- 1Centro Interdisciplinar de Neurociências de Valparaíso, Faculdade de Ciências, Universidade de Valparaíso, Valparaíso, Chile
- 2Departamento de Neurociência, Faculdade de Medicina, Universidade do Chile, Santiago, Chile
- 3Núcleo Milenio Biologia NUMIND de Distúrbios Neuropsiquiátricos, Universidade de Valparaíso, Valparaíso, Chile
- 4Departamento de Biologia Celular e Molecular, Faculdade de Ciências Biológicas, Pontifícia Universidade Católica do Chile, Santiago, Chile
- 5Departamento de Farmácia e Centro Interdisciplinar de Neurociência, Faculdade de Química, Pontifícia Universidade Católica do Chile, Santiago, Chile
A força dos comportamentos orientados a objetivos é regulada pelos neurônios da dopamina no mesencéfalo. Disfunções dos circuitos dopaminérgicos são observadas na dependência de drogas e no transtorno obsessivo-compulsivo. O comportamento compulsivo é uma característica que os dois distúrbios compartilham, que está associada a uma neurotransmissão de dopamina aumentada. A atividade dos neurônios da dopamina no mesencéfalo é regulada principalmente pela ação homeostática da dopamina através dos receptores D2 (D2R) que diminuem o disparo dos neurônios, bem como a síntese e liberação de dopamina. A transmissão de dopamina também é regulada por sistemas heterotransmissores de neurotransmissores, como o sistema kappa opioid, entre outros. Grande parte do nosso conhecimento atual do sistema kappa opioide e sua influência na transmissão da dopamina vem de modelos animais pré-clínicos de doenças cerebrais. Em 1988, usando microdiálise cerebral, foi demonstrado que a ativação aguda dos Receptores Opióides Kappa (KOR) diminui os níveis sinápticos de dopamina no estriado. Esse efeito inibitório do KOR se opõe à influência facilitadora dos medicamentos de abuso na liberação de dopamina, levando à proposição do uso de agonistas do KOR como terapia farmacológica para a ingestão compulsiva de medicamentos. Surpreendentemente, 30 anos depois, os antagonistas da KOR são propostos para tratar a dependência de drogas. O que pode ter acontecido durante esses anos que gerou essa mudança drástica de paradigma? As evidências coletadas sugerem que o efeito do KOR nos níveis de dopamina sináptica é complexo, dependendo da frequência da ativação do KOR e do momento com outros estímulos recebidos pelos neurônios da dopamina, bem como das diferenças de sexo e espécie. Inversamente ao seu efeito agudo, a ativação crônica da KOR parece facilitar a neurotransmissão da dopamina e os comportamentos mediados pela dopamina. As ações opostas exercidas pela ativação aguda versus crônica da KOR têm sido associadas a um efeito aversivo inicial e recompensador tardio, durante a exposição a drogas de abuso. Os comportamentos compulsivos induzidos pela ativação repetida do D2R também são potencializados pela co-ativação sustentada do KOR, que se correlaciona com os níveis sinápticos diminuídos de dopamina e D2R sensibilizado. Assim, a ativação dependente do tempo do KOR afeta diretamente os níveis de dopamina, afetando o ajuste de comportamentos motivados. Esta revisão analisa a contribuição do sistema kappa opioide para os correlatos dopaminérgicos de comportamentos compulsivos.
Introdução
Sistema Dopaminérgico em Comportamentos Compulsivos
Compulsão é a impossibilidade de parar automaticamente para executar uma ação habitual com resultado conhecido, apesar das consequências adversas (Robbins e outros, 2012) Comportamentos compulsivos são características do transtorno obsessivo-compulsivo (TOC) e da dependência de drogas, entre outras doenças psiquiátricas. O comportamento de verificação é muito comum nos distúrbios do espectro obsessivo-compulsivo, sendo caracterizado pela repetição constante de uma determinada rotina, de maneira estereotipada ou ritualística (Williams et al., 2013) Uma ampla gama de comportamentos normais (por exemplo, verificação, limpeza, lavagem das mãos etc.) pode se tornar compulsiva em pacientes com TOC e, em geral, surge em resposta a pensamentos obsessivos e angustiantes que induzem ansiedade. Da mesma forma, procurar e consumir drogas de abuso torna-se compulsivo em viciados em drogas Como no TOC, a ansiedade desempenha um papel fundamental no desencadeamento do consumo compulsivo de drogas em usuários experientes de drogas. As mesmas deficiências no processamento de recompensas e punições são observadas em ambas as condições (Figee e outros, 2016), o que levou alguns autores a discutir o TOC como um vício comportamental (Holden, 2001).
Um possível mecanismo que leva ao comportamento compulsivo está enquadrado na teoria do vício em sensibilização por incentivo, que é que uma motivação ampliada ("querer") para a droga se desenvolve durante o vício sem desenvolver um efeito prazeroso ampliado ("gostar") (Berridge et al., 1989; Berridge e Robinson, 2016) A sensibilização duradoura do circuito de recompensa / motivação está envolvida na indução de sensibilização a incentivos associada à procura de drogas. O circuito de recompensa / motivação é composto por neurônios da dopamina no mesencéfalo substantia nigra (SN) e área tegmentar ventral (ATV), que têm como alvo as camadas dorsal e ventral do estriado, respectivamente. Os neurônios da dopamina que se projetam no estriado ventral ou no núcleo accumbens (NAc) têm sido tradicionalmente relacionados a comportamentos orientados a objetivos, enquanto os neurônios da dopamina que se projetam no estriado dorsal têm sido associados à aquisição de hábitos (Everitt e Robbins, 2005; Sábio, 2009; Yager et al., 2015; Volkow et al., 2017).
A sensibilização do circuito de recompensa / motivação é observada em roedores como o aumento gradual da atividade locomotora induzida pela administração repetida de uma dose fixa de medicamento potencialmente viciante (Pierce e Kalivas, 1997; Robinson e Berridge, 2001) A sensibilização locomotora é um fenômeno suportável, pois é observável após semanas, meses e até um ano após a retirada do medicamento (Robinson e Berridge, 1993) Sugeriu-se cedo que a sensibilização do circuito de recompensa / motivação contribui para a busca compulsiva de drogas (Robinson e Berridge, 1993) Consequentemente, a sensibilização locomotora facilita a auto-administração de cocaína em busca de reintegração (De Vries e outros, 2002) Além disso, ratos com acesso prolongado à autoadministração de cocaína apresentam maior resposta locomotora à cocaína do que ratos com acesso limitado (Ferrario et al., 2005) Além disso, as alterações neuroquímicas subjacentes à sensibilização locomotora a psicoestimulantes também são observadas na busca compulsiva de drogas (Steketee e Kalivas, 2011; Giuliano et al., 2019) Esses dados apóiam a correspondência proposta anteriormente entre sensibilização locomotora e busca compulsiva de drogas observada em humanos (Robinson e Berridge, 1993; Vanderschuren e Kalivas, 2000) Mecanisticamente, a administração repetida de drogas de abuso sensibiliza os circuitos mesolímbicos de dopamina, aumentando a neurotransmissão dopaminérgica. Os psicoestimulantes, como cocaína ou anfetaminas, que bloqueiam o transportador de dopamina da membrana plasmática (DAT), induzem um grande aumento de dopamina no espaço sináptico no estriado e NAc, ativando a locomoção (Steketee e Kalivas, 2011) Como na toxicodependência, a sensibilização do circuito de recompensa / motivação da dopamina contribui para comportamentos compulsivos observados no TOC. De fato, a ativação repetida dos receptores da dopamina D2 (D2Rs) é suficiente para induzir a sensibilização locomotora e verificar o comportamento em ratos e camundongos (Szechtman e outros, 1998; Szechtman e outros, 1999; Sun et al., 2019) A administração repetida de quinpirol, um agonista D2R / D3R, é um modelo aceito de TOC, uma vez que recapitula a validade facial, através de um incremento de verificação compulsiva e comportamento estereotipado, validade preditiva, como observado por uma diminuição de comportamentos compulsivos após tratamento crônico com recaptação de serotonina inibidores (SRI) e validade de construto, uma vez que as estruturas cerebrais envolvidas neste modelo são compartilhadas com as da patologia (Stuchlik et al., 2016; Szechtman e outros, 2017). Em resumo, a ativação repetida da transmissão de dopamina, seja pelos mecanismos pré-sináptico (liberação de dopamina) ou pós-sináptico (ativação de D2R), leva à sensibilização locomotora e comportamentos compulsivos.
O sistema kappa opioid é um dos sistemas mais preponderantes que controlam a transmissão de dopamina no circuito de recompensa / motivação. As evidências mostram que a transmissão kappa-opióide se opõe aos efeitos da dopamina; a ativação aguda de receptores kappa opioides (KORs) neutraliza a atividade locomotora induzida por psicoestimulantes (Gray et al., 1999) Por outro lado, a ativação repetida do KOR mantém e aprimora a busca compulsiva e habitual de medicamentos (Koob, 2013) O consumo de drogas de abuso induz uma transmissão kappa opióide homeostática aumentada, provavelmente contribuindo para os estados emocionais negativos da disforia (Koob, 2013) desencadeando o uso compulsivo de drogas (Chavkin e Koob, 2016) De fato, o bloqueio do KOR impediu o restabelecimento da nicotina por estresse, mas não induzido por drogas (Jackson et al., 2013), cocaína (Beardsley et al., 2005) e etanol (Sperling e outros, 2010) De acordo com esse achado, o bloqueio da KOR reverte alterações dopaminérgicas no estriado dorsolateral de ratos sensibilizados com anfetaminas, sem modificar sua resposta locomotora aprimorada à droga (Azocar et al., 2019). Assim, o sistema KOR parece melhorar o reforço negativo, aumentando o valor da droga. No TOC, o reforço negativo é desencadeado por obsessões, que fortalecem uma determinada compulsão para evitar essa obsessão. Embora não tenha sido testado diretamente, o reforço negativo pode desempenhar um papel na sensibilização ao quinpirol. De fato, o D2R está envolvido na geração de reforço negativo. Por exemplo, evitar locais para áreas pareadas com retirada de morfina não foi desenvolvido em camundongos sem a longa isoforma de D2R (Smith et al., 2002) e o tratamento repetido com quinpirol durante o período de abstinência restabelece a busca de cocaína e heroína em um paradigma de auto-administração, um efeito relacionado à locomoção sensibilizada ao quinpirol (De Vries e outros, 2002), sugerindo mecanismos compartilhados entre psicoestimulantes e sensibilização induzida por quinpirol. Além disso, a introdução da gaiola doméstica, mas não uma gaiola nova, na arena de campo aberto reduz a sensibilização locomotora e o comportamento de verificação compulsiva (Szechtman e outros, 2001), indicando que as dicas de segurança / familiares podem competir com as sugestões ambientais negativas que favorecem a sensibilização. Da mesma forma que a sensibilização induzida por psicoestimulante, a ativação repetida do KOR facilita a sensibilização locomotora (Escobar e outros, 2017) e comportamento de verificação compulsiva (Perreault et al., 2007) induzida pela administração repetida de quinpirol. Ainda não se sabe se esse efeito potencializador é uma conseqüência do reforço negativo reforçado.
A análise minuciosa realizada recentemente mostra que o efeito do sistema kappa-opióide na transmissão dopaminérgica é complexo: depende da via da dopamina envolvida (Margolis e outros, 2006; Margolis e outros, 2008) e no momento entre a ativação do receptor KOR e a ativação do receptor de dopamina (Chartoff et al., 2016) Consistente com essa complexidade, o potencial uso terapêutico de ligantes KOR tem sido amplamente discutido. Foi proposto que o agonista de KOR pode ser clinicamente útil durante a fase de uso de drogas, atenuando a hiperdopaminergia induzida por drogas (Shippenberg et al., 2007) Por outro lado, um antagonista da KOR pode ser útil no tratamento da síndrome de abstinência induzida por um aumento na expressão da dinorfina após o consumo repetido de drogas (Pequenino e koob, 2010) Por conseguinte, foi proposto que o agonista parcial do KOR (Béguin et al., 2012) poderia ser uma opção terapêutica para tratar a ingestão compulsiva de drogas e os sintomas de abstinência em indivíduos dependentes (Chartoff et al., 2016; Callaghan et al., 2018) Nesta revisão, analisamos a modulação dependente do tempo / contexto de correlatos dopaminérgicos de sensibilização comportamental e compulsividade.
Crosstalk Anatômico e Funcional entre os Sistemas Opioide Kappa e Dopaminérgico nas Regiões Striatal e Midbrain
Regiões estriatais
Os KORs são receptores acoplados à proteína Gi / o altamente expressos no sistema de dopamina no mesencéfalo (Mansour e outros, 1996) Esses receptores pertencem à família de receptores opióides compostos por mu (MOR), delta (DOR) e kappa (KORs). Os agonistas endógenos para esses receptores são endorfinas, encefalina e dinorfina, respectivamente. No estriado, a dinorfina é sintetizada por neurônios de tamanho médio (MSNs) contendo dopamina D1receptor (D1R) que possuem axônios recorrentes ativando KORs do mesmo núcleo (Mansour e outros, 1995) Imagens de microscopia eletrônica de NAc de rato mostram que os KORs são encontrados predominantemente em estruturas pré-sinápticas contendo DAT, enquanto uma proporção menor de KORs localiza-se em dendritos em oposição ao DAT (Svingos et al., 2001; Kivell e outros, 2014) Estudos imunofluorescentes que caracterizam preparações pré-sinápticos-sinaptossomais de NAc mostram que KORs e D2Rs coexistem preferencialmente em sinaptossomas contendo a enzima sintetizadora de dopamina, tirosina hidroxilase (TH) (Escobar e outros, 2017) Além disso, os KORs são abundantes nos corpos celulares do NAc e do estriado e colocalizam-se com os D2Rs em uma subpopulação celular (Escobar e outros, 2017) Com informações genéticas e moleculares, sugeriu-se que 20% da ligação total de KOR no estriado é observada nos terminais DA (Van't Veer et al., 2013). Além disso, Tejeda et ai. (2017) mostrou que os MSNs D1R e D2R expressam KOR com uma preferência maior por D1R contendo MSNs (Tejeda et al., 2017) Esses dados anatômicos indicam que os KORs estão presentes pré e pós-sinapticamente, regulando a neurotransmissão de dopamina no circuito de recompensa / motivação.
Várias abordagens experimentais mostram que a ativação de KORs inibe a liberação de dopaminae A ativação aguda de KORs por injeção sistêmica ou infusão local de agonistas diminui os níveis extracelulares de dopamina no NAc (Di Chiara e Imperato, 1988; Spanagel et al., 1992; Fuentealba et al., 2006) e estriado dorsal (Gehrke e outros, 2008) Apoiando uma ação inibidora tônica dos KORs sobre a neurotransmissão da dopamina, a infusão direta do antagonista seletivo e duradouro da KOR nor-binaltorphimine (nor-BNI) (Broadbear e outros, 1994) aumenta os níveis basais de dopamina no NAc (Spanagel et al., 1992) e liberação de dopamina no estriado dorsal (Azocar et al., 2019) Evidências finais da inibição tônica de KOR da dopamina foram demonstradas em camundongos knockout para KOR, que exibiram níveis extracelulares aumentados de dopamina no estriado e NAc (Chefer et al., 2005) Os mecanismos responsáveis pela inibição de KOR da liberação de dopamina não estão completamente elucidados. Porém, é bem sabido que a ativação dos KORs leva ao aumento do K + e diminuição das condutâncias do Ca2 +, induzindo assim a hiperpolarização celular e o bloqueio da liberação do neurotransmissor vesicular (Bruchas e Chavkin, 2010; Margolis e Karkhanis, 2019).
Adicionalmente, em vitro e in vivo dados funcionais sugerem que os KORs modificam os níveis extracelulares de dopamina modulando a atividade do DAT. Por exemplo, a ativação de KORs em células EM4 que co-expressam KORs e DAT, leva a um aumento da captação de dopamina medida por voltametria (Kivell e outros, 2014). A ex vivo A análise também usando voltametria em tecidos desagregados mostrou que uma injeção sistêmica do agonista KOR U-69593 aumentou a captação de dopamina no NAc (Thompson et al., 2000) Um artigo recente semelhante mostra que o nor-BNI bloqueia o aumento da captação de dopamina no estriado ventral e dorsal, induzido por uma injeção sistêmica aguda de MP1104, um agonista misto dos receptores opióides Kappa / Delta (Atigari et al., 2019) No entanto, o efeito da ativação de KOR na captação de dopamina ainda não foi totalmente elucidado. A administração sistêmica do agonista parcial KOR nalmefene diminuiu a dose de captação de dopamina no estriado de forma dependente, quantificada por voltametria cíclica de varredura rápida (FSCV) (Rose et al., 2016) Usando uma microdiálise sem fluxo líquido em ratos machos adultos, o bloqueio da KOR foi acompanhado por um aumento na fração de extração (Ed), que é uma medida indireta da captação de dopamina (Chefer et al., 2006; Azocar et al., 2019), sugerindo que a ativação tônica do KOR exerce um controle inibitório sobre a atividade do DAT (captação de dopamina). Esses resultados destacam o papel complexo da atividade endógena da KOR na captação de dopamina no controle dos níveis extracelulares da dopamina. Abordagens de resolução temporal mais alta, como o FSCV, falharam em mostrar um efeito do KOR na captação de dopamina (Ebner e outros, 2010; Ehrich e outros, 2015; Hoffman et al., 2016), sugerindo que a atividade DAT de aprimoramento de KOR nas regiões estriatais precisa de um período de incubação., o aprimoramento mediado por KOR da atividade de DAT pode ser explicado por um aumento no número de DAT nas membranas celulares induzida pela ativação de KOR, conforme relatado em sinaptossomas estriados e células linhas (Kivell e outros, 2014).
Regiões do mesencéfalo
Os ensaios autorradiográficos realizados no cérebro médio de ratos mostram ligação significativa para os KORs no eixo rostrocaudal do SN e VTA (Speciale et al., 1993) Por outro lado, os dados da microscopia eletrônica mostram que os terminais contendo dinorfina são sinapses diretamente nos dendritos positivos para TH no SN e no VTA (Sesack e Pickel, 1992), sugerindo que os KORs se localizam nos compartimentos somatodendríticos dos neurônios da dopamina. Os MSNs estriatais contendo D1R são uma das entradas de dinorfina nos neurônios da dopamina no mesencéfalo. Curiosamente, o bloqueio dos KORs não modifica o efeito inibitório dos D1R-MSNs para os neurônios da dopamina VTA, indicando que essa inibição é mediada pelo GABA (Edwards e outros, 2017) Os KORs modulam respostas somatodendríticas dos neurônios da dopamina no mesencéfalo. Estudos eletrofisiológicos mostram que a ativação de KORs no ATV hiperpolariza e diminui a taxa de disparo espontâneo dos neurônios da dopamina (Margolis e outros, 2003) Consequentemente, a infusão de agonistas da KOR diminui o efluxo somatodendrítico da dopamina (Smith et al., 1992; Dalman e O'Malley, 1999) No entanto, esse efeito inibitório dos KORs nos neurônios da dopamina parece ser dependente do circuito. A infusão de agonistas kappa-opióides no ATV diminui a liberação de dopamina no córtex pré-frontal medial (mPFC) (Margolis e outros, 2006) mas não no NAc (Devine e outros, 1993; Margolis e outros, 2006). Além disso, Margolis et ai. (2006) descobriram que os KORs inibem os neurônios da dopamina VTA que se projetam para o mPFC e a amígdala basolateral, mas não aqueles que se projetam para o NAc. Nesse mesmo ano, Ford et ai. (2006) mostraram que a aplicação de agonistas de KOR no banho em cortes de ATV de camundongo induzia uma corrente externa mais alta nos neurônios da dopamina que se projetam para o NAc em comparação com aqueles que se projetam para a amígdala basolateral, indicando que os KORs exercem uma maior inibição dos neurônios de dopamina que se projetam para o NAc do que para a amígdala. Além disso, a ativação do KOR diminui a amplitude de excitação (Margolis e outros, 2005) e inibitórios (Ford et al., 2007) correntes pós-sinápticas nos neurônios da dopamina no mesencéfalo. As diferenças entre as espécies e as projeções eferentes complexas de ATV para mPFC e NAc (Van Bockstaele e Pickel, 1995; Carr e Sesack, 2000) tornam difícil estabelecer se os KORs inibem seletivamente algumas das populações de dopamina neuronal na ATV. No entanto, os dados resumidos aqui indicam que os KORs estão no somatório e nos terminais dos neurônios da dopamina, bem como nas entradas que os regulam, assim posicionados de maneira requintada para controlar a atividade sináptica dos neurônios da dopamina no mesencéfalo.
Papel das KORs no controle da neurotransmissão da dopamina na sensibilização induzida por psicoestimulantes e comportamentos compulsivos
A dependência de drogas é um processo que envolve a busca inicial de drogas impulsivas associada aos seus efeitos de reforço positivo. Por outro lado, a compulsividade é um traço de personalidade observável em viciados em drogas. Várias neuroadaptações nas vias dopaminérgicas foram propostas para explicar a busca e a ingestão compulsivas de drogas após a exposição repetida a drogas de abuso (Everitt e Robbins, 2005; Koob e Volkow, 2016) Uma das hipóteses propostas que impulsionam a ingestão compulsiva de drogas é a sensibilização de seus efeitos negativos de reforço (Koob, 2013) O controle inibitório do sistema kappa opioide na liberação de dopamina pode contribuir para as propriedades de reforço negativo das drogas de abuso. No entanto, as consequências da ativação do KOR na neurotransmissão de dopamina e na busca compulsiva de drogas parecem ser complexas e aparentemente contraditórias. De fato, a liberação de dopamina induzida por anfetamina e cocaína é atenuada pela administração concomitante de agonistas do KOR (Heidbreder e Shippenberg, 1994; Maisonneuve et al., 1994; Thompson et al., 2000) e até diminuem a auto-administração de cocaína (Negus e outros, 1997) Além disso, os KORs exercem um feedback inibitório sobre a liberação de dopamina da via mesolímbica em resposta à ativação sustentada do D1R pós-sináptico, como ocorre com a exposição repetida a psicoestimulantes (Cole et al., 1995; Nestler, 2001) Paradoxalmente, a ativação dos KORs também pode facilitar a liberação de dopamina no caminho da recompensa / motivação (Fuentealba et al., 2006; Fuentealba et al., 2007) e consumo de psicoestimulantes (Wee et al., 2009). Fuentealba et ai. (2007) mostraram que após quatro dias a administração de U69593, um agonista do KOR, aumentou a liberação de dopamina induzida por anfetamina no NAc. Recentemente, foi demonstrado que o bloqueio dos KORs reverte as alterações na liberação e captação de dopamina no estriado dorsal que ocorrem durante a sensibilização locomotora induzida pela anfetamina (Azocar et al., 2019) No total, esses dados sugerem que a ativação dos KORs também pode contribuir para propriedades de reforço positivo da droga de abuso (Chartoff et al., 2016).
Além disso, a ativação dos KORs também parece contribuir para a busca compulsiva de drogas; O bloqueio dos KORs reduz a cocaína (Wee et al., 2009), heroína (Schlosburg et al., 2013) e metanfetamina (Whitfield e outros, 2015) ingestão em ratos com acesso ilimitado ao medicamento (Wee et al., 2009) Esse efeito também é evidenciado na busca por drogas induzidas pelo estresse. Por exemplo, os ratos KOR knockout não mostraram preferência por cocaína após estresse forçado na natação (McLaughlin e outros, 2006a) O bloqueio de KORs atenua a preferência do local da nicotina induzida pela exposição forçada ao estresse no nado (Smith et al., 2012) Curiosamente, o bloqueio do KOR atenua a busca de cocaína e nicotina induzida pelo estresse, mas não afetou a busca induzida por um desafio às drogas (Beardsley et al., 2005; Jackson et al., 2013) O efeito KOR do facilitador induzido pelo estresse parece ser mediado pelo circuito de recompensa / motivação (Shippenberg et al., 2007; Pequenino e koob, 2010) Em um elegante estudo realizado pela Dra. Kauer e seu grupo, foi demonstrado que o bloqueio de KORs na ATV, anteriormente ou após um estresse agudo, inibe o restabelecimento da busca pela cocaína, um efeito associado ao resgate de pacientes a longo prazo. potencialização de sinapses inibitórias nos neurônios da dopamina (Graziane e outros, 2013; Polter e outros, 2014).
A facilitação da ingestão de psicoestimulantes exercida pelos KORs parece depender de uma janela de tempo em relação à exposição a drogas. A administração do agonista KOR U50488 1 h antes da exposição à cocaína potencializa a preferência pelo local da cocaína e a liberação relativa de dopamina evocada pela cocaína no NAc, enquanto os efeitos opostos são observados quando administrados 15 minutos antes (McLaughlin e outros, 2006a; Ehrich e outros, 2014) Usando auto-estimulação intracraniana Chartoff et ai. (2016) observaram que o agonista KOR Salvinorin A tem um efeito inicial aversivo e recompensador retardado, acompanhado por uma diminuição e um aumento na liberação estimulada de dopamina na NAc, respectivamente. Juntos, esses dados indicam um efeito dependente do tempo da ativação do KOR nas propriedades recompensadoras da cocaína e apontam para a ativação do KOR mediada pelo estresse como um elemento-chave para o desenvolvimento da busca compulsiva por drogas.
Sensibilização Locomotora Induzida por Quinpirol e Comportamento Compulsivo
Os fatos de que o sistema dopamina está envolvido na geração de sensibilização e compulsividade são fortalecidos pelo comportamento observado em roedores tratados com o agonista D2R, quinpirol. Resumidamente, os D2Rs são receptores acoplados a Gi amplamente expressos no circuito de recompensa / motivação; são expressos somatodendriticamente e nos terminais axonais dos neurônios da dopamina (Sesack et al., 1994) e sua ativação diminui os níveis extracelulares de dopamina (Imperato e Di Chiara, 1988) No estriado, os D2Rs também estão localizados pós-sinapticamente em neurônios espinhosos médios (Sesack et al., 1994) e sua ativação inibe a via indireta, permitindo a atividade locomotora.
O Dr. Henry Szechtman começou a estudar os efeitos do quinpirol no comportamento de ratos no final da década de 1980. Suas descobertas iniciais mostraram que a administração aguda de quinpirol tem efeito dose-dependente sobre a atividade locomotora. Em doses baixas (0.03 mg / kg), diminui a atividade locomotora, enquanto em doses mais altas (> 0.5 mg / kg), aumenta. (Eilam e Szechtman, 1989) Esses efeitos estão associados à ativação de D2Rs pré-sinápticos de alta afinidade e D2Rs pós-sinápticos de baixa afinidade, respectivamente (Usiello et al., 2000) Inesperadamente, a administração repetida (quinzenalmente) de quinpirol induz um aumento gradual e sustentado da locomoção, semelhante à sensibilização locomotora induzida por psicoestimulantes (Szechtman e outros, 1993; Szechtman e outros, 1994) Demonstrou-se que o efeito de sensibilização locomotora depende de D2Rs, uma vez que camundongos deficientes para esse receptor não desenvolvem sensibilização locomotora ao quinpirol (Escobar e outros, 2015).
No início da década de 90, Szechtman e Eilam relataram que, junto com a sensibilização locomotora, os ratos desenvolveram um comportamento estereotipado, que é reforçado a cada administração de quinpirol (Eilam e Szechtman, 1989; Szechtman e outros, 1993) Hoje, a administração repetida de quinpirol é um modelo validado para TOC (Szechtman e outros, 1999; Szechtman e outros, 2001; Eilam e Szechtman, 2005; Stuchlik et al., 2016; Szechtman e outros, 2017), com base na observação de que o comportamento de ratos se torna cada vez mais estruturado e inflexível, remanescente do comportamento ritual característico do comportamento de verificação compulsiva (Szechtman e outros, 1998; Szechtman e outros, 2017) Estudos recentes mostram que o quinpirol repetido também induz comportamentos compulsivos em camundongos, como verificação compulsiva (Sun et al., 2019), inflexibilidade comportamental e mastigação compulsiva (Asaoka et al., 2019), este último revertido pelo bloqueio de D2Rs no estriado, apoiando ainda mais a ativação repetida de D2Rs para induzir comportamentos compulsivos. Juntos, os dados apontam para um papel crucial dos D2Rs nas vias da dopamina no mesencéfalo para induzir sensibilização e compulsão locomotoras. A administração repetida de quinpirol inicia o comportamento estereotipado induzido pela cocaína (Thompson et al., 2010) e os efeitos locomotores da anfetamina (Cope e outros, 2010), reforçando a ideia de que a ativação de D2Rs está subjacente à sensibilização induzida por psicoestimulantes e sugerindo um mecanismo compartilhado entre quinpirol e sensibilização induzida por psicoestimulantes. Curiosamente, o efeito sensibilizante da ativação repetida de D2Rs parece ser mais forte do que o induzido por psicoestimulantes, uma vez que todos os ratos tratados com quinpirol desenvolvem sensibilização locomotora (Escobar e outros, 2015), enquanto cerca de sessenta por cento dos ratos sensibilizam com anfetamina (Escobar e outros, 2012; Casanova e outros, 2013).
A sensibilização comportamental induzida pela ativação repetida de D2Rs é acompanhada de adaptações no circuito de recompensa / motivação. Os ratos sensibilizados com quinpirol apresentam menor tônus dopaminérgico no NAc, observado como diminuição basal (Koeltzow e outros, 2003) e estimulou a liberação de dopamina tônica e fásica (Escobar e outros, 2015), indicando diminuição da capacidade de liberação de dopamina do circuito mesencéfalo da dopamina. Os níveis de dopamina sináptica no NAc são controlados pela atividade dos neurônios DAT e dopamina (Goto e Grace, 2008), qual in vivo consiste em disparos tônicos e de rajada (Wilson et al., 1977; Grace e Bunney, 1980) Relatórios anteriores mostram que ratos sensibilizados a quinpirol exibem um número menor de neurônios dopaminérgicos em disparos tônicos e de rajada no ATV (Sésia et al., 2013) Juntos, esses dados indicam que a diminuição na liberação de dopamina observada após a sensibilização ao quinpirol é resultado de uma diminuição na atividade geral dos neurônios da dopamina. O comportamento compulsivo e a atividade locomotora sensibilizada induzida pelo tratamento repetido com quinpirol podem ser consequência da sensibilização dos D2Rs, devido à diminuição do tônus dopaminérgico no NAc. De fato, ratos sensibilizados a quinpirol mostram um aumento na ligação da dopamina D2R (Culver e outros, 2008) e um aumento no estado de afinidade desses receptores (Perreault et al., 2007), apoiando esta hipótese.
Interações KOR-Dopamina em Comportamentos Compulsivos Induzidos por Quinpirol
Os estudos iniciais sobre o papel da KOR em comportamentos compulsivos induzidos por D2R também vieram do laboratório de Szechtman. Este grupo examinou a administração concomitante do agonista KOR U69593 com quinpirol na atividade locomotora. Especificamente, os autores administraram injeções subcutâneas a ratos com uma mistura de U69593 e quinpirol, até que 8 a 10 injeções fossem concluídas. Ao contrário do efeito hipolocomotor do U69593 sozinho, foi observada hiperlocomoção quando administrado concomitantemente com doses baixas (pré-sinápticas) e altas (pós-sinápticas) de quinpirol. U69593 mudou o efeito hipolocomotor de uma dose pré-sináptica de quinpirol para hiperlocomoção e aumentou o efeito hiperlocomotor de uma dose pós-sináptica de quinpirol (Perreault et al., 2006) A co-ativação de KORs também acelerou a indução da sensibilização locomotora e potencializou o efeito da ativação de D2Rs, uma vez que a locomoção máxima alcançada pelo duplo tratamento duplica o efeito locomotor induzido apenas pelo quinpirol (Perreault et al., 2006; Escobar e outros, 2017) A co-ativação de KORs também acelera a aquisição do comportamento de verificação compulsiva (Perreault et al., 2007) Esses efeitos potenciadores dos KORs nos comportamentos induzidos por quinpirol requerem ativação repetida dos KORs. De fato, a injeção aguda do agonista KOR U69593 não modificou ainda mais a atividade locomotora em ratos sensibilizados com quinpirol (Escobar e outros, 2017) O mecanismo de KOR potencializando a sensibilização induzida por D2R é desconhecido. Uma possibilidade é que o próprio sistema opióide kappa endógeno esteja mediando a sensibilização dependente de D2R. No entanto, essa possibilidade foi descartada, mostrando que a pré-administração de norBNI não modificou a sensibilização locomotora ao quinpirol, sugerindo que a dinorfina não é liberada na ativação D2R a jusante (Escobar e outros, 2017) Esses dados não descartam que a dinorfina possa ter um papel importante na sensibilização de comportamentos compulsivos, por exemplo, o estresse induz a liberação de dinorfina e a ativação de KORs, o que facilita comportamentos compulsivos (McLaughlin e outros, 2003; McLaughlin e outros, 2006a; McLaughlin e outros, 2006b).
A diafonia entre D2Rs e KORs é complexa e parece depender se a ativação de ambos os receptores é coincidente ou separada temporalmente. Dados anatômicos indicam que a diafonia entre D2Rs e KORs pode ocorrer de forma pré-sináptica nos axônios e na soma dos neurônios da dopamina, bem como pós-sinapticamente nos MSNs do estriado. Embora não descarte um papel para os KORs localizados em axônios de outros sistemas neuroquímicos, os dados anatômicos apontam fortemente para um papel direto dos KORs que regulam os D2Rs. Aguda ou repetida, a ativação dos KORs diminui a função inibidora dos D2Rs nos neurônios da dopamina. Estudos de eletrofisiologia mostraram que a ativação aguda de KOR nos neurônios da dopamina do ATV e SN inibe a corrente pós-sináptica inibidora mediada por D2R, um efeito mediado pelos mecanismos pré e pós-sinápticos, pois a KOR diminui a liberação de dopamina e a dinorfina bloqueia o efeito inibitório da dopamina aplicada no banho (Ford et al., 2007) Estudos neuroquímicos mostraram que a ativação repetida de KORs bloqueia a inibição induzida por D2R da liberação de dopamina no NAc (Fuentealba et al., 2006) Além disso, a ativação aguda coincidente de D2Rs e KORs diminui a inibição da liberação de dopamina no NAc em comparação com o efeito de cada receptor sozinho (Escobar e outros, 2017) Assim, os KORs pré-sinápticos não agem de maneira aditiva ou em sinergia com os D2Rs pré-sinápticos; por outro lado, os KORs inibem ou ocluem o efeito inibitório de D2R. Esse mecanismo poderia explicar o efeito de ativação locomotora de uma dose aguda de agonistas da KOR concomitante a uma dose baixa de quinpirol (Perreault et al., 2006).
Um estudo recente mostra que a ativação do KOR no ATV medeia o comportamento compulsivo medido como inibição comportamental e enterramento de mármore (Abraão et al., 2017), reforçando a ideia de que a ativação dos KORs é realmente um gatilho para a compulsividade. Dados publicados por Margolis et ai. (2006; 2008) indicam que a interação KORs e D2Rs deve ocorrer nos neurônios da dopamina direcionados ao mPFC (Margolis e outros, 2006; Margolis e outros, 2008) A despeito de, Ford et ai. (2006; 2007) descobriram que a inibição de KORs de IPSC mediada por D2R ocorre em neurônios de dopamina direcionados ao NAc (Ford et al., 2006; Ford et al., 2007) Juntos, esses dados mostram que a interação KOR com D2R no compartimento somatodendrítico dos neurônios da dopamina pode surgir como resultado de uma diafonia no mesmo neurônio da dopamina. Se isso acontece nas projeções mesolímbica ou mesocortical ainda é controverso.
Notavelmente, o KOR foi encontrado nos MSNs do NAc (Escobar e outros, 2017; Tejeda et al., 2017), indicando que a potencialização do comportamento compulsivo induzido por D2R também pode surgir por ações diretas nas células-alvo dos neurônios da dopamina. A esse respeito, vale ressaltar que a administração repetida de U69593 aumenta a quantidade de D2Rs no estado de alta afinidade (Perreault et al., 2007) Dados neuroquímicos indicam que níveis extracelulares diminuídos de dopamina estão associados à sensibilização de D2Rs. A co-ativação de KORs não diminui ainda mais os níveis extracelulares de dopamina no NAc já diminuídos pela ativação repetida de D2Rs (Escobar e outros, 2017), descartando um papel para os KORs pré-sinápticos acelerando ou potencializando a sensibilização de D2Rs na NAc por meio desse mecanismo. Portanto, os KORs desencadeiam mecanismos moleculares lentos que sensibilizam ainda mais os efeitos neuroquímicos e comportamentais dos D2Rs, sugerindo que o aprimoramento da sensibilização da locomoção pode ser devido a um efeito pós-sináptico adaptativo e não pré-sináptico. A este respeito, a ativação repetida dos KORs pode desencadear a inibição da via estriada indireta D2R, que alterna o equilíbrio D1R / D2R para D1R, induzindo a compulsividade (Figura 1).
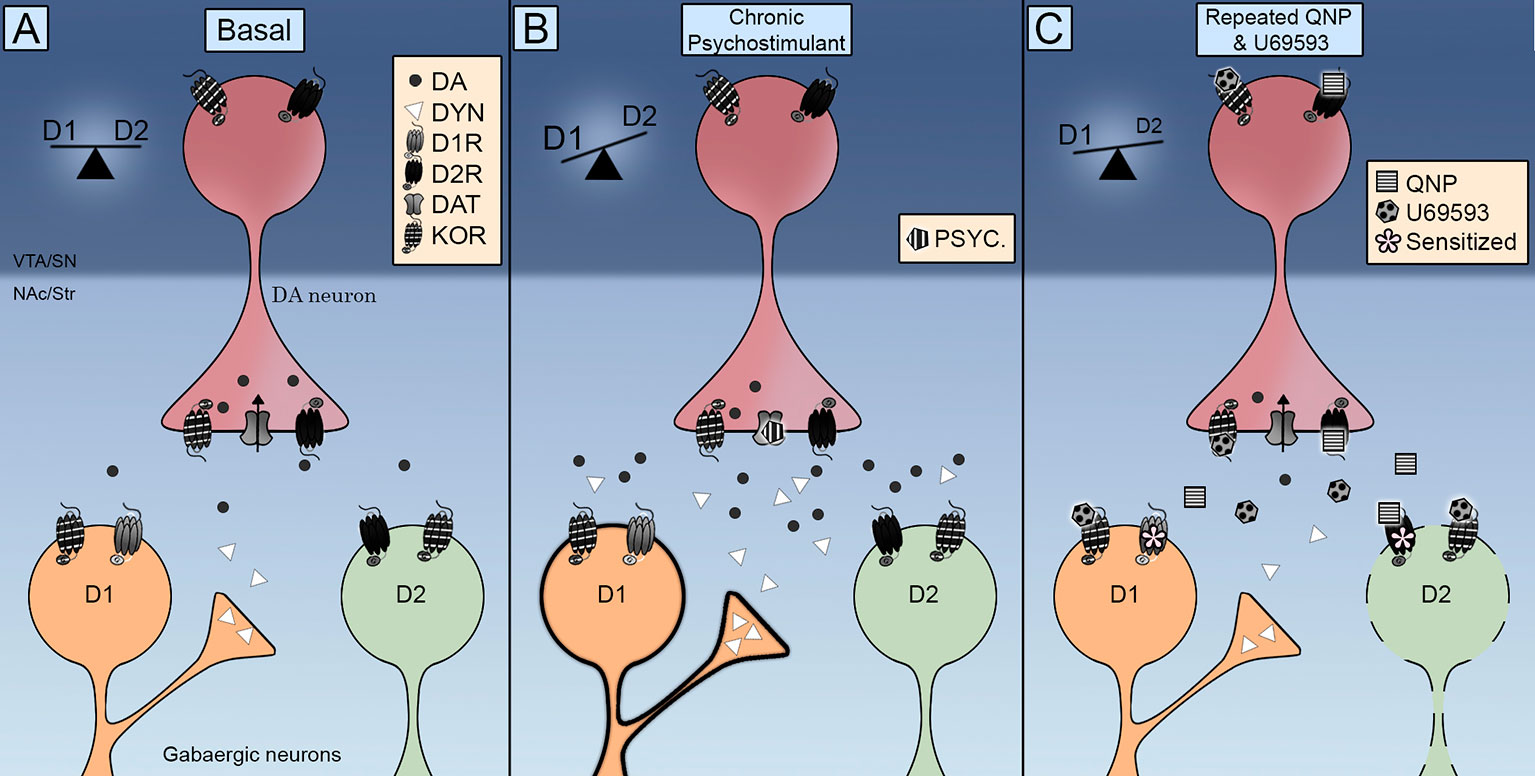 Figura 1 Esquema integrativo do controle dos receptores opioides Kappa (KOR) em vias estriadas diretas (D1R) e indiretas (D2R). (UMA) Os KOR estão localizados pré-sinapticamente nos terminais da dopamina e pós-sinapticamente nos neurônios de tamanho médio (MSNs). Sua ativação controla os níveis extracelulares de dopamina e sua localização promove a interação com o transportador de dopamina (DAT) e os receptores de dopamina D2. (B) A exposição repetida a um psicoestimulante é acompanhada por um aumento nos níveis extracelulares de dopamina e na dinorfina. A ativação dos receptores D1 e D2 muda a balança para a via direta D1R, promovendo a sensibilização locomotora. (C) A coadministração de quinpirol e U69593 é acompanhada por uma diminuição nos níveis extracelulares de dopamina. A ativação concomitante dos receptores KOR e D2 debilita a via indireta D2, induzindo o comportamento compulsivo.
Figura 1 Esquema integrativo do controle dos receptores opioides Kappa (KOR) em vias estriadas diretas (D1R) e indiretas (D2R). (UMA) Os KOR estão localizados pré-sinapticamente nos terminais da dopamina e pós-sinapticamente nos neurônios de tamanho médio (MSNs). Sua ativação controla os níveis extracelulares de dopamina e sua localização promove a interação com o transportador de dopamina (DAT) e os receptores de dopamina D2. (B) A exposição repetida a um psicoestimulante é acompanhada por um aumento nos níveis extracelulares de dopamina e na dinorfina. A ativação dos receptores D1 e D2 muda a balança para a via direta D1R, promovendo a sensibilização locomotora. (C) A coadministração de quinpirol e U69593 é acompanhada por uma diminuição nos níveis extracelulares de dopamina. A ativação concomitante dos receptores KOR e D2 debilita a via indireta D2, induzindo o comportamento compulsivo.
Diferenças sexuais de interações KOR-dopamina em comportamentos compulsivos
Estudos clínicos mostraram diferenças sexuais no comportamento compulsivo, incluindo busca compulsiva de drogas. Observa-se um início mais precoce dos sintomas do TOC nos homens em comparação com as mulheres (Mathis e outros, 2011), com mulheres mostrando maior prevalência de sintomas de contaminação e limpeza (Labad e outros, 2008) Em relação às diferenças entre os sexos na dependência de drogas, as evidências clínicas indicam que, embora o uso de drogas seja mais prevalente nos homens, as mulheres apresentam uma progressão mais rápida do que os homens na busca compulsiva por drogas (Hernández-Ávila et al., 2004; Fattore e Melis, 2016).
Ultimamente, as evidências pré-clínicas destacaram fortemente as bases neurobiológicas subjacentes às diferenças entre os sexos no abuso de drogas observadas em estudos clínicos (Becker e Chartoff, 2019) Observações iniciais usando microdiálise sem fluxo líquido mostraram que a concentração extracelular de dopamina no estriado dorsal varia durante o ciclo estral, com níveis mais elevados de próestro e estro em comparação com o diestro. Além disso, enquanto a ovariectomia diminui a concentração extracelular de dopamina estriatal em ratos fêmeas, a castração de ratos machos não modifica a concentração extracelular estriada de dopamina (Xiao e Becker, 1994), sugerindo um papel importante dos hormônios ovários na atividade da dopamina. Além disso, os hormônios femininos regulam a resposta aos psicoestimulantes. Cedo in vitro experimentos mostraram que o estradiol e a progesterona restauram a liberação de dopamina induzida por anfetamina do tecido estriatal obtido de ratas ovariectomizadas (Becker e Ramirez, 1981) Mais recentemente, estudos rápidos de voltametria cíclica de varredura mostraram que as mulheres exibem maior liberação e captação de dopamina estimulada eletricamente em comparação aos homens (Walker et al., 2000) Essas diferenças entre os sexos na neurotransmissão por dopamina podem ser responsáveis pela maior procura de cocaína e anfetamina observada nas mulheres. (Roberts e outros, 1989; Cox et al., 2013).
A regulação da KOR nos níveis extracelulares da dopamina também mostra diferenças entre os sexos (Chartoff e Mavrikaki, 2015) Usando autoestimulação intracraniana e voltametria cíclica, Conway et ai. (2019) mostraram que a menor sensibilidade ao efeito anedônico agudo de um agonista de KOR observado em ratos fêmeas em comparação com ratos machos é acompanhada por uma inibição atenuada da liberação estimulada de dopamina no NAc (Conway et al., 2019) Foi sugerido que o estradiol contribui para a inibição embotada da liberação de dopamina observada em ratos fêmeas após a ativação do KOR (Abraão et al., 2018) Embora a diafonia entre KORs e sinalização de dopamina tenha sido estudada em homens (Tejeda e Bonci, 2019), faltam pesquisas sobre essa interação e seu impacto no processo de dependência de mulheres (Chartoff e Mavrikaki, 2015) Em ratos fêmeas, a administração aguda do agonista KOR U69593 atenuou a hiperlocomoção induzida por cocaína em ratos controle e ovariectomizados. Curiosamente, a administração repetida de U69593 atenuou a hiperlocomoção induzida por cocaína de maneira dependente do estradiol (Puig-Ramos et al., 2008) Esses dados sugerem que o estradiol inicia as ações da KOR em ratos fêmeas, um efeito que pode estar relacionado às diferenças sexuais na resposta ao estresse (Puig-Ramos et al., 2008) Se em ratos fêmeas a ativação repetida de KORs facilita a liberação estriada de dopamina, como observado em machos, é uma questão sem resposta.
Embora seja observada uma facilitação na liberação de dopamina induzida por psicoestimulante em fêmeas em comparação com ratos machos, as diferenças sexuais nos mecanismos de dopamina subjacentes à sensibilização locomotora por anfetaminas não foram totalmente elucidadas (Becker, 1999) A exposição repetida à anfetamina induz uma maior atividade locomotora em ambos os adolescentes (Mathews e McCormick, 2007) e ratos adultos do sexo feminino (Milesi-Hallé et al., 2007), com ratos adolescentes do sexo feminino mostrando uma sensibilização locomotora mais robusta após exposição repetida à anfetamina. A ativação neonatal do receptor D2 potencializou a sensibilização comportamental induzida por anfetaminas apenas em ratos fêmeas (Brown et al., 2011) Como mencionado anteriormente, em ratos machos, a exposição repetida ao agonista D2 induz sensibilização locomotora e comportamento do tipo compulsivo (Dvorkin e outros, 2006) Além disso, a co-ativação do KOR potencializa a sensibilização locomotora induzida pela exposição repetida ao quinpirol, facilitando o controle inibitório dos receptores D2 na liberação de DA no NAc (Escobar e outros, 2017) Diferenças entre os sexos, como a menor sensibilidade observada ao efeito inibitório do KOR na liberação de dopamina em mulheres (Conway et al., 2019) pode ser responsável por uma contribuição diferencial da KOR na busca compulsiva de drogas.
Conclusões
Como os KORs modulam a sinalização de dopamina para elaborar comportamentos motivados e quando isso resulta em um comportamento compulsivo sensibilizado? Dados anatômicos mostram que os KORs estão primorosamente posicionados para controlar a atividade sináptica dos neurônios da dopamina no mesencéfalo. Dados funcionais indicam que os KORs controlam o funcionamento de DAT e D2R, bem como a taxa de disparo dos neurônios dopaminérgicos. A evidência inicial que mostra que a ativação aguda de KORs diminui a liberação de dopamina induzida por drogas de abuso foi complementada com dados indicando que a ativação repetida da KOR facilita a liberação de dopamina e a busca compulsiva por drogas. A sinalização da dopamina equilibra as vias de saída direta e indireta de áreas estriatais (Figura 1A) A estimulação crônica com psicoestimulantes aumenta a liberação de dopamina, ativando D1R e D2R (Figura 1B) ou quinpirol que ativam apenas D2R (Figura 1C) resulta em sensibilização locomotora e comportamentos compulsivos por uma via indireta D2R debilitada, alternando assim a balança para a via direta D1R. A transmissão de KOR é aumentada durante a ingestão crônica de psicoestimulantes pelo aumento da dinorfina nos neurônios D1 estriados (Figura 1B) Uma transmissão KOR aprimorada é emulada no modelo farmacológico do TOC através da administração de U69593. Essa ativação KOR concomitante debilita ainda mais a via indireta D2 (Figura 1C) Pesquisas futuras devem ser realizadas para elucidar completamente as consequências da ativação do KOR na atividade DAT, entender o papel do sistema KOR endógeno na compulsividade induzida pelo quinpirol e determinar a contribuição do sistema KOR para as diferenças sexuais observadas nos comportamentos compulsivos.
Contribuições do autor
AE, MA e JF contribuíram para a concepção do manuscrito. AE e JF escreveram o primeiro rascunho do manuscrito com a contribuição de MA. MA e JC contribuíram para a revisão crítica e edição do manuscrito. Todos os autores o aprovaram para publicação.
Métodos
O trabalho dos autores citados nesta revisão foi apoiado pelos números de concessão do FONDECYT: 1110352 e 1150200 à MA; 1141088 para JF; Concessão DIPOG 391340281 a JF; FONDECYT Pós-doutorado 3170497 para JC e 3190843 para AE.
Conflito de interesses
Os autores declaram que a pesquisa foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.
Atualmente, o editor de manipulação está organizando um Tópico de Pesquisa com um dos autores JF e confirma a ausência de qualquer outra colaboração.
Referências
Abraham, AD, Fontaine, HM, Song, AJ, Andrews, MM, Baird, MA, Kieffer, BL, et al. (2017). A ativação do receptor opióide Kappa nos neurônios da dopamina interrompe a inibição comportamental. Neuropsychopharmacology 43 (2), 362-372. doi: 10.1038 / npp.2017.133
Abraham, AD, Schattauer, SS, Reichard, KL, Cohen, JH, Fontaine, HM, Song, AJ, et al. (2018). A regulação estrogênica do GRK2 inativa a analgesia mediadora da sinalização do receptor kappa opióide, mas não a aversão. J. Neurosci. 38 (37), 8031-8043. doi: 10.1523 / JNEUROSCI.0653-18.2018
Asaoka, N., Nishitani, N., Kinoshita, H., Nagai, Y., Hatakama, H., Nagayasu, K., et al. (2019). Um antagonista do receptor A2A da adenosina melhora múltiplos sintomas de psicose induzida por quinpirol repetida. eNeuro 6 (1), 1-16. ENEURO.0366-18.2019. doi: 10.1523 / ENEURO.0366-18.2019
Atigari, DV, Uprety, R., Pasternak, GW, Majumdar, S., Kivell, BM (2019). MP1104, um agonista misto do receptor opióide kappa-delta tem propriedades anti-cocaína com efeitos colaterais reduzidos em ratos. Neurofarmacologia 150, 217 – 228. doi: 10.1016 / j.neuropharm.2019.02.010
Azocar, VH, Sepúlveda, G., Ruiz, C., Aguilera, C., Andrés, ME, Fuentealba, JA (2019). O bloqueio do receptor kappa-opióide reverte as alterações na dinâmica dopaminérgica dorsolateral do estriado durante a sensibilização à anfetamina. J. Neurochem. 148, 348-358. doi: 10.1111 / jnc.14612
Béguin, C., Potuzak, J., Xu, W., Liu-Chen, LY, Streicher, JM, Groer, CE, et al. (2012). Propriedades diferenciais de sinalização no receptor opióide kappa da 12-epi-salvinorina A e seus análogos. Bioorg. Med. Química Lett. 15; 22 (2), 1023-1026. doi: 10.1016 / j.bmcl.2011.11.128
Beardsley, PM, Howard, JL, Shelton, KL, Carroll, FI (2005). Efeitos diferenciais do novo antagonista dos receptores opióides kappa, JDTic, na reintegração da procura de cocaína induzida por estressores de pedal versus primos de cocaína e seus efeitos antidepressivos em ratos. Psicofármaco. (Berl) 183, 118–126. doi: 10.1007/s00213-005-0167-4
Becker, JB, Chartoff, E. (2019). Diferenças de sexo nos mecanismos neurais mediando recompensa e dependência. Neuropsychopharmacology 44, 166–183. doi: 10.1038/s41386-018-0125-6
Becker, JB, Ramirez, VD (1981). As diferenças entre os sexos na liberação de anfetaminas estimularam as catecolaminas do tecido estriado de ratos in vitro. Cérebro Res. 204, 361–372. doi: 10.1016/0006-8993(81)90595-3
Becker, JB (1999). Diferenças de gênero na função dopaminérgica no estriado e núcleo accumbens. Pharmacol. Biochem. Behav. 64, 803–812. doi: 10.1016/S0091-3057(99)00168-9
Berridge, KC, Robinson, TE (2016). Gostar, querer e a teoria de incentivo-sensibilização do vício. Sou. Psychol. 71, 670-679. doi: 10.1037 / amp0000059
Berridge, KC, Venier, IL, Robinson, TE (1989). Análise da reatividade gustativa da afagia induzida por 6-hidroxidopamina: implicações para as hipóteses de excitação e anedonia da função da dopamina. Behav. Neurosci. 103, 36 – 45. doi: 10.1037 / 0735-7044.103.1.36
Broadbear, JH, Negus, SS, Butelman, ER, Costa, BR, Woods, JH (1994). Efeitos diferenciais da norbinaltorfimina administrada sistemicamente (nor-BNI) nos agonistas kappa-opióides no ensaio de contorção em camundongos. Psychopharmacology 115, 311 – 319. doi: 10.1007 / BF02245071
Brown, RW, Perna, MK, Noel, DM, Whittemore, JD, Lehmann, J., Smith, ML (2011). Sensibilização locomotora de anfetamina e preferência de lugar condicionada em ratos machos e fêmeas adolescentes tratados neonatalmente com quinpirol. Behav. Pharmacol. 22, 374–378. doi: 10.1097/FBP.0b013e328348737b
Bruchas, MR. Chavkin, C. (2010). Cascatas de quinase e sinalização dirigida a ligantes no receptor kappa opióide. Psicofármaco. (Berl) 210, 137–147. doi: 10.1007/s00213-010-1806-y
Callaghan, CK, Rouine, J., O'Mara, SM (2018). Potenciais papéis dos receptores opióides na motivação e no transtorno depressivo maior. Prog. Cérebro Res. 239, 89-119. doi: 10.1016 / bs.pbr.2018.07.009
Carr, DB, Sesack, SR (2000). Os neurônios contendo GABA na área tegmentar ventral do rato se projetam para o córtex pré-frontal. Sinapse 38 (2), 114–123. doi: 10.1002/1098-2396(200011)38:2<114::AID-SYN2>3.0.CO;2-R
Casanova, JP, Velis, GP, Fuentealba, JA (2013). A sensibilização locomotora de anfetamina é acompanhada de uma liberação aumentada de dopamina estimulada por K + no córtex pré-frontal medial do rato. Behav. Cérebro Res. 237, 313 – 317. doi: 10.1016 / j.bbr.2012.09.052
Chartoff, EH, Mavrikaki, M. (2015). Diferenças de sexo na função dos receptores kappa opioides e seu potencial impacto no vício. Frente. Neurosci. 9, 466. doi: 10.3389 / fnins.2015.00466
Chartoff, EH, Ebner, SR, Sparrow, A., Potter, D., Baker, PM, Ragozzino, ME, et al. (2016). O tempo relativo entre a ativação do receptor kappa opioide e a cocaína determina o impacto na recompensa e na liberação de dopamina. Neuropsychopharmacology 41, 989 – 1002. doi: 10.1038 / npp.2015.226
Chavkin, C., Koob, GF (2016). Dinorfina, disforia e dependência: o estresse do vício. Neuropsychopharmacology 41, 373 – 374. doi: 10.1038 / npp.2015.258
Chefer, VI, Czyzyk, T., Bolan, EA, Moron, J., Pintar, JE, Shippenberg, T. S. (2005). Os sistemas de receptores kapa-opióides endógenos regulam a dinâmica da dopamina mesoacumbal e a vulnerabilidade à cocaína. J. Neurosci. 25, 5029 – 5037. doi: 10.1523 / JNEUROSCI.0854-05.2005
Chefer, VI, Zapata, A., Shippenberg, TS, Bungay, PM (2006). A microdiálise quantitativa sem fluxo líquido permite a detecção de aumentos e diminuições na captação de dopamina no núcleo accumbens de camundongo. J. Neurosci. Métodos 155, 187 – 193. doi: 10.1016 / j.jneumeth.2005.12.018
Cole, RL, Konradi, C., Douglass, J., Hyman, SE (1995). Adaptação neuronal à anfetamina e dopamina: mecanismos moleculares da regulação do gene da prodinorfina no estriado de ratos. Neurônio 14, 813–823. doi: 10.1016/0896-6273(95)90225-2
Conway, SM, Puttick, D., Russell, S., Potter, D., Roitman, MF, Chartoff, E. H. (2019). As mulheres são menos sensíveis do que os homens aos efeitos motivacionais e supressores de dopamina da ativação do receptor opioide kappa. Neurofarmacologia 146, 231 – 241. doi: 10.1016 / j.neuropharm.2018.12.002
Os dados foram analisados por meio de questionários, entrevistas e entrevistas. O tratamento com quinpirol neonatal melhora a ativação locomotora e a liberação de dopamina no núcleo accumbens em resposta ao tratamento com anfetaminas na idade adulta. Sinapse 64, 289 – 300. doi: 10.1002 / syn.20729
Cox, BM, Young, AB, Ver, RE, Reichel, CM (2013). Diferenças de sexo na busca de metanfetamina em ratos: impacto da ocitocina. Psiconeuroendocrinologia 38, 2343 – 2353. doi: 10.1016 / j.psyneuen.2013.05.005
Culver, KE, Szechtman, H., Levant, B. (2008). Ligação alterada do receptor do tipo dopamina D2 em ratos com sensibilização comportamental ao quinpirol: efeitos do pré-tratamento com Ro 41-1049. EUR. J. Pharmacol. 592, 67 – 72. doi: 10.1016 / j.ejphar.2008.06.101
Dalman, FC, O'Malley, KL (1999). Tolerância e dependência kappa-opioide em culturas de neurônios dopaminérgicos do mesencéfalo. J. Neurosci. 19, 5750–5757. doi: 10.1523/JNEUROSCI.19-14-05750.1999
De Vries, TJ, Schoffelmeer, AN, Binnekade, R., Raasø, H., Vanderschuren, LJ (2002). A recaída ao comportamento de busca de cocaína e heroína mediada pelos receptores da dopamina D2 depende do tempo e está associada à sensibilização comportamental. Neuropsychopharmacology 26, 18–26. doi: 10.1016/S0893-133X(01)00293-7
Devine, DP, Leone, P., Pocock, D., Wise, RA (1993). Envolvimento diferencial dos receptores tegmentares ventrais mu, delta e kappa opioide na modulação da liberação basal de dopamina mesolímbica: estudos de microdiálise in vivo. J. Pharmacol. Exp. Ther. 266, 1236-1246.
Di Chiara, G., Imperato, A. (1988). Efeitos opostos dos agonistas opióides mu e kappa na liberação de dopamina no núcleo accumbens e no caudado dorsal de ratos em movimento livre. J. Pharmacol. Exp. Ther. 244, 1067 – 1080. doi: 10.1073 / pnas.85.14.5274
Dvorkin, A., Perreault, ML, Szechtman, H. (2006). Desenvolvimento e organização temporal da verificação compulsiva induzida por injeções repetidas do agonista da dopamina quinpirol em um modelo animal de transtorno obsessivo-compulsivo. Behav. Cérebro Res. 169, 303 – 311. doi: 10.1016 / j.bbr.2006.01.024
Ebner, SR, Roitman, MF, Potter, DN, Rachlin, AB, Chartoff, EH (2010). Os efeitos depressivos do agonista do receptor kappa opioide salvinorina A estão associados à diminuição da liberação de dopamina fásica no núcleo accumbens. Psicofármaco. (Berl) 210, 241–252. doi: 10.1007/s00213-010-1836-5
Edwards, NJ, Tejeda, HA, Pignatelli, M., Zhang, S., McDevitt, RA, Wu, J., et al. (2017). A especificidade do circuito na arquitetura inibitória do ATV regula o comportamento induzido pela cocaína. Nat. Neurosci. 20 (3), 438-448. doi: 10.1038 / nn.4482
Ehrich, JM, Phillips, PEM, Chavkin, C. (2014). A ativação do receptor opióide Kappa potencializa o aumento induzido pela cocaína na liberação evocada de dopamina registrada in vivo no núcleo accumbens do camundongo. Neuropsychopharmacology 39, 3036 – 3048. doi: 10.1038 / npp.2014.157
Ehrich, JM, Messinger, DI, Knakal, CR, Kuhar, JR, Schattauer, SS, Bruchas, MR, et al. (2015). Aversão induzida por receptor opióide Kappa requer ativação de MAPK p38 em neurônios da dopamina VTA. J. Neurosci. 35, 12917 – 12931. doi: 10.1523 / JNEUROSCI.2444-15.2015
Eilam, D., Szechtman, H. (1989). Efeito bifásico do agonista quinpirol D-2 na locomoção e movimentos. EUR. J. Pharmacol. 161, 151–157. doi: 10.1016/0014-2999(89)90837-6
Eilam, D., Szechtman, H. (2005). Comportamento induzido por psicoestimulante como modelo animal de transtorno obsessivo-compulsivo: uma abordagem etológica da forma de rituais compulsivos. Espectro do CNS. 10, 191-202. doi: 10.1017 / S109285290001004X
Escobar, AP, Cornejo, FA, Andrés, ME, Fuentealba, JA (2012). O tratamento repetido com o agonista do receptor opioide kappa U69593 reverte a liberação aumentada de dopamina induzida por K + no núcleo accumbens, mas não a expressão da sensibilização locomotora em ratos sensibilizados com anfetamina. Neurochem. Int. 60 (4), 344-349. doi: 10.1016 / j.neuint.2012.01.014
Escobar, AP, Cornejo, FA, Olivares-Costa, M., González, M., Fuentealba, JA, Gysling, K., et al. (2015). A neurotransmissão reduzida de dopamina e glutamato no núcleo accumbens de ratos sensibilizados com quinpirol sugere a função inibidora do autoreceptor D2. J. Neurochem. 134, 1081-1090. doi: 10.1111 / jnc.13209
Escobar, AP, González, MP, Meza, RC, Noches, V., Henny, P., Gysling, K., et al. (2017). Mecanismos de potenciação do receptor kappa opióide da função do receptor D2 da dopamina na sensibilização locomotora induzida por quinpirol em ratos. Int. J. Neuropsychopharmacol. 20, 660-669. doi: 10.1093 / ijnp / pyx042
Everitt, BJ, Robbins, TW (2005). Sistemas neurais de reforço para dependência de drogas: de ações a hábitos e compulsão. Nat. Neurosci. 8, 1481 – 1489. doi: 10.1038 / nn1579
Fattore, L., Melis, M. (2016). Diferenças de sexo em comportamentos impulsivos e compulsivos: um foco na dependência de drogas. Viciado. Biol. 21 (5), 1043-1051. doi: 10.1111 / adb.12381
Ferrario, CR, Gorny, G., Crombag, HS, Li, Y., Kolb, B., Robinson, TE (2005). Plasticidade neural e comportamental associada à transição do uso controlado para escalado de cocaína. Biol. Psi. 58 (9), 751-9.
Figee, M., Pattij, T., Willuhn, I., Luigjes, J., van den Brink, W., Goudriaan, A., et al. (2016). Compulsividade em transtorno obsessivo-compulsivo e vícios. EUR. Neuropsychopharmacol. 26, 856 – 868. doi: 10.1016 / j.euroneuro.2015.12.003
Ford, CP, Mark, GP, Williams, JT (2006). As propriedades e a inibição opióide dos neurônios da dopamina mesolímbica variam de acordo com a localização do alvo. J. Neurosci. 26, 2788 – 2797. doi: 10.1523 / JNEUROSCI.4331-05.2006
Ford, CP, Beckstead, MJ, Williams, JT (2007). Inibição opióide Kappa das correntes pós-sinápticas inibidoras da dopamina somatodendrítica. J. Neurophysiol. 97, 883 – 891. doi: 10.1152 / jn.00963.2006
Fuentealba, JA, Gysling, K., Magendzo, K., Andrés, ME (2006). A administração repetida do agonista seletivo do receptor kappa-opióide U-69593 aumenta os níveis extracelulares estimulados de dopamina no núcleo accumbens do rato. J. Neurosci. Res. 84, 450 – 459. doi: 10.1002 / jnr.20890
Fuentealba, JA, Gysling, K., Andrés, ME (2007). Aumento da resposta locomotora à anfetamina induzida pela administração repetida do agonista seletivo do receptor kappa-opióide U-69593. Sinapse 61, 771 – 777. doi: 10.1002 / syn.20424
Gehrke, BJ, Chefer, VI, Shippenberg, TS (2008). Efeitos da administração aguda e repetida de salvinorina A na função da dopamina no estriado dorsal de ratos. Psicofármaco. (Berl) 197, 509–517. doi: 10.1007/s00213-007-1067-6
Giuliano, C., Belin, D., Everitt, BJ (2019). O consumo compulsivo de álcool resulta de uma falha em desativar o controle estriado dorsolateral sobre o comportamento. J. Neurosci. 39 (9), 1744-1754. doi: 10.1523 / JNEUROSCI.2615-18.2018
Goto, Y., Grace, AA (2008). Processamento de informações límbicas e corticais no núcleo accumbens. Tendências Neurosci. 31, 552 – 558. doi: 10.1016 / j.tins.2008.08.002
Grace, AA, Bunney, BS (1980). Neurônios da dopamina nigral: registro e identificação intracelular com injeção de L-dopa e histofluorescência. Ciência 210, 654 – 656. doi: 10.1126 / science.7433992
Gray, AM, Rawls, SM, Shippenberg, TS, McGinty, JF (1999). O agonista κ-opioide, U-69593, diminui os comportamentos agudos evocados por anfetaminas e os níveis de dopamina e glutamato dependentes de cálcio no estriado ventral. J. Neurochem. 73, 1066 – 1074. doi: 10.1046 / j.1471-4159.1999.0731066.x
Graziane, NM, Polter, AM, Briand, LA, Pierce, RC, Kauer, JA (2013). Os receptores opioides Kappa regulam a busca pela cocaína induzida pelo estresse e a plasticidade sináptica. Neurônio 77, 942 – 954. doi: 10.1016 / j.neuron.2012.12.034
Heidbreder, CA, Shippenberg, TS (1994). O U-69593 evita a sensibilização à cocaína ao normalizar a dopamina Accumbens basal. Neuroreport 5, 1797–1800. doi: 10.1097/00001756-199409080-00028
Hernandez-Avila, CA 1., Rounsaville, BJ, Kranzler, HR (2004). Mulheres dependentes de opióides, maconha e álcool mostram progressão mais rápida para o tratamento de abuso de substâncias. Álcool de Drogas Depende. 74 (3), 265–272. doi: 10.1016 / j.drugalcdep.2004.02.001
Hoffman, AF, Spivak, CE, Lupica, CR (2016). Liberação aprimorada de dopamina por inibidores de transporte de dopamina, descrita por um modelo de difusão restrito e voltametria cíclica de varredura rápida. ACS Chem. Neurosci. 7, 700-709. doi: 10.1021 / acschemneuro.5b00277
Holden, C. (2001). Vícios "comportamentais": eles existem? Ciência 294, 980 – 982. doi: 10.1126 / science.294.5544.980
Imperato, A., Di Chiara, G. (1988). Efeitos de agonistas e antagonistas dos receptores D-1 e D-2 aplicados localmente estudados com diálise cerebral. EUR. J. Pharmacol. 156, 385–393. doi: 10.1016/0014-2999(88)90284-1
Jackson, KJ, McLaughlin, JP, Carroll, FI, Damaj, MI (2013). Efeitos do antagonista dos receptores opióides kappa, norbinaltorfimina, no estresse e na reposição induzida por drogas da preferência de locais condicionados à nicotina em camundongos. Psicofármaco. (Berl) 226, 763–768. doi: 10.1007/s00213-012-2716-y
Kivell, B., Uzelac, Z., Sundaramurthy, S., Rajamanickam, J., Ewald, A., Chefer, V., et al. (2014). A salvinorina A regula a função do transportador de dopamina através de um receptor opioide kappa e de um mecanismo dependente de ERK1 / 2. Neurofarmacologia 86, 228 – 240. doi: 10.1016 / j.neuropharm.2014.07.016
Koeltzow, TE, Austin, JD, Vezina, P. (2003). A sensibilização comportamental ao quinpirol não está associada ao aumento do excesso de dopamina no núcleo accumbens. Neurofarmacologia 44, 102–110. doi: 10.1016/S0028-3908(02)00328-3
Koob, GF, Volkow, ND (2016). Neurobiologia do vício: uma análise neurocircuito. Lancet Psychiatry 3, 760–773. doi: 10.1016/S2215-0366(16)00104-8
Koob, GF (2013). O vício é um déficit de recompensa e transtorno de excesso de estresse. Frente. Psiquiatria 4, 72. doi: 10.3389 / fpsyt.2013.00072
Labad, J. 1., Menchon, JM, Alonso, P., Segalas, C., Jimenez, S., Jaurrieta, N., et al. (2008). Diferenças de gênero nas dimensões dos sintomas obsessivo-compulsivos. Deprime a ansiedade. 25 (10), 832-838. doi: 10.1002 / da.20332
Maisonneuve, IM, Archer, S., Glick, SD (1994). O U50,488, um agonista do receptor opioide kappa, atenua os aumentos induzidos pela cocaína na dopamina extracelular no núcleo accumbens de ratos. Neurosci. Lett. 181, 57–60. doi: 10.1016/0304-3940(94)90559-2
Mansour, A., Fox, CA, Akil, H., Watson, SJ (1995). Expressão de mRNA de receptores opióides no SNC de ratos: implicações anatômicas e funcionais. Tendências Neurosci. 18 (1), 22–29. doi: 10.1016/0166-2236(95)93946-U
Mansour, A., Burke, S., Pavlic, RJ, Akil, H., Watson, SJ (1996). Localização imuno-histoquímica do receptor kappa 1 clonado no SNC do rato e na hipófise. Neuroscience 71, 671–690. doi: 10.1016/0306-4522(95)00464-5
Margolis, EB, Karkhanis, AN (2019). Contribuições dopaminérgicas celulares e de circuito à aversão mediada pelo receptor kappa opióide. Neurochem. Int. 129, 104504. doi: 10.1016 / j.neuint.2019.104504
Margolis, EB, Hjelmstad, GO, Bonci, A., Fields, HL (2003). Os agonistas Kappa-opióides inibem diretamente os neurônios dopaminérgicos do mesencéfalo. J. Neurosci. 23, 9981–9986. doi: 10.1523/JNEUROSCI.23-31-09981.2003
Margolis, EB, Hjelmstad, GO, Bonci, A., Fields, HL (2005). Os agonistas Kappa e os opióides Mu inibem a entrada glutamatérgica nos neurônios da área tegmentar ventral. J. Neurophysiol. 93, 3086 – 3093. doi: 10.1152 / jn.00855.2004
Margolis, EB, Lock, H., Chefer, VI, Shippenberg, TS, Hjelmstad, GO, Fields, HL (2006). Os opióides Kappa controlam seletivamente os neurônios dopaminérgicos que se projetam no córtex pré-frontal. Proc. Natl. Acad. Sci. EUA 103, 2938 – 2942. doi: 10.1073 / pnas.0511159103
Margolis, EB, Mitchell, JM, Ishikawa, J., Hjelmstad, GO, Fields, HL (2008). Neurônios da dopamina no mesencéfalo: o alvo da projeção determina a duração do potencial de ação e a inibição do receptor da dopamina D (2). J. Neurosci. 28, 8908 – 8913. doi: 10.1523 / JNEUROSCI.1526-08.2008
Mathews, IZ, McCormick, CM (2007). Ratos fêmeas e machos no final da adolescência diferem dos adultos na atividade locomotora induzida por anfetaminas, mas não na preferência condicionada por anfetaminas. Behav. Pharmacol. 18, 641–650. doi: 10.1097/FBP.0b013e3282effbf5
Mathis, MA 1., Pd, A., Funaro, G., RC, T., Moraes, I., AR, T., et al. (2011). Diferenças de gênero no transtorno obsessivo-compulsivo: uma revisão de literatura. Braz. J. Psiquiatria 33 (4), 390-399. doi: 10.1590 / S1516-44462011000400014
McLaughlin, JP, Marton-Popovici, M., Chavkin, C. (2003). O antagonismo do receptor opioide Kappa e a interrupção do gene da prodinorfina bloqueiam as respostas comportamentais induzidas pelo estresse. J. Neurosci. 23 (13), 5674-5683. doi: 10.1523 / JNEUROSCI.23-13-05674.2003
McLaughlin, JP, Land, BB, Li, S., Pintar, JE, Chavkin, C. (2006a). A ativação prévia dos receptores kappa opióides por U50,488 imita o estresse repetido na natação forçada para potencializar o condicionamento da preferência no local da cocaína. Neuropsychopharmacology 31, 787 – 794. doi: 10.1038 / sj.npp.1300860
McLaughlin, JP, Li, S., Valdez, J., Chavkin, TA, Chavkin, C. (2006b). A derrota social respostas comportamentais induzidas pelo estresse são mediadas pelo sistema opióide kappa endógeno. Neuropsychopharmacology 31 (6), 1241–1248. doi: 10.1038 / sj.npp.1300872
Milesi-Hallé, A., McMillan, DE, Laurenzana, EM, Byrnes-Blake, KA, Owens, SM (2007). Diferenças entre os sexos na resposta comportamental induzida por (+) - anfetamina e (+) - metanfetamina em ratos machos e fêmeas Sprague-Dawley. Pharmacol. Biochem. Behav. 86, 140 – 149. doi: 10.1016 / j.pbb.2006.12.018
Negus, SS, Mello, NK, Portoghese, PS, Lin, CE (1997). Efeitos de kappa opioides na auto-administração de cocaína por macacos rhesus. J. Pharmacol. Exp. Ther. 282, 44-55.
Nestler, EJ (2001). Bases moleculares da plasticidade a longo prazo subjacente à dependência. Nat. Rev. Neurosci. 2, 119 – 128. doi: 10.1038 / 35053570
Perreault, ML, Graham, D., Bisnaire, L., Simms, J., Hayton, S., Szechtman, H. (2006). O agonista Kappa-opioide U69593 potencializa a sensibilização locomotora ao quinpirol agonista D2 / D3: mecanismos pré e pós-sinápticos. Neuropsychopharmacology 31, 1967 – 1981. doi: 10.1038 / sj.npp.1300938
Perreault, ML, Seeman, P., Szechtman, H. (2007). A estimulação do receptor Kappa-opióide acelera a patogênese da verificação compulsiva no modelo de sensibilização ao quinpirol do transtorno obsessivo-compulsivo (TOC). Behav. Neurosci. 121, 976 – 991. doi: 10.1037 / 0735-7044.121.5.976
Pierce, RC, Kalivas, PW (1997). Um modelo de circuitos da expressão de sensibilização comportamental a psicoestimulantes do tipo anfetamina. Cérebro Res. Cérebro Res. Rev. 25, 192–216. doi: 10.1016/S0165-0173(97)00021-0
Polter, AM, Bispo, RA, Briand, LA, Graziane, NM, Pierce, RC, Kauer, JA (2014). O bloco pós-esforço de receptores opioides Kappa resgata a potencialização a longo prazo de sinapses inibitórias e evita o restabelecimento da procura de cocaína. Biol. Psiquiatria 76, 785 – 793. doi: 10.1016 / j.biopsych.2014.04.019
Puig-Ramos, A., Santiago, GS, Segarra, AC (2008). O U-69593, um agonista do receptor opióide kappa, diminui a sensibilização comportamental induzida por cocaína em ratos fêmeas. Behav. Neurosci. 122, 151 – 160. doi: 10.1037 / 0735-7044.122.1.151
Robbins, TW, Gillan, CM, Smith, DG, de Wit, S., Ersche, KD (2012). Endofenótipos neurocognitivos de impulsividade e compulsividade: em direção à psiquiatria dimensional. Tendências Cognit. Sci. 16, 81 – 91. doi: 10.1016 / j.tics.2011.11.009
Roberts, DCS, Bennett, SAL, Vickers, GJ (1989). O ciclo estral afeta a auto-administração de cocaína em um esquema de proporção progressiva em ratos. Psicofármaco. (Berl) 98, 408 – 411. doi: 10.1007 / BF00451696
Robinson, TE, Berridge, KC (1993). A base neural do desejo por drogas: uma teoria de estímulo-sensibilização do vício. Cérebro Res. Cérebro Res. Rev. 18, 247–291. doi: 10.1016/0165-0173(93)90013-P
Robinson, TE, Berridge, KC (2001). Sensibilização por incentivos e dependência. Vício 96, 103 – 114. doi: 10.1046 / j.1360-0443.2001.9611038.x
Rose, JH, Karkhanis, AN, Steiniger-Brach, B., Jones, SR (2016). Efeitos distintos do nalmefeno nas taxas de captação de dopamina e na atividade dos receptores kappa opioides no núcleo accumbens após a exposição crônica intermitente ao etanol. Int. J. Mol. Sci. 17, 1216. doi: 10.3390 / ijms17081216
Schlosburg, JE, Whitfield, TW, Jr., Park, PE, Crawford, EF, George, O., Vendruscolo, LF, et al. (2013). O antagonismo a longo prazo dos receptores opióides κ impede a escalada e o aumento da motivação para a ingestão de heroína. J. Neurosci. 33 (49), 19384-19392. doi: 10.1523 / JNEUROSCI.1979-13.2013
Sesack, SR, Pickel, VM (1992). Localização ultraestrutural dupla da imunorreatividade da encefalina e tirosina hidroxilase na área tegmentar ventral do rato: múltiplos substratos para interações ópio-dopamina. J. Neurosci. 12, 1335–a1350. doi: 10.1523/JNEUROSCI.12-04-01335.1992
Sesack, SR, Aoki, C., Pickel, VM (1994). Localização ultraestrutural da imunorreatividade do receptor D2 em neurônios da dopamina no cérebro e seus alvos estriatais. J. Neurosci. 14, 88–106. doi: 10.1523/JNEUROSCI.14-01-00088.1994
Sesia, T., Bizup, B., Grace, AA (2013). Avaliação de modelos animais de transtorno obsessivo-compulsivo: correlação com a atividade dos neurônios dopaminérgicos fásicos. Int. J. Neuropsychopharmacol. 16, 1295-1307. doi: 10.1017 / S146114571200154X
Shippenberg, TS, Zapata, A., Chefer, VI (2007). Dinorfina e a fisiopatologia da toxicodependência. Pharmacol. Ter. 116, 306-321. doi: 10.1016 / j.pharmthera.2007.06.011
Smith, JA, Loughlin, SE, Leslie, FM (1992). inibição kappa-opióide da liberação de [3H] dopamina a partir de culturas de células dissociadas mesencefálicas ventrais de rato. Mol. Pharmacol. 42.
Smith, JW, Fetsko, LA, Xu, R., Wang, Y. (2002). Os camundongos knockout para o receptor Dopamine D2L exibem déficits nas propriedades de reforço positivas e negativas da morfina e no aprendizado de evitação. Neuroscience 113 (4), 755–765. doi: 10.1016/S0306-4522(02)00257-9
Smith, JS, Schindler, AG, Martinelli, E., Gustin, RM, Bruchas, MR, Chavkin, C. (2012). A ativação induzida por estresse do sistema receptor de dinorfina / κ-opioide na amígdala potencializa a preferência por locais condicionados à nicotina. J. Neurosci. 32, 1488 – 1495. doi: 10.1523 / JNEUROSCI.2980-11.2012
Spanagel, R., Herz, A., Shippenberg, TS (1992). Os sistemas opióides endógenos tonicamente ativos opostos modulam a via dopaminérgica mesolímbica. Proc. Natl. Acad. Sci. 89, 2046 – 2050. doi: 10.1073 / pnas.89.6.2046
Speciale, SG, Manaye, KF, Sadeq, M., Alemão, DC (1993). Receptores opióides nas regiões dopaminérgicas do mesencéfalo do rato. II Autoradiografia Kappa e receptor delta. J. Transm. Neural. Seção Gen. 91, 53 – 66. doi: 10.1007 / BF01244918
Sperling, RE, Gomes, SM, Sypek, EI, Carey, AN, McLaughlin, JP (2010). Mediação kappa-opióide endógena da potenciação induzida por estresse da preferência e autoadministração condicionadas ao etanol. Psicofármaco. (Berl) 210 (2), 199–209. doi: 10.1007/s00213-010-1844-5
Steketee, JD, Kalivas, PW (2011). Procura de drogas: sensibilização comportamental e recaída no comportamento de busca de drogas sibley DR, ed. Pharmacol. Rev. 63, 348-365. doi: 10.1124 / pr.109.001933
Stuchlik, A., Radostová, D., Hatalova, H., Vales, K., Nekovarova, T., Koprivova, J., et al. (2016). Validade do modelo de rato com sensibilização por quinpirol do TOC: ligando evidências de estudos animais e clínicos. Frente. Behav. Neurosci. 10, 209. doi: 10.3389 / fnbeh.2016.00209
Sun, T., Song, Z., Tian, Y., Tian, W., Zhu, C., Ji, G., et al. (2019). A entrada da amígdala basolateral no córtex pré-frontal medial controla o comportamento de verificação do tipo transtorno obsessivo-compulsivo. Proc. Natl. Acad. Sci. EUA 116, 3799 – 3804. doi: 10.1073 / pnas.1814292116
Svingos, AL, Chavkin, C., Colago, EEO, Pickel, VM (2001). Coexpressão maior de receptores opióides - e do transportador de dopamina nos perfis axonais do núcleo accumbens. Sinapse 42, 185 – 192. doi: 10.1002 / syn.10005
Szechtman, H., Talangbayan, H., Eilam, D. (1993). Componentes ambientais e comportamentais da sensibilização induzida pelo agonista da dopamina quinpirol. Behav. Pharmacol. 4, 405–410. doi: 10.1097/00008877-199308000-00014
Szechtman, H., Talangbayan, H., Canaran, G., Dai, H., Eilam, D. (1994). Dinâmica da sensibilização comportamental induzida pelo agonista da dopamina quinpirol e um mecanismo central de controle de energia proposto. Psicofármaco. (Berl) 115, 95 – 104. doi: 10.1007 / BF02244757
Szechtman, H., Sulis, W., Eilam, D. (1998). Quinpirol induz o comportamento de verificação compulsiva em ratos: um modelo animal potencial de transtorno obsessivo-compulsivo (TOC). Behav. Neurosci. 112, 1475 – 1485. doi: 10.1037 / 0735-7044.112.6.1475
Szechtman, H., Culver, K., Eilam, D. (1999). Papel dos sistemas de dopamina no transtorno obsessivo-compulsivo (TOC): implicações de um novo modelo animal induzido por psicoestimulante. pol. J. Pharmacol. 51, 55-61.
Szechtman, H., Eckert, MJ, Tse, WS, Boersma, JT, Bonura, CA, JZ, M., et al. (2001) Comportamento de verificação compulsiva de ratos sensibilizados a quinpirol como modelo animal de Transtorno Obsessivo-Compulsivo (TOC): forma e controle. BMC Neurosci. 2, 4. doi: 10.1186/1471-2202-2-4
Szechtman, H., Ahmari, SE, Beninger, RJ, Eilam, D. Harvey, BH, Edemann-Callesen, H., et al. (2017). Transtorno obsessivo-compulsivo: percepções de modelos animais. Neurosci. Biobehav. Rev. 76, 254 – 279. doi: 10.1016 / j.neubiorev.2016.04.019
Tejeda, HA, Bonci, A. (2019). Controle da dinâmica da dopamina por dinorfina / receptor opióide kappa: implicações para estados afetivos negativos e transtornos psiquiátricos. Cérebro Res. 1713, 91 – 101. doi: 10.1016 / j.brainres.2018.09.023
Tejeda, HA, Wu, J., Kornspun, AR, Pignatelli, M., Kashtelyan, V., Krashes, M. J., et al. (2017). A modulação do receptor kappa-opioide específico da via e da célula do equilíbrio de excitação-inibição bloqueia diferencialmente a atividade dos neurônios acumbens d1 e d2. Neurônio 93 (1), 147-163. doi: 10.1016 / j.neuron.2016.12.005
Thompson, AC, Zapata, A., Justice, JB, Vaughan, RA, Sharpe, LG, Shippenberg, TS (2000). A ativação do receptor Kappa-opióide modifica a captação de dopamina no núcleo accumbens e se opõe aos efeitos da cocaína. J. Neurosci. 20, 9333–9340. doi: 10.1523/JNEUROSCI.20-24-09333.2000
Thompson, D., Martini, L., Whistler, JL (2010). A proporção alterada de receptores de dopamina D1 e D2 no estriado de camundongos está associada à sensibilização comportamental à cocaína. PloS One 5, e11038. doi: 10.1371 / journal.pone.0011038
Usiello, A., Baik, J.-H., Rougé-Pont, F., Picetti, R., Dierich, A., LeMeur, M., et al. (2000) Funções distintas das duas isoformas dos receptores da dopamina D2. Natureza 408, 199 – 203. doi: 10.1038 / 35041572
Van Bockstaele, EJ, Pickel, VM (1995). Os neurônios que contêm GABA na área tegmentar ventral projetam-se para o núcleo accumbens no cérebro de ratos. Cérebro Res. 682 (1-2), 215–221. doi: 10.1016/0006-8993(95)00334-M
Van't Veer, A., Bechtholt, AJ, Onvani, S., Potter, D., Wang, Y., Liu-Chen, LY, et al. (2013). A ablação dos receptores kappa-opióides dos neurônios da dopamina do cérebro tem efeitos semelhantes aos ansiolíticos e aumenta a plasticidade induzida pela cocaína. Neuropsychopharmacology 38 (8), 1585-1597. doi: 10.1038 / npp.2013.58
Vanderschuren, LJ, Kalivas, PW (2000). Alterações na transmissão dopaminérgica e glutamatérgica na indução e expressão de sensibilização comportamental: uma revisão crítica de estudos pré-clínicos. Psicofármaco. (Berl) 151 (2-3), 99–120. doi: 10.1007 / s002130000493
Volkow, ND, Wise, RA, Baler, R. (2017). O sistema motriz da dopamina: implicações para a dependência de drogas e alimentos. Nat. Rev. Neurosci. 18, 741-752. doi: 10.1038 / nrn.2017.130
Walker, QD, Rooney, MB, Wightman, RM, Kuhn, CM (2000). A liberação e captação de dopamina são maiores no estriado do sexo feminino do que no masculino, conforme medido por voltametria cíclica rápida. Neuroscience 95, 1061–1070. doi: 10.1016/S0306-4522(99)00500-X
Wee, S., Koob, GF (2010). O papel do sistema opióide da dinorfina-κ nos efeitos reforçadores das drogas de abuso. Psicofármaco. (Berl) 210, 121–135. doi: 10.1007/s00213-010-1825-8
Wee, S., Orio, L., Ghirmai, S., Cashman, JR, Koob, GF (2009). A inibição dos receptores kappa opióides atenuou o aumento da ingestão de cocaína em ratos com acesso prolongado à cocaína. Psicofármaco. (Berl) 205, 565–575. doi: 10.1007/s00213-009-1563-y
Whitfield, TW, Jr., Schlosburg, JE, Wee, S., Gould, A., George, O., Grant, Y., et al. (2015). κ Receptores opióides no núcleo accumbens mediam a escalada da ingestão de metanfetamina. J. Neurosci. 35 (10), 4296-4305. doi: 10.1523 / JNEUROSCI.1978-13.2015
Williams, MT, Mugno, B., Franklin, M., Faber, S. (2013). Dimensões dos sintomas no transtorno obsessivo-compulsivo: fenomenologia e resultados do tratamento com exposição e prevenção ritual. Psicopatologia 46, 365 – 376. doi: 10.1159 / 000348582
Wilson, CJ, Young, SJ, Groves, PM (1977). Propriedades estatísticas dos trens de pico neuronal na substância negra: tipos de células e suas interações. Cérebro Res. 136, 243–260. doi: 10.1016/0006-8993(77)90801-0
Wise, RA (2009). Papéis para dopamina nigrostriatal - não apenas mesocorticolímbica - em recompensa e dependência. Tendências Neurosci. 32, 517 – 524. doi: 10.1016 / j.tins.2009.06.004
Xiao, L., Becker, JB (1994). Determinação quantitativa por microdiálise da concentração extracelular de dopamina estriada em ratos machos e fêmeas: efeitos do ciclo estral e gonadectomia. Neurosci. Lett. 180, 155–158. doi: 10.1016/0304-3940(94)90510-X
Palavras-chave: receptor opióide kappa, dopamina, compulsividade, anfetamina, quinpirol, sensibilização locomotora
Citação: Escobar AdP, Casanova JP, Andrés ME e Fuentealba JA (2020) Crosstalk entre sistemas Kappa Opioid e Dopamine em comportamentos compulsivos. Frente. Pharmacol. 11: 57. doi: 10.3389 / fphar.2020.00057
Recebido: 16 de outubro de 2019; Aceito: 22 de janeiro de 2020;
Publicado: 18 February 2020.
Editado por:
Gonzalo E. Yevenes, Universidade de Concepcion, Chile
Revisados pela:
Luis Geraldo Aguayo, Universidade de Concepcion, Chile
Hugo Tejeda, Instituto Nacional de Abuso de Drogas (NIDA), Estados Unidos
Cecília Scorza, Instituto de Investigações Biológicas Clemente Estable (IIBCE), Uruguai
Copyright © 2020 Escobar, Casanova, Andrés e Fuentealba. Este é um artigo de acesso aberto distribuído sob os termos do Licença Creative Commons Attribution (CC BY). O uso, distribuição ou reprodução em outros fóruns é permitido, desde que o (s) autor (es) original (ais) e o proprietário dos direitos autorais sejam creditados e que a publicação original desta revista seja citada, de acordo com a prática acadêmica aceita. Não é permitida a utilização, distribuição ou reprodução que não esteja em conformidade com estes termos.
* Correspondência: José Antonio Fuentealba, [email protegido]
†ORCID: José Antonio Fuentealba, orcid.org/0000-0003-0775-0675