantz, SL (2014). Sexskillnader som svar på amfetamin hos vuxna long evans råttor som utför en fördröjningsdiskonteringsuppgift. Pharmacol. Biochem. Behav. 118, 1-9. doi: 10.1016 / j.pbb.2013.12.021
Feja, M. och Koch, M. (2014). Ventral medial prefrontal cortex inaktivering försämrar impulskontrollen men påverkar inte fördröjningsrabattning hos råttor. Behav. Brain Res. 264, 230-239. doi: 10.1016 / j.bbr.2014.02.013
Graves, SM, Rafeyan, R., Watts, J. och Napier, TC (2012). Mirtazapin, och mirtazapinliknande föreningar som möjligt farmakoterapi för missbrukstörningar: bevis från bänken och sängen. Pharmacol. Ther. 136, 343-353. doi: 10.1016 / j.pharmthera.2012.08.013
Holt, DD, Green, L. och Myerson, J. (2003). Är diskontering impulsiv? Bevis från tidsmässig och sannolikhet diskontering i spelande och icke-spelande högskolestudenter. Behav. processer 64, 355–367. doi: 10.1016/S0376-6357(03)00141-4
Hosking, JG, Cocker, PJ och Winstanley, CA (2014). Dissocierbara bidrag från främre cingulära cortex och basolateral amygdala på en gnagare kostnad / nytta beslutsfattande uppgift av kognitiv ansträngning. Neuropsychopharmacology 39, 1558-1567. doi: 10.1038 / npp.2014.27
Housden, CR, O'Sullivan, SS, Joyce, EM, Lees, AJ och Roiser, JP (2010). Intakt belöningsinlärning men förhöjd fördröjningsrabatt hos patienter med Parkinsons sjukdom med impulsivt-kompulsivt spektrumbeteende. Neuropsychopharmacology 35, 2155-2164. doi: 10.1038 / npp.2010.84
Jimura, K., Myerson, J., Hilgard, J., Braver, TS och Green, L. (2009). Är människor verkligen mer tålmodiga än andra djur? bevis från mänsklig diskontering av verkliga likvida belöningar. Psychon. Tjur. Varv. 16, 1071-1075. doi: 10.3758 / PBR.16.6.1071
Johnson, PS, Madden, GJ, Brewer, AT, Pinkston, JW och Fowler, SC (2011). Effekter av akut pramipexol på preferens för spelliknande scheman för förstärkning hos råttor. Psykofarmakologi (Berl) 231, 11–18. doi: 10.1007/s00213-010-2006-5
Keesey, RE (1962). Relationen mellan pulsfrekvens, intensitet och varaktighet och graden av respons för intrakraniell stimulering. J. Comp. Physiol. Psychol. 55, 671-678. doi: 10.1037 / h0049222
Koffarnus, MN, Newman, AH, Grundt, P., Rice, KC och Woods, JH (2011). Effekter av selektiva dopaminergiska föreningar vid en fördröjningsdiskonteringsuppgift. Behav. Pharmacol. 22, 300-311. doi: 10.1097 / FBP.0b013e3283473bcb
Kraplin, A., Dshemuchadse, M., Behrendt, S., Scherbaum, S., Goschke, T. och Buhringer, G. (2014). Dysfunktionell beslutsfattande vid patologisk spel: mönsterspecifika och impulsivitetens roll. Psykiatrisk Res. 215, 675-682. doi: 10.1016 / j.psychres.2013.12.041
Ledgerwood, DM, Alessi, SM, Phoenix, N. och Petry, NM (2009). Behavioral bedömning av impulsivitet hos patologiska spelare med och utan substansanvändningsstörningar jämfört med friska kontroller. Drogalkohol Beroende. 105, 89-96. doi: 10.1016 / j.drugalcdep.2009.06.011
Leroi, I., Barraclough, M., McKie, S., Hinvest, N., Evans, J., Elliott, R. och McDonald, K. (2013). Dopaminerg påverkan på verkställande funktion och impulsivt beteende vid impulskontrollstörningar vid Parkinsons sjukdom. J. Neuropsychol. 7, 306-325. doi: 10.1111 / jnp.12026
Madden, GJ, Johnson, PS, Brewer, AT, Pinkston, JW och Fowler, SC (2010). Effekter av pramipexol på impulsivt val hos manliga wistar-råttor. Exp. Clin. Psychopharmacol. 18, 267-276. doi: 10.1037 / a0019244
Madden, GJ, Petry, NM och Johnson, PS (2009). Patologiska spelare rabatt probabilistiska belöningar mindre brant än matchade kontroller. Exp. Clin. Psychopharmacol. 17, 283-290. doi: 10.1037 / a0016806
McCown, TJ, Napier, TC och Breese, GR (1986). Effekter av kronisk elektrodimplantation på dopaminerga neuroner in vivo-. Pharmacol. Biochem. Behav 25, 63–69. doi: 10.1016/0091-3057(86)90231-5
Mejia-Toiber, J., Boutros, N., Markou, A. och Semenova, S. (2014). Impulsivt val och ångestliknande beteende hos vuxna råttor utsatta för kronisk intermittent etanol under ungdom och vuxen ålder. Behav. Brain Res. 266, 19-28. doi: 10.1016 / j.bbr.2014.02.019
Michalczuk, R., Bowden-Jones, H., Verdejo-Garcia, A. och Clark, L. (2011). Impulsivitet och kognitiv snedvridning hos patologiska spelare som besöker Storbritanniens nationella problemspelsklinik: en preliminär rapport. Psychol. Med. 41, 2625-2635. doi: 10.1017 / S003329171100095X
Miedl, SF, Peters, J. och Buchel, C. (2012). Ändrade neurala belöningsrepresentationer hos patologiska spelare som avslöjas av fördröjning och sannolikhetsdiskontering. Båge. Gen. Psykiatri 69, 177-186. doi: 10.1001 / archgenpsychiatry.2011.1552
Milenkova, M., Mohammadi, B., Kollewe, K., Schrader, C., Fellbrich, A., Wittfoth, M., et al. (2011). Intertemporal val vid Parkinsons sjukdom. Mov. Disord. 26, 2004-2010. doi: 10.1002 / mds.23756
Minamimoto, T., La Camera, G. och Richmond, BJ (2009). Mätning och modellering av interaktionen mellan belöningsstorlek, fördröjning till belöning och mättnadsnivå på motivation hos apor. J. Neurophysiol. 101, 437-447. doi: 10.1152 / jn.90959.2008
Olds, J. och Milner, P. (1954). Positiv förstärkning producerad genom elektrisk stimulering av septal-området och andra områden av råtthjärnan. J. Comp. Physiol. Psychol. 47, 419-427. doi: 10.1037 / h0058775
Papini, MR och Dudley, RT (1997). Konsekvenserna av överraskande belöningsutelämnanden. Rev. Gen. Psychol. 1, 175-197. doi: 10.1037 / 1089-2680.1.2.175
Personer, AL, Tedford, SE och Napier, TC (2013). "Prekliniska utvärderingar av serotonergiska föreningar med blandfunktion för behandling av spelbesvär. program nr. 635.12, "in Neurovetenskapens mötesplanerare (San Diego, CA: Samhälle för neurovetenskap).
Petry, NM (2011). Diskontering av probabilistiska belöningar är förknippad med speluppehåll hos behandlingssökande patologiska spelare. J. Abnorm. Psychol. 121, 151-159. doi: 10.1037 / a0024782
Pirch, JH, Napier, TC och Corbus, MJ (1981). Hjärnstimulering som en cue för händelsesrelaterade potentialer i råttcortex: amfetamin effekter. Int. J. Neurosci. 15, 217-222.
Pizzagalli, DA, Evins, AE, Schetter, EC, Frank, MJ, Pajtas, PE, Santesso, DL, et al. (2008). Endos av en dopaminagonist försvårar förstärkning av lärande hos människor: beteendemässig bevisning från ett laboratoriebaserat mått på belöningsrespons. Psykofarmakologi (Berl) 196, 221–232. doi: 10.1007/s00213-007-0957-y
Rachlin, H., Raineri, A. och Cross, D. (1991). Subjektiv sannolikhet och fördröjning. J. Exp. Anal. Behav. 55, 233-244. doi: 10.1901 / jeab.1991.55-233
Rasmussen, EB, advokat, SR och Reilly, W. (2010). Procent kroppsfett är relaterad till fördröjning och sannolikhet diskontering för mat hos människor. Behav. processer 83, 23-30. doi: 10.1016 / j.beproc.2009.09.001
Reynolds, B., Richards, JB, Horn, K. och Karraker, K. (2004). Försenad diskontering och sannolikhet diskontering som relaterad till cigarettrökstatus hos vuxna. Behav. processer 65, 35–42. doi: 10.1016/S0376-6357(03)00109-8
Riba, J., Kramer, UM, Heldmann, M., Richter, S. och Munte, TF (2008). Dopaminagonist ökar riskupptagningen men blockerar belöningsrelaterad hjärnaktivitet. PLoS. ETT 3: e2479. doi: 10.1371 / journal.pone.0002479
Rokosik, SL och Napier, TC (2011). Intrakranial självstimulering som en positiv förstärkare för att studera impulsivitet i ett sannolikhetsdiskonteringsparadigm. J. Neurosci. metoder 198, 260-269. doi: 10.1016 / j.jneumeth.2011.04.025
Rokosik, SL och Napier, TC (2012). Pramipexolinducerad ökad probabilistisk diskontering: jämförelse mellan en gnagarmodell av Parkinsons sjukdom och kontroller. Neuropsychopharmacology 37, 1397-1408. doi: 10.1038 / npp.2011.325
Roll, JM, McSweeney, FK, Johnson, KS och Weatherly, JN (1995). Satiety bidrar lite till in-session minskningar i att svara. Lära sig. Motiv. 26, 323–341. doi: 10.1016/0023-9690(95)90018-7
Schuck-Paim, C., Pompilio, L. och Kacelnik, A. (2004). Statligt beroende beslut orsakar uppenbara överträdelser av rationalitet i djurval. PLoS. Biol. 2: e402. doi: 10.1371 / journal.pbio.0020402
Spengos, K., Grips, E., Karachalios, G., Tsivgoulis, G. och Papadimitriou, G. (2006). Reversibelt patologiskt spel under behandling med pramipexol. Nervenarzt 77, 958–960. doi: 10.1007/s00115-006-2128-5
St Onge, JR och Floresco, SB (2009). Dopaminerg modulering av riskbaserat beslutsfattande. Neuropsychopharmacology 34, 681-697. doi: 10.1038 / npp.2008.121
Stopper, CM och Floresco, SB (2011). Bidrag från kärnan accumbens och dess delregioner till olika aspekter av riskbaserad beslutsfattande. Cogn. Påverka. Behav. Neurosci. 11, 97-112. doi: 10.3758 / s13415-010-0015-9
Szamosi, A., Nagy, H. och Keri, S. (2013). Fördröja diskontering av belöning och caudat kärnvolym hos individer med alfa-synuklein-gen duplicering före och efter utvecklingen av Parkinsons sjukdom. Neurodegener. Dis. 11, 72-78. doi: 10.1159 / 000341997
Tedford, SE, Persons, AL, Chen, L., Rokosik, SL och Napier, TC (2012). "Intrakranial självstimulering som positiv förstärkare för att studera impulsivitet i ett fördröjt diskonteringsparadigm hos råttor. programnummer 926.03, "in Neurovetenskapens mötesplanerare (New Orleans, LA: Samhälle för neurovetenskap).
Tehovnik, EJ och Sommer, MA (1997). Effektiv spridning och tidsåtgång av neuralt inaktivering orsakad av lidokaininjektion i apa cerebral cortex. J. Neurosci. metoder 74, 17–26. doi: 10.1016/S0165-0270(97)02229-2
Treadway, MT, Buckholtz, JW, Schwartzman, AN, Lambert, WE och Zald, DH (2009). Värt "EEfRT"? Ansträngningsutgifterna för belöningsuppgift som ett objektivt mått på motivation och anhedoni. PLoS. ETT 4: e6598. doi: 10.1371 / journal.pone.0006598
Trowill, JA, Panksepp, J. och Gandelman, R. (1969). En incitament modell av givande hjärnstimulering. Psychol. Varv. 76, 264-281. doi: 10.1037 / h0027295
Voon, V., Sohr, M., Lang, AE, Potenza, MN, Siderowf, AD, Whetteckey, J., et al. (2011). Impulskontrollstörningar vid Parkinsons sjukdom: en studie med flera centre fallstudier. Ann. Neurol. 69, 986-996. doi: 10.1002 / ana.22356
Wardle, MC, Treadway, MT, Mayo, LM, Zald, DH och de, WH (2011). Förstärkt ansträngning: Effekter av d-amfetamin på mänskligt ansträngningsbaserat beslutsfattande. J. Neurosci. 31, 16597-16602. doi: 10.1523 / JNEUROSCI.4387-11.2011
Weintraub, D., Koester, J., Potenza, MN, Siderowf, AD, Stacy, M., Voon, V., et al. (2010). Impulskontrollstörningar i Parkinsons sjukdom: En tvärsnittsstudie av 3090-patienter. Båge. Neurol. 67, 589-595. doi: 10.1001 / archneurol.2010.65
Klokt, RA (1996). Beroendeframkallande läkemedel och hjärnstimulansbelöning. Annu. Rev. Neurosci. 19, 319-340. doi: 10.1146 / annurev.neuro.19.1.319
Klokt, RA (2005). Forebrain-substrat av belöning och motivation. J. Comp. Neurol. 493, 115-121. doi: 10.1002 / cne.20689
Wise, RA, Bauco, P., Carlezon, WA Jr. och Trojniar, W. (1992). Självstimulerande och läkemedelsbelöningsmekanismer. Ann. NY Acad. sci. 654, 192-198. doi: 10.1111 / j.1749-6632.1992.tb25967.x
Wise, RA och Bozarth, MA (1981). Hjärnunderlag för förstärkning och självbehandling av droger. Prog. Neuropsychopharmacol. 5, 467-474. doi: 10.1016 / 0364-7722 (81) 90028-X
Wise, RA, och Rompre, PP (1989). Hjärndopamin och belöning. Annu. Rev. Psychol. 40, 191-225. doi: 10.1146 / annurev.psych.40.1.191
Zeeb, FD, Robbins, TW och Winstanley, CA (2009). Serotonerg och dopaminerg modulering av spelbeteende som bedömts med hjälp av en ny råttspeluppgift. Neuropsychopharmacology 34, 2329-2343. doi: 10.1038 / npp.2009.62
Nyckelord: kostnad / nytta beslutsfattande, diskontering, ansträngningsbaserad beslutsfattande, spelande, intrakranial självstimulering
Citat: Tedford SE, Holtz NA, Personer AL och Napier TC (2014) Ett nytt tillvägagångssätt för att bedöma spelliknande beteenden i laboratorierådor: Användning av intrakranial självstimulering som positiv förstärkare. Främre. Behav. Neurosci. 8: 215. doi: 10.3389 / fnbeh.2014.00215
Mottagen: 06 mars 2014; Godkänd: 27 maj 2014;
Publicerad online: 11 June 2014.
Redigerad av:
Patrick Anselme, Universitetet i Liège, Belgien
Recenserad av:
Christelle Baunez, National National de la Recherche Scientifique, Frankrike
Yueqiang Xue, University of Tennessee Health Science Center, USA
