Neurosci avant. 2015 Mar 18; 9: 86. doi: 10.3389 / fnins.2015.00086. eCollection 2015.
Terbeck S1, Akkus F2, Chesterman LP3, Hasler G2.
Abstract
Dans la présente revue, nous donnons un aperçu de l'implication de l'activité et de la densité du récepteur métabotropique du glutamate 5 (mGluR5) dans l'anxiété pathologique, les troubles de l'humeur et la dépendance. Plus précisément, nous décrirons les études sur le mGluR5 chez l'homme qui ont utilisé la tomographie par émission de positrons (TEP) et combiné les résultats avec la recherche préclinique sur les animaux. Cette vue combinée de différentes approches méthodologiques - des approches neurobiologiques de base aux études humaines - pourrait donner une vue plus complète et cliniquement pertinente de la fonction mGluR5 en santé mentale que la vue sur les données précliniques seules. Nous examinerons également les données de recherche actuelles sur mGluR5 selon les critères du domaine de recherche (RDoC). Premièrement, nous avons trouvé des preuves d'une activité glutamate anormale liée aux systèmes de valence positif et négatif, ce qui suggérerait que l'intervention antagoniste mGluR5 a des effets anti-addictifs, anti-dépressifs et anxiolytiques importants. Deuxièmement, il est prouvé que le mGluR5 joue un rôle important dans les systèmes de fonctionnement social et la réponse au stress social. Enfin, le rôle important de mGluR5 dans l'homéostasie du sommeil suggère que ce récepteur du glutamate pourrait jouer un rôle important dans le domaine des systèmes d'excitation et de modulation de RDoC. Le glutamate était auparavant principalement étudié dans des études non humaines, mais la recherche clinique initiale sur la TEP sur l'homme soutient désormais également l'hypothèse que, en médiant l'excitabilité cérébrale, la neuroplasticité et la cognition sociale, une activité anormale du glutamate métabotrope pourrait prédisposer les individus à un large éventail de problèmes psychiatriques.
Introduction
Le glutamate est le principal neurotransmetteur excitateur dans le cerveau et de nombreux chercheurs ont suggéré qu'il joue un rôle important dans diverses maladies mentales et médicales. En effet, les chercheurs mentionnent souvent le potentiel de développement d'un traitement pharmacologique à base de glutamate ionotrope ou métabotrope pour de nombreux troubles psychiatriques, sous la forme d'agonistes ou d'antagonistes. Les connaissances sur le système glutamatergique ont considérablement progressé au cours de la dernière décennie, grâce aux avancées technologiques dans l'imagerie des récepteurs et des émetteurs chez l'homme. Bien qu'il existe un grand nombre d'études expérimentales sur l'intervention du glutamate dans divers troubles psychiatriques, il n'existe pas de revues systématiques axées sur la combinaison des résultats d'études de recherche préclinique sur des animaux avec d'autres méthodes neuroscientifiques, telles que la tomographie par émission de positrons (TEP) humaine. . Ainsi, par rapport aux avis précédents (Swanson et al., 2005; Pittenger et al., 2006; Kalivas, 2009; Brennan et al., 2012; Luykx et al., 2012; Riaza Bermudo-Soriano et al., 2012), cette revue examine les études humaines, principalement la recherche sur le PET, et combine ces résultats avec des résultats précliniques.
Tomographie par émission de positrons (PET)
La TEP est une technique d'imagerie médicale nucléaire, sensible et non invasive utilisée pour visualiser la distribution, la concentration et la fonction des récepteurs. Pour identifier les récepteurs du cerveau humain, il faut développer des ligands de récepteurs marqués par radio (traceurs). Le scanner détecte les rayons gamma émis par le traceur, qui est introduit dans le corps. Un traceur PET mGluR5 adapté aux études sur l'homme a été mis au point avec succès à l'Institut Paul Scherrer (PSI) de Villigen (Suisse) et à l'Ecole Polytechnique Fédérale de Suisse (ETH) à Zurich, en Suisse. ABP688 est un antagoniste non sélectif et hautement sélectif, qui se lie à un site allostérique du mGluR5. 11C-ABP688 a montré une sélectivité élevée pour mGluR5 et une absorption élevée dans les régions cérébrales riches en récepteurs. La première description de ces caractéristiques de 11C-ABP688 chez l’animal a déjà été publiée (Ametamey et al., 2006). Nous avons vu des résultats prometteurs provenant d'études menées chez le rat utilisant une sonde bêta pour estimer la cinétique de ce traceur. De plus, lorsque nous avons effectué la première étude TEP chez l’homme, nous avons estimé la cinétique chez l’homme (Ametamey et al., 2006), nous avons trouvé des résultats comparables à ceux des études chez le rat (Soares et Law, 2009). Nous avons maintenant utilisé avec succès le traceur développé pour la recherche sur mGluR5 chez des volontaires sains et des patients psychiatriques. Les techniques PET fournissent des informations sur la densité relative des récepteurs dans la zone du cerveau examinée. Il n'y a aucune information concernant la concentration du neurotransmetteur (c'est-à-dire, si une densité de récepteurs élevée pourrait entraîner ou être la conséquence d'une action accrue ou réduite des neurotransmetteurs). La recherche TEP fournit des informations sur les anomalies du niveau des récepteurs chez les patients humains vivants et permet ainsi d’évaluer quels récepteurs doivent être ciblés par un traitement pharmaceutique. Au contraire, la spectroscopie par résonance magnétique (SRM) fournit in vivo informations biochimiques sur le tissu examiné (Hasler et al., 2009) et fournit donc des informations sur la quantité relative de métabolite du glutamate dans des groupes de patients. Les résultats des recherches PET et MRS peuvent apporter des preuves supplémentaires aux études pré-cliniques sur les animaux. La recherche animale est la méthode de recherche préclinique à la pointe de la technologie. Divers modèles animaux de conditions psychiatriques ont été établis. Les études chez l'animal permettent de tester des modèles mécaniques et de tester de nouveaux produits pharmaceutiques dans ces modèles afin de réduire les risques potentiels pour l'homme. Un des inconvénients de la recherche animale en psychiatrie pourrait toutefois résider dans le développement de modèles animaux adaptés à différentes conditions psychiatriques, en particulier, des conditions impliquant des aspects cognitifs humains internes ou uniques pourraient entraîner des problèmes. Ainsi, un nouveau médicament qui montre une amélioration du comportement des animaux pourrait ne pas aider les aspects psychologiques internes de l’état psychiatrique.
Cet article examinera donc si les résultats de la recherche préclinique chez l'animal et de la recherche TEP en psychiatrie humaine forment une vision cohérente de l'implication de mGluR5 dans les troubles mentaux. Cette revue ne traitera que de l'activité de mGluR5 car de nombreuses données humaines sur la TEP sont disponibles pour ce type de récepteur. mGluR5 constitue une cible prometteuse pour le développement de médicaments, car un traceur PET peut mesurer la liaison de mGluR5 chez l'homme, offrant ainsi une meilleure compréhension de ses fonctions chez l'homme. En effet, les recherches suggèrent que les médicaments ciblant les récepteurs métabotropiques du glutamate du groupe I figurent parmi les agents les plus prometteurs actuellement en développement pour le traitement des maladies psychiatriques (Krystal et al., 2010).
Dans cette revue, nous commençons par une brève introduction du système du glutamate et de mGluR5, puis nous évaluons l'implication de mGluR5 dans les troubles anxieux et de l'humeur et la dépendance en comparant des essais précliniques antérieurs à des études PET récentes. Nous décrirons ensuite une approche permettant de déterminer l'efficacité des interventions basées sur mGluR5 en contribuant à des changements d'apprentissage et du fonctionnement social et en diminuant l'excitabilité dans diverses régions du cerveau.
Le système glutamatergique et mGluR5
Le glutamate régule la fonction du système nerveux central grâce aux actions des récepteurs ionotropes et métabotropes. L'implication du glutamate dans diverses conditions psychiatriques et médicales a fait l'objet d'un examen approfondi. Cependant, les travaux antérieurs portaient principalement sur les récepteurs ionotropes du glutamate. Contrairement aux actions rapides et directes des récepteurs ionotropes, les trois groupes de récepteurs métabotropes (mGlu) modifient l'activité neuronale par le biais d'une signalisation couplée à la protéine G. Les groupes de mGluR se distinguent par leurs propriétés de signalisation pharmacologiques et intracellulaires. mGluR5, qui a été cloné pour la première fois chez des animaux sous 1992 et quelques années plus tard chez l'homme, appartient aux récepteurs métabotropes du groupe I (Olive, 2005). Ses actions sont principalement excitatrices (Meldrum, 2000). Cleva et Olive (2011) ont décrit de fortes liaisons et interactions de récepteurs entre mGluR5 et le récepteur NMDA, suggérant que mGluR5 pourrait également être largement impliqué dans la médiation de la plasticité neurale ainsi que dans les processus d'apprentissage et de mémoire. En outre, il existe certaines preuves que l’activation de mGluR5 améliore le GABA, en particulier dans le noyau accumbens (Hoffpauir et Gleason, 2002). Ainsi, il est suggéré que l'activité du récepteur métabotrope du glutamate puisse moduler les voies de signalisation excitatrices et inhibitrices (GABA). Une densité élevée de récepteurs mGluR5 a été identifiée principalement dans les régions du cerveau antérieur, du striatum et des régions limbiques, y compris l'amygdale et l'hippocampe (Swanson et al., 2005). En utilisant des techniques de biologie moléculaire avancées pour déterminer l’expression de l’ARNm de mGluR5 dans le cerveau des rongeurs, les recherches ont montré que les régions du bulbe olfactif, du striatum dorsal, du noyau accumbens, du septum latéral et de l’hippocampe présentent les niveaux les plus élevés d’expression de mGluR5 (Abe et al., 1992) (Voir la figure 1).
FIGURE 1
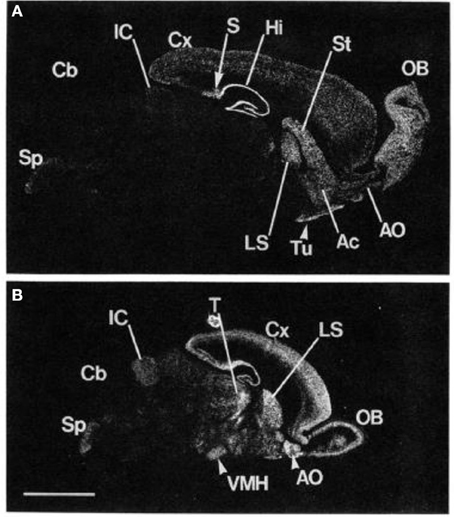
Figure 1. Localisation de l'ARNm de mGluR5 dans le cerveau des rats adultes et 6 âgés de sur place hybridation. Images de film négatives de la section sagittale du cerveau de rat adulte (A) et le cerveau de rat d'un jour 6 (B) sont indiqués. OB, noyau olfactif principal; Ac, noyau accumbens; Tutubercule olfactif; Ststriatum; Hihippocampe; S, subiculum; Cx, cortex cérébral; LS, noyau septal latéral; IC, colliculus inférieur; Cbcortex cérébelleux; Sp, le noyau trijumeau de la colonne vertébrale; Tle thalamus; VMH, noyau hypothalamique ventromédial. Barre d'échelle, 4 mm. De Abe et al. (1992). Remarque: la figure et sa légende sont reproduites avec autorisation.
Les récepteurs pré-synaptiques mGluR5 participent à la régulation de la plasticité synaptique et aux modifications de l’excitabilité neuronale pour le maintien de l’homéostasie (Schoepp, 2001). Fait important, l’expression significative de mGluR5 peut déjà être déterminée lors de la période prénatale de la semaine de gestation (GW) 9 (Boer et al., 2010). De plus, l’expression de mGluR5 était beaucoup plus élevée chez les animaux plus jeunes que chez les adultes (Romano et al., 1996), suggérant qu'une intervention précoce ciblant mGluR5 pourrait avoir des effets préventifs dans les troubles du développement neurologique.
En raison de ses fonctions dans différents processus neuronaux, des anomalies flagrantes du système du glutamate entraînent de graves troubles neurologiques (par exemple, des crises d'épilepsie), alors que de plus petits changements peuvent contribuer à diverses affections psychiatriques (Yüksel et Öngür, 2010). Les efforts visant à développer des médicaments ciblant sélectivement mGluR5 ont commencé à la fin des 90. À ce jour, divers agonistes et antagonistes de mGluR5 ont été mis au point (voir Lea et Faden, 2006 pour plus d'informations concernant le développement de médicaments spécifiques). Par exemple, 3- [2-méthyl-1,3-thiazol-4yl] éthynyl] pyridine (MTEP) est un antagoniste hautement sélectif et non compétitif de mGluR5 qui atteint une occupation du récepteur complète h après l'injection de rats avec une dose de 1 mg / kg (Anderson et al., 2003). Table 1 donne un aperçu de certains médicaments mGluR5 utilisés chez les animaux et de ceux précédemment utilisés dans les premiers essais cliniques sur l'homme.
TABLEAU 1
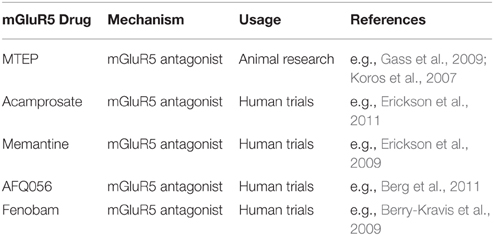
Tableau 1. Sélection de médicaments agissant mGluR5.
Troubles psychiatriques et intervention à base de glutamate chez l'homme
Les troubles psychiatriques sont très hétérogènes et la comorbidité est fréquente. Cependant, les traitements pharmacologiques psychiatriques courants reposent sur relativement peu de mécanismes physiopathologiques, par exemple l'augmentation de la disponibilité de la monoamine dans l'anxiété et la dépression. Il est donc urgent d'améliorer et de faire progresser les traitements psychiatriques, et une intervention pharmacologique à base de glutamate métabotropique constitue un développement prometteur à cet égard (Agid et al., 2007).
La plupart des essais humains ont été menés sur des cas de syndrome du x fragile (FXS). La théorie de FXS basée sur mGluR5 postule que l’absence de protéine de retard mental X fragile (FMRP) entraîne une signalisation glutamatergique excessive via mGluR5 (Bear et al., 2004). Cela conduit à une traduction accrue de l'ARNm local à la synapse, car FMRP n'est pas présent pour réguler le processus. Enfin, cela affaiblit la synapse et entraîne une augmentation du nombre d'épines dendritiques immatures plus longues, ce qui pourrait expliquer la déficience intellectuelle constatée chez les patients atteints de FXS. Cette déficience est associée à des symptômes d'humeur et d'anxiété et présente généralement des caractéristiques communes aux troubles du spectre autistique, notamment des retards dans le développement de la parole et du langage, une altération de la théorie de l'esprit, une altération du traitement social et émotionnel, ainsi qu'un comportement répétitif (Garber et al., 2008). Les preuves préliminaires et indirectes que l'antagoniste de mGluR5 peut améliorer la sociabilité dans FXS (Burket et al., 2014) suscite l’espoir que les médicaments ciblant mGluR5 pourraient être utilisés en clinique dans les affections psychiatriques prévalentes associées à des troubles du système pour des processus sociaux tels que l’autisme, la schizophrénie et la dépression. De plus, le phénotype de comportement répétitif observable dans FXS pourrait suggérer une pathophysiologie partagée parmi d'autres troubles psychiatriques tels que le trouble obsessionnel-compulsif (TOC) et la dépendance.
Bien que les connaissances scientifiques et les recherches sur mGluR5 se soient accrues, les efforts de développement de médicaments ont été relativement infructueux (Agid et al., 2007). Les médicaments qui ciblent les récepteurs ionotropes produisent généralement de nombreux effets secondaires et les stratégies actuelles de développement de médicaments n’ont pas encore produit de cibles sélectives pour les récepteurs ionotropes susceptibles de réduire les effets secondaires potentiels. Par exemple, les antagonistes des récepteurs ionotropes produisent des effets secondaires chez l’homme, notamment des troubles de la mémoire, des épisodes psychotiques et des accidents vasculaires cérébraux (Swanson et al., 2005). Les effets indésirables peuvent être dus au fait que les récepteurs ionotropes du glutamate ont une distribution omniprésente, alors que les récepteurs métabotropes sont plus inégaux et sélectivement distribués (Krystal et al., 2010). En conséquence, le développement récent de médicaments s'est concentré sur des composés ciblant les récepteurs métabotropes, en supposant que ces médicaments seraient associés à moins d'effets secondaires que ceux qui se lient aux récepteurs ionotropes à action rapide.
Dans les sections suivantes, nous examinerons les résultats d'études sur la TEP chez l'homme concernant l'implication de mGluR5 dans les troubles de l'humeur et la toxicomanie et comparerons ces résultats à ceux d'études animales. De plus, nous décrirons certains emplacements du cerveau pour l'activité de mGluR5 chez l'homme et suggérerons enfin une approche de la manière dont l'activité mGluR5 directe et indirecte pourrait être impliquée dans les syndromes psychiatriques humains.
mGluR5, anxiété pathologique et troubles de l'humeur
Anxiété Pathologique
L’anxiété pathologique survient dans les troubles anxieux, y compris le trouble anxieux généralisé, le trouble panique (les troubles psychiatriques les plus répandus dans le monde, First et Gibbon, 1997), mais aussi dans d’autres affections psychiatriques prévalentes telles que la dépression et le trouble obsessionnel-compulsif (TOC). Ces troubles psychiatriques entraînent une altération importante du fonctionnement social et professionnel, entraînant des coûts pour la santé et des souffrances pour les patients (First et Gibbon, 1997). En règle générale, l’anxiété peut être associée à une excitabilité cérébrale excessive (Harvey et Shahid, 2012).
Recherche neurobiologique préclinique
Les résultats d'un grand nombre d'essais précliniques chez l'animal ont permis de déterminer l'effet du traitement par antagoniste de mGluR5 sur l'anxiété. Swanson et al. (2005) ont examiné des études chez l'animal portant sur des médicaments ciblant le mGluR5 sur les comportements anxieux. Ils ont conclu que le traitement antagoniste de mGluR5 entraînait principalement des réponses anxiolytiques chez des animaux de laboratoire. Des effets tels que la diminution de la peur conditionnée au gel, une acceptation accrue des chocs et des punitions et une augmentation des interactions sociales ont été observés. Par exemple, une dose unique de 2-méthyl-6- (phényléthynyl) pyridine (MPEP) a prolongé le temps passé par les rats dans le bras ouvert d'un labyrinthe expérimental, sans affecter le comportement moteur ni la planification (Tatarczyńska et al., 2001). Krystal et al. (2010) ont examiné des études précliniques sur des animaux portant sur des antagonistes du mGluR5 (MTEP, MPEP, fenobam) dans des modèles murins d'anxiété. Ces études ont utilisé différentes mesures de résultats, telles que l’extinction du conditionnement de la peur et des réponses dans le labyrinthe en hauteur, pour évaluer l’efficacité des traitements médicamenteux. Parmi les études examinées, 88.45% a signalé un effet anxiolytique avec les antagonistes de mGluR5 (Krystal et al., 2010). Plus récemment, une autre revue de recherche sur l'anxiété chez des modèles animaux, examinant l'effet d'une intervention antagoniste ionotrope et métabotropique du récepteur du glutamate, a été publiée (Riaza Bermudo-Soriano et al., 2012). En ce qui concerne mGluR5, les auteurs ont répertorié les études 43 sur l'anxiété chez l'animal et toutes sauf deux ont démontré des effets anxiolytiques.
Études humaines
Les premières preuves de l'hypothèse selon laquelle la fonction du glutamate est anormale dans les troubles anxieux sont issues de la recherche sur le SRM. Par exemple, en utilisant la spectroscopie de résonance magnétique 1H à haut champ et à un seul voxel, les chercheurs ont constaté que, par rapport aux témoins sains, les patients présentant un trouble d'anxiété sociale présentaient des taux de glutamate significativement plus élevés dans le cortex cingulaire antérieur (ACC) (Phan et al., 2005). De plus, des recherches ont révélé une augmentation de la concentration globale de glutamate chez les patients 10 atteints de phobie sociale (Pollack et al., 2008). Comme indiqué précédemment, ces études n'ont toutefois pas permis de déterminer quels récepteurs du glutamate étaient associés à une activité excessive du glutamate.
En utilisant la méthodologie de recherche PET, nous avons récemment été le premier à montrer des relations entre mGluR5 et l’anxiété, dans le Trouble dépressif majeur (MDD), et le TOC. Dans une étude, nous avons étudié le rapport de volume de distribution (DVR) mGluR5 chez des patients 10 présentant un contrôle sain de TOC et de 10 à l’aide de [11C] ABP688 PET (Akkus et al., 2014). Nous avons utilisé l'échelle obsessionnelle-compulsive de Yale-Brown (Y-BOCS) comme mesure clinique de la gravité des symptômes du TOC. Nous avons observé des corrélations positives significatives entre les scores d’obsession Y-BOCS et le mGluR5 DVR dans les régions de l’amygdale, de l’ACC et du cortex orbitofrontal médial (Akkus et al., 2014). Ces zones du cerveau ont déjà été impliquées dans la physiopathologie des TOC. En effet, les anomalies cérébrales structurelles de l'amygdale, de l'ACC et du cortex orbitofrontal ont été associées de manière constante au TOC (Rosenberg et Keshavan, 1998; Szeszko et al., 2008; Van den Heuvel et al., 2009). Étant donné que les études d'imagerie structurelle ont fourni des preuves d'une corrélation positive entre la gravité des TOC et le volume de matière grise (Zarei et al., 2011), une augmentation de la liaison de mGluR5 dans les TOC peut refléter une augmentation de la densité des neurones. Bien que, dans le DSM-5, le TOC ne soit plus considéré comme un trouble anxieux, la plupart des patients atteints de TOC présentent des symptômes d’anxiété. Dans un échantillon clinique relativement important, nous avons précédemment montré que les patients souffrant de TOC obsédés avaient une prévalence particulièrement élevée de symptômes et de troubles anxieux (Hasler et al., 2005). Dans cet article, nous avons suggéré que les obsessions impliquant le stress, l’anxiété ou les conflits pourraient donc être associées à une neurotransmission glutamatergique accrue dans l’amygdale, le CAC et le cortex orbitofrontal.
Pris ensemble, les résultats d'études récentes utilisant différentes méthodologies de recherche confirment l'hypothèse selon laquelle la fonction du glutamate serait anormale dans des zones clés du système limbique dans les troubles de l'humeur liés à l'anxiété. La fonction anormale est également susceptible d'être liée au récepteur mGluR5. Étant donné que la recherche pré-clinique et celle sur la TEP ont montré des résultats cohérents, nous suggérons que le traitement antagoniste par mGluR5 produise des effets anxiolytiques significatifs chez les patients souffrant d’anxiété pathologique.
Trouble dépressif majeur (TDM)
Recherche neurobiologique préclinique
Dans leur avis, Krystal et al. (2010) a décrit les résultats de huit études portant sur les antagonistes du mTEP et du MPEP, antagonistes de mGluR5, dans des modèles animaux de dépression. Par rapport aux taux de succès du traitement des troubles anxieux, les auteurs ont indiqué que seul 62.5 – 75% avait trouvé un effet antidépresseur net, même si le traitement avec la kétamine, un antagoniste ionotrope de NMDA, avait entraîné des effets antidépresseurs rapides (même chez les patients résistants).Pittenger et al., 2006).
Études humaines
Une revue récente a examiné les études 13 MRS sur les troubles de l'humeur. Les auteurs ont rapporté que ces études ont systématiquement mis en évidence la réduction du glutamate dans le TDM (Hasler et al., 2007; Yüksel et Öngür, 2010). En particulier, des taux réduits de glutamate dans l'ACC, le cortex préfrontal dorsolatéral gauche, le cortex préfrontal dorsomédial, le cortex préfrontal ventromédial, l'amygdale et l'hippocampe ont été mis en évidence. Un examen ultérieur d'études sur des études utilisant le SRM pour examiner le glutamate dans le TDM a confirmé que la concentration de glutamate dans l'ACC était systématiquement réduite (Luykx et al., 2012). Certains chercheurs ont observé une augmentation du glutamate dans le cortex occipital, plus élevée dans le sous-groupe mélancolique de patients atteints de TDM (Sanacora et al., 2008).
Dans une étude précédente, nous avions obtenu des images TEP de la liaison du récepteur mGluR5 chez des sujets non médicamentés 11 avec des témoins sains MDD et 11 (Deschwanden et al., 2011). Nous avons constaté une diminution de la liaison régionale de mGluR5 dans le cortex préfrontal, le cortex cingulaire, l'insula, le thalamus et l'hippocampe des patients souffrant de dépression. De plus, la sévérité de la dépression était négativement corrélée à la liaison de mGluR5 dans l'hippocampe. Nous avons suggéré que ces résultats indiquent une diminution de la neurotransmission de mGluR5 dans la dépression, possiblement à la suite de modifications basales ou compensatoires de l'activité du système du glutamate. En outre, nous avons examiné la quantité d’expression de mGluR5 dans des échantillons de cerveau post-mortem de sujets déprimés 15 et de témoins appariés à 15 (Deschwanden et al., 2011). Nous avons observé une expression réduite de mGluR5 dans le cortex préfrontal dans des échantillons prélevés chez des personnes dépressives. Une expression réduite des récepteurs NMDA dans le cerveau post-mortem de patients déprimés a également été rapportée (Feyissa et al., 2009). En effet, il a déjà été suggéré que les propriétés antidépressives des antagonistes de mGluR5 pourraient impliquer une inhibition du récepteur NMDA. Cela pourrait induire une neurotransmission et / ou une induction de l’expression du gène du facteur neurotrope dérivé du cerveau dans l’hippocampe (Legutko et al., 2006). En outre, une étude récente a montré que la privation de sommeil augmentait la disponibilité de mGluR5 chez l’homme en bonne santé (Hefti et al., 2013). Dans le cortex cingulaire antérieur, l'insula, le lobe temporal médial, le gyrus parahippocampique, le striatum et l'amygdale, cette augmentation était significativement corrélée à l'efficacité de la privation de sommeil, reflétée par une somnolence subjective accrue. Cette étude suggère qu'une augmentation de mGluR5 pourrait être un mécanisme neurobiologique expliquant l'efficacité antidépressive élevée de la privation de sommeil. Les études précliniques confirment notre association hypothétique entre le mGluR5, le sommeil et la dépression. En particulier, une étude chez des souris knock-out mGluR5 fournit des preuves importantes que le mGluR5 est impliqué dans le façonnement de la stabilité des transitions de l'état de sommeil NREM-sommeil paradoxal, de l'activité d'onde lente NREM et de la réponse homéostatique à la perte de sommeilAhnaou et al., 2015).
Pour résumer, il existe des preuves convergentes d'études animales, post mortem, MRS et PET indiquant que le système central du glutamate joue un rôle important dans la physiopathologie du TDM. Cependant, les preuves suggèrent que l'antagonisme de mGluR5 pourrait ne pas aider directement les patients atteints de TDM. Ceci est conforme à notre découverte d'une expression réduite de mGluR5 chez les patients atteints de TDM. On pourrait supposer que les médicaments qui ciblent le système mGluR5 pourraient être particulièrement utiles chez les patients déprimés souffrant d'anxiété comorbide, de troubles de la dépendance et / ou de troubles du rythme circadien. En outre, les dysfonctionnements dans les systèmes de processus sociaux sont généralement associés à la DMD. De faibles compétences sociales sont apparues comme un facteur de risque important de la dépression (Segrin, 2000). Ces déficits, y compris les comportements paralinguistiques et linguistiques, ainsi que les altérations de l’expression faciale, du regard, de la posture et des gestes, sont comparables à ceux observés dans le syndrome d’effet FX et le trouble du spectre autistique. En outre, en ce qui concerne les compétences sociales, des expériences sur des souris exposées à un stress social chronique ont montré que le couplage Homer1 / mGluR5 était interrompu, ce qui suggère que mGluR5 modère de nuit la vulnérabilité dépressive au stress social (Wagner et al., Sous presse). De plus, dans les modèles murins de déficits sociaux, la suppression de mGluR5 a conduit à la normalisation des interactions sociales (Chung et al., 2015). Ensemble, ces études fournissent des preuves précliniques que mGluR5 joue un rôle important dans les causes sociales de la dépression et les déficits sociaux fréquemment observés chez les patients déprimés. En conséquence, les médicaments ciblant mGluR5 peuvent jouer un rôle important dans la prévention de la dépression chez les jeunes présentant un déficit social et peuvent contribuer à traiter les déficits sociaux et le fonctionnement psychosocial faible des patients atteints de TDM.
mGluR5 et toxicomanie
La toxicomanie se caractérise par une consommation continue de drogues malgré des conséquences négatives, des tentatives répétées d’arrêter ou de réduire la consommation de drogues et des symptômes de tolérance et de sevrage. Bien que le système dopaminergique joue un rôle clé dans le traitement de la récompense aiguë (Kalivas et Volkow, 2005), de plus en plus de preuves impliquent une neurotransmission glutamatergique dans le traitement de la toxicomanie (Krystal et al., 2010).
Recherche neurobiologique préclinique
Dans 2001, une étude fondamentale sur mGluR5 et la dépendance a été publiée (Chiamulera et al., 2001). Les auteurs ont montré que des souris dépourvues du récepteur mGluR5 ne parvenaient pas à s'auto-administrer par voie intraveineuse de cocaïne malgré une augmentation des taux de dopamine extracellulaire dans le noyau accumbens après une injection aiguë. De nombreuses études de suivi chez l’animal ont montré que les antagonistes des récepteurs mGluR5, MPEP et MTEP, réduisent l’auto-administration de médicaments entraînant une dépendance tels que la cocaïne et la nicotine (Kalivas, 2009). Olive (2005) ont passé en revue des études sur des animaux liées à la toxicomanie et ont suggéré que mGluR5 pourrait être impliqué dans le développement, la perception des récompenses et la rechute de la consommation de drogues telles que la cocaïne, la morphine, la nicotine et l’éthanol. Les auteurs ont décrit des preuves d'études animales montrant que les antagonistes du mGluR5 réduisaient l'auto-administration du médicament, ainsi que le comportement de recherche du médicament. Par exemple, il a été découvert récemment, dans un modèle de trouble de la frénésie alimentaire chez le babouin, que le MTEP diminuait la consommation de bonbons sans modifier le comportement de recherche de bonbons (Bisaga et al., 2008). Les recherches ont suggéré que la diminution de l'apport alimentaire associée à l'utilisation d'antagonistes des récepteurs mGluR5 pourrait être liée à une réduction de la valeur enrichissante des stimuli de renforcement (Bisaga et al., 2008). De plus, des recherches ont montré une augmentation significative des niveaux d’ARNm de mGluR5 dans le noyau accumbens et le striatum dorsolatéral après administration répétée de cocaïne chez le rat de laboratoire (Bisaga et al., 2008).
Études humaines
En utilisant une autoradiographie dans des échantillons de tissu cérébral post-mortem provenant de patients souffrant de troubles alcooliques et de témoins en bonne santé, les recherches ont révélé une densité de liaison 30 – 40% supérieure mGluR1 / 5 dans l’hippocampe et le striatum chez les patients souffrant de troubles alcooliques (Kupila et al., 2012). Cette découverte suggère que la densité des récepteurs mGluR5 pourrait être augmentée dans certaines zones du cerveau de patients toxicomanes. Récemment, nous avons utilisé la TEP pour mesurer la liaison du récepteur mGluR5 dans le cerveau des personnes ayant une dépendance à la nicotine (Akkus et al., 2013). Nous avons constaté une réduction globale du mGluR5 DVR dans la substance grise des fumeurs de 14 par rapport aux non-fumeurs (voir la figure 2). Les réductions les plus importantes ont été observées dans les cortex orbitofrontal médial droit et gauche. Nous avons suggéré que cette diminution de la liaison aux récepteurs mGluR5 pourrait constituer une adaptation durable aux augmentations chroniques du glutamate induites par une administration chronique de nicotine. Cette adaptation semble être spécifique à la dépendance à la nicotine puisqu'elle n'a pas été trouvée chez les consommateurs chroniques de cocaïne non-fumeurs (Hulka et al., 2014).
FIGURE 2
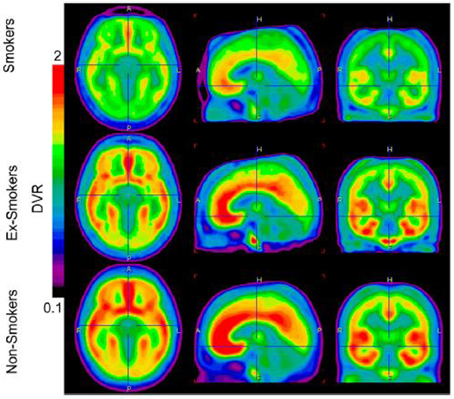
Figure 2. L'image montre l'absorption moyenne de mGluR5 DVR par le cerveau dans les trois groupes de diagnostic. L'absorption du cerveau est visible réduite chez les groupes de fumeurs et d'anciens fumeurs. Les images sont calculées avec la version du logiciel PMOD 3 (PMOD Technologies). De Akkus et al. (2013). Remarque: la figure et sa légende sont reproduites avec autorisation.
En effet, il a été suggéré que la régulation négative de mGluR5 représente une neuroadaptation compensatoire (Kalivas, 2009), un stimulant de la récompense induite par le médicament (Rutten et al., 2011), ou un facteur médiant les effets des signaux contextuels dans les réponses comportementales conditionnées (Tronci et al., 2010). Figure 3 présente une approche du dysfonctionnement mGluR5 dans la toxicomanie.
FIGURE 3
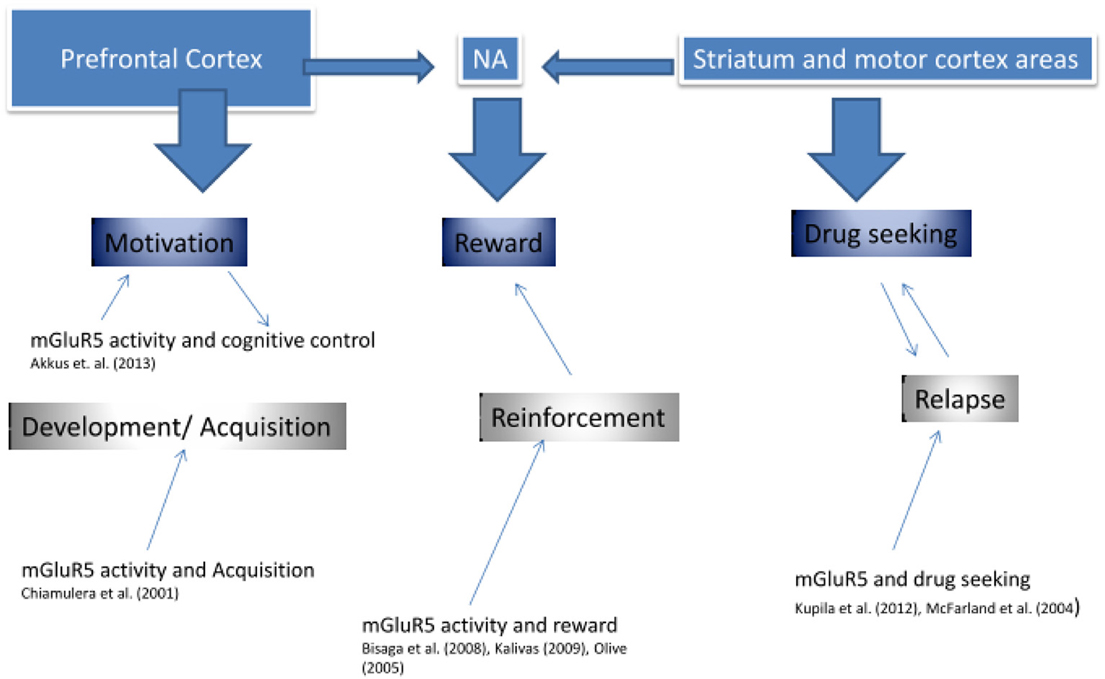
Figure 3. Implication de mGluR5 dans la dépendance.
Comme le montre la figure 3, Les recherches précédentes soutiennent l’idée que mGluR5 est impliqué dans trois phases clés de la dépendance, dans le développement et l’acquisition, dans la valeur de renforcement de la drogue, ainsi que dans la rechute de la dépendance. On pourrait suggérer que chaque fonction est principalement décrite par différentes régions du cerveau, qui présentent également une densité élevée de récepteurs mGluR5, et dont la densité réduite de mGluR5 a été démontrée dans notre recherche sur la TEP (Deschwanden et al., 2011). Kalivas (2009) développé un vaste modèle de dépendance au glutamate. L'auteur a suggéré que la dépendance était associée à un dysfonctionnement de l'homéostasie du glutamate entre des zones clés du circuit cérébral cortico-strié, y compris l'amygdale, le noyau accumbens (NA), le cortex préfrontal et le cortex moteur. Comme nous le suggérons dans la figure 3Premièrement («composante de motivation»), le cortex préfrontal pourrait être un moteur de la motivation et du contrôle cognitif au cours des phases initiales du développement de la toxicomanie. L’étude préclinique de Chiamulera et al. (2001) suggère que mGluR5 est nécessaire dans cette phase de développement de la dépendance en médiant les propriétés gratifiantes des drogues d'abus. Deuxièmement («composant de récompense»), il a été démontré que le Nucleus Accumbens (NA) influence la valeur de récompense du médicament, non seulement via la dopamine, mais également via l’activité mGluR5 (Bisaga et al., 2008). En particulier, une altération de la transmission du glutamate entre le cortex pré-limique et l'AN a été suggérée, de telle sorte que la recherche de drogue a été initiée par une plus grande confiance dans le comportement acquis associé au système de récepteurs NMDA / mGluR5 (Kalivas, 2009). Et troisièmement («composant de recherche de médicaments»), certaines recherches soutiennent que mGluR5 est impliqué dans une plus grande dépendance vis-à-vis des processus moteurs - réduisant le contrôle cognitif - via le striatum (Kupila et al., 2012). Nos propres recherches publiées et non publiées suggèrent que la régulation négative de mGluR5 est associée à un risque accru de rechute des anciens fumeurs (Akkus, PNAS). Cette régulation négative peut être une modification pathogène ou insuffisante. Dans l’ensemble, les recherches actuelles suggèrent que les médicaments ciblant mGluR5 pourraient améliorer les traitements des troubles de la toxicomanie à différents stades de leur développement.
a lieu
Les preuves examinées suggèrent une forte implication de mGluR5 dans les troubles anxieux, les TOC, le TDM, ainsi que la toxicomanie, et un traitement par des médicaments ciblant mGluR5 pourrait également être bénéfique chez l'homme. Cependant, la pathogenèse des symptômes de l'humeur et de l'anxiété associés au TOC peut différer des symptômes de l'humeur et de l'anxiété non liés au TOC. En conséquence, la présence de symptômes obsessionnels compulsifs peut être un facteur prédictif important de la réponse antidépressive et anxiolytique à des médicaments ciblant mGluR5. En outre, nous avons suggéré que les médicaments ciblant le système mGluR5 pourraient contribuer à accroître la résistance au stress social et à améliorer les déficits sociaux liés à la dépression. Étant donné que le stress social est de loin le principal facteur de risque non génétique de la dépression et que les déficits sociaux sont étroitement liés à la réduction du fonctionnement social et de la qualité de la vie, les résultats d'études animales revêtent une grande pertinence scientifique et clinique.
L'Institut national de la santé mentale (NIMH) a récemment lancé un projet novateur et à la pointe de la technologie: les critères du domaine de la recherche (RDoC). Cela reflète la mise en œuvre de la stratégie NIMH 1.4 «Développer, à des fins de recherche, de nouvelles méthodes de classification des troubles mentaux basées sur les dimensions du comportement observable et les mesures neurobiologiques.» (http://www.nimh.nih.gov/research-priorities/strategic-objectives/strategic-objective-1.shtml). Cette initiative est le résultat de la restructuration de catégories de DSM rigides, dont la plupart ont également été développées avant la recherche en neurosciences (Morris et Cuthbert, 2012). RDoC décrit cinq domaines ou constructions; Système de valence négative, Système de valence positive, Systèmes cognitifs, Systèmes pour processus sociaux, Systèmes d'éveil / régulation. Conformément aux preuves examinées sur l'implication de mGluR5 dans les MDD et les TOC, nous proposons que l'activité de mGluR5 soit directement associée au système de valence négative, qui implique les facteurs observables de peur, de menace, de menace persistante, de perte et de non-frustration frustrée. récompense. Ainsi, comme le suggèrent les études examinées, un traitement antagoniste avec mGluR5 devrait réduire ces symptômes observables. De plus, nous suggérons que le traitement par mGluR5 pourrait également être bénéfique dans les «troubles de valence positive» tels que la dépendance et la dépression, via une activité anormale de mGluR5 sur la structure et la fonction cérébrales liées au récepteur NMDA glutamatergique fonctionnellement lié à l'apprentissage par récompense . Par exemple, Simonyi et al. (2010) ont examiné de nombreuses études sur des animaux utilisant des antagonistes des récepteurs mGluR5 chez des souris knock-out pour déterminer le rôle de mGluR5 dans l'apprentissage et la mémoire. L’apprentissage inhibiteur, tel que l’apprentissage par évitement passif, est une tâche bien établie sur des modèles animaux et utilisée pour étudier les processus d’apprentissage de l’hippocampe. De nombreuses études ont montré qu’elle dépendait du récepteur mGluR5 (Simonyi et al., 2010). Par exemple, des recherches ont démontré une hyperexpression de la protéine mGluR5 dans CA3 au cours de la potentialisation à court et à long terme de CA1 chez le rat (Riedel et al., 2000). Hyman (2005) a présenté un modèle biologique de dépendance intégrant des processus neuronaux anormaux d'apprentissage et de mémoire constituant les éléments de base de la dépendance. Les auteurs ont proposé cette potentialisation à long terme, notamment les modifications de la disponibilité des récepteurs du glutamate et la régulation de l'expression des gènes en tant que mécanismes potentiellement importants pour les modifications induites par le médicament détectées dans les circuits anormaux associés à la toxicomanie. Enfin, les études sur mGluR5 et l’homéostasie du sommeil (Hefti et al., 2013; Ahnaou et al., 2015) suggèrent un rôle important du mGluR5 dans le domaine des systèmes d'excitation et de modulation de RDoC.
Figure 4 résume les processus proposés pour médier les actions de mGluR5 dans les troubles de l'humeur et la toxicomanie. Le haut de la figure 4 décrit les régions du cerveau mGluR5 à haute densité; amygdale, hippocampe, striatum, NA et cortex préfrontal. Les syndromes psychiatriques sont appariés à ces régions. L'activité de mGluR5, suggérée dans l'amygdale, pourrait être un médiateur de l'excitation émotionnelle primaire, telle que l'anxiété et la dépression. Nous avons décrit comment deux voies possibles; le fonctionnement social et l'apprentissage peuvent jouer un rôle médiateur dans les autres processus. Ainsi, l'activité de mGluR5 s'est avérée impliquée dans l'apprentissage et, par conséquent, via l'activité dans l'hippocampe, NA et le striatum pourrait être impliquée dans la mémoire et la récompense, le contrôle cognitif et la motivation, ainsi que dans l'addiction (voir aussi la figure). 3). Enfin, nous avons suggéré comment mGluR5 pourrait être lié à la réponse au stress social et aux déficits sociaux, qui pourraient être pertinents pour un large éventail de conditions psychiatriques. Table 2 donne un aperçu de la zone de dégradation RDoC, du tableau clinique associé et de la participation de mGluR5.
FIGURE 4
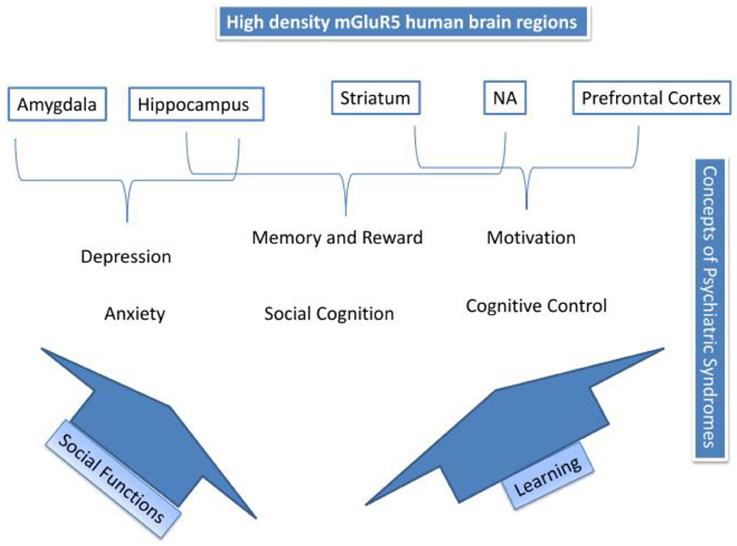
Figure 4. Résumé des mécanismes de mGluR5.
TABLEAU 2
![]()
Tableau 2. RDoC et mGluR5.
Remarques finales
Cette revue a décrit l'implication de mGluR5 dans les troubles de l'humeur, les TOC et la dépendance, et a comparé la recherche préclinique à la recherche humaine, en particulier la recherche PET. Nous avons donc comparé différentes approches méthodologiques, telles que la recherche sur les animaux, la MRS et les études PET. Nous suggérons qu'un puissant effet anxiolytique direct serait présent si le traitement antagoniste mGluR5 était initié lors d'essais cliniques sur l'homme (RDoC). Une suractivité de mGluR5 a également été rapportée dans FXS, caractérisé par des déficits sociaux importants. En conséquence, l'activité de mGluR5 peut non seulement normaliser l'activité dans les systèmes de valence négative et les systèmes d'éveil, mais également atténuer les déficiences des systèmes de processus sociaux (RDoC). Cela revêt une grande importance clinique, car un fonctionnement social médiocre est un résultat important des maladies psychiatriques courantes telles que les TOC, la dépression et la toxicomanie, entraînant d’énormes souffrances personnelles et des coûts indirects importants pour la société. Enfin, il a également été démontré que mGluR5 avait une implication significative dans la toxicomanie, ce qui pourrait être causé principalement par l’augmentation de la valeur de récompense de la drogue. Une intervention antagoniste de mGluR5 serait donc plus efficace pour le traitement de l’anxiété pathologique et de la toxicomanie, ainsi que pour améliorer la résilience au stress social et le fonctionnement social.
Déclaration de conflit d'intérêts
Terbeck, Chesterman et Akkus n’ont pas de conflit d’intérêts. Le Dr Hasler a reçu une subvention de Novartis, qui produit et teste des médicaments ciblant mGluR5.
Bibliographie
Abe, T., H. Sugihara, H. Nawa, R. Shigemoto, N. Mizuno et S. Nakanishi (1992). Caractérisation moléculaire d'un nouveau récepteur métabotropique du glutamate mGluR5 couplé à la transduction du signal inositol phosphate / Ca2 +. J. Biol. Chem. 267, 13361 – 13368.
Résumé PubMed | Texte intégral | Google Scholar
Agid, Y., Buzsáki, G., Diamond, DM, Frackowiak, R., Giedd, J., Girault, J.-A., et al. (2007). Comment améliorer la découverte de médicaments pour les troubles psychiatriques? Nat. Rev. Drug Discov. 6, 189 – 201. doi: 10.1038 / nrd2217
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Akkus, F., Ametamey, SM, Treyer, V., Burger, C., A. Johayem, A., Umbricht, D., et al. (2013). Réduction globale marquée de la liaison du récepteur mGluR5 chez les fumeurs et les ex-fumeurs déterminée par tomographie par émission de positrons [11C] ABP688. Proc. Natl. Acad. Sci. Etats-Unis. 110, 737 – 742. doi: 10.1073 / pnas.1210984110
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Akkus, F., Terbeck, S., Ametamey, SM, Rufer, MD, Treyer, V., Burger, C. et al. (2014). Liaison du récepteur métabotropique du glutamate à 5 chez les patients présentant un trouble obsessionnel-compulsif. Int. J. Neuropsychopharmacol. 17, 1915 – 1922. doi: 10.1017 / S1461145714000716
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Ametamey, SM, Kessler, LJ, Honer, M., Wyss, MT, Buck, A., Hintermann, S., et al. (2006). La radiosynthèse et l'évaluation préclinique de 11CABP688 en tant que sonde d'imagerie du sous-type de récepteur métabotropique du glutamate, 5. J. Nucl. Med. 47, 698 – 705.
Résumé PubMed | Texte intégral | Google Scholar
Anderson, JJ, Bradbury, MJ, Giracello, DR, Chapman, DF, Holtz, G., Roppe, J., et al. (2003). in vivo occupation du récepteur des antagonistes du récepteur mGlu5 par le nouveau radioligand [3 H] 3-méthoxy-5- (pyridine-2-yléthynyl) pyridine). EUR. J. Pharmacol. 473, 35–40. doi: 10.1016/S0014-2999(03)01935-6
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Ahnaou, A., Raeymaekers, L., Steckler, T. et Drinkenbrug, WHIM (2015). Pertinence du récepteur métabotropique du glutamate (mGluR5) dans la régulation du cycle de sommeil NREM-REM et de l'homéostasie: mise en évidence chez la souris mGluR5 (- / -). Comportement Brain Res. 282, 218 – 226. doi: 10.1016 / j.bbr.2015.01.009
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Bear, MF, Huber, KM et Warren, ST (2004). La théorie mGluR du retard mental X fragile. Tendances Neurosci. 27, 370 – 377. doi: 10.1016 / j.tins.2004.04.009
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
D. Berg, J. Godau, C. Trenkwalder, K. Eggert, I. Csoti, A. Storch, et al. (2011). Traitement AFQ056 des dyskinésies induites par la lévodopa: résultats des essais comparatifs randomisés sur 2. Mov. Désordre. 26, 1243 – 1250. doi: 10.1002 / mds.23616
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Berry-Kravis, E., D. Hessl, S. Coffey, C. Hervey, A. Schneider, J. Yuhas et autres. (2009). Essai pilote à dose unique en ouvert sur le fenobam chez des adultes atteints du syndrome de l’X fragile. J. Med. Genet. 46, 266 – 271. doi: 10.1136 / jmg.2008.063701
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Bisaga, A., Danysz, W. et Foltin, RW (2008). L'antagonisme des récepteurs glutamatergiques NMDA et mGluR5 diminue la consommation d'aliments dans le modèle babouin du syndrome de la frénésie alimentaire. EUR. Neuropsychopharmacol. 18, 794 – 802. doi: 10.1016 / j.euroneuro.2008.05.004
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Boer K., Encha-Razavi, F., Sinico, M. et Aronica, E. (2010). Distribution différentielle des récepteurs métabotropes du glutamate du groupe I dans le cortex humain en développement. Brain Res. 1324, 24 – 33. doi: 10.1016 / j.brainres.2010.02.005
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Brennan, BP, Rauch, SL, Jensen, JE et Pope, HG (2012). Une revue critique des études de spectroscopie par résonance magnétique du trouble obsessionnel-compulsif. Biol. Psychiatrie 73, 31 – 24. doi: 10.1016 / j.biopsych.2012.06.023
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Burket, JA, Benson, AD, Tang, AH et Deutsch, SI (2014). La rapamycine améliore la sociabilité dans le modèle murin BTBR de troubles du spectre autistique. Brain Res. Taureau. 100, 70 – 75. doi: 10.1016 / j.brainresbull.2013.11.005
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Chiamulera, C., MP Epping-Jordan, A. Zocchi, C. Marcon, C. Cottiny, S. Tacconi et al. (2001). Les effets stimulants et stimulants locomoteurs de la cocaïne sont absents chez les souris mutantes nulles mGluR5. Nat. Neurosci. 4, 873 – 874. Doi: 10.1038 / nn0901-873
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
W. Chung, SY Choi, E. Lee, H. Park, H. J. Kang., H. Park et al. (2015). Les déficits sociaux chez les souris mutantes IRSp53 sont améliorés par la suppression de NMDAR et de mGluR5. Nat. Neurosci. 18, 435 – 442. doi: 10.1038 / nn.3927
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Cleva, RM et Olive, MF (2011). Modulateurs allostériques positifs des récepteurs métabotropiques du glutamate de type 5 (mGluR5) et leur potentiel thérapeutique pour le traitement des troubles du système nerveux central. Molécules 16, 2097 – 2106. doi: 10.3390 / molécules16032097
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Deschwanden, A., Karolewicz, B., Feyissa, AM, Treyer, V., Ametamey, SM, Johayem, A. et al. (2011). Réduction de la densité 5 du récepteur métabotropique du glutamate dans la dépression majeure déterminée par [(11) C] ABP688 PET et une étude post mortem. Un m. J. Psychiatrie 168, 727 – 734. doi: 10.1176 / appi.ajp.2011.09111607
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Erickson, CA, Mullett, JE et McDougle, CJ (2009). La mémantine en ouvert dans le syndrome de l'X fragile. J. Autism Dev. Désordre. 39, 1629 – 1635. Paramètres: 10.1007 / s10803-009-0807-3
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Erickson, CA, Early, M., Stigler, KA, Wink, LK, Mullett, JE et McDougle, CJ (2011). Une étude pilote naturaliste ouverte sur l'acamprosate chez les jeunes atteints de trouble autistique. J. Enfant Adolesc. Psychopharmacol. 21, 565 – 569. doi: 10.1089 / cap.2011.0034
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Premièrement, MB et Gibbon, M. (1997). Guide de l'utilisateur pour l'entrevue clinique structurée pour les troubles de l'axe I du DSM-IV SCID-I: version clinicien. New York, NY: American Psychiatric Pub.
Feyissa, AM, Chandran, A., Stockmeier, CA et Karolewicz, B. (2009). Réduction des niveaux de sous-unités NR2A et NR2B du récepteur NMDA et de PSD-95 dans le cortex préfrontal dans la dépression majeure. Programme. Neuropsychopharmacol. Biol. Psychiatrie 33, 70 – 75. doi: 10.1016 / j.pnpbp.2008.10.005
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Garber, KB, J. Visootsak et Warren, ST (2008). Syndrome de l'X fragile. EUR. J. Hum. Genet. 16, 666 – 672. doi: 10.1038 / ejhg.2008.61
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Gass, JT, Osborne, député, Watson, NL, Brown, JL et Olive, MF (2009). L'antagonisme de mGluR5 atténue le renforcement de la méthamphétamine et empêche le rétablissement du comportement de recherche de méthamphétamine chez le rat. Neuropsychopharmacology. 34, 820 – 833. doi: 10.1038 / npp.2008.140
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Harvey, BH et Shahid, M. (2012). Les récepteurs métabotropes et ionotropes du glutamate en tant que cibles neurobiologiques dans les troubles anxieux et liés au stress: focus sur la pharmacologie et les modèles de traduction précliniques. Pharmacol. Biochem. Comportement. 100, 775 – 800. doi: 10.1016 / j.pbb.2011.06.014
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Hasler, G., LaSalle-Ricci, VH, Ronquillo, JG, Crawley, SA, Cochran, LW, Kazuba, D. et al. (2005). Les dimensions des symptômes du trouble obsessionnel-compulsif montrent des relations spécifiques avec la comorbidité psychiatrique. Res psychiatrie. 135, 121 – 132. doi: 10.1016 / j.psychres.2005.03.003
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Hasler, G., JW van der Veen, T. Tumonis, N. Meyers, J. Shen et Drevets, WC (2007). Réduction des taux de glutamate / glutamine préfrontal et d'acide γ-aminobutyrique dans la dépression majeure, déterminée par spectroscopie par résonance magnétique proton. Cambre. Psychiatrie générale 64, 193 – 200. doi: 10.1001 / archpsyc.64.2.193
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Hasler, G., van der Veen, JW, Geraci, M., Shen, J., D., Pine et Drevets, WC (2009). Niveaux d'acide gamma-aminobutyrique corticaux préfrontal dans le trouble panique, déterminés par spectroscopie à résonance magnétique du proton. Biol. Psychiatrie 65, 273 – 275. doi: 10.1016 / j.biopsych.2008.06.023
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Hefti, K., Holst, SC, J. Sovago, V. Bachmann, A. Buck, A. Ametamey, SM et al. (2013). Augmentation de la disponibilité du sous-type de 5 du récepteur métabotropique du glutamate dans le cerveau humain après une nuit sans sommeil. Biol. Psychiatrie 73, 161 – 168. doi: 10.1016 / j.biopsych.2012.07.030
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Hyman, SE (2005). Dépendance: une maladie de l'apprentissage et de la mémoire. Un m. J. Psychiatrie 162, 1414 – 1422. doi: 10.1176 / appi.ajp.162.8.1414
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Hoffpauir, BK et Gleason, EL (2002). L'activation de mGluR5 module la fonction du récepteur GABAA dans les cellules amacrines rétiniennes. J. Neurophysiol. 88, 1766 – 1776.
Résumé PubMed | Texte intégral | Google Scholar
Hulka, LM, Treyer, V., Scheidegger, Preller, KH, Vonmoos, M., Baumgartner, MR, et al. (2014). Le tabagisme, mais pas la consommation de cocaïne, est associé à la densité 5 du récepteur métabotropique du glutamate cérébral inférieur chez l'homme. Mol. Psychiatrie 19, 625 – 632. doi: 10.1038 / mp.2013.51
CrossRef Full Text | Google Scholar
Kalivas, PW et Volkow, ND (2005). La base neurale de la dépendance: une pathologie de la motivation et du choix. Un m. J. Psychiatrie 162, 1403 – 1413. doi: 10.1176 / appi.ajp.162.8.1403
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Kalivas, PW (2009). L'hypothèse d'homéostasie du glutamate de la dépendance. Nat. Rev. Neurosci. 10, 561 – 572. doi: 10.1038 / nrn2515
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Krystal, JH, Mathew, SJ, D'Souza, DC, Garakani, A., Gunduz-Bruce, H., et Charney, DS (2010). Applications psychiatriques potentielles des agonistes et antagonistes des récepteurs métabotropiques du glutamate. Drogues du SNC 24, 669–693. doi: 10.2165/11533230-000000000-00000
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Kupila, J., Kärkkäinen, O., Laukkanen, V., Tupala, E., Tiihonen, J. et Storvik, M. (2012). Densité des récepteurs mGluR1 / 5 dans le cerveau de sujets alcooliques: étude par autoradiographie sur tout l'hémisphère. Res psychiatrie. 12, 149 – 155. doi: 10.1016 / j.pscychresns.2012.04.003
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Koros, E., H. Rosenbrock, G. Birk, C. Weiss et Sams-Dodd, F. (2007). Le MTEP, un antagoniste sélectif du récepteur mGlu5, similaire aux antagonistes du récepteur NMDA, induit l'isolement social chez le rat. Neuropsychopharmacology 32, 562 – 576. doi: 10.1038 / sj.npp.1301133
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Lea, PM et Faden, AI (2006). Les récepteurs métabotropiques du glutamate de sous-type 5 antagonistes MPEP et MTEP. CNS Drug Rev. 12, 149 – 166. Doi: 10.1111 / j.1527-3458.2006.00149.x
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Legutko, B., Szewczyk, B., L. Pomierny-Chamioło, G., Nowak et A. Pilc (2006). Effet du traitement MPEP sur l'expression des gènes du facteur neurotrophique dérivé du cerveau. Pharmacol. Représentant. 58, 427 – 430.
Résumé PubMed | Texte intégral | Google Scholar
Luykx, JJ, Laban, KG, van den Heuvel, député, Boks, MPM, Mandl, RCW, Kahn, RS, et al. (2012). Régulation à la baisse du glutamate spécifique à une région et à un état dans le trouble dépressif majeur: méta-analyse des résultats de (1) H-MRS. Neurosci. Biobehav. Tour. 36, 198 – 205. doi: 10.1016 / j.neubiorev.2011.05.014
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Meldrum, BS (2000). Le glutamate en tant que neurotransmetteur dans le cerveau: examen de la physiologie et de la pathologie. J. Nutr. 130, 1007.
Résumé PubMed | Texte intégral | Google Scholar
Morris, SE et Cuthbert, BN (2012). Critères du domaine de recherche: systèmes cognitifs, circuits neuronaux et dimensions du comportement. Dialogues Clin. Neurosci. 14, 29 – 37.
Résumé PubMed | Texte intégral | Google Scholar
Olive, MF (2005). Récepteurs mGlu5: neuroanatomie, pharmacologie et rôle dans la toxicomanie. Curr. Psychiatrie Rev. 1, 197 – 214. doi: 10.2174 / 1573400054065578
CrossRef Full Text | Google Scholar
Phan, KL, Fitzgerald, DA, Cortese, BM, Seraji-Bozorgzad, N., Tancer, ME et Moore, GJ (2005). Neurochimie du cingulum antérieur dans le trouble d'anxiété sociale: 1H-MRS chez 4 Tesla. Neuroreport 16, 183–186. doi: 10.1097/00001756-200502080-00024
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Pittenger, C., Krystal, JH et Coric, V. (2006). Les médicaments modulant le glutamate en tant que nouveaux agents pharmacothérapeutiques dans le traitement du trouble obsessionnel-compulsif. NeuroRx 3, 69 – 81. doi: 10.1016 / j.nurx.2005.12.006
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Pollack, MH, Jensen, JE, Simon, NM, Kaufmann, RE et Renshaw, PF (2008). Etude MRS à haut champ sur le GABA, le glutamate et la glutamine dans le trouble d'anxiété sociale?: Réponse au traitement par le lévétiracétam. Programme. Neuropsychopharmacol. Biol. Psychiatrie 32, 739 – 743. doi: 10.1016 / j.pnpbp.2007.11.023
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Riaza Bermudo-Soriano, C., Perez-Rodriguez, MM, Vaquero-Lorenzo, C. et Baca-Garcia, E. (2012). Nouvelles perspectives dans le glutamate et l'anxiété. Pharmacol. Biochem. Comportement. 100, 752 – 774. doi: 10.1016 / j.pbb.2011.04.010
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Riedel, G., Casabona, G., Platt, B., Macphail, EM et Nicoletti, F. (2000). Augmentation de l'expression de la protéine du récepteur mGlu5 dans l'hippocampe de rat en fonction du temps et du sous-région, provoquée par le conditionnement. Neuropharmacologie 39, 1943–1951. doi: 10.1016/S0028-3908(00)00037-X
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Rutten, K., De Vry, J., Bruckmann, W. et Tzschentke, TM (2011). Le blocage pharmacologique ou l'inactivation génétique du récepteur NOP potentialise l'effet de récompense de la morphine chez le rat. La drogue dépend de l'alcool. 114, 253 – 256. doi: 10.1016 / j.drugalcdep.2010.10.004
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Romano, C., van den Pol, AN et O'Malley, KL (1996). Expression développementale précoce améliorée du récepteur métabotropique du glutamate mGluR5 dans le cerveau de rat: protéines, variantes d'épissage d'ARNm et distribution régionale. J. Compar. Neurol. 367, 403 – 412.
Rosenberg, DR et Keshavan, MS (1998). Vers un modèle neurodéveloppemental du trouble obsessionnel-compulsif. Biol. Psychiatrie 43, 623–640. doi: 10.1016/S0006-3223(97)00443-5
CrossRef Full Text | Google Scholar
Sanacora, G., Zarate, CA, Krystal, JH et Manji, HK (2008). Cibler le système glutamatergique pour développer de nouvelles thérapies améliorées pour les troubles de l'humeur. Nat. Rev. Drug Discov. 7, 426 – 437. doi: 10.1038 / nrd2462
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Schoepp, DD (2001). Dévoiler les fonctions des récepteurs présynaptiques du glutamate métabotropique dans le système nerveux central. J. Pharmacol. Exp. Ther. 299, 12 – 20.
Résumé PubMed | Texte intégral | Google Scholar
Segrin, C. (2000). Déficits en compétences sociales associés à la dépression. Clin. Psychol. Tour. 20, 379–403. doi: 10.1016/S0272-7358(98)00104-4
CrossRef Full Text | Google Scholar
Simonyi, A., Schachtman, TR et Christoffersen, GR (2010). Récepteur métabotropique du glutamate, sous-type 5, antagoniste de l'apprentissage et de la mémoire. EUR. J. Pharmacol. 639, 17 – 25. doi: 10.1016 / j.ejphar.2009.12.039
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Soares, DP et Law, M. (2009). Spectroscopie par résonance magnétique du cerveau: examen des métabolites et applications cliniques. Clin. Radiol. 64, 12 – 21. doi: 10.1016 / j.crad.2008.07.002
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Le juge en chef Swanson, MM. Bures, Johnson, MP, Linden, A.-M., Monn, JA et Schoepp, DD (2005). Les récepteurs métabotropiques du glutamate en tant que nouvelles cibles pour les troubles anxieux et de stress. Nat. Rev. Drug Discov. 4, 131 – 144. doi: 10.1038 / nrd1630
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Szeszko, PR, Christian, C., Macmaster, F., Lencz, T., Mirza, Y., Taormina, SP, et al. (2008). Altérations structurelles de la matière grise dans le trouble obsessionnel-compulsif pédiatrique naïve à la drogue psychotrope: étude optimisée de la morphométrie à base de voxel. Un m. J. Psychiatrie 165, 1299 – 1307. doi: 10.1176 / appi.ajp.2008.08010033
CrossRef Full Text | Google Scholar
E. Tatarczyńska, A. Klodzińska, E. E. Chojnacka-Wójcik, A. Palucha, F. Gasparini, R. Kuhn et coll. (2001). Les effets potentiels anxiolytiques et antidépresseurs de MPEP, un antagoniste puissant, sélectif et systémique du récepteur mGlu5. Le fr. J. Pharmacol. 132, 1423 – 1430. doi: 10.1038 / sj.bjp.0703923
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Tronci, V., S. Vronskaya, N. Montgomery, D. Mura et Balfour, DJK (2010). Les effets de l'antagoniste du récepteur mGluR5 6-méthyl-2- (phényléthynyl) -pyridine (MPEP) sur les réponses comportementales à la nicotine. Psychopharmacologie 211, 33–42. doi: 10.1007/s00213-010-1868-x
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Van den Heuvel, OA, Remijnse, PL, D. Mataix-Cols, H. Vrenken, HJ Groenewegen, Uylings, HBM, et al. (2009). Les principaux symptômes du trouble obsessionnel-compulsif sont véhiculés par des systèmes neuronaux partiellement distincts. Cerveau 132, 853 – 868. doi: 10.1093 / brain / awn267
CrossRef Full Text | Google Scholar
Wagner, KV, J. Hartmann, C. Labermaier, AS Häusl, G. Zhao, D. Harbich et al. (dans la presse). L'activité Homer1 / mGluR5 modère la vulnérabilité au stress social chronique. Neuropsychopharmacology. doi: 10.1038 / npp.2014.308
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Yüksel, C. et Öngür, D. (2010). Études par spectroscopie à résonance magnétique des anomalies liées au glutamate dans les troubles de l'humeur. Biol. Psychiatrie 68, 785 – 794. doi: 10.1016 / j.biopsych.2010.06.016
CrossRef Full Text | Google Scholar
Zarei, M., D. Mataix-Cols, I. Heyman, M., Hough, J., Doherty, L. Burge, et al. (2011). Modifications du volume de la matière grise et de la microstructure de la substance blanche chez les adolescents présentant un trouble obsessionnel-compulsif. Biol. Psychiatrie 70, 1083 – 1090. doi: 10.1016 / j.biopsych.2011.06.032
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Mots-clés: mGluR5, PET, troubles de l'humeur, dépendance, anxiété
Citation: Terbeck S, Akkus F, Chesterman LP et Hasler G (2015) Le rôle du récepteur métabotropique du glutamate 5 dans la pathogenèse des troubles de l'humeur et de la toxicomanie: combinaison de preuves précliniques et d'études de tomographie par émission de positrons (PET) humaine. De face. Neurosci. 9: 86. doi: 10.3389 / fnins.2015.00086
Reçu: 07 Janvier 2015; Accepté: 27 février 2015;
Publié: 18 March 2015.
Édité par:
Ashok Kumar, Université de Floride, États-Unis
Commenté par:
Karthik Bodhinathan, Institut de recherche médicale Sanford Burnham, États-Unis
Amber M. Muehlmann, Université de Floride, États-Unis
Droits d'auteur © 2015 Terbeck, Akkus, Chesterman et Hasler. Ceci est un article en accès libre distribué selon les termes de la Licence d'attribution Creative Commons (CC BY). L'utilisation, la distribution ou la reproduction sur d'autres forums est autorisée, à condition que l'auteur original ou le donneur de licence soit crédité et que la publication originale de ce journal soit citée conformément à la pratique académique reconnue. Aucune utilisation, distribution ou reproduction n’est autorisée si elle n’est pas conforme à ces conditions.
* Correspondance: Sylvia Terbeck, École de psychologie, Faculté des sciences de la santé et des sciences humaines, Université de Plymouth, PL4 8AA, Plymouth, Royaume-Uni. [email protected]
Troubles psychiatriques et intervention à base de glutamate chez l'homme
Les troubles psychiatriques sont très hétérogènes et la comorbidité est fréquente. Cependant, les traitements pharmacologiques psychiatriques courants reposent sur relativement peu de mécanismes physiopathologiques, par exemple l'augmentation de la disponibilité de la monoamine dans l'anxiété et la dépression. Il est donc urgent d'améliorer et de faire progresser les traitements psychiatriques, et une intervention pharmacologique à base de glutamate métabotropique constitue un développement prometteur à cet égard (Agid et al., 2007).
La plupart des essais humains ont été menés sur des cas de syndrome du x fragile (FXS). La théorie de FXS basée sur mGluR5 postule que l’absence de protéine de retard mental X fragile (FMRP) entraîne une signalisation glutamatergique excessive via mGluR5 (Bear et al., 2004). Cela conduit à une traduction accrue de l'ARNm local à la synapse, car FMRP n'est pas présent pour réguler le processus. Enfin, cela affaiblit la synapse et entraîne une augmentation du nombre d'épines dendritiques immatures plus longues, ce qui pourrait expliquer la déficience intellectuelle constatée chez les patients atteints de FXS. Cette déficience est associée à des symptômes d'humeur et d'anxiété et présente généralement des caractéristiques communes aux troubles du spectre autistique, notamment des retards dans le développement de la parole et du langage, une altération de la théorie de l'esprit, une altération du traitement social et émotionnel, ainsi qu'un comportement répétitif (Garber et al., 2008). Les preuves préliminaires et indirectes que l'antagoniste de mGluR5 peut améliorer la sociabilité dans FXS (Burket et al., 2014) suscite l’espoir que les médicaments ciblant mGluR5 pourraient être utilisés en clinique dans les affections psychiatriques prévalentes associées à des troubles du système pour des processus sociaux tels que l’autisme, la schizophrénie et la dépression. De plus, le phénotype de comportement répétitif observable dans FXS pourrait suggérer une pathophysiologie partagée parmi d'autres troubles psychiatriques tels que le trouble obsessionnel-compulsif (TOC) et la dépendance.
Bien que les connaissances scientifiques et les recherches sur mGluR5 se soient accrues, les efforts de développement de médicaments ont été relativement infructueux (Agid et al., 2007). Les médicaments qui ciblent les récepteurs ionotropes produisent généralement de nombreux effets secondaires et les stratégies actuelles de développement de médicaments n’ont pas encore produit de cibles sélectives pour les récepteurs ionotropes susceptibles de réduire les effets secondaires potentiels. Par exemple, les antagonistes des récepteurs ionotropes produisent des effets secondaires chez l’homme, notamment des troubles de la mémoire, des épisodes psychotiques et des accidents vasculaires cérébraux (Swanson et al., 2005). Les effets indésirables peuvent être dus au fait que les récepteurs ionotropes du glutamate ont une distribution omniprésente, alors que les récepteurs métabotropes sont plus inégaux et sélectivement distribués (Krystal et al., 2010). En conséquence, le développement récent de médicaments s'est concentré sur des composés ciblant les récepteurs métabotropes, en supposant que ces médicaments seraient associés à moins d'effets secondaires que ceux qui se lient aux récepteurs ionotropes à action rapide.
Dans les sections suivantes, nous examinerons les résultats d'études sur la TEP chez l'homme concernant l'implication de mGluR5 dans les troubles de l'humeur et la toxicomanie et comparerons ces résultats à ceux d'études animales. De plus, nous décrirons certains emplacements du cerveau pour l'activité de mGluR5 chez l'homme et suggérerons enfin une approche de la manière dont l'activité mGluR5 directe et indirecte pourrait être impliquée dans les syndromes psychiatriques humains.
mGluR5, anxiété pathologique et troubles de l'humeur
Anxiété Pathologique
L’anxiété pathologique survient dans les troubles anxieux, y compris le trouble anxieux généralisé, le trouble panique (les troubles psychiatriques les plus répandus dans le monde, First et Gibbon, 1997), mais aussi dans d’autres affections psychiatriques prévalentes telles que la dépression et le trouble obsessionnel-compulsif (TOC). Ces troubles psychiatriques entraînent une altération importante du fonctionnement social et professionnel, entraînant des coûts pour la santé et des souffrances pour les patients (First et Gibbon, 1997). En règle générale, l’anxiété peut être associée à une excitabilité cérébrale excessive (Harvey et Shahid, 2012).
Recherche neurobiologique préclinique
Les résultats d'un grand nombre d'essais précliniques chez l'animal ont permis de déterminer l'effet du traitement par antagoniste de mGluR5 sur l'anxiété. Swanson et al. (2005) ont examiné des études chez l'animal portant sur des médicaments ciblant le mGluR5 sur les comportements anxieux. Ils ont conclu que le traitement antagoniste de mGluR5 entraînait principalement des réponses anxiolytiques chez des animaux de laboratoire. Des effets tels que la diminution de la peur conditionnée au gel, une acceptation accrue des chocs et des punitions et une augmentation des interactions sociales ont été observés. Par exemple, une dose unique de 2-méthyl-6- (phényléthynyl) pyridine (MPEP) a prolongé le temps passé par les rats dans le bras ouvert d'un labyrinthe expérimental, sans affecter le comportement moteur ni la planification (Tatarczyńska et al., 2001). Krystal et al. (2010) ont examiné des études précliniques sur des animaux portant sur des antagonistes du mGluR5 (MTEP, MPEP, fenobam) dans des modèles murins d'anxiété. Ces études ont utilisé différentes mesures de résultats, telles que l’extinction du conditionnement de la peur et des réponses dans le labyrinthe en hauteur, pour évaluer l’efficacité des traitements médicamenteux. Parmi les études examinées, 88.45% a signalé un effet anxiolytique avec les antagonistes de mGluR5 (Krystal et al., 2010). Plus récemment, une autre revue de recherche sur l'anxiété chez des modèles animaux, examinant l'effet d'une intervention antagoniste ionotrope et métabotropique du récepteur du glutamate, a été publiée (Riaza Bermudo-Soriano et al., 2012). En ce qui concerne mGluR5, les auteurs ont répertorié les études 43 sur l'anxiété chez l'animal et toutes sauf deux ont démontré des effets anxiolytiques.
Études humaines
Les premières preuves de l'hypothèse selon laquelle la fonction du glutamate est anormale dans les troubles anxieux sont issues de la recherche sur le SRM. Par exemple, en utilisant la spectroscopie de résonance magnétique 1H à haut champ et à un seul voxel, les chercheurs ont constaté que, par rapport aux témoins sains, les patients présentant un trouble d'anxiété sociale présentaient des taux de glutamate significativement plus élevés dans le cortex cingulaire antérieur (ACC) (Phan et al., 2005). De plus, des recherches ont révélé une augmentation de la concentration globale de glutamate chez les patients 10 atteints de phobie sociale (Pollack et al., 2008). Comme indiqué précédemment, ces études n'ont toutefois pas permis de déterminer quels récepteurs du glutamate étaient associés à une activité excessive du glutamate.
En utilisant la méthodologie de recherche PET, nous avons récemment été le premier à montrer des relations entre mGluR5 et l’anxiété, dans le Trouble dépressif majeur (MDD), et le TOC. Dans une étude, nous avons étudié le rapport de volume de distribution (DVR) mGluR5 chez des patients 10 présentant un contrôle sain de TOC et de 10 à l’aide de [11C] ABP688 PET (Akkus et al., 2014). Nous avons utilisé l'échelle obsessionnelle-compulsive de Yale-Brown (Y-BOCS) comme mesure clinique de la gravité des symptômes du TOC. Nous avons observé des corrélations positives significatives entre les scores d’obsession Y-BOCS et le mGluR5 DVR dans les régions de l’amygdale, de l’ACC et du cortex orbitofrontal médial (Akkus et al., 2014). Ces zones du cerveau ont déjà été impliquées dans la physiopathologie des TOC. En effet, les anomalies cérébrales structurelles de l'amygdale, de l'ACC et du cortex orbitofrontal ont été associées de manière constante au TOC (Rosenberg et Keshavan, 1998; Szeszko et al., 2008; Van den Heuvel et al., 2009). Étant donné que les études d'imagerie structurelle ont fourni des preuves d'une corrélation positive entre la gravité des TOC et le volume de matière grise (Zarei et al., 2011), une augmentation de la liaison de mGluR5 dans les TOC peut refléter une augmentation de la densité des neurones. Bien que, dans le DSM-5, le TOC ne soit plus considéré comme un trouble anxieux, la plupart des patients atteints de TOC présentent des symptômes d’anxiété. Dans un échantillon clinique relativement important, nous avons précédemment montré que les patients souffrant de TOC obsédés avaient une prévalence particulièrement élevée de symptômes et de troubles anxieux (Hasler et al., 2005). Dans cet article, nous avons suggéré que les obsessions impliquant le stress, l’anxiété ou les conflits pourraient donc être associées à une neurotransmission glutamatergique accrue dans l’amygdale, le CAC et le cortex orbitofrontal.
Pris ensemble, les résultats d'études récentes utilisant différentes méthodologies de recherche confirment l'hypothèse selon laquelle la fonction du glutamate serait anormale dans des zones clés du système limbique dans les troubles de l'humeur liés à l'anxiété. La fonction anormale est également susceptible d'être liée au récepteur mGluR5. Étant donné que la recherche pré-clinique et celle sur la TEP ont montré des résultats cohérents, nous suggérons que le traitement antagoniste par mGluR5 produise des effets anxiolytiques significatifs chez les patients souffrant d’anxiété pathologique.
Trouble dépressif majeur (TDM)
Recherche neurobiologique préclinique
Dans leur avis, Krystal et al. (2010) a décrit les résultats de huit études portant sur les antagonistes du mTEP et du MPEP, antagonistes de mGluR5, dans des modèles animaux de dépression. Par rapport aux taux de succès du traitement des troubles anxieux, les auteurs ont indiqué que seul 62.5 – 75% avait trouvé un effet antidépresseur net, même si le traitement avec la kétamine, un antagoniste ionotrope de NMDA, avait entraîné des effets antidépresseurs rapides (même chez les patients résistants).Pittenger et al., 2006).
Études humaines
Une revue récente a examiné les études 13 MRS sur les troubles de l'humeur. Les auteurs ont rapporté que ces études ont systématiquement mis en évidence la réduction du glutamate dans le TDM (Hasler et al., 2007; Yüksel et Öngür, 2010). En particulier, des taux réduits de glutamate dans l'ACC, le cortex préfrontal dorsolatéral gauche, le cortex préfrontal dorsomédial, le cortex préfrontal ventromédial, l'amygdale et l'hippocampe ont été mis en évidence. Un examen ultérieur d'études sur des études utilisant le SRM pour examiner le glutamate dans le TDM a confirmé que la concentration de glutamate dans l'ACC était systématiquement réduite (Luykx et al., 2012). Certains chercheurs ont observé une augmentation du glutamate dans le cortex occipital, plus élevée dans le sous-groupe mélancolique de patients atteints de TDM (Sanacora et al., 2008).
Dans une étude précédente, nous avions obtenu des images TEP de la liaison du récepteur mGluR5 chez des sujets non médicamentés 11 avec des témoins sains MDD et 11 (Deschwanden et al., 2011). Nous avons constaté une diminution de la liaison régionale de mGluR5 dans le cortex préfrontal, le cortex cingulaire, l'insula, le thalamus et l'hippocampe des patients souffrant de dépression. De plus, la sévérité de la dépression était négativement corrélée à la liaison de mGluR5 dans l'hippocampe. Nous avons suggéré que ces résultats indiquent une diminution de la neurotransmission de mGluR5 dans la dépression, possiblement à la suite de modifications basales ou compensatoires de l'activité du système du glutamate. En outre, nous avons examiné la quantité d’expression de mGluR5 dans des échantillons de cerveau post-mortem de sujets déprimés 15 et de témoins appariés à 15 (Deschwanden et al., 2011). Nous avons observé une expression réduite de mGluR5 dans le cortex préfrontal dans des échantillons prélevés chez des personnes dépressives. Une expression réduite des récepteurs NMDA dans le cerveau post-mortem de patients déprimés a également été rapportée (Feyissa et al., 2009). En effet, il a déjà été suggéré que les propriétés antidépressives des antagonistes de mGluR5 pourraient impliquer une inhibition du récepteur NMDA. Cela pourrait induire une neurotransmission et / ou une induction de l’expression du gène du facteur neurotrope dérivé du cerveau dans l’hippocampe (Legutko et al., 2006). En outre, une étude récente a montré que la privation de sommeil augmentait la disponibilité de mGluR5 chez l’homme en bonne santé (Hefti et al., 2013). Dans le cortex cingulaire antérieur, l'insula, le lobe temporal médial, le gyrus parahippocampique, le striatum et l'amygdale, cette augmentation était significativement corrélée à l'efficacité de la privation de sommeil, reflétée par une somnolence subjective accrue. Cette étude suggère qu'une augmentation de mGluR5 pourrait être un mécanisme neurobiologique expliquant l'efficacité antidépressive élevée de la privation de sommeil. Les études précliniques confirment notre association hypothétique entre le mGluR5, le sommeil et la dépression. En particulier, une étude chez des souris knock-out mGluR5 fournit des preuves importantes que le mGluR5 est impliqué dans le façonnement de la stabilité des transitions de l'état de sommeil NREM-sommeil paradoxal, de l'activité d'onde lente NREM et de la réponse homéostatique à la perte de sommeilAhnaou et al., 2015).
Pour résumer, il existe des preuves convergentes d'études animales, post mortem, MRS et PET indiquant que le système central du glutamate joue un rôle important dans la physiopathologie du TDM. Cependant, les preuves suggèrent que l'antagonisme de mGluR5 pourrait ne pas aider directement les patients atteints de TDM. Ceci est conforme à notre découverte d'une expression réduite de mGluR5 chez les patients atteints de TDM. On pourrait supposer que les médicaments qui ciblent le système mGluR5 pourraient être particulièrement utiles chez les patients déprimés souffrant d'anxiété comorbide, de troubles de la dépendance et / ou de troubles du rythme circadien. En outre, les dysfonctionnements dans les systèmes de processus sociaux sont généralement associés à la DMD. De faibles compétences sociales sont apparues comme un facteur de risque important de la dépression (Segrin, 2000). Ces déficits, y compris les comportements paralinguistiques et linguistiques, ainsi que les altérations de l’expression faciale, du regard, de la posture et des gestes, sont comparables à ceux observés dans le syndrome d’effet FX et le trouble du spectre autistique. En outre, en ce qui concerne les compétences sociales, des expériences sur des souris exposées à un stress social chronique ont montré que le couplage Homer1 / mGluR5 était interrompu, ce qui suggère que mGluR5 modère de nuit la vulnérabilité dépressive au stress social (Wagner et al., Sous presse). De plus, dans les modèles murins de déficits sociaux, la suppression de mGluR5 a conduit à la normalisation des interactions sociales (Chung et al., 2015). Ensemble, ces études fournissent des preuves précliniques que mGluR5 joue un rôle important dans les causes sociales de la dépression et les déficits sociaux fréquemment observés chez les patients déprimés. En conséquence, les médicaments ciblant mGluR5 peuvent jouer un rôle important dans la prévention de la dépression chez les jeunes présentant un déficit social et peuvent contribuer à traiter les déficits sociaux et le fonctionnement psychosocial faible des patients atteints de TDM.
mGluR5 et toxicomanie
La toxicomanie se caractérise par une consommation continue de drogues malgré des conséquences négatives, des tentatives répétées d’arrêter ou de réduire la consommation de drogues et des symptômes de tolérance et de sevrage. Bien que le système dopaminergique joue un rôle clé dans le traitement de la récompense aiguë (Kalivas et Volkow, 2005), de plus en plus de preuves impliquent une neurotransmission glutamatergique dans le traitement de la toxicomanie (Krystal et al., 2010).
Recherche neurobiologique préclinique
Dans 2001, une étude fondamentale sur mGluR5 et la dépendance a été publiée (Chiamulera et al., 2001). Les auteurs ont montré que des souris dépourvues du récepteur mGluR5 ne parvenaient pas à s'auto-administrer par voie intraveineuse de cocaïne malgré une augmentation des taux de dopamine extracellulaire dans le noyau accumbens après une injection aiguë. De nombreuses études de suivi chez l’animal ont montré que les antagonistes des récepteurs mGluR5, MPEP et MTEP, réduisent l’auto-administration de médicaments entraînant une dépendance tels que la cocaïne et la nicotine (Kalivas, 2009). Olive (2005) ont passé en revue des études sur des animaux liées à la toxicomanie et ont suggéré que mGluR5 pourrait être impliqué dans le développement, la perception des récompenses et la rechute de la consommation de drogues telles que la cocaïne, la morphine, la nicotine et l’éthanol. Les auteurs ont décrit des preuves d'études animales montrant que les antagonistes du mGluR5 réduisaient l'auto-administration du médicament, ainsi que le comportement de recherche du médicament. Par exemple, il a été découvert récemment, dans un modèle de trouble de la frénésie alimentaire chez le babouin, que le MTEP diminuait la consommation de bonbons sans modifier le comportement de recherche de bonbons (Bisaga et al., 2008). Les recherches ont suggéré que la diminution de l'apport alimentaire associée à l'utilisation d'antagonistes des récepteurs mGluR5 pourrait être liée à une réduction de la valeur enrichissante des stimuli de renforcement (Bisaga et al., 2008). De plus, des recherches ont montré une augmentation significative des niveaux d’ARNm de mGluR5 dans le noyau accumbens et le striatum dorsolatéral après administration répétée de cocaïne chez le rat de laboratoire (Bisaga et al., 2008).
Études humaines
En utilisant une autoradiographie dans des échantillons de tissu cérébral post-mortem provenant de patients souffrant de troubles alcooliques et de témoins en bonne santé, les recherches ont révélé une densité de liaison 30 – 40% supérieure mGluR1 / 5 dans l’hippocampe et le striatum chez les patients souffrant de troubles alcooliques (Kupila et al., 2012). Cette découverte suggère que la densité des récepteurs mGluR5 pourrait être augmentée dans certaines zones du cerveau de patients toxicomanes. Récemment, nous avons utilisé la TEP pour mesurer la liaison du récepteur mGluR5 dans le cerveau des personnes ayant une dépendance à la nicotine (Akkus et al., 2013). Nous avons constaté une réduction globale du mGluR5 DVR dans la substance grise des fumeurs de 14 par rapport aux non-fumeurs (voir la figure 2). Les réductions les plus importantes ont été observées dans les cortex orbitofrontal médial droit et gauche. Nous avons suggéré que cette diminution de la liaison aux récepteurs mGluR5 pourrait constituer une adaptation durable aux augmentations chroniques du glutamate induites par une administration chronique de nicotine. Cette adaptation semble être spécifique à la dépendance à la nicotine puisqu'elle n'a pas été trouvée chez les consommateurs chroniques de cocaïne non-fumeurs (Hulka et al., 2014).
FIGURE 2
Figure 2. L'image montre l'absorption moyenne de mGluR5 DVR par le cerveau dans les trois groupes de diagnostic. L'absorption du cerveau est visible réduite chez les groupes de fumeurs et d'anciens fumeurs. Les images sont calculées avec la version du logiciel PMOD 3 (PMOD Technologies). De Akkus et al. (2013). Remarque: la figure et sa légende sont reproduites avec autorisation.
En effet, il a été suggéré que la régulation négative de mGluR5 représente une neuroadaptation compensatoire (Kalivas, 2009), un stimulant de la récompense induite par le médicament (Rutten et al., 2011), ou un facteur médiant les effets des signaux contextuels dans les réponses comportementales conditionnées (Tronci et al., 2010). Figure 3 présente une approche du dysfonctionnement mGluR5 dans la toxicomanie.
FIGURE 3
Figure 3. Implication de mGluR5 dans la dépendance.
Comme le montre la figure 3, Les recherches précédentes soutiennent l’idée que mGluR5 est impliqué dans trois phases clés de la dépendance, dans le développement et l’acquisition, dans la valeur de renforcement de la drogue, ainsi que dans la rechute de la dépendance. On pourrait suggérer que chaque fonction est principalement décrite par différentes régions du cerveau, qui présentent également une densité élevée de récepteurs mGluR5, et dont la densité réduite de mGluR5 a été démontrée dans notre recherche sur la TEP (Deschwanden et al., 2011). Kalivas (2009) développé un vaste modèle de dépendance au glutamate. L'auteur a suggéré que la dépendance était associée à un dysfonctionnement de l'homéostasie du glutamate entre des zones clés du circuit cérébral cortico-strié, y compris l'amygdale, le noyau accumbens (NA), le cortex préfrontal et le cortex moteur. Comme nous le suggérons dans la figure 3Premièrement («composante de motivation»), le cortex préfrontal pourrait être un moteur de la motivation et du contrôle cognitif au cours des phases initiales du développement de la toxicomanie. L’étude préclinique de Chiamulera et al. (2001) suggère que mGluR5 est nécessaire dans cette phase de développement de la dépendance en médiant les propriétés gratifiantes des drogues d'abus. Deuxièmement («composant de récompense»), il a été démontré que le Nucleus Accumbens (NA) influence la valeur de récompense du médicament, non seulement via la dopamine, mais également via l’activité mGluR5 (Bisaga et al., 2008). En particulier, une altération de la transmission du glutamate entre le cortex pré-limique et l'AN a été suggérée, de telle sorte que la recherche de drogue a été initiée par une plus grande confiance dans le comportement acquis associé au système de récepteurs NMDA / mGluR5 (Kalivas, 2009). Et troisièmement («composant de recherche de médicaments»), certaines recherches soutiennent que mGluR5 est impliqué dans une plus grande dépendance vis-à-vis des processus moteurs - réduisant le contrôle cognitif - via le striatum (Kupila et al., 2012). Nos propres recherches publiées et non publiées suggèrent que la régulation négative de mGluR5 est associée à un risque accru de rechute des anciens fumeurs (Akkus, PNAS). Cette régulation négative peut être une modification pathogène ou insuffisante. Dans l’ensemble, les recherches actuelles suggèrent que les médicaments ciblant mGluR5 pourraient améliorer les traitements des troubles de la toxicomanie à différents stades de leur développement.
a lieu
Les preuves examinées suggèrent une forte implication de mGluR5 dans les troubles anxieux, les TOC, le TDM, ainsi que la toxicomanie, et un traitement par des médicaments ciblant mGluR5 pourrait également être bénéfique chez l'homme. Cependant, la pathogenèse des symptômes de l'humeur et de l'anxiété associés au TOC peut différer des symptômes de l'humeur et de l'anxiété non liés au TOC. En conséquence, la présence de symptômes obsessionnels compulsifs peut être un facteur prédictif important de la réponse antidépressive et anxiolytique à des médicaments ciblant mGluR5. En outre, nous avons suggéré que les médicaments ciblant le système mGluR5 pourraient contribuer à accroître la résistance au stress social et à améliorer les déficits sociaux liés à la dépression. Étant donné que le stress social est de loin le principal facteur de risque non génétique de la dépression et que les déficits sociaux sont étroitement liés à la réduction du fonctionnement social et de la qualité de la vie, les résultats d'études animales revêtent une grande pertinence scientifique et clinique.
L'Institut national de la santé mentale (NIMH) a récemment lancé un projet novateur et à la pointe de la technologie: les critères du domaine de la recherche (RDoC). Cela reflète la mise en œuvre de la stratégie NIMH 1.4 «Développer, à des fins de recherche, de nouvelles méthodes de classification des troubles mentaux basées sur les dimensions du comportement observable et les mesures neurobiologiques.» (http://www.nimh.nih.gov/research-priorities/strategic-objectives/strategic-objective-1.shtml). Cette initiative est le résultat de la restructuration de catégories de DSM rigides, dont la plupart ont également été développées avant la recherche en neurosciences (Morris et Cuthbert, 2012). RDoC décrit cinq domaines ou constructions; Système de valence négative, Système de valence positive, Systèmes cognitifs, Systèmes pour processus sociaux, Systèmes d'éveil / régulation. Conformément aux preuves examinées sur l'implication de mGluR5 dans les MDD et les TOC, nous proposons que l'activité de mGluR5 soit directement associée au système de valence négative, qui implique les facteurs observables de peur, de menace, de menace persistante, de perte et de non-frustration frustrée. récompense. Ainsi, comme le suggèrent les études examinées, un traitement antagoniste avec mGluR5 devrait réduire ces symptômes observables. De plus, nous suggérons que le traitement par mGluR5 pourrait également être bénéfique dans les «troubles de valence positive» tels que la dépendance et la dépression, via une activité anormale de mGluR5 sur la structure et la fonction cérébrales liées au récepteur NMDA glutamatergique fonctionnellement lié à l'apprentissage par récompense . Par exemple, Simonyi et al. (2010) ont examiné de nombreuses études sur des animaux utilisant des antagonistes des récepteurs mGluR5 chez des souris knock-out pour déterminer le rôle de mGluR5 dans l'apprentissage et la mémoire. L’apprentissage inhibiteur, tel que l’apprentissage par évitement passif, est une tâche bien établie sur des modèles animaux et utilisée pour étudier les processus d’apprentissage de l’hippocampe. De nombreuses études ont montré qu’elle dépendait du récepteur mGluR5 (Simonyi et al., 2010). Par exemple, des recherches ont démontré une hyperexpression de la protéine mGluR5 dans CA3 au cours de la potentialisation à court et à long terme de CA1 chez le rat (Riedel et al., 2000). Hyman (2005) a présenté un modèle biologique de dépendance intégrant des processus neuronaux anormaux d'apprentissage et de mémoire constituant les éléments de base de la dépendance. Les auteurs ont proposé cette potentialisation à long terme, notamment les modifications de la disponibilité des récepteurs du glutamate et la régulation de l'expression des gènes en tant que mécanismes potentiellement importants pour les modifications induites par le médicament détectées dans les circuits anormaux associés à la toxicomanie. Enfin, les études sur mGluR5 et l’homéostasie du sommeil (Hefti et al., 2013; Ahnaou et al., 2015) suggèrent un rôle important du mGluR5 dans le domaine des systèmes d'excitation et de modulation de RDoC.
Figure 4 résume les processus proposés pour médier les actions de mGluR5 dans les troubles de l'humeur et la toxicomanie. Le haut de la figure 4 décrit les régions du cerveau mGluR5 à haute densité; amygdale, hippocampe, striatum, NA et cortex préfrontal. Les syndromes psychiatriques sont appariés à ces régions. L'activité de mGluR5, suggérée dans l'amygdale, pourrait être un médiateur de l'excitation émotionnelle primaire, telle que l'anxiété et la dépression. Nous avons décrit comment deux voies possibles; le fonctionnement social et l'apprentissage peuvent jouer un rôle médiateur dans les autres processus. Ainsi, l'activité de mGluR5 s'est avérée impliquée dans l'apprentissage et, par conséquent, via l'activité dans l'hippocampe, NA et le striatum pourrait être impliquée dans la mémoire et la récompense, le contrôle cognitif et la motivation, ainsi que dans l'addiction (voir aussi la figure). 3). Enfin, nous avons suggéré comment mGluR5 pourrait être lié à la réponse au stress social et aux déficits sociaux, qui pourraient être pertinents pour un large éventail de conditions psychiatriques. Table 2 donne un aperçu de la zone de dégradation RDoC, du tableau clinique associé et de la participation de mGluR5.
FIGURE 4
Figure 4. Résumé des mécanismes de mGluR5.
TABLEAU 2
![]()
Tableau 2. RDoC et mGluR5.
Remarques finales
Cette revue a décrit l'implication de mGluR5 dans les troubles de l'humeur, les TOC et la dépendance, et a comparé la recherche préclinique à la recherche humaine, en particulier la recherche PET. Nous avons donc comparé différentes approches méthodologiques, telles que la recherche sur les animaux, la MRS et les études PET. Nous suggérons qu'un puissant effet anxiolytique direct serait présent si le traitement antagoniste mGluR5 était initié lors d'essais cliniques sur l'homme (RDoC). Une suractivité de mGluR5 a également été rapportée dans FXS, caractérisé par des déficits sociaux importants. En conséquence, l'activité de mGluR5 peut non seulement normaliser l'activité dans les systèmes de valence négative et les systèmes d'éveil, mais également atténuer les déficiences des systèmes de processus sociaux (RDoC). Cela revêt une grande importance clinique, car un fonctionnement social médiocre est un résultat important des maladies psychiatriques courantes telles que les TOC, la dépression et la toxicomanie, entraînant d’énormes souffrances personnelles et des coûts indirects importants pour la société. Enfin, il a également été démontré que mGluR5 avait une implication significative dans la toxicomanie, ce qui pourrait être causé principalement par l’augmentation de la valeur de récompense de la drogue. Une intervention antagoniste de mGluR5 serait donc plus efficace pour le traitement de l’anxiété pathologique et de la toxicomanie, ainsi que pour améliorer la résilience au stress social et le fonctionnement social.
Déclaration de conflit d'intérêts
Terbeck, Chesterman et Akkus n’ont pas de conflit d’intérêts. Le Dr Hasler a reçu une subvention de Novartis, qui produit et teste des médicaments ciblant mGluR5.
Bibliographie
Abe, T., H. Sugihara, H. Nawa, R. Shigemoto, N. Mizuno et S. Nakanishi (1992). Caractérisation moléculaire d'un nouveau récepteur métabotropique du glutamate mGluR5 couplé à la transduction du signal inositol phosphate / Ca2 +. J. Biol. Chem. 267, 13361 – 13368.
Résumé PubMed | Texte intégral | Google Scholar
Agid, Y., Buzsáki, G., Diamond, DM, Frackowiak, R., Giedd, J., Girault, J.-A., et al. (2007). Comment améliorer la découverte de médicaments pour les troubles psychiatriques? Nat. Rev. Drug Discov. 6, 189 – 201. doi: 10.1038 / nrd2217
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Akkus, F., Ametamey, SM, Treyer, V., Burger, C., A. Johayem, A., Umbricht, D., et al. (2013). Réduction globale marquée de la liaison du récepteur mGluR5 chez les fumeurs et les ex-fumeurs déterminée par tomographie par émission de positrons [11C] ABP688. Proc. Natl. Acad. Sci. Etats-Unis. 110, 737 – 742. doi: 10.1073 / pnas.1210984110
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Akkus, F., Terbeck, S., Ametamey, SM, Rufer, MD, Treyer, V., Burger, C. et al. (2014). Liaison du récepteur métabotropique du glutamate à 5 chez les patients présentant un trouble obsessionnel-compulsif. Int. J. Neuropsychopharmacol. 17, 1915 – 1922. doi: 10.1017 / S1461145714000716
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Ametamey, SM, Kessler, LJ, Honer, M., Wyss, MT, Buck, A., Hintermann, S., et al. (2006). La radiosynthèse et l'évaluation préclinique de 11CABP688 en tant que sonde d'imagerie du sous-type de récepteur métabotropique du glutamate, 5. J. Nucl. Med. 47, 698 – 705.
Résumé PubMed | Texte intégral | Google Scholar
Anderson, JJ, Bradbury, MJ, Giracello, DR, Chapman, DF, Holtz, G., Roppe, J., et al. (2003). in vivo occupation du récepteur des antagonistes du récepteur mGlu5 par le nouveau radioligand [3 H] 3-méthoxy-5- (pyridine-2-yléthynyl) pyridine). EUR. J. Pharmacol. 473, 35–40. doi: 10.1016/S0014-2999(03)01935-6
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Ahnaou, A., Raeymaekers, L., Steckler, T. et Drinkenbrug, WHIM (2015). Pertinence du récepteur métabotropique du glutamate (mGluR5) dans la régulation du cycle de sommeil NREM-REM et de l'homéostasie: mise en évidence chez la souris mGluR5 (- / -). Comportement Brain Res. 282, 218 – 226. doi: 10.1016 / j.bbr.2015.01.009
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Bear, MF, Huber, KM et Warren, ST (2004). La théorie mGluR du retard mental X fragile. Tendances Neurosci. 27, 370 – 377. doi: 10.1016 / j.tins.2004.04.009
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
D. Berg, J. Godau, C. Trenkwalder, K. Eggert, I. Csoti, A. Storch, et al. (2011). Traitement AFQ056 des dyskinésies induites par la lévodopa: résultats des essais comparatifs randomisés sur 2. Mov. Désordre. 26, 1243 – 1250. doi: 10.1002 / mds.23616
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Berry-Kravis, E., D. Hessl, S. Coffey, C. Hervey, A. Schneider, J. Yuhas et autres. (2009). Essai pilote à dose unique en ouvert sur le fenobam chez des adultes atteints du syndrome de l’X fragile. J. Med. Genet. 46, 266 – 271. doi: 10.1136 / jmg.2008.063701
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Bisaga, A., Danysz, W. et Foltin, RW (2008). L'antagonisme des récepteurs glutamatergiques NMDA et mGluR5 diminue la consommation d'aliments dans le modèle babouin du syndrome de la frénésie alimentaire. EUR. Neuropsychopharmacol. 18, 794 – 802. doi: 10.1016 / j.euroneuro.2008.05.004
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Boer K., Encha-Razavi, F., Sinico, M. et Aronica, E. (2010). Distribution différentielle des récepteurs métabotropes du glutamate du groupe I dans le cortex humain en développement. Brain Res. 1324, 24 – 33. doi: 10.1016 / j.brainres.2010.02.005
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Brennan, BP, Rauch, SL, Jensen, JE et Pope, HG (2012). Une revue critique des études de spectroscopie par résonance magnétique du trouble obsessionnel-compulsif. Biol. Psychiatrie 73, 31 – 24. doi: 10.1016 / j.biopsych.2012.06.023
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Burket, JA, Benson, AD, Tang, AH et Deutsch, SI (2014). La rapamycine améliore la sociabilité dans le modèle murin BTBR de troubles du spectre autistique. Brain Res. Taureau. 100, 70 – 75. doi: 10.1016 / j.brainresbull.2013.11.005
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Chiamulera, C., MP Epping-Jordan, A. Zocchi, C. Marcon, C. Cottiny, S. Tacconi et al. (2001). Les effets stimulants et stimulants locomoteurs de la cocaïne sont absents chez les souris mutantes nulles mGluR5. Nat. Neurosci. 4, 873 – 874. Doi: 10.1038 / nn0901-873
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
W. Chung, SY Choi, E. Lee, H. Park, H. J. Kang., H. Park et al. (2015). Les déficits sociaux chez les souris mutantes IRSp53 sont améliorés par la suppression de NMDAR et de mGluR5. Nat. Neurosci. 18, 435 – 442. doi: 10.1038 / nn.3927
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Cleva, RM et Olive, MF (2011). Modulateurs allostériques positifs des récepteurs métabotropiques du glutamate de type 5 (mGluR5) et leur potentiel thérapeutique pour le traitement des troubles du système nerveux central. Molécules 16, 2097 – 2106. doi: 10.3390 / molécules16032097
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Deschwanden, A., Karolewicz, B., Feyissa, AM, Treyer, V., Ametamey, SM, Johayem, A. et al. (2011). Réduction de la densité 5 du récepteur métabotropique du glutamate dans la dépression majeure déterminée par [(11) C] ABP688 PET et une étude post mortem. Un m. J. Psychiatrie 168, 727 – 734. doi: 10.1176 / appi.ajp.2011.09111607
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Erickson, CA, Mullett, JE et McDougle, CJ (2009). La mémantine en ouvert dans le syndrome de l'X fragile. J. Autism Dev. Désordre. 39, 1629 – 1635. Paramètres: 10.1007 / s10803-009-0807-3
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Erickson, CA, Early, M., Stigler, KA, Wink, LK, Mullett, JE et McDougle, CJ (2011). Une étude pilote naturaliste ouverte sur l'acamprosate chez les jeunes atteints de trouble autistique. J. Enfant Adolesc. Psychopharmacol. 21, 565 – 569. doi: 10.1089 / cap.2011.0034
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Premièrement, MB et Gibbon, M. (1997). Guide de l'utilisateur pour l'entrevue clinique structurée pour les troubles de l'axe I du DSM-IV SCID-I: version clinicien. New York, NY: American Psychiatric Pub.
Feyissa, AM, Chandran, A., Stockmeier, CA et Karolewicz, B. (2009). Réduction des niveaux de sous-unités NR2A et NR2B du récepteur NMDA et de PSD-95 dans le cortex préfrontal dans la dépression majeure. Programme. Neuropsychopharmacol. Biol. Psychiatrie 33, 70 – 75. doi: 10.1016 / j.pnpbp.2008.10.005
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Garber, KB, J. Visootsak et Warren, ST (2008). Syndrome de l'X fragile. EUR. J. Hum. Genet. 16, 666 – 672. doi: 10.1038 / ejhg.2008.61
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Gass, JT, Osborne, député, Watson, NL, Brown, JL et Olive, MF (2009). L'antagonisme de mGluR5 atténue le renforcement de la méthamphétamine et empêche le rétablissement du comportement de recherche de méthamphétamine chez le rat. Neuropsychopharmacology. 34, 820 – 833. doi: 10.1038 / npp.2008.140
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Harvey, BH et Shahid, M. (2012). Les récepteurs métabotropes et ionotropes du glutamate en tant que cibles neurobiologiques dans les troubles anxieux et liés au stress: focus sur la pharmacologie et les modèles de traduction précliniques. Pharmacol. Biochem. Comportement. 100, 775 – 800. doi: 10.1016 / j.pbb.2011.06.014
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Hasler, G., LaSalle-Ricci, VH, Ronquillo, JG, Crawley, SA, Cochran, LW, Kazuba, D. et al. (2005). Les dimensions des symptômes du trouble obsessionnel-compulsif montrent des relations spécifiques avec la comorbidité psychiatrique. Res psychiatrie. 135, 121 – 132. doi: 10.1016 / j.psychres.2005.03.003
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Hasler, G., JW van der Veen, T. Tumonis, N. Meyers, J. Shen et Drevets, WC (2007). Réduction des taux de glutamate / glutamine préfrontal et d'acide γ-aminobutyrique dans la dépression majeure, déterminée par spectroscopie par résonance magnétique proton. Cambre. Psychiatrie générale 64, 193 – 200. doi: 10.1001 / archpsyc.64.2.193
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Hasler, G., van der Veen, JW, Geraci, M., Shen, J., D., Pine et Drevets, WC (2009). Niveaux d'acide gamma-aminobutyrique corticaux préfrontal dans le trouble panique, déterminés par spectroscopie à résonance magnétique du proton. Biol. Psychiatrie 65, 273 – 275. doi: 10.1016 / j.biopsych.2008.06.023
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Hefti, K., Holst, SC, J. Sovago, V. Bachmann, A. Buck, A. Ametamey, SM et al. (2013). Augmentation de la disponibilité du sous-type de 5 du récepteur métabotropique du glutamate dans le cerveau humain après une nuit sans sommeil. Biol. Psychiatrie 73, 161 – 168. doi: 10.1016 / j.biopsych.2012.07.030
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Hyman, SE (2005). Dépendance: une maladie de l'apprentissage et de la mémoire. Un m. J. Psychiatrie 162, 1414 – 1422. doi: 10.1176 / appi.ajp.162.8.1414
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Hoffpauir, BK et Gleason, EL (2002). L'activation de mGluR5 module la fonction du récepteur GABAA dans les cellules amacrines rétiniennes. J. Neurophysiol. 88, 1766 – 1776.
Résumé PubMed | Texte intégral | Google Scholar
Hulka, LM, Treyer, V., Scheidegger, Preller, KH, Vonmoos, M., Baumgartner, MR, et al. (2014). Le tabagisme, mais pas la consommation de cocaïne, est associé à la densité 5 du récepteur métabotropique du glutamate cérébral inférieur chez l'homme. Mol. Psychiatrie 19, 625 – 632. doi: 10.1038 / mp.2013.51
CrossRef Full Text | Google Scholar
Kalivas, PW et Volkow, ND (2005). La base neurale de la dépendance: une pathologie de la motivation et du choix. Un m. J. Psychiatrie 162, 1403 – 1413. doi: 10.1176 / appi.ajp.162.8.1403
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Kalivas, PW (2009). L'hypothèse d'homéostasie du glutamate de la dépendance. Nat. Rev. Neurosci. 10, 561 – 572. doi: 10.1038 / nrn2515
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Krystal, JH, Mathew, SJ, D'Souza, DC, Garakani, A., Gunduz-Bruce, H., et Charney, DS (2010). Applications psychiatriques potentielles des agonistes et antagonistes des récepteurs métabotropiques du glutamate. Drogues du SNC 24, 669–693. doi: 10.2165/11533230-000000000-00000
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Kupila, J., Kärkkäinen, O., Laukkanen, V., Tupala, E., Tiihonen, J. et Storvik, M. (2012). Densité des récepteurs mGluR1 / 5 dans le cerveau de sujets alcooliques: étude par autoradiographie sur tout l'hémisphère. Res psychiatrie. 12, 149 – 155. doi: 10.1016 / j.pscychresns.2012.04.003
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Koros, E., H. Rosenbrock, G. Birk, C. Weiss et Sams-Dodd, F. (2007). Le MTEP, un antagoniste sélectif du récepteur mGlu5, similaire aux antagonistes du récepteur NMDA, induit l'isolement social chez le rat. Neuropsychopharmacology 32, 562 – 576. doi: 10.1038 / sj.npp.1301133
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Lea, PM et Faden, AI (2006). Les récepteurs métabotropiques du glutamate de sous-type 5 antagonistes MPEP et MTEP. CNS Drug Rev. 12, 149 – 166. Doi: 10.1111 / j.1527-3458.2006.00149.x
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Legutko, B., Szewczyk, B., L. Pomierny-Chamioło, G., Nowak et A. Pilc (2006). Effet du traitement MPEP sur l'expression des gènes du facteur neurotrophique dérivé du cerveau. Pharmacol. Représentant. 58, 427 – 430.
Résumé PubMed | Texte intégral | Google Scholar
Luykx, JJ, Laban, KG, van den Heuvel, député, Boks, MPM, Mandl, RCW, Kahn, RS, et al. (2012). Régulation à la baisse du glutamate spécifique à une région et à un état dans le trouble dépressif majeur: méta-analyse des résultats de (1) H-MRS. Neurosci. Biobehav. Tour. 36, 198 – 205. doi: 10.1016 / j.neubiorev.2011.05.014
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Meldrum, BS (2000). Le glutamate en tant que neurotransmetteur dans le cerveau: examen de la physiologie et de la pathologie. J. Nutr. 130, 1007.
Résumé PubMed | Texte intégral | Google Scholar
Morris, SE et Cuthbert, BN (2012). Critères du domaine de recherche: systèmes cognitifs, circuits neuronaux et dimensions du comportement. Dialogues Clin. Neurosci. 14, 29 – 37.
Résumé PubMed | Texte intégral | Google Scholar
Olive, MF (2005). Récepteurs mGlu5: neuroanatomie, pharmacologie et rôle dans la toxicomanie. Curr. Psychiatrie Rev. 1, 197 – 214. doi: 10.2174 / 1573400054065578
CrossRef Full Text | Google Scholar
Phan, KL, Fitzgerald, DA, Cortese, BM, Seraji-Bozorgzad, N., Tancer, ME et Moore, GJ (2005). Neurochimie du cingulum antérieur dans le trouble d'anxiété sociale: 1H-MRS chez 4 Tesla. Neuroreport 16, 183–186. doi: 10.1097/00001756-200502080-00024
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Pittenger, C., Krystal, JH et Coric, V. (2006). Les médicaments modulant le glutamate en tant que nouveaux agents pharmacothérapeutiques dans le traitement du trouble obsessionnel-compulsif. NeuroRx 3, 69 – 81. doi: 10.1016 / j.nurx.2005.12.006
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Pollack, MH, Jensen, JE, Simon, NM, Kaufmann, RE et Renshaw, PF (2008). Etude MRS à haut champ sur le GABA, le glutamate et la glutamine dans le trouble d'anxiété sociale?: Réponse au traitement par le lévétiracétam. Programme. Neuropsychopharmacol. Biol. Psychiatrie 32, 739 – 743. doi: 10.1016 / j.pnpbp.2007.11.023
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Riaza Bermudo-Soriano, C., Perez-Rodriguez, MM, Vaquero-Lorenzo, C. et Baca-Garcia, E. (2012). Nouvelles perspectives dans le glutamate et l'anxiété. Pharmacol. Biochem. Comportement. 100, 752 – 774. doi: 10.1016 / j.pbb.2011.04.010
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Riedel, G., Casabona, G., Platt, B., Macphail, EM et Nicoletti, F. (2000). Augmentation de l'expression de la protéine du récepteur mGlu5 dans l'hippocampe de rat en fonction du temps et du sous-région, provoquée par le conditionnement. Neuropharmacologie 39, 1943–1951. doi: 10.1016/S0028-3908(00)00037-X
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Rutten, K., De Vry, J., Bruckmann, W. et Tzschentke, TM (2011). Le blocage pharmacologique ou l'inactivation génétique du récepteur NOP potentialise l'effet de récompense de la morphine chez le rat. La drogue dépend de l'alcool. 114, 253 – 256. doi: 10.1016 / j.drugalcdep.2010.10.004
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Romano, C., van den Pol, AN et O'Malley, KL (1996). Expression développementale précoce améliorée du récepteur métabotropique du glutamate mGluR5 dans le cerveau de rat: protéines, variantes d'épissage d'ARNm et distribution régionale. J. Compar. Neurol. 367, 403 – 412.
Rosenberg, DR et Keshavan, MS (1998). Vers un modèle neurodéveloppemental du trouble obsessionnel-compulsif. Biol. Psychiatrie 43, 623–640. doi: 10.1016/S0006-3223(97)00443-5
CrossRef Full Text | Google Scholar
Sanacora, G., Zarate, CA, Krystal, JH et Manji, HK (2008). Cibler le système glutamatergique pour développer de nouvelles thérapies améliorées pour les troubles de l'humeur. Nat. Rev. Drug Discov. 7, 426 – 437. doi: 10.1038 / nrd2462
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Schoepp, DD (2001). Dévoiler les fonctions des récepteurs présynaptiques du glutamate métabotropique dans le système nerveux central. J. Pharmacol. Exp. Ther. 299, 12 – 20.
Résumé PubMed | Texte intégral | Google Scholar
Segrin, C. (2000). Déficits en compétences sociales associés à la dépression. Clin. Psychol. Tour. 20, 379–403. doi: 10.1016/S0272-7358(98)00104-4
CrossRef Full Text | Google Scholar
Simonyi, A., Schachtman, TR et Christoffersen, GR (2010). Récepteur métabotropique du glutamate, sous-type 5, antagoniste de l'apprentissage et de la mémoire. EUR. J. Pharmacol. 639, 17 – 25. doi: 10.1016 / j.ejphar.2009.12.039
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Soares, DP et Law, M. (2009). Spectroscopie par résonance magnétique du cerveau: examen des métabolites et applications cliniques. Clin. Radiol. 64, 12 – 21. doi: 10.1016 / j.crad.2008.07.002
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Le juge en chef Swanson, MM. Bures, Johnson, MP, Linden, A.-M., Monn, JA et Schoepp, DD (2005). Les récepteurs métabotropiques du glutamate en tant que nouvelles cibles pour les troubles anxieux et de stress. Nat. Rev. Drug Discov. 4, 131 – 144. doi: 10.1038 / nrd1630
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Szeszko, PR, Christian, C., Macmaster, F., Lencz, T., Mirza, Y., Taormina, SP, et al. (2008). Altérations structurelles de la matière grise dans le trouble obsessionnel-compulsif pédiatrique naïve à la drogue psychotrope: étude optimisée de la morphométrie à base de voxel. Un m. J. Psychiatrie 165, 1299 – 1307. doi: 10.1176 / appi.ajp.2008.08010033
CrossRef Full Text | Google Scholar
E. Tatarczyńska, A. Klodzińska, E. E. Chojnacka-Wójcik, A. Palucha, F. Gasparini, R. Kuhn et coll. (2001). Les effets potentiels anxiolytiques et antidépresseurs de MPEP, un antagoniste puissant, sélectif et systémique du récepteur mGlu5. Le fr. J. Pharmacol. 132, 1423 – 1430. doi: 10.1038 / sj.bjp.0703923
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Tronci, V., S. Vronskaya, N. Montgomery, D. Mura et Balfour, DJK (2010). Les effets de l'antagoniste du récepteur mGluR5 6-méthyl-2- (phényléthynyl) -pyridine (MPEP) sur les réponses comportementales à la nicotine. Psychopharmacologie 211, 33–42. doi: 10.1007/s00213-010-1868-x
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Van den Heuvel, OA, Remijnse, PL, D. Mataix-Cols, H. Vrenken, HJ Groenewegen, Uylings, HBM, et al. (2009). Les principaux symptômes du trouble obsessionnel-compulsif sont véhiculés par des systèmes neuronaux partiellement distincts. Cerveau 132, 853 – 868. doi: 10.1093 / brain / awn267
CrossRef Full Text | Google Scholar
Wagner, KV, J. Hartmann, C. Labermaier, AS Häusl, G. Zhao, D. Harbich et al. (dans la presse). L'activité Homer1 / mGluR5 modère la vulnérabilité au stress social chronique. Neuropsychopharmacology. doi: 10.1038 / npp.2014.308
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Yüksel, C. et Öngür, D. (2010). Études par spectroscopie à résonance magnétique des anomalies liées au glutamate dans les troubles de l'humeur. Biol. Psychiatrie 68, 785 – 794. doi: 10.1016 / j.biopsych.2010.06.016
CrossRef Full Text | Google Scholar
Zarei, M., D. Mataix-Cols, I. Heyman, M., Hough, J., Doherty, L. Burge, et al. (2011). Modifications du volume de la matière grise et de la microstructure de la substance blanche chez les adolescents présentant un trouble obsessionnel-compulsif. Biol. Psychiatrie 70, 1083 – 1090. doi: 10.1016 / j.biopsych.2011.06.032
Résumé PubMed | Texte intégral | CrossRef Full Text | Google Scholar
Mots-clés: mGluR5, PET, troubles de l'humeur, dépendance, anxiété
Citation: Terbeck S, Akkus F, Chesterman LP et Hasler G (2015) Le rôle du récepteur métabotropique du glutamate 5 dans la pathogenèse des troubles de l'humeur et de la toxicomanie: combinaison de preuves précliniques et d'études de tomographie par émission de positrons (PET) humaine. De face. Neurosci. 9: 86. doi: 10.3389 / fnins.2015.00086
Reçu: 07 Janvier 2015; Accepté: 27 février 2015;
Publié: 18 March 2015.
Édité par:
Ashok Kumar, Université de Floride, États-Unis
Commenté par:
Karthik Bodhinathan, Institut de recherche médicale Sanford Burnham, États-Unis
Amber M. Muehlmann, Université de Floride, États-Unis
Droits d'auteur © 2015 Terbeck, Akkus, Chesterman et Hasler. Ceci est un article en accès libre distribué selon les termes de la Licence d'attribution Creative Commons (CC BY). L'utilisation, la distribution ou la reproduction sur d'autres forums est autorisée, à condition que l'auteur original ou le donneur de licence soit crédité et que la publication originale de ce journal soit citée conformément à la pratique académique reconnue. Aucune utilisation, distribution ou reproduction n’est autorisée si elle n’est pas conforme à ces conditions.
* Correspondance: Sylvia Terbeck, École de psychologie, Faculté des sciences de la santé et des sciences humaines, Université de Plymouth, PL4 8AA, Plymouth, Royaume-Uni. [email protected]