 Stephanie E. Tedford1,2*, Nathan A. Holtz1,2, Amanda L. Personnes1,2 † ainsi que T. Celeste Napier1,2,3
Stephanie E. Tedford1,2*, Nathan A. Holtz1,2, Amanda L. Personnes1,2 † ainsi que T. Celeste Napier1,2,3- 1Département de pharmacologie, Centre de toxicomanie et de comportement compulsif, Centre médical de l'Université Rush, Chicago, IL, États-Unis.
- 2Département de pharmacologie, Centre médical de l'Université Rush, Chicago, IL, USA
- 3Département de psychiatrie, Centre médical de l'Université Rush, Chicago, IL, États-Unis
Le jeu pathologique est une manifestation des troubles du contrôle des impulsions. Les fondements biologiques de ces troubles restent insaisissables et le traitement est loin d'être idéal. Les modèles animaux de troubles du contrôle des impulsions constituent un outil de recherche essentiel pour comprendre cette pathologie et développer les médicaments. Il est difficile de modéliser de tels comportements complexes, mais par sa déconstruction, les scientifiques ont résumé chez les animaux les aspects critiques du jeu. L'un des aspects du jeu est la prise de décision coûts-avantages, dans laquelle on évalue les coûts et les avantages attendus d'un plan d'action. La prise de décision basée sur le risque et la récompense, basée sur les délais et sur les efforts représente tous des choix coûts / bénéfices. Ces caractéristiques sont étudiées chez l'homme et ont été traduites en protocoles animaux pour mesurer les processus de prise de décision. Traditionnellement, le renforcement positif utilisé dans les études sur les animaux est la nourriture. Nous décrivons ici comment l'auto-stimulation intracrânienne peut être utilisée pour des tâches décisionnelles coût / bénéfice et récapitulons nos récentes études montrant comment les thérapies pharmacologiques modifient ces comportements chez le rat de laboratoire. Nous proposons que ces modèles pourraient être utiles dans le dépistage de nouveaux composés pour leur capacité à promouvoir et à prévenir certains aspects du comportement de jeu.
Introduction
Le jeu problématique ou inadapté, y compris la condition extrême appelée jeu pathologique, est caractérisé par des comportements qui persistent souvent sur de longues périodes. Le jeu problématique peut avoir un impact négatif important sur le bien-être personnel, professionnel et financier. Au cours des deux dernières décennies, les possibilités de jeu ont augmenté grâce aux modifications de la législation et à l’introduction de nouveaux lieux de jeu (par exemple, les jeux sur Internet). En conséquence, la prévalence du jeu problématique a augmenté. Il n'existe pas de traitement approuvé par la FDA pour ce trouble. Il est donc essentiel de mieux comprendre ces comportements afin de développer des thérapies efficaces.
Le jeu problématique est un phénomène complexe, qui inclut des niveaux accrus de prise de décision impulsive (Alessi et Petry, 2003; Dixon et al., 2003; Holt et al., 2003; Kraplin et al., 2014) résultant d’évaluations défavorables des coûts / avantages. Les évaluations cliniques de la prise de décision, qui utilisent souvent des enquêtes et des outils informatiques interactifs, ont joué un rôle déterminant dans la détermination de profils de prise de décision sous-optimaux dans diverses pathologies, y compris les joueurs pathologiques (Ledgerwood et al., 2009; Madden et al., 2009; Michalczuk et al., 2011; Petry, 2011; Miedl et al., 2012). Les évaluations cliniques reposent souvent sur trois aspects différents, bien que superposés, de la prise de décision coûts / avantages, à savoir: (i) le niveau de risque lié à l’obtention d’une récompense (prise de décision risque / récompense), (ii) une retard subi avant la remise de la récompense (prise de décision basée sur le retard), et (iii) la quantité d'effort requise pour obtenir une récompense (prise de décision basée sur l'effort). Plusieurs tâches ont été développées pour mesurer ces caractéristiques critiques de la prise de décision sous-optimale afin de mieux comprendre les processus qui comprennent le jeu problématique. Dans ces tâches, le sujet choisit entre une récompense petite ou grande, chacune associée à des éventualités de réponse spécifiques. Dans la prise de décision risque / récompense (c.-à-d. Réduction de probabilité), les sujets choisissent entre une petite récompense générée de manière cohérente avec des probabilités élevées (par exemple, 100% de probabilité de recevoir $ 10) et une récompense importante livrée avec des probabilités variables (par exemple, 10 – 80 % de probabilité de recevoir $ 100). Dans les études cliniques et précliniques, l’absence de récompense attendue est un événement aversif qui entraîne des réponses physiologiques correspondantes (Douglas et Parry, 1994; Papini et Dudley, 1997). La préférence accordée à l'option plus large et «risquée» par rapport à l'option petite et certaine est considérée comme reflétant une prise de décision sous-optimale en termes de risque / rendement et a été rapportée pour plusieurs pathologies humaines présentant une impulsivité accrue (Reynolds et al., 2004; Rasmussen et al., 2010; Dai et al., 2013). Dans la prise de décision basée sur le retard (c.-à-d. Réduction du retard, mesure du choix impulsif), la petite récompense est livrée peu de temps après la sélection de l’option, tandis que la récompense la plus importante est délivrée à la suite d’un retard variable (par exemple, $ 10 maintenant ou moins). $ 100 en semaines 2). Les individus qui font preuve d'une forte impulsivité manifestent leur préférence pour les récompenses immédiatement disponibles (même si elles sont plus petites), par rapport aux récompenses différées (même si elles sont plus grandes), bien que cette dernière option puisse être plus avantageuse pour l'individu (Crean et al., 2000; Reynolds et al., 2004; Bickel et al., 2012). Dans la prise de décision basée sur l'effort, le sujet choisit entre une petite récompense donnée après de petits efforts, ou une récompense importante après un effort plus important. Dans cette tâche, la préférence individuelle pour l'option d'effort élevé / grande récompense et le «point» où l'individu bascule vers l'option d'effort faible / petite récompense sont déterminés. Des études sur la prise de décision basée sur l'effort chez les joueurs humains doivent encore être menées, mais seraient d'un intérêt significatif pour évaluer la fonction cognitive dans cette population.
Les protocoles de prise de décision utilisés dans les évaluations cliniques peuvent être modifiés pour étudier la prise de décision chez le rat de laboratoire. Ces modèles sont essentiels pour explorer les aspects comportementaux et neuropharmacologiques du jeu pathologique. Chez le rat, la prise de décision peut être évaluée en plaçant l'animal dans une chambre de conditionnement et en lui permettant de choisir entre deux leviers (ou deux trémies nez-poke) mis à disposition en même temps. La modalité de récompense établie pour le renforçateur positif dans ces tâches de rongeur est la nourriture (Stopper et Floresco, 2011; Eubig et al., 2014). Nous discutons ici une nouvelle méthode utilisée dans notre laboratoire qui utilise la stimulation électrique directe des voies de récompense du cerveau (auto-stimulation intracrânienne; ICSS) pour évaluer la prise de décision coût / bénéfice chez le rat et l’apport des neurotransmetteurs monoaminergiques dans la prise de décision (Rokosik et Napier, 2011, 2012; Tedford et al., 2012; Personnes et al., 2013).
Auto-stimulation intracrânienne
Un renforçateur opérant est un stimulus qui, lorsqu'il est subordonné à une action, augmente la probabilité que cette action se reproduise. L'autostimulation intracrânienne (ICSS) est un comportement opérant dans lequel les animaux auto-administrent une stimulation électrique à des régions cérébrales connues pour être impliquées dans le renforcement positif. ICSS a été étudié pour la première fois dans les 1950 lorsque James Olds et Peter Milner (Olds et Milner, 1954) ont déterminé que les rats retournaient à plusieurs reprises à un endroit dans une boîte où ils recevaient une stimulation électrique vers des régions du cerveau liées à la récompense. Ils ont permis aux rats de travailler pour cette stimulation électrique du cerveau (EBS) en répondant à un manipulandum opérant (par exemple, en appuyant sur un levier, en faisant tourner une roue) (Olds et Milner, 1954). La découverte de cette technique a joué un rôle déterminant dans la cartographie des voies de récompense à travers le cerveau. Bien que de nombreuses régions du cerveau puissent être utilisées pour soutenir l'ICSS (Olds et Milner, 1954; Wise et Bozarth, 1981; Sage, 1996), il est bien établi que la stimulation du faisceau médial du cerveau antérieur (MFB) favorise des sorties comportementales profondes et fiables (Corbett et Wise, 1980; Pirch et al., 1981; McCown et al., 1986; Tehovnik et Sommer, 1997). Les paramètres de courant de stimulation peuvent être manipulés pour affecter la valeur de renforcement de l'EBS et donc modifier le comportement ICSS. Ces paramètres incluent l'intensité (c.-à-d. Les ampères) du courant électrique et la fréquence du courant (c.-à-d. Le hertz). Les élévations des deux paramètres entraînent généralement une excitation accrue des neurones pertinents pour la récompense, soit en augmentant le nombre de neurones engagés par la stimulation (ampères) (Keesey, 1962; Wise et al., 1992) ou en augmentant la fréquence à laquelle une population déterminée de neurones déclenche (hertz) (Sage et Rompre, 1989; Sage, 2005). Les manipulations d'intensité de courant modifient le nombre de neurones activés, c'est-à-dire que des intensités de courant plus importantes affectent une plus grande population de neurones que des courants plus petits. Ainsi, lorsque ce paramètre est maintenu constant, la population de neurones excités par EBS est relativement similaire quelle que soit la fréquence actuelle. Le paramètre de stimulation de choix pour ces protocoles est la fréquence actuelle, car cette sélection nous permet de manipuler la cadence de déclenchement du même groupe de neurones avec des effets minimaux sur le temps ou l’espace de l’intégration de stimulation. En manipulant ces paramètres EBS, nous avons développé des modèles sophistiqués de prise de décision coût / bénéfice qui utilisent l’ICSS (Rokosik et Napier, 2011, 2012; Tedford et al., 2012; Personnes et al., 2013). Cette application représente une rupture radicale avec le stimulus de renforcement (c’est-à-dire, l’alimentation) utilisé traditionnellement pour évaluer la prise de décision chez les rongeurs. ICSS peut offrir plusieurs avantages expérimentaux par rapport aux méthodes de renforcement traditionnelles. Pour faciliter la réponse opérée des aliments, l’apport quotidien est souvent limité (Feja et Koch, 2014; Hosking et al., 2014; Mejia-Toiber et al., 2014). Cette pratique peut confondre les mesures de résultats, car il existe un chevauchement important des systèmes neurobiologiques qui sont modifiés lors de restrictions alimentaires chroniques et de ceux qui interviennent dans la prise de décision impulsive (Schuck-Paim et al., 2004; Minamimoto et al., 2009). De plus, les animaux renforcés avec de la nourriture sont de plus en plus rassasiés tout au long de la session, ce qui diminue la valeur du renforcement des aliments (Bizo et al., 1998), bien que cet effet puisse dépendre de la taille du renfort (Roll et al., 1995). Contrairement au renforcement des aliments, la valeur de renforcement de l'EBS reste stable tout au long d'une session, permettant des évaluations comportementales plus complètes et cohérentes (Trowill et al., 1969). Cette fonctionnalité permet aux sessions de tests de se répéter plusieurs fois par jour, ce qui peut être bénéfique lors de l’étude des effets des thérapies pharmacologiques, en particulier des traitements médicamenteux chroniques. Nos études publiées sur la réduction de probabilité (décrites ci-dessous) ont été menées plusieurs fois par jour au cours de traitements chroniques par un agoniste de la dopamine (pramipexole). Nous proposons que cet avantage procédural est plus applicable à la condition humaine et fournit ainsi des résultats de traduction améliorés. À ce jour, des études similaires évaluant les effets des agonistes de la dopamine sur la prise de décision impulsive utilisant un aliment récompense n’ont évalué que les traitements médicamenteux en phase aiguë (St Onge et Floresco, 2009; Zeeb et al., 2009; Madden et al., 2010; Johnson et al., 2011; Koffarnus et al., 2011) et il sera très intéressant de comparer les résultats comportementaux à la suite d'un traitement médicamenteux aigu et chronique entre ces différents renforçants. Si l'ICSS présente plusieurs avantages par rapport au renforcement des aliments, l'ICSS présente également plusieurs inconvénients. Par exemple, l'ICSS nécessite une chirurgie et une récupération cérébrales invasives, et des stades de la tête mal ajustés peuvent entraîner la perte de sujets tout au long du paradigme comportemental. Malgré ces inconvénients, nous pensons que l’ICSS est une alternative viable au renforcement des aliments et présente des avantages considérables pour le renforcement des aliments dans ces tâches comportementales.
Les tâches de prise de décisions en matière de coûts et d’avantages nécessitent de choisir entre des options associées à des ampleurs de récompense variables. En conséquence, les renforçateurs utilisés dans ces tâches doivent démontrer la capacité de produire de tels changements dans l’ampleur de la récompense, puis les rats doivent être en mesure de faire la distinction entre l’option de petit renforçateur (SR) et celle de grand renforçateur (LR). Dans les procédures qui utilisent le renforcement alimentaire, ceci est obtenu en modifiant le nombre de granulés alimentaires obtenus après une réponse. Dans ICSS, l'EBS peut être modifié en modifiant l'intensité du courant de stimulation ou la fréquence du courant. Figure 1 illustre la réponse du levier-presse obtenue lorsque l'intensité du courant varie (c'est-à-dire que la fréquence du courant est maintenue constante; 1A) ou lorsque la fréquence du courant varie (l’intensité du courant est maintenue constante; Figure 1B). Lorsque l'un ou l'autre paramètre est modifié, les rats présentent un effet de levier modéré en appuyant sur de petites valeurs d'EBS et des taux de pression de levier plus élevés pour des valeurs de EBS élevées, ce qui suggère que la valeur de renforcement de la stimulation la plus grande est supérieure (que l'intensité ou la fréquence du courant soit manipulée ou non). EBS peut donc être adapté pour les renforçateurs petits et grands nécessaires aux protocoles de prise de décision coûts / avantages. Ces valeurs de renforcement peuvent être déterminées chez des rats individuels en générant des courbes de réponse au taux de pression de levier stables pour chaque animal (Rokosik et Napier, 2011, 2012). Alternativement, une courbe de population peut être générée à partir d’un groupe de rats à partir duquel une valeur normalisée SR et LR peut être déterminée (Tedford et al., 2012; Personnes et al., 2013). Cette dernière approche fournit un moyen plus efficace et rapide de dériver les valeurs SR et LR. Dans une deuxième série d'études, nous avons utilisé des manipulations d'intensité ou de fréquence du courant pour établir les valeurs de RS / LR dans une tâche d'actualisation de probabilité (prise de décision en fonction du risque / rendement). Les modifications des valeurs de renforcement d'intensité de courant (c.-à-d. Que la fréquence du courant est maintenue constante) et les valeurs de fréquence actuelles (c.-à-d. L'intensité du courant est maintenue constante) produisent toutes deux un comportement d'actualisation significatif chez le rat (Figures 1C, D). En partie sur la base de la pente de la courbe d’actualisation, il a été déterminé que la fréquence du courant était le paramètre approprié pour manipuler les valeurs de ferraillage. Une fois qu'il est établi que les rats peuvent faire la distinction entre les fréquences de courant normalisées utilisées pour les RS et les LR, ils peuvent être testés selon l'un des paradigmes de prise de décision à médiation ICSS suivants: (i) prise de décision en matière de risque / rendement (Rokosik et Napier, 2011, 2012), ii) prise de décision en fonction des délais (Tedford et al., 2012), ou (iii) une prise de décision basée sur l'effort (Personnes et al., 2013).
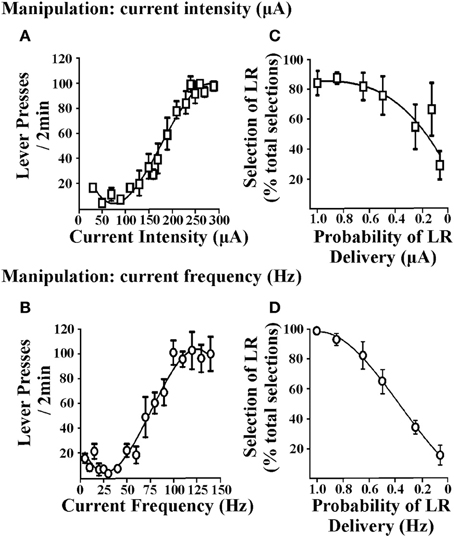
Figure 1. Effets des paramètres de stimulation cérébrale sur la réponse de la presse à levier et la réduction de probabilité. Les deux paramètres EBS testés étaient l'intensité du courant et la fréquence du courant. Levier de rats pressé pour EBS (dans un rapport de renforcement de ratio 1 fixe) dans lequel chaque minute 2, un paramètre d'EBS était manipulé et l'autre paramètre maintenu constant. (A) Manipulation de l'intensité du courant. Des intensités de courant allant de 10 à 350 μA ont été présentées dans un ordre randomisé (n = 6); la fréquence actuelle a été maintenue à 100 Hz. (B) Manipulation de la fréquence actuelle. Les fréquences courantes allant de 5 à 140 Hz ont été présentées dans un ordre aléatoire (n = 3); l'intensité du courant a été maintenue constante à un niveau individualisé et déterminé lors de sessions de formation antérieures. La manipulation de l'intensité du courant ou de la fréquence du courant a produit des modèles similaires de réponse par pression sur le levier. Les données sont exprimées en moyenne ± sem pour les trois dernières sessions consécutives. Les rats ont ensuite été entraînés à la tâche de réduction de probabilité et les valeurs des renforcements petits et grands ont été déterminées individuellement pour chaque animal en calculant les intensités de courant de stimulation effectives et les fréquences des courants obtenues à partir de la courbe de réponse EBS versus la courbe de réponse à pression au levier ayant provoqué 60 et 90% des taux de réponse maximaux presse-levier, respectivement. Varier la magnitude de l'intensité du courant (C) ou fréquence actuelle (D) a entraîné l’actualisation du grand renforçateur (LR) à mesure que la probabilité d’accouchement diminuait (c’est-à-dire une diminution du pourcentage de sélection du levier associé au LR par rapport au total des sélections). Les données sont exprimées en moyenne ± sem pour le premier jour de l'actualisation en utilisant l'intensité actuelle et en 2 pour l'actualisation en utilisant la fréquence actuelle. Figure modifiée de Rokosik et Napier (2011) et réimprimé avec la permission de l'éditeur.
Validation de l'utilisation de l'ICSS pour évaluer les mesures d'impulsivité et la prise de décision
Le développement de nouveaux modèles animaux nécessite un examen minutieux de la validité. Ainsi, lors de la conception de ces tâches décisionnelles à médiation ICSS, nous nous sommes efforcés de vérifier la validité apparente et la validité de construction, et de déterminer la probabilité de validité prédictive.
La validité apparente fait référence à la mesure dans laquelle un test semble subjectivement mesurer le phénomène auquel il est destiné. La conception de chaque tâche décisionnelle à médiation ICSS était basée sur les protocoles actuels utilisés chez l'homme pour la réduction de délai et de probabilité (Rasmussen et al., 2010; Leroi et al., 2013) et d’autres tâches décisionnelles axées sur l’effort (Treadway et al., 2009; Buckholtz et al., 2010; Wardle et al., 2011). Chez les humains, les mesures de la prise de décision en termes de coûts / avantages découlent du fait de demander aux individus de choisir entre plusieurs options disponibles avec des imprévus spécifiques placés sur chaque sélection (c.-à-d. Un risque, un délai ou un effort). Nous imitons ce scénario en présentant aux rats deux leviers étendus simultanément, dans lesquels une sélection de l'un ou l'autre des leviers est associée à des récompenses petites ou plus grandes, également livrées dans des paramètres particuliers de contingence. Ainsi, chacune de nos tâches décisionnelles à médiation ICSS démontre une validité apparente.
La validité de construction fait référence à la capacité du paradigme à évaluer avec précision ce qu'il se propose de mesurer. Dans la prise de décision basée sur le risque / récompense et le délai, la préférence pour la récompense la plus importante diminue au fur et à mesure que la probabilité d'accouchement diminue, ou que le retard dans la livraison de la récompense augmente, respectivement. Dans la prise de décision basée sur l'effort, les individus manifestent une préférence initiale pour l'option d'effort élevé / grande récompense lorsque l'effort associé à la récompense importante est jugé raisonnable. Un changement de préférence au faible effort / à la petite récompense est observé lorsque le gros effort ne vaut plus la dépense énergétique. Il est bien établi que les rongeurs présentent des schémas similaires de prise de décision en termes de risque / rendement, basés sur les délais et sur l'effort, par rapport aux humains (Rachlin et al., 1991; Buelow et Suhr, 2009; Jimura et al., 2009), et nous avons observé ces profils dans chacune de nos tâches (Rokosik et Napier, 2011, 2012; Tedford et al., 2012; Personnes et al., 2013) (par exemple, voir la figure 2).
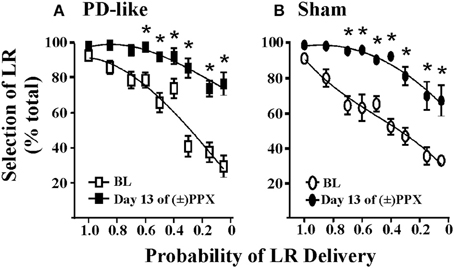
Figure 2. Effets du pramipexole sur la prise de décision en matière de risque / récompense en utilisant une tâche d’actualisation de la probabilité. Chronique (±) PPX diminue l'actualisation dans les situations de type PD (A) et contrôle simulé (B) les rats. Décrivant brièvement la tâche, de type PD (n = 11) et contrôle simulé (n = 10) des rats ont été formés à la tâche d’actualisation de probabilité en utilisant ICSS. Les probabilités associées à la livraison du gros renforçant (LR) ont été présentées dans un ordre pseudo-randomisé. Une fois qu'un comportement stable a été observé, les rats ont été traités de manière chronique avec des injections deux fois par jour de 2 en mg / kg (±) PPX pendant les jours 13. Les données présentées ont été collectées à partir du moment où nous avons observé l'effet maximal le dernier jour de traitement (c.-à-d. 6 h après l'injection) et sont comparées au niveau de référence avant traitement (BL). Le pourcentage de sélection du rapport de résolution (c.-à-d. Le ratio de choix libre) par rapport à la probabilité que le rapport de requête ait été livré est indiqué. Un rmANOVA à deux voies avec post hoc Newman-Keuls a révélé une augmentation significative du pourcentage de sélection de la LR incertaine après un traitement chronique au PPX (*p <0.05) pour les deux groupes de rats PD et simulés. Bien que les moyennes du groupe indiquent une augmentation induite par PPX dans la prise de décision risque / récompense sous-optimale, deux rats dans chaque groupe ont montré une augmentation de moins de 20% par rapport à la ligne de base à la plus faible probabilité testée; par conséquent, certains rats semblaient insensibles à la capacité du médicament à modifier la réduction de probabilité. Figure modifiée de Rokosik et Napier (2012) et réimprimé avec la permission de l'éditeur.
La validité prédictive fait référence à la capacité des modèles à prévoir les relations futures, et nous postulons que nos modèles peuvent être utilisés pour prédire la capacité de nouveaux traitements pharmacologiques à modifier la prise de décision coût / bénéfice. Autrement dit, en démontrant la preuve de concept en reproduisant les effets des agents pharmacologiques sur les comportements de prise de décision déjà établis chez l'homme, nous proposons que nos modèles puissent être efficaces pour prédire comment d'autres médicaments peuvent médier ces comportements en clinique. . Par exemple, un sous-ensemble de patients atteints de la maladie de Parkinson (MP) qui sont traités avec des thérapies par agonistes dopaminergiques démontrent une prévalence accrue du comportement de jeu (Weintraub et al., 2010) et l’escompte accru des prises de décision fondées sur les retards (Housden et al., 2010; Milenkova et al., 2011; Voon et al., 2011; Leroi et al., 2013; Szamosi et al., 2013). Ainsi, notre laboratoire s’est efforcé de modéliser la MP chez le rat et d’étudier les effets du pramipexole, un agoniste de la dopamine couramment employé et associé à des comportements de jeu (Weintraub et al., 2010), sur la prise de décision coût / bénéfice chez le rat utilisant la tâche d’actualisation de probabilité (prise de décision risque / récompense) (Rokosik et Napier, 2012). Pour ce faire, les rats ont été rendus «semblables à la PD» par lésion sélective de terminaux dopaminergiques dans le striatum dorsolatéral. via perfusions bilatérales de 6-OHDA, tandis que les rats témoins recevaient des perfusions du véhicule 6-OHDA (Rokosik et Napier, 2012). Les neurones du striatum dorsolatéral des rats traités par 6-OHDA présentent une diminution de la tyrosine hydroxylase (Rokosik et Napier, 2012), un marqueur de la dopamine. Les rats de type PD présentent des perturbations motrices similaires à celles des humains atteints de MP à un stade précoce, qui peuvent être inversés en fonction de la dose avec le traitement au pramipexole. La dose de pramipexole que nous avons administrée pour étudier la prise de décision en matière de risque / récompense atténue les déficits moteurs et est donc pertinente du point de vue thérapeutique (Rokosik et Napier, 2012). Bien que nous n'ayons trouvé aucune différence dans le comportement «à risque» de base entre les rats témoins et les rats analogues à la MP, le traitement chronique au pramipexole augmente la sélection de la LR à risque dans les deux groupes de rats lorsque les probabilités d'accouchement étaient faibles (Figures 1). 2A, B), indiquant que le pramipexole induit une prise de décision risque / rendement non optimale. Ces données concordent avec les études évaluant les effets du pramipexole chez l’homme (Spengos et al., 2006; Pizzagalli et al., 2008; Riba et al., 2008). Néanmoins, nous déduisons la validité prédictive de nos modèles de rongeurs pour indiquer d'autres agents pharmacologiques susceptibles d'intervenir dans la prise de décision coût / bénéfice chez l'homme.
Nous avons également testé la mirtazapine, un antidépresseur atypique, dans le cadre d'une prise de décision basée sur les efforts. Les dépendances comportementales et la toxicomanie partagent de nombreuses caractéristiques qui se chevauchent, y compris une prise de décision sous-optimale, et de nouvelles études sur des humains et des animaux non-humains montrent que la mirtazapine est efficace pour réduire les comportements motivés par des drogues maltraitées (par exemple, opiacés et psychostimulants), même ceux associés à rechute pendant les périodes d’abstinence (pour examen, voir Graves et al., 2012). Les données recueillies lors de notre tâche de prise de décision reposant sur les efforts de médiation ICSS indiquent que la mirtazapine a effectivement réduit la préférence pour un effort élevé / LR, passant à un effort faible / SR, ce qui suggère que l'effort requis pour le LR n'était plus ", ou que la valeur de récompense de la LR était diminuée (Personnes et al., 2013). Ces résultats suggèrent qu'il pourrait être intéressant d'étudier les effets de la mirtazapine sur la prise de décision sous-optimale chez les joueurs problématiques en clinique.
Conclusion
En résumé, nous avons utilisé l’ICSS comme agent de renforcement positif dans plusieurs nouvelles tâches conçues pour mesurer des aspects distincts, mais qui se chevauchent, de la prise de décision en matière de coûts / avantages, présentés dans le jeu problématique. Ces mesures peuvent être utilisées pour explorer plus avant la contribution de divers substrats neuroanatomiques et de systèmes de neurotransmetteurs dans le jeu problématique. Les tâches à médiation ICSS constituent une alternative viable au renforcement des aliments dans ces paradigmes opérationnels complexes. Nous pensons que la validité de ces tâches indique qu’elles peuvent contribuer au dépistage des médicaments susceptibles de provoquer des troubles du contrôle de l’impulsion, tels que le jeu pathologique, et à l’identification des médicaments susceptibles de réduire ces troubles.
Déclaration de conflit d'intérêts
Le Dr Napier a bénéficié du soutien de la recherche des Instituts nationaux de la santé, de la Fondation Michael J. Fox et du Centre national de jeu responsable. Le D r Napier a reçu une indemnité pour: la consultation d’un centre d’éducation pour la santé à but non lucratif et de cabinets d’avocats sur des questions liées à la toxicomanie et aux troubles du contrôle de l’impulsion; prendre la parole sur les dépendances lors d'assemblées publiques locales, d'écoles secondaires publiques, d'organismes communautaires à but non lucratif et de réunions professionnelles de tribunaux pour toxicomanes; fournir des revues de subventions pour les Instituts nationaux de la santé et d'autres agences; et des conférences académiques et des séances scientifiques. La Dre Napier est membre de l'Illinois Alliance on Problem Gambling et elle fournit des conseils d'expert sur le développement de médicaments à la Cures Within Research Foundation. Les docteurs Holtz, Persons et Mme Tedford déclarent que la recherche a été menée en l'absence de relations commerciales ou financières qui pourraient être interprétées comme un conflit d'intérêts potentiel.
Remerciements
Ce travail a été soutenu par le Centre national pour le jeu responsable, la Fondation Michael J. Fox, la Fondation Daniel F. et Ada L. Rice et USPHSGs NS074014 à T. Celeste Napier et DA033121 à Stephanie E. Tedford et T. Celeste Napier .
Bibliographie
Alessi, SM et Petry, NM (2003). La gravité du jeu pathologique est associée à l’impulsivité dans une procédure d’escompte de retard. Comportement Les processus 64, 345–354. doi: 10.1016/S0376-6357(03)00150-5
Bickel, WK, Jarmolowicz, DP, Mueller, ET, Koffarnus, MN et Gatchalian, KM (2012). Remises excessives sur les agents de renforcement retardés en tant que processus trans-maladie contribuant à la toxicomanie et à d'autres vulnérabilités liées à la maladie: preuves émergentes. Pharmacol Ther. 134, 287 – 297. doi: 10.1016 / j.pharmthera.2012.02.004
Bizo, LA, Bogdanov, SV et Killeen, PR (1998). La satiété entraîne une diminution de la réponse instrumentale au cours de la session. J. Exp. Psychol. Anim. Comportement Processus. 24, 439 – 452. doi: 10.1037 / 0097-7403.24.4.439
Buckholtz, JW, Treadway, MT, Cowan, RL, Woodward, ND, Li, R., Ansari, MS, et al. (2010). Différences de réseau dopaminergique dans l'impulsivité humaine. Sciences 329, 532. doi: 10.1126 / science.1185778
Buelow, MT, et Suhr, JA (2009). Construire la validité de la tâche de jeu de l'Iowa. Neuropsychol. Tour. 19, 102 – 114. Paramètres: 10.1007 / s11065-009-9083-4
Corbett, D., et Wise, RA (1980). Autostimulation intracrânienne en relation avec les systèmes dopaminergiques ascendants du cerveau moyen: une étude de cartographie des électrodes mobiles. Brain Res. 185, 1–15. doi: 10.1016/0006-8993(80)90666-6
Crean, JP, de, WH et Richards, JB (2000). L’escompte des récompenses en tant que mesure du comportement impulsif dans une population ambulatoire psychiatrique. Exp. Clin. Psychopharmacol. 8, 155 – 162. doi: 10.1037 / 1064-1297.8.2.155
Dai, Z., Harrow, SE, Song, X., Rucklidge, J. et Grace, R. (2013). Réductions sur le jeu, les retards et les probabilités chez les adultes avec et sans TDAH. J. Atten. Désordre. doi: 10.1177 / 1087054713496461. [Epub ahead of print].
Dixon, MR, Marley, J. et Jacobs, EA (2003). Retarder l'escompte par les joueurs pathologiques. J. Appl. Comportement Anal. 36, 449 – 458. doi: 10.1901 / jaba.2003.36-449
Douglas, VI et Parry, PA (1994). Effets de récompense et de non récompense sur la frustration et l'attention dans le trouble du déficit de l'attention. J. anormal. Child Psychol. 22, 281 – 302. doi: 10.1007 / BF02168075
Eubig, PA, Noé, TE, Floresco, SB, Sable, JJ et Schantz, SL (2014). Différences entre les sexes en réponse à l'amphétamine chez des rats adultes à long evans effectuant une opération de remise à plus tard. Pharmacol. Biochem. Comportement. 118, 1 – 9. doi: 10.1016 / j.pbb.2013.12.021
Feja, M. et Koch, M. (2014). L'inactivation du cortex préfrontal médial ventral affecte le contrôle des impulsions mais n'affecte pas la réduction du délai chez les rats. Comportement Brain Res. 264, 230 – 239. doi: 10.1016 / j.bbr.2014.02.013
Graves, SM, Rafeyan, R., Watts, J. et Napier, TC (2012). La mirtazapine et des composés analogues à la mirtazapine pourraient être utilisés en pharmacothérapie pour traiter les troubles liés à l’abus de substances psychoactives: données probantes au banc et au chevet du patient. Pharmacol. Ther. 136, 343 – 353. doi: 10.1016 / j.pharmthera.2012.08.013
Holt, DD, Green, L. et Myerson, J. (2003). L'actualisation est-elle impulsive? Preuves tirées de l’escompte temporel et de la probabilité de probabilité chez les étudiants de niveau universitaire qui jouent ou non. Comportement Les processus 64, 355–367. doi: 10.1016/S0376-6357(03)00141-4
Hosking, JG, Cocker, PJ et Winstanley, CA (2014). Contributions dissociables du cortex cingulaire antérieur et de l'amygdale basolatérale sur une tâche décisionnelle de coût / bénéfice chez un rongeur nécessitant un effort cognitif. Neuropsychopharmacology 39, 1558 – 1567. doi: 10.1038 / npp.2014.27
Housden, CR, O'Sullivan, SS, Joyce, EM, Lees, AJ et Roiser, JP (2010). Apprentissage de récompense intact mais réduction des délais élevés chez les patients atteints de la maladie de Parkinson avec des comportements à spectre impulsif-compulsif. Neuropsychopharmacology 35, 2155 – 2164. doi: 10.1038 / npp.2010.84
Jimura, K., Myerson, J., Hilgard, J., Braver, TS et Green, L. (2009). Les gens sont-ils vraiment plus patients que les autres animaux? la preuve obtenue par la remise par l'homme de prix réels en liquide. Psychon. Taureau. Tour. 16, 1071 – 1075. doi: 10.3758 / PBR.16.6.1071
Johnson, PS, Madden, GJ, Brewer, AT, Pinkston, JW et Fowler, SC (2011). Effets du pramipexole aigu sur la préférence pour des schémas de renforcement similaires à ceux du jeu chez le rat. Psychopharmacologie (Berl) 231, 11–18. doi: 10.1007/s00213-010-2006-5
Keesey, RE (1962). La relation entre la fréquence, l'intensité et la durée du pouls et le taux de réponse à la stimulation intracrânienne. J. Comp. Physiol. Psychol. 55, 671 – 678. doi: 10.1037 / h0049222
Koffarnus, MN, Newman, AH, Grundt, P., Rice, KC et Woods, JH (2011). Effets de composés dopaminergiques sélectifs sur une tâche de réduction de délai. Comportement Pharmacol. 22, 300 – 311. doi: 10.1097 / FBP.0b013e3283473bcb
Kraplin, A., Dshemuchadse, M., S. Behrendt, S., Scherbaum, S., T. Goschke et G. Buhringer (2014). Prise de décision dysfonctionnelle dans le jeu pathologique: spécificité des modèles et rôle de l'impulsivité. Res psychiatrie. 215, 675 – 682. doi: 10.1016 / j.psychres.2013.12.041
Ledgerwood, DM, Alessi, SM, Phoenix, N. et Petry, NM (2009). Évaluation comportementale de l'impulsivité chez les joueurs pathologiques avec et sans antécédents de trouble lié à l'utilisation de substances par rapport à des témoins sains. La drogue dépend de l'alcool. 105, 89 – 96. doi: 10.1016 / j.drugalcdep.2009.06.011
Leroi, I., Barraclough, M., McKie, S., Hinvest, N., Evans, J., Elliott, R., et McDonald, K. (2013). Influences dopaminergiques sur la fonction exécutive et le comportement impulsif dans les troubles du contrôle des impulsions dans la maladie de Parkinson. J. Neuropsychol. 7, 306 – 325. doi: 10.1111 / jnp.12026
Madden, GJ, Johnson, PS, Brewer, AT, Pinkston, JW et Fowler, SC (2010). Effets du pramipexole sur le choix impulsif chez le rat wistar mâle. Exp. Clin. Psychopharmacol. 18, 267 – 276. doi: 10.1037 / a0019244
Madden, GJ, Petry, NM et Johnson, PS (2009). Les joueurs pathologiques ont des remises probabilistes moins élevées que les témoins appariés. Exp. Clin. Psychopharmacol. 17, 283 – 290. doi: 10.1037 / a0016806
McCown, TJ, Napier, TC et Breese, GR (1986). Effets de l'implantation chronique d'électrodes sur les neurones dopaminergiques in vivo. Pharmacol. Biochem. Comportement 25, 63–69. doi: 10.1016/0091-3057(86)90231-5
Mejia-Toiber, J., N. Boutros, A. Markou et S. Semenova (2014). Choix impulsif et comportement semblable à l'anxiété chez les rats adultes exposés à l'éthanol intermittent chronique à l'adolescence et à l'âge adulte. Comportement Brain Res. 266, 19 – 28. doi: 10.1016 / j.bbr.2014.02.019
Michalczuk, R., H. Bowden-Jones, A. Verdejo-Garcia et L. Clark (2011). Impulsivité et distorsions cognitives chez les joueurs pathologiques fréquentant la clinique britannique du jeu problématique: rapport préliminaire. Psychol. Med. 41, 2625 – 2635. doi: 10.1017 / S003329171100095X
Miedl, SF, Peters, J. et Buchel, C. (2012). Représentations altérées de la récompense neuronale chez les joueurs pathologiques révélées par un délai et une réduction de probabilité. Cambre. Psychiatrie générale 69, 177 – 186. doi: 10.1001 / archgenpsychiatry.2011.1552
Milenkova, M., Mohammadi, B., Kollewe, K., Schrader, C., Fellbrich, A., Wittfoth, M., et al. (2011). Choix intertemporel dans la maladie de Parkinson. Mov. Désordre. 26, 2004 – 2010. doi: 10.1002 / mds.23756
Minamimoto, T., La Camera, G. et Richmond, BJ (2009). Mesurer et modéliser l'interaction entre la taille de la récompense, le délai de récompense et le niveau de satisfaction sur la motivation chez les singes. J. Neurophysiol. 101, 437 – 447. doi: 10.1152 / jn.90959.2008
Olds, J. et Milner, P. (1954). Renforcement positif produit par une stimulation électrique de la zone septale et d'autres régions du cerveau de rat. J. Comp. Physiol. Psychol. 47, 419 – 427. doi: 10.1037 / h0058775
Papini, MR et Dudley, RT (1997). Conséquences des omissions de récompense surprenantes. Rev. Gen. Psychol. 1, 175 – 197. doi: 10.1037 / 1089-2680.1.2.175
Persons, AL, Tedford, SE et Napier, TC (2013). «Évaluations précliniques de composés sérotoninergiques à fonctions mixtes pour le traitement des troubles du jeu. numéro de programme 635.12 ”, dans Neuroscience Meeting Planner (San Diego, CA: Society for Neuroscience).
Petry, NM (2011). L'abattement des récompenses probabilistes est associé à l'abstinence du jeu chez les joueurs pathologiques en quête de traitement. J. anormal. Psychol. 121, 151 – 159. doi: 10.1037 / a0024782
Pirch, JH, Napier, TC et Corbus, MJ (1981). Stimulation cérébrale comme indice des potentiels liés à un événement dans le cortex de rat: effets de l'amphétamine. Int. J. Neurosci. 15, 217 – 222.
Pizzagalli, DA, AE, Schetter, EC, Frank, MJ, Pajtas, PE, Santesso, DL, et al. (2008). Une seule dose d'un agoniste de la dopamine altère l'apprentissage par renforcement chez l'homme: mise en évidence du comportement à partir d'une mesure en laboratoire de la réactivité à la récompense. Psychopharmacologie (Berl) 196, 221–232. doi: 10.1007/s00213-007-0957-y
Rachlin, H., Raineri, A. et Cross, D. (1991). Probabilité et retard subjectifs. J. Exp. Anal. Comportement. 55, 233 – 244. doi: 10.1901 / jeab.1991.55-233
Rasmussen, EB, avocat, SR, et Reilly, W. (2010). Le pourcentage de graisse corporelle est lié au retard et à la probabilité d'escompte pour l'alimentation chez l'homme. Comportement Les processus 83, 23 – 30. doi: 10.1016 / j.beproc.2009.09.001
Reynolds, B., Richards, JB, Horn, K. et Karraker, K. (2004). Retard et remise de probabilité en fonction du statut de tabagisme chez les adultes. Comportement Les processus 65, 35–42. doi: 10.1016/S0376-6357(03)00109-8
Riba, J., Kramer, UM, Heldmann, M., Richter, S. et Munte, TF (2008). L'agoniste de la dopamine augmente la prise de risque mais atténue l'activité cérébrale liée aux récompenses. PLoS. UN 3: e2479. doi: 10.1371 / journal.pone.0002479
Rokosik, SL et Napier, TC (2011). L'auto-stimulation intracrânienne en tant que renforçateur positif pour étudier l'impulsivité dans un paradigme de réduction de probabilité. J. Neurosci. Les méthodes 198, 260 – 269. doi: 10.1016 / j.jneumeth.2011.04.025
Rokosik, SL et Napier, TC (2012). Réduction probabiliste accrue induite par le pramipexole: comparaison entre un modèle de rongeur de la maladie de Parkinson et des témoins. Neuropsychopharmacology 37, 1397 – 1408. doi: 10.1038 / npp.2011.325
Roll, JM, McSweeney, FK, Johnson, KS et Weatherly, JN (1995). La satiété contribue peu à la diminution de la réponse au sein de la session. Apprendre. Motiv. 26, 323–341. doi: 10.1016/0023-9690(95)90018-7
Schuck-Paim, C., L. Pompilio et A. Kacelnik (2004). Les décisions dépendant de l'État provoquent des violations apparentes de la rationalité dans le choix des animaux. PLoS. Biol. 2: e402. doi: 10.1371 / journal.pbio.0020402
Spengos, K., Grips, E., G. Karachalios, G. Tsivgoulis et G. Papadimitriou (2006). Jeu pathologique réversible sous traitement par le pramipexole. Nervenarzt 77, 958–960. doi: 10.1007/s00115-006-2128-5
St Onge, JR et Floresco, SB (2009). Modulation dopaminergique de la prise de décision basée sur le risque. Neuropsychopharmacology 34, 681 – 697. doi: 10.1038 / npp.2008.121
Stopper, CM et Floresco, SB (2011). Contributions du noyau accumbens et de ses sous-régions à différents aspects de la prise de décision fondée sur les risques. Cogn. Affecter. Comportement Neurosci. 11, 97 – 112. Paramètres: 10.3758 / s13415-010-0015-9
Szamosi, A., Nagy, H., et Keri, S. (2013). Retarder l'actualisation de la récompense et du volume du noyau caudé chez les individus atteints de duplication du gène alpha-synucléine avant et après le développement de la maladie de Parkinson. Neurodégénére. Dis. 11, 72 – 78. doi: 10.1159 / 000341997
Tedford, SE, Personnes, AL, L. Chen, Rokosik, SL et Napier, TC (2012). «L'auto-stimulation intracrânienne est le renforcement positif pour étudier l'impulsivité dans un paradigme d'actualisation différée chez le rat. programme n ° 926.03, ”dans Neuroscience Meeting Planner (Nouvelle-Orléans, LA: Society for Neuroscience).
Tehovnik, EJ et Sommer, MA (1997). Étalement efficace et délai d'inactivation neuronale causée par l'injection de lidocaïne dans le cortex cérébral de singe. J. Neurosci. Les méthodes 74, 17–26. doi: 10.1016/S0165-0270(97)02229-2
Treadway, MT, Buckholtz, JW, Schwartzman, AN, Lambert, WE et Zald, DH (2009). Vaut le «EEfRT»? La dépense d'effort pour la tâche de récompenses comme mesure objective de la motivation et de l'anhédonie. PLoS. UN 4: e6598. doi: 10.1371 / journal.pone.0006598
Trowill, JA, Panksepp, J. et Gandelman, R. (1969). Un modèle d'incitation à la stimulation cérébrale enrichissante. Psychol. Tour. 76, 264 – 281. doi: 10.1037 / h0027295
Voon, V., Sohr, M., Lang, AE, Potenza, MN, Siderowf, AD, Whetteckey, J., et al. (2011). Troubles du contrôle des impulsions dans la maladie de Parkinson: étude multicentrique cas-contrôle. Ann. Neurol. 69, 986 – 996. doi: 10.1002 / ana.22356
Wardle, MC, Treadway, MT, Mayo, LM, Zald, DH et de, WH (2011). Accroître l'effort: effets de la d-amphétamine sur la prise de décision basée sur l'effort humain. J. Neurosci. 31, 16597 – 16602. doi: 10.1523 / JNEUROSCI.4387-11.2011
Weintraub, D., Koester, J., Potenza, MN, Siderowf, AD, Stacy, M., Voon, V., et al. (2010). Troubles du contrôle des impulsions dans la maladie de Parkinson: étude transversale sur des patients traités par 3090. Cambre. Neurol. 67, 589 – 595. doi: 10.1001 / archneurol.2010.65
Sage, RA (1996). Drogues addictives et récompense de stimulation cérébrale. Annu. Rev. Neurosci. 19, 319 – 340. doi: 10.1146 / annurev.neuro.19.1.319
Sage, RA (2005). Substrat du cerveau antérieur de récompense et de motivation. J. Comp. Neurol. 493, 115 – 121. doi: 10.1002 / cne.20689
Wise, RA, P. Bauco, Carlezon, WA Jr. et Trojniar, W. (1992). Mécanismes d'auto-stimulation et de récompense des médicaments. Ann. NY Acad. Sci. 654, 192 – 198. fichier: 10.1111 / j.1749-6632.1992.tb25967.x
Wise, RA et Bozarth, MA (1981). Substrats du cerveau pour le renforcement et l’auto-administration du médicament. Programme. Neuropsychopharmacol. 5, 467 – 474. Paramètres: 10.1016 / 0364-7722 (81) 90028-X
Wise, RA et Rompre, PP (1989). Dopamine cérébrale et récompense. Annu. Rev. Psychol. 40, 191 – 225. doi: 10.1146 / annurev.psych.40.1.191
Zeeb, FD, Robbins, TW et Winstanley, CA (2009). Modulation sérotoninergique et dopaminergique du comportement de jeu évaluée à l'aide d'une nouvelle tâche de jeu chez le rat. Neuropsychopharmacology 34, 2329 – 2343. doi: 10.1038 / npp.2009.62
Mots-clés: prise de décision coût / bénéfice, actualisation, prise de décision basée sur l'effort, jeu, autostimulation intracrânienne
Citation: Tedford SE, Holtz NA, Persons AL et Napier TC (2014) Une nouvelle approche pour évaluer le comportement de jeu chez le rat de laboratoire: utiliser l'auto-stimulation intracrânienne comme renforçant positif. De face. Comportement Neurosci. 8: 215. doi: 10.3389 / fnbeh.2014.00215
Reçu: 06 March 2014; Accepté: 27 May 2014;
Publié en ligne: 11 June 2014.
Édité par:
Patrick Anselme, Université de Liège, Belgique
Commenté par:
Christelle BaunezCentre National de la Recherche Scientifique, France
Yueqiang Xue, Centre des sciences de la santé de l'Université du Tennessee, États-Unis
