 Fabrizio Sanna1*,
Fabrizio Sanna1*,  Jessica Bratzu1,
Jessica Bratzu1,  Maria A. Piludu2,
Maria A. Piludu2,  Maria G. Corda2,
Maria G. Corda2,  Maria R. Melis1,
Maria R. Melis1,  Osvaldo Giorgi2 e
Osvaldo Giorgi2 e  Antonio Argiolas1,3
Antonio Argiolas1,3- 1Departamento de Ciências Biomédicas, Seção de Neurociência e Farmacologia Clínica, e Centro de Excelência para a Neurobiologia de Dependências, Universidade de Cagliari, Cagliari, Itália
- 2Departamento de Ciências da Vida e do Meio Ambiente, Seção de Ciências Farmacêuticas, Farmacológicas e Nutracêuticas, Universidade de Cagliari, Cagliari, Itália
- 3Instituto de Neurociência, Conselho Nacional de Pesquisa, Seção Cagliari, Cittadella Universitaria, Cagliari, Itália
Os ratos de raça alta romana (RHA) e de baixa evitação (RLA), que diferem por uma aquisição rápida versus fraca da resposta de evitação ativa na caixa de transporte, exibem diferenças na atividade sexual quando colocadas na presença de uma Rato feminino receptivo. De fato, ratos RHA apresentam níveis mais elevados de motivação sexual e desempenho copulatório do que os ratos RLA, que persistem também após repetidas atividades sexuais. Estas diferenças foram correlacionadas a um tom mais elevado do sistema dopaminérgico mesolímbico de ratos RHA vs. ratos RLA, revelado pelo maior aumento de dopamina encontrado no dialisado obtido a partir do núcleo accumbens de RHA do que ratos RLA durante a actividade sexual. Este trabalho mostra que a dopamina extracelular e a noradrenalina (NA) também aumentam no dialisado do córtex pré-frontal medial (mPFC) de ratos RHA e RLA machos colocados na presença de um rato fêmea inacessível e mais marcadamente durante a interação sexual direta. Tais aumentos em dopamina (e seu metabolito principal ácido 3,4-dihidroxifenilacético, DOPAC) e NA foram encontrados em animais sexualmente ingênuos e experientes, mas foram mais altos: (i) em ratos RHA que em RLA; e (ii) em ratos RHA e RLA sexualmente experientes do que em suas contrapartes ingênuas. Finalmente, as diferenças em dopamina e NA no mPFC ocorreram concomitantemente àquelas em atividade sexual, uma vez que os ratos RHA apresentaram maiores níveis de motivação sexual e desempenho copulatório do que os ratos RLA tanto nas condições sexualmente ingênuas quanto nas experimentadas. Esses resultados sugerem que um tom dopaminérgico mais alto também ocorre no mPFC, juntamente com um aumento do tônus noradrenérgico, que pode estar envolvido nos diferentes padrões copulatórios encontrados em ratos RHA e RLA, como sugerido para o sistema dopaminérgico mesolímbico.
Introdução
As linhas de ratos exóticas romanas (RHA) e de baixa evitação (RLA), originalmente selecionadas para a aquisição rápida e fraca, respectivamente, da resposta de evitação ativa na caixa de transporte (Bignami, 1965; Broadhurst e Bignami, 1965; Driscoll e Bättig, 1982; Fernández-Teruel e outros, 2002; Giorgi et al., 2007) exibem diferenças significativas na atividade sexual quando colocadas na presença de um rato fêmea sexualmente receptivo. De fato, ratos RHA mostram níveis mais altos de motivação sexual do que ratos RLA, como revelado pelo maior número de ereções penianas sem contato mostradas por esses ratos quando colocados na presença de uma fêmea receptiva inacessível, e melhores desempenhos copulatórios do que ratos RLA. pelas diferenças significativas em numerosos parâmetros copulatórios encontrados principalmente no primeiro teste copulatório entre as duas linhagens de ratos, mas também após cinco testes copulatórios, embora atenuados (Sanna et al., 2014a). Entre as diferenças mais pronunciadas encontradas entre os ratos RHA e RLA está a percentagem de ratos que iniciam a montagem e intromissão e que ejaculam no primeiro teste copulatório, 80% de ratos RHA contra 40% de ratos RLA, respectivamente (Sanna et al., 2014a). Um grande corpo de evidências experimentais sugere que um tom dopaminérgico funcionalmente diferente está envolvido nas diferenças de cópula entre ratos RHA e RLA. De fato, os diferentes padrões copulatórios de ratos RHA e RLA foram modificados de forma diferente pela apomorfina, um agonista misto de receptor de dopamina D1 / D2 administrado em doses que facilitam o comportamento sexual, e pelo haloperidol, um antagonista do receptor de dopamina D2 administrado em doses baixas que inibem o comportamento sexual (Sanna et al., 2014b). Assim, os ratos RLA foram mais sensíveis à facilitação e inibição do comportamento sexual induzido por apomorfina e haloperidol, respectivamente, como demonstrado pelas maiores modificações induzidas pelas duas drogas, principalmente quando administradas nas doses mais baixas em diferentes parâmetros copulatórios em Ratos RLA com respeito a ratos RHA (Sanna et al., 2014b). Talvez mais importante para este estudo, os diferentes padrões copulatórios de ratos RHA e RLA também estão relacionados a diferenças na atividade dos neurônios dopaminérgicos mesolímbicos, cuja atividade é bem conhecida por aumentar durante o comportamento sexual (Pfaus et al., 1990; Pleim et al., 1990; Pfaus e Phillips, 1991; Damsma et al., 1992; Wenkstern e outros, 1993; Balfour e outros, 2004; Pitchers et al., 2010, 2013; Beloate et al., 2016). De fato, apesar de ratos RHA e RLA sexualmente ingênuos e sexualmente experientes terem valores basais similares de dopamina extracelular no dialisato do nucleus accumbens, as concentrações de dopamina extracelular e ácido 3,4-dihidroxifenilacético (DOPAC, um dos seus principais metabólitos) ), foram encontrados a aumentar diferencialmente nos dialisados obtidos a partir do shell nucleus accumbens de ratos RHA e RLA virgens e experimentados sexualmente em ambas as fases, antecipatória e consumatória do comportamento sexual. As diferenças acima foram mais marcantes entre os ratos sexualmente ingênuos RHA e RLA, mas persistiram entre ratos RHA e RLA sexualmente experientes, embora tendendo a diminuir nestes ratos, como encontrado com as diferenças no comportamento sexual (Sanna et al., 2015).
O papel funcional do aumento da atividade dopaminérgica no núcleo accumbens observado durante a atividade sexual (seja nas fases apetitiva e consumatória do comportamento sexual) ainda é motivo de debate, assim como o observado durante a alimentação que, como a atividade sexual, tem forte valência motivacional. Assim, embora a dopamina no nucleus accumbens esteja envolvida na motivação e os neurónios dopaminérgicos mesolímbicos sejam habitualmente referidos como neurónios recompensadores, nos últimos anos da 15, estudos recentes apoiam a hipótese de que a dopamina mesolímbica não está envolvida na expressão primária de comportamentos motivados ou recompensadores, mas sim com aprendizagem e memória de associações de estímulo-recompensa (Agmo et al., 1995; Berridge e Robinson, 1998; Ikemoto e Panksepp, 1999; Pitchers et al., 2013, 2014; Beloate et al., 2016; Salamone et al., 2016). Em consonância com essa hipótese, o bloqueio de receptores de dopamina no nucleus accumbens ou a inativação de neurônios dopaminérgicos na área tegmentar ventral foram recentemente encontrados incapazes de alterar a expressão de aspectos apetitivos e consumatórios do comportamento copulatório em ratos machos (Pitchers et al., 2013, 2014; Beloate et al., 2016).
No entanto, um papel de outras áreas do cérebro contendo dopamina nas diferenças acima no comportamento sexual entre ratos RHA e RLA não pode ser descartado. Nesse sentido, sabe-se que a dopamina exerce efeitos facilitatórios sobre as fases antecipatória e consumatória do comportamento sexual em animais de laboratório e também em humanos, não apenas no nucleus accumbens (Everitt, 1990; Pfaus et al., 1990; Hull e outros, 1991; Pfaus e Everitt, 1995; Melis e Argiolas, 2011), mas também em outras áreas do cérebro, como a área pré-óptica medial, o hipotálamo e seus núcleos (isto é, núcleo paraventricular (PVN)); Pfaus e Phillips, 1991; Argiolas e Melis, 1995, 2005, 2013; Hull e outros, 1995, 1999; Melis e Argiolas, 1995; Melis et al., 2003; Succu et al., 2007; Pfaus, 2010). Outra área que contém dopamina e pode desempenhar um papel no comportamento sexual é o córtex pré-frontal medial (mPFC; Fernández-Guasti e outros, 1994; Agmo e Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles e outros, 1998, 2007; Kakeyama et al., 2003; Balfour e outros, 2006; Afonso et al., 2007; Davis et al., 2010; Febo, 2011). De fato, nesta área do cérebro são encontradas as terminações nervosas dos neurônios dopaminérgicos mesocorticais, com seus corpos celulares localizados na área tegmentar ventral como neurônios dopaminérgicos mesolímbicos. Quanto ao nucleus accumbens, o papel exato dessa área cerebral no comportamento sexual está longe de ser claro. De fato, as lesões do mPFC são geralmente encontradas incapazes de alterar o comportamento sexual de ratos machos com uma fêmea sexualmente receptiva (Fernández-Guasti e outros, 1994; Agmo e Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles e outros, 1998, 2007; Kakeyama et al., 2003; Balfour e outros, 2006; Afonso et al., 2007), nem a expressão de preferência de lugar condicionado por recompensa sexual (Davis et al., 2010). No entanto, as lesões mPFC, que não alteraram a aparência de preferência condicionada por recompensa sexual, aboliram nos mesmos animais a capacidade de formar uma aversão condicionada à atividade sexual quando associada a estímulos aversivos (Davis et al., 2010) e queima de células seletivas durante comportamentos de aproximação de um rato macho em direção a uma fêmea sexualmente receptiva inacessível foram medidos no mPFC de ratos machos (Febo, 2011). Essas descobertas levaram a sugerir que a ativação de mPFC durante o comportamento sexual desempenha um papel na integração de informações externas e internas para a execução e controle de comportamentos direcionados por objetivos, e não na expressão de respostas inatas a reforçadores naturais (ver Goto e Grace, 2005). Assim, juntamente com o nucleus accumbens, o mPFC faz parte de um complexo sistema neural envolvido na modulação do comportamento motivado (comportamento direcionado por objetivos), que requer a integração de informações cognitivas do mPFC, informações emocionais da amígdala e contextualização. informação relacionada do hipocampo, no nucleus accumbens (Goto e Grace, 2005).
Evidências experimentais sugerem que a liberação de dopamina no mPFC está envolvida na regulação adaptativa do comportamento motivado, e acredita-se que a desregulação desses mecanismos desempenhe um papel em condições patológicas ou mal-adaptativas, como transtornos psiquiátricos como esquizofrenia, déficit de atenção e hiperatividade ( TDAH), depressão (Dunlop e Nemeroff, 2007; Masana et al., 2011), ou abuso de substâncias e comportamento de jogo (Everitt e Robbins, 2005). Curiosamente, a atividade da dopamina liberada no mPFC pode ser influenciada pela noradrenalina (NA), que está presente no mPFC em níveis mais elevados do que a dopamina e, em particular, pelo transportador de NA (NET), que não é apenas mais abundante do que o transportador de dopamina (DAT) no mPFC (Carboni et al., 1990, 2006; Gresch e outros, 1995; Westernik et al., 1998), mas também mostra uma afinidade pela dopamina ainda maior do que a de NA (Horn, 1973). Uma vez que a dopamina e o NA frequentemente cooperam em muitas funções mPFC, desde a memória de trabalho e formação de conjuntos atencionais e mudança para a aprendizagem reversa, inibição de resposta e resposta ao estresse (ver Robbins e Arnsten, 2009), isso levanta a possibilidade de que a dopamina, sozinha ou em conjunto com NA, no mPFC também pode desempenhar um papel nas diferenças comportamentais entre ratos RHA e RLA, incluindo aqueles encontrados no comportamento sexual.
Para testar esta hipótese, a atividade do sistema dopaminérgico mesocortical e do sistema noradrenérgico no mPFC foi estudada nas duas linhagens de ratos RHA e RLA por meio de microdiálise intracerebral. Resumidamente, a dopamina (e seu principal metabólito DOPAC) e NA foram medidos em dialisados obtidos dos compartimentos pré-límbico (PrL) e infralímbico (IL) do CPMM de RHA sexualmente ingênua (por exemplo, nunca exposta a uma fêmea receptiva) e sexualmente experientes. RLA ratos (por exemplo, que foram submetidos a cinco testes preliminares de cópula e apresentam níveis constantes de atividade copulatória) quando colocados em presença de uma fêmea receptiva inacessível e durante interação sexual direta por cromatografia líquida de alta pressão acoplada com detecção eletroquímica (HPLC-ECD).
Materiais e Métodos
Animais
Ratos forrageiros RHA e RLA (N = 30 para cada linha, pesando ≈300 g no início do trabalho experimental) eram todos da colônia fundada em 1998 na Universidade de Cagliari, Itália (Giorgi et al., 2007). Os procedimentos utilizados para a reprodução selectiva da colónia da Sardenha já foram descritos em pormenor (Giorgi et al., 2005).
Ratos ovariectomizados com estímulo SD SD (250-300g no início do trabalho experimental) utilizados em todos os experimentos, foram obtidos da Envigo (San Pietro al Natisone, Itália). Os animais foram aclimatados quatro por gaiola (38 cm × 60 cm × 20 cm) para as instalações de alojamento do Departamento de Ciências Biomédicas da Universidade de Cagliari por um mínimo de 10 dias antes do início dos experimentos, a 24 ° C, umidade 60%, inverteu o ciclo de luz / escuridão 12 h (luzes apagadas de 08: 00 h para 20: 00 h), água e alimentos de laboratório padrão ad libitum. Os animais foram manipulados diariamente para 1-2 min durante todo o período de habituação, a fim de limitar o estresse de manipulação durante os experimentos; além disso, o contato com o pessoal de manutenção da casa de animais era restrito a um único atendente e a cama nas gaiolas de casa nunca era trocada no dia anterior ou no dia dos experimentos. Todas as experiências foram realizadas entre 10: 00 he 18: 00 h. Este estudo foi realizado de acordo com as recomendações das diretrizes das Comunidades Européias, da Diretiva de setembro 22, 2010 (2010 / 63 / EU) e da Legislação italiana (DL de março 4, 2014, n. 26). O protocolo foi aprovado pelo Comitê de Ética para Experimentação Animal da Universidade de Cagliari (Autorização No. 361 / 2016-PR, abril 08, 2016 para FS).
Grupos Experimentais
Foram utilizados ratos RHA e RLA sexualmente ingénuos e sexualmente experientes. Ratos sexualmente ingênuos foram ratos nunca expostos a ovários sexualmente receptivos-ovariectomizados e estradiol + progesterona-fêmea; ratos sexualmente experientes foram ratos que já foram submetidos a cinco testes de cópula consecutivos de 60 min em intervalos de 3 dias com uma fêmea receptiva (Sanna et al., 2014a,b). As fêmeas foram introduzidas em estro pelo tratamento com benzoato de estradiol subcutâneo (200 μg / rato em óleo de amendoim) e progesterona (0.5 mg / rato em óleo de amendoim), 48 he 6 h antes dos testes de cópula, respectivamente. O estro foi verificado por coloração de May-Grunwald-Giemsa e exame microscópico dos esfregaços vaginais 1 h antes dos experimentos. De acordo com estudos anteriores (Sanna et al., 2014a,b, 2015) cinco testes preliminares de copulação foram suficientes para ter ratos romanos machos de ambas as linha mostrando níveis constantes de atividade copulatória: por exemplo, ratos RHA e RLA sexualmente experientes satisfizeram o critério de pelo menos uma ejaculação alcançada em cada um dos dois últimos testes (um rato RHA e dois ratos RLA que não satisfizeram este critério foram descartados nesta fase). Dois dias após esses testes copulatórios preliminares, ratos romanos sexualmente experientes foram submetidos à cirurgia estereotáxica para o implante de uma sonda de microdiálise no mPFC, conforme descrito a seguir (Sanna et al., 2015).
Microdiálise no mPFC durante o Comportamento Sexual
No dia anterior à microdiálise, ratos RHA e RLA sexualmente ingênuos ou experientes foram posicionados em um aparelho estereotáxico (Stoelting Co., Wood Dale, IL, EUA) sob anestesia com isoflurano (1.5% –2%; Harvard Apparatus, Holliston, MA, EUA) e implantado com sonda de microdiálise vertical caseira (membrana de diálise ≈3 mm de superfície livre; Melis et al., 2003), e dirigida unilateralmente nos compartimentos mPFC, PrL e IL (coordenadas: 3.0 mm anterior e 0.7 mm lateral ao bregma e 5.5 mm ventral à dura-máter; Paxinos e Watson, 2004). No dia da experiência, os animais foram transferidos durante a fase escura do ciclo para a gaiola de acasalamento (45 cm × 30 cm × 24 cm), que foi localizado em uma sala à prova de som iluminada por uma luz vermelha fraca e contendo dentro de outro pequena caixa de acrílico (15 cm × 15 cm × 15 cm) com furos 25 (Ø 2 mm) nas paredes verticais para permitir a interação visual, olfativa e acústica, mas não direta. Após um período de habituação de 2 h, a sonda de microdiálise foi ligada a uma bomba de microinfusão CMA / 100 (Harvard Apparatus, Holliston, MA, EUA) com tubo de polietileno e perfundida com uma solução de Ringer (147 mM NaCl, 3 mM KCl e 1.2 mM CaCl2, pH 6.5), com um caudal de 2.5 μl / min. Após um período de equilíbrio de 2 h do meio de perfusão com o fluido extracelular, foram recolhidas alíquotas de 37.5 μl de dialisado a cada 15 min durante a experiência em tubos de polietileno arrefecido com gelo para a medição da concentração de dopamina, DOPAC e NA, conforme descrito abaixo. Após a recolha de pelo menos quatro aliquotas de dialisado, introduziu-se um rato fêmea receptivo na pequena gaiola localizada no interior da gaiola de acoplamento para 30 min. Durante estes 30 min, foram recolhidas outras duas alíquotas de dialisado. Nessas condições, os ratos machos não podem interagir diretamente com a fêmea, mas mostram ereções sem contato (veja abaixo). Após este período, a pequena gaiola foi removida, a cópula foi permitida para 75 min, e outras cinco alíquotas de dialisado foram coletadas. No final deste período, a fêmea foi então removida da gaiola de acasalamento e uma outra alíquota de dialisato foi coletada (Pfaus e Everitt, 1995; Melis et al., 2003; Sanna et al., 2015). Parâmetros sexuais relacionados às fases antecipatória e consumatória do comportamento sexual foram registrados ao longo do experimento (ver abaixo).
Comportamento Sexual
Vários parâmetros de motivação sexual e desempenho copulatório relacionados às fases antecipatória e consumatória do comportamento sexual foram registrados ao longo do experimento por um observador que não estava ciente das condições experimentais específicas, por exemplo, que não conheciam a linha e o nível da experiência sexual. dos animais utilizados nesse experimento (ver abaixo). Resumidamente, a latência para a primeira ereção sem contato (NCPEL, cronometrada a partir da introdução da fêmea receptiva na pequena gaiola interna) e sua frequência (NCPEF, o número de ereções penianas sem contato que ocorrem no período em que a fêmea é presente na gaiola interna) foram registrados. Estas ereções penianas mediadas por feromônios, que ocorrem em ratos machos sexualmente potentes na presença de uma fêmea receptora inacessível, são consideradas um importante índice de excitação sexual (Sachs et al., 1994; Sachs, 2000; Melis et al., 2003). Quando a interação sexual foi permitida, por exemplo, durante a cópula, a latência para montar e intromitar (ML e IL, cronometrado desde a remoção da pequena gaiola interior até a primeira montagem ou a primeira intromissão, respectivamente); freqüência de montagem e intromissão (MF e IF, o número de montagens e intromissões, respectivamente, na primeira série de atividade copulatória e durante todo o período de copulação); latência à ejaculação (EL, desde a primeira intromissão da primeira série até a ejaculação); frequência de ejaculação (EF, o número total de ejaculações durante o teste de cópula) e intervalo pós-ejaculatório (PEI, cronometrado desde a primeira ejaculação até a próxima intromissão), foram registrados. Além disso, eficácia copulatória (CE; o número de intromissões de uma determinada série dividido pela soma do número de montarias e de intromissões na mesma série) e intervalo inter-intromissão (III; a relação entre a latência da ejaculação de uma dada série e o número de intromissões naquela série) também foram calculados para a primeira série de atividade copulatória (Sachs e Barfield, 1976; Meisel e Sachs, 1994; Melis et al., 2003; Sanna et al., 2014a,b, 2015).
Determinação das concentrações de dopamina, DOPAC e noradrenalina no dialisado do mPFC
As concentrações de dopamina, DOPAC e NA foram medidas em 20 μL da mesma alíquota de dialisato do mPFC por cromatografia líquida de alta pressão (HPLC) acoplada à detecção eletroquímica usando uma célula dupla 4011 (Coulochem II, ESA, Cambridge, MA, EUA) como já descrito (Melis et al., 2003). A detecção foi realizada no modo de redução em + 350 e −180 mV. A HPLC foi equipada com uma coluna Supelcosil C18 (7.5 cm × 3.0 mm id, tamanho de partícula 3 μm; Supelco, Supelchem, Milão, Itália), eluída com citrato 0.06 M / acetato pH 4.2, contendo metanol 20% v / v, 0.1 mM EDTA, 1 μM trietilamina e 0.03 mM dodecil sulfato de sódio como fase móvel, a uma taxa de fluxo de 0.6 mL / min e temperatura ambiente. A sensibilidade do ensaio foi 0.125 pg para dopamina, 0.1 pg para DOPAC e 0.2 pg para NA.
Histologia
No final das experiências, os ratos foram mortos por decapitação, os cérebros foram imediatamente removidos do crânio e imersos em formaldeído aquoso 4% durante os dias 12-15. Após este período, prepararam-se secções cerebrais corais de 40 μm com micrótomo de congelação, coradas com Vermelho Neutro e inspeccionadas num microscópio de contraste de fase. A posição da ponta da sonda foi então localizada no mPFC seguindo o trato da sonda através de uma série de seções do cérebro (veja Figura 1). Apenas ratos considerados como tendo a parte ativa da membrana dialisante posicionada corretamente nos compartimentos PrL e IL do CPFm foram considerados para a avaliação estatística dos resultados (um rato de cada grupo experimental foi descartado nesta fase).
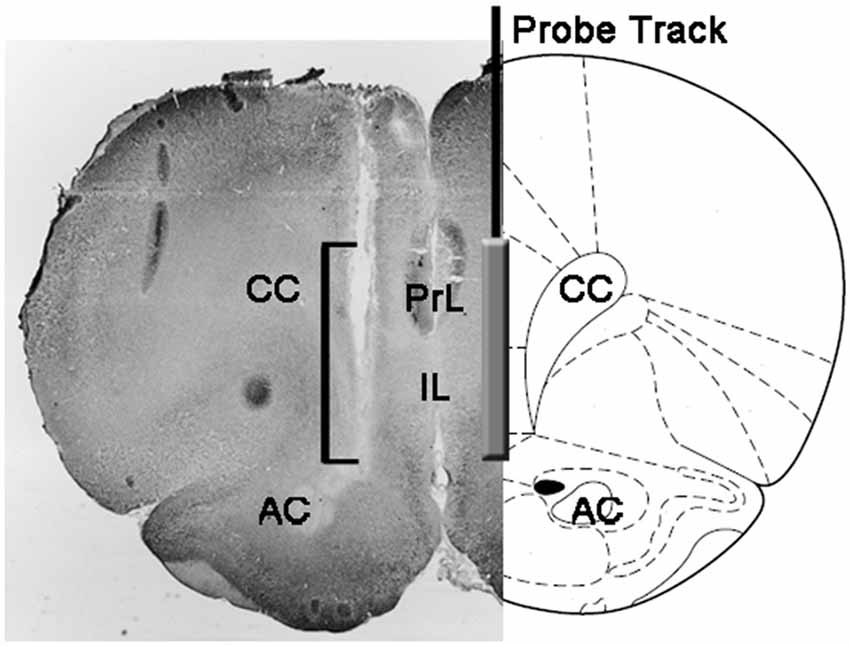 Figura 1. Representação esquemática de uma seção coronal do cérebro de rato mostrando a trilha da sonda de microdiálise nas porções PrL e IL do córtex pré-frontal medial (mPFC; Paxinos e Watson, 2004). O colchete na micro-fotografia indica a parte da secção Neutra vermelha manchada mostrando a parte ativa da membrana dialisante da sonda de microdiálise. Abreviaturas: PrL, área pré-límbica; IL, área infralimbica; CA, comissura anterior; CC, corpo caloso.
Figura 1. Representação esquemática de uma seção coronal do cérebro de rato mostrando a trilha da sonda de microdiálise nas porções PrL e IL do córtex pré-frontal medial (mPFC; Paxinos e Watson, 2004). O colchete na micro-fotografia indica a parte da secção Neutra vermelha manchada mostrando a parte ativa da membrana dialisante da sonda de microdiálise. Abreviaturas: PrL, área pré-límbica; IL, área infralimbica; CA, comissura anterior; CC, corpo caloso.
Estatísticas
Análises estatísticas de dados bioquímicos (dopamina, DOPAC e NA) e comportamentais (NCPEL e NCPEF, ML, IL e EL, MF, IF e EF, e PEI) foram realizadas incluindo todos os sujeitos experimentais (como tradicionalmente feito em estudos de ratos machos). comportamento copulatório) ou excluir aqueles indivíduos que não copularam para ejaculação durante o experimento de microdiálise. No primeiro caso, quando todos os animais foram incluídos nas análises, os animais que não apresentaram NCPE dentro do período em que a fêmea estava inacessível ou não montou ou intrometeu ou ejaculou com a fêmea disponível receberam as respectivas pontuações completas: 1800 s if o macho não exibiu NCPE no 30 min em que a fêmea estava inacessível; 900 s para ML e IL se o macho não montou ou intrometeu com a fêmea disponível; 1800 s para EL se o macho não atingir a ejaculação e 600 s para PEI se o macho não intrometeu após a primeira ejaculação. No segundo caso, os animais que não copularam para ejaculação foram excluídos das análises. Isto foi feito para avaliar se as diferenças no comportamento sexual podem estar correlacionadas com as diferenças nas concentrações de dopamina extracelular, DOPAC e NA no mPFC das linhas romanas de ratos durante as duas fases da atividade sexual, excluindo possíveis interferências confusas devido à inclusão de sujeitos que não apresentaram comportamento sexual, atribuindo-lhes valores fixos para os parâmetros sexuais analisados.
Diversas análises estatísticas foram realizadas com ANOVAs para detectar e melhor caracterizar possíveis diferenças entre as condições experimentais utilizadas. Antes de realizar ANOVAs, os conjuntos de dados de cada uma das diferentes variáveis experimentais foram inspecionados quanto à homogeneidade de variâncias entre os quatro grupos experimentais (feito com todos os animais ou com ratos copulando apenas para ejaculação) com o teste de Bartlett ou Levine dependendo do caso. Quando foram encontradas diferenças significativas nas variâncias, os conjuntos de dados foram transformados em logaritmo (isto é, os valores experimentais de Y foram alterados para os valores de Log Y), novamente verificados quanto à homogeneidade das variâncias e depois analisados por ANOVAs. Resumidamente, os parâmetros comportamentais obtidos antes (por exemplo, com a fêmea inacessível) e durante a primeira série de atividade copulatória (da primeira montada / intromissão à primeira montaria / intromissão após a primeira ejaculação) de ratos RHA e RLA virgens e sexualmente experientes durante a microdiálise foram analisadas por ANOVAs bidirecionais, usando a linha de rato e o nível de experiência sexual entre os fatores dos sujeitos (os valores médios dos parâmetros comportamentais são relatados na tabela 1 e F valores e níveis de significância de análises de ANOVA na Tabela 2).
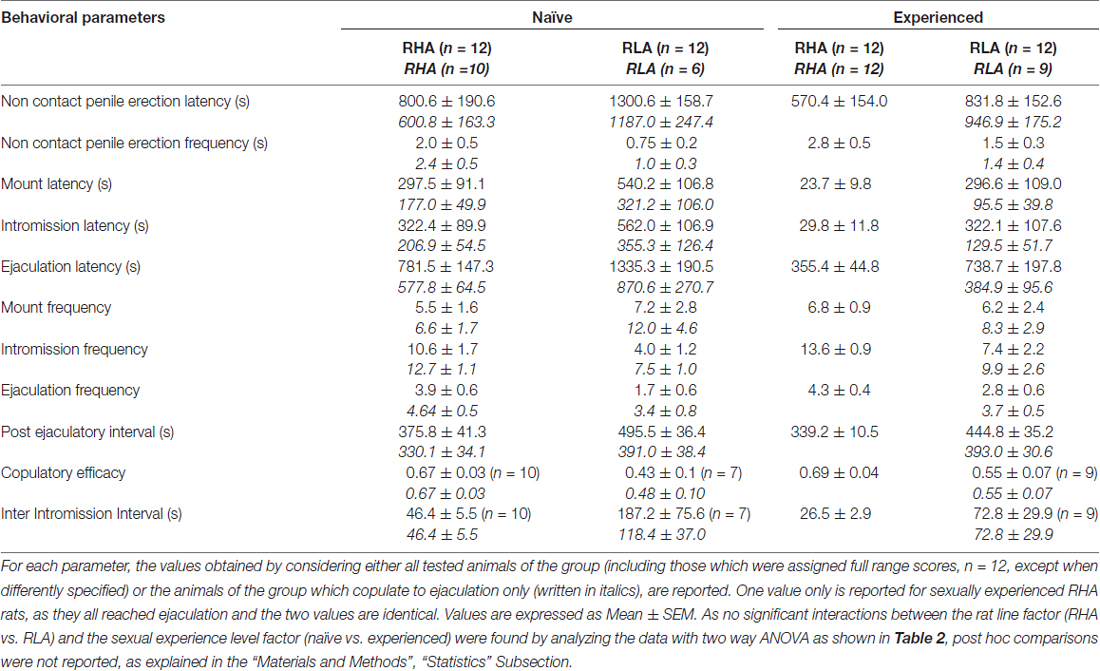 tabela 1. Parâmetros copulatórios medidos na primeira série de atividade copulatória (que começa com a primeira montagem / intromissão quando a fêmea é acessível ao macho e termina após o intervalo pós-ejaculatório com a primeira montagem / intromissão da segunda série) e pênis sem contato ereções de ratos RHA e RLA sexualmente ingênuos e experientes.
tabela 1. Parâmetros copulatórios medidos na primeira série de atividade copulatória (que começa com a primeira montagem / intromissão quando a fêmea é acessível ao macho e termina após o intervalo pós-ejaculatório com a primeira montagem / intromissão da segunda série) e pênis sem contato ereções de ratos RHA e RLA sexualmente ingênuos e experientes.
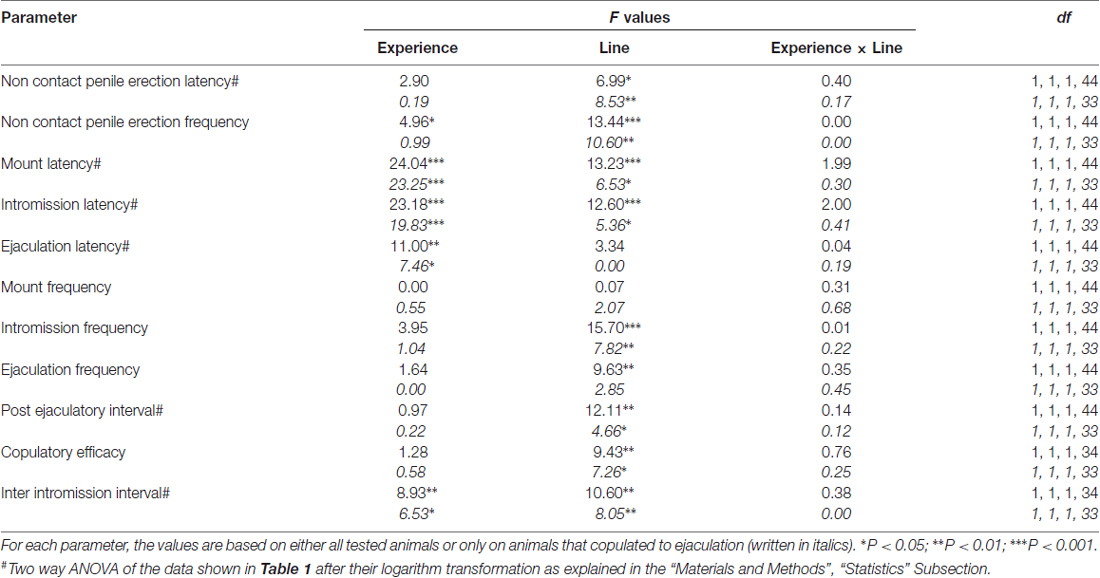 tabela 2. F valores e níveis de significância de ANOVA two-way realizados em dados reportados na Tabela 1 usando a linha de rato (RHA vs. RLA) e o nível de experiência sexual (ingênuo versus experiente) entre os fatores dos sujeitos.
tabela 2. F valores e níveis de significância de ANOVA two-way realizados em dados reportados na Tabela 1 usando a linha de rato (RHA vs. RLA) e o nível de experiência sexual (ingênuo versus experiente) entre os fatores dos sujeitos.
Além disso, uma análise global dos dados obtidos de cada rato durante a microdiálise foi feita calculando-se primeiro as AUCs obtidas pela plotagem dos valores das concentrações de dopamina, DOPAC ou NA ou o número de erecções, montagens, intromissões e ejaculações sem contato vs tempo (iniciando 1 h após a introdução do rato macho na gaiola de acasalamento até o final do experimento - 180 min dividido em frações de 15 min para os valores neuroquímicos - ou no momento em que a fêmea foi introduzida na gaiola de acasalamento para os parâmetros comportamentais) e então comparando os valores calculados por ANOVAs bidirecionais usando a linha de rato e o nível de experiência sexual entre os fatores dos sujeitos (valores médios das AUCs dos parâmetros neuroquímicos e comportamentais são relatados na Tabela 4 e F valores e níveis de significância de análises de ANOVA na Tabela 5). Finalmente, um ponto mais detalhado para apontar a análise de cada conjunto de dados (ou seja, os valores das concentrações de dopamina, DOPAC, NA e os números de ereções sem contato, montes, intromissões e ejaculações versus tempo) foi realizado por fatorial. ANOVAs para medidas repetidas, usando a linha de rato e o nível de experiência sexual entre os fatores do sujeito e o tempo (isto é, frações de dialisato) como dentro do fator de sujeitos (o F valores e níveis de significância dessas análises ANOVA são relatados na Tabela 6). Análises semelhantes foram conduzidas considerando também apenas os valores basais dos parâmetros neuroquímicos (últimas quatro alíquotas de dialisato coletadas antes da introdução na gaiola de acasalamento da fêmea receptora), cujos valores médios são relatados na tabela. 3. Como primeiro, mas não as interações de segunda ordem foram encontradas ao realizar as ANOVAs fatoriais gerais para medidas repetidas, em particular as interações Linha × Tempo e Experiência × Tempo para os parâmetros neuroquímicos (ver Tabela 6), ANOVAs bidirecionais com a linha ou o nível de experiência sexual como fatores entre fator e tempo como dentro do fator sujeito foram realizados nestes conjuntos de dados comparando diretamente ratos sexualmente ingênuos ou experientes RHA vs. RLA, ou experimentados sexualmente vs. ratos RHA naïves ou ratos sexualmente experientes vs. RLA naïves. Os resultados do post hoc contrastes em pares realizados usando o teste HSD de Tukey em interações significativas reveladas por essas ANOVAs de duas vias são relatados na Figura 2. Em todos os outros casos, post hoc comparações não foram relatadas, como ANOVAs não conseguiu revelar qualquer interação significativa entre a linha de rato, o nível de experiência sexual e o tempo. As análises estatísticas foram realizadas com Graph Pad 5 (PRISM, São Diego, CA, EUA) e STATISTICA 12 (Statsoft, Tulsa, EUA), com o nível de significância P <0.05.
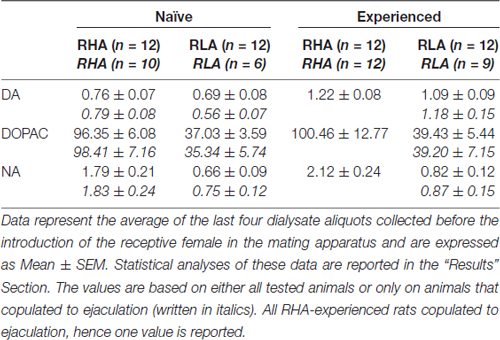 tabela 3. Concentrações de dopamina basal (DA), ácido 3,4-Diidroxifenilacético (DOPAC) e noradrenalina (NA) no dialisado do córtex pré-frontal medial (mPFC) de ratos RHA e RLA sexualmente ingênuos e experientes.
tabela 3. Concentrações de dopamina basal (DA), ácido 3,4-Diidroxifenilacético (DOPAC) e noradrenalina (NA) no dialisado do córtex pré-frontal medial (mPFC) de ratos RHA e RLA sexualmente ingênuos e experientes.
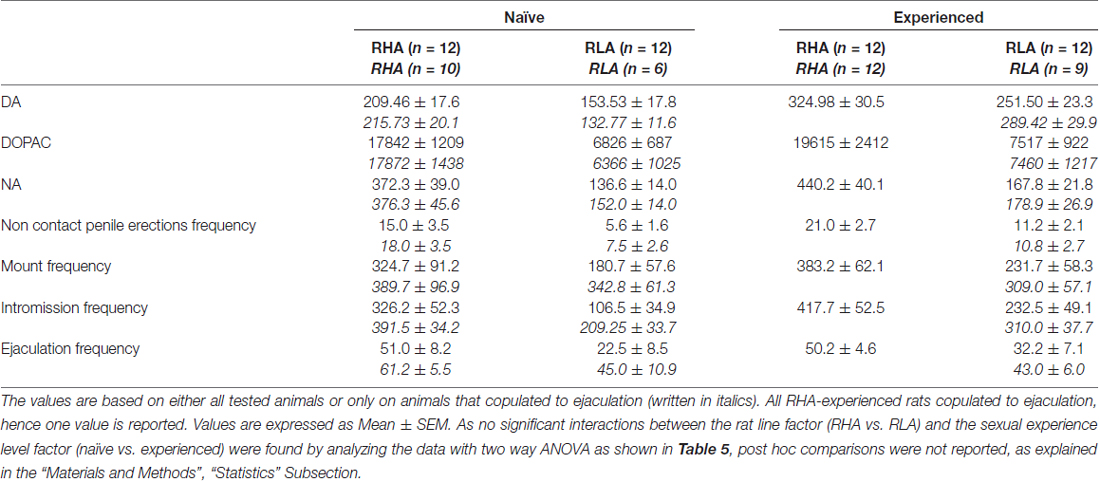 tabela 4. Avaliação global das diferenças nas concentrações de DA, DOPAC e NA e no comportamento sexual entre ratos RHA e RLA sexualmente ingénuos e experientes através da análise das AUC médias obtidas a partir dos resultados apresentados nas Figuras 2 e 3.
tabela 4. Avaliação global das diferenças nas concentrações de DA, DOPAC e NA e no comportamento sexual entre ratos RHA e RLA sexualmente ingénuos e experientes através da análise das AUC médias obtidas a partir dos resultados apresentados nas Figuras 2 e 3.
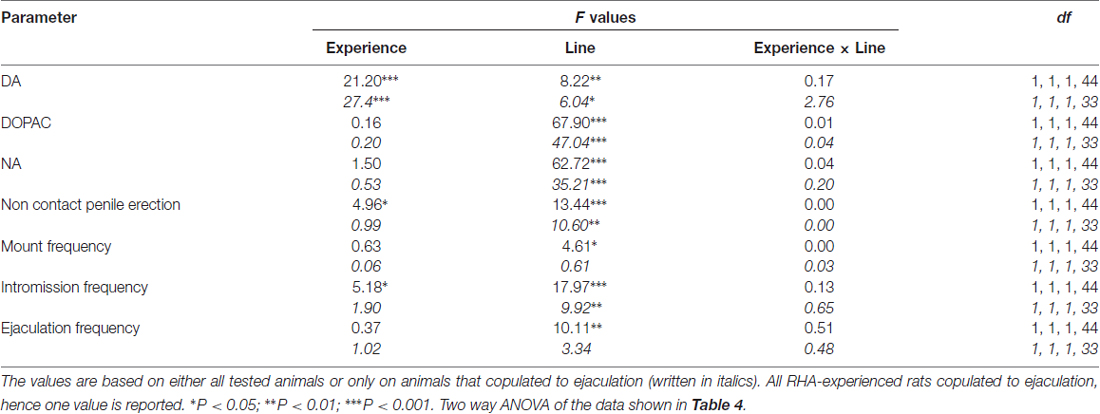 tabela 5. F valores e níveis de significância de ANOVAs bidirecionais realizados em dados relatados na Tabela 4 usando a linha de rato (RHA vs. RLA) e o nível de experiência sexual (ingênuo versus experiente) entre os fatores dos sujeitos.
tabela 5. F valores e níveis de significância de ANOVAs bidirecionais realizados em dados relatados na Tabela 4 usando a linha de rato (RHA vs. RLA) e o nível de experiência sexual (ingênuo versus experiente) entre os fatores dos sujeitos.
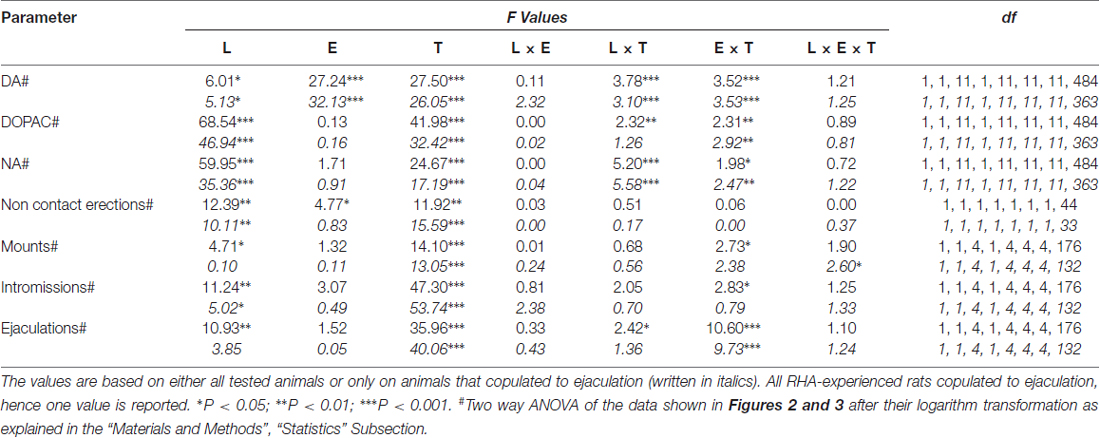 tabela 6. F valores e níveis de significância de ANOVAs fatoriais gerais para medidas repetidas realizadas nos resultados mostrados nas Figuras 2 e 3 usando a linha de rato (L; RHA vs. RLA) e o nível de experiência sexual (E) (ingênuo versus experiente) entre os fatores dos sujeitos e o tempo (T) (frações dialisadas) como dentro do fator sujeito.
tabela 6. F valores e níveis de significância de ANOVAs fatoriais gerais para medidas repetidas realizadas nos resultados mostrados nas Figuras 2 e 3 usando a linha de rato (L; RHA vs. RLA) e o nível de experiência sexual (E) (ingênuo versus experiente) entre os fatores dos sujeitos e o tempo (T) (frações dialisadas) como dentro do fator sujeito.
 Figura 2. Concentrações de dopamina (DA), DOPAC e noradrenalina (NA) nos dialisados mPFC de naïve sexualmente (ÁS) e experientes ratos machos RHA e RLA (B, D, F) durante a atividade sexual com uma mulher receptiva. Sexualmente ingênuos (ou seja, nunca expostos antes a uma mulher sexualmente receptiva) e sexualmente experientes (que foram submetidos a cinco testes de cópula) de ambas as linhas com uma sonda de microdiálise estereotaxicamente implantada no mPFC foram colocados individualmente na gaiola de acasalamento. A sonda de microdiálise foi perfundida com o tampão de diálise, conforme descrito na seção “Materiais e métodos”. Após a coleta de quatro alíquotas de dialisato para a determinação dos valores basais, uma fêmea receptiva inacessível foi então colocada dentro da pequena gaiola do aparato de acasalamento (tempo = 0, inaccess.fe). Após 30 min, a pequena gaiola foi removida e a cópula foi permitida para 75 min (cópula), após o que a fêmea foi removida da gaiola de acasalamento (fe.remov). Durante o experimento, ereções sem contato e parâmetros copulatórios foram medidos, e alíquotas de dialisato coletaram cada 15 min e analisaram para dopamina, DOPAC e NA como descrito na seção “Materiais e Métodos”. Os valores são médias ± SEM dos valores obtidos por todos os ratos 12 por grupo (RHA_all = linhas verdes sólidas, RLA_all = linhas castanhas sólidas) ou aqueles que copularam apenas para ejaculação (RHA_cop = linhas vermelhas tracejadas; RLA_cop = linhas azuis tracejadas). Em ratos RHA experientes, todos os animais atingiram a cópula e os valores são idênticos. *P <0.05 em relação aos valores basais (sem feminino) do grupo (verde para RHA_todos; vermelho para RHA_cop, marrom para RLA_todos, azul para RLA_cop); #P <0.05 em relação aos valores correspondentes do grupo RLA (vermelho, RHA_all vs. RLA_all; verde, RHA_cop vs. RLA_cop); §P <0.05 em relação aos valores correspondentes ao tempo dos ratos sexualmente ingênuos (verde, experiente RHA_all vs. ingênuo RHA_all; vermelho, experiente RHA_cop vs. ingênuo RHA_cop; marrom, experiente RLA_all vs. ingênuo RLA_all; azul, experiente RLA_cop vs. naïve RLA_cop) (ANOVAs de duas vias feitas nos dados mostrados após sua transformação logarítmica, conforme explicado em “Materiais e Métodos”, Subseção “Estatísticas”, seguido pelos testes HSD de Tukey).
Figura 2. Concentrações de dopamina (DA), DOPAC e noradrenalina (NA) nos dialisados mPFC de naïve sexualmente (ÁS) e experientes ratos machos RHA e RLA (B, D, F) durante a atividade sexual com uma mulher receptiva. Sexualmente ingênuos (ou seja, nunca expostos antes a uma mulher sexualmente receptiva) e sexualmente experientes (que foram submetidos a cinco testes de cópula) de ambas as linhas com uma sonda de microdiálise estereotaxicamente implantada no mPFC foram colocados individualmente na gaiola de acasalamento. A sonda de microdiálise foi perfundida com o tampão de diálise, conforme descrito na seção “Materiais e métodos”. Após a coleta de quatro alíquotas de dialisato para a determinação dos valores basais, uma fêmea receptiva inacessível foi então colocada dentro da pequena gaiola do aparato de acasalamento (tempo = 0, inaccess.fe). Após 30 min, a pequena gaiola foi removida e a cópula foi permitida para 75 min (cópula), após o que a fêmea foi removida da gaiola de acasalamento (fe.remov). Durante o experimento, ereções sem contato e parâmetros copulatórios foram medidos, e alíquotas de dialisato coletaram cada 15 min e analisaram para dopamina, DOPAC e NA como descrito na seção “Materiais e Métodos”. Os valores são médias ± SEM dos valores obtidos por todos os ratos 12 por grupo (RHA_all = linhas verdes sólidas, RLA_all = linhas castanhas sólidas) ou aqueles que copularam apenas para ejaculação (RHA_cop = linhas vermelhas tracejadas; RLA_cop = linhas azuis tracejadas). Em ratos RHA experientes, todos os animais atingiram a cópula e os valores são idênticos. *P <0.05 em relação aos valores basais (sem feminino) do grupo (verde para RHA_todos; vermelho para RHA_cop, marrom para RLA_todos, azul para RLA_cop); #P <0.05 em relação aos valores correspondentes do grupo RLA (vermelho, RHA_all vs. RLA_all; verde, RHA_cop vs. RLA_cop); §P <0.05 em relação aos valores correspondentes ao tempo dos ratos sexualmente ingênuos (verde, experiente RHA_all vs. ingênuo RHA_all; vermelho, experiente RHA_cop vs. ingênuo RHA_cop; marrom, experiente RLA_all vs. ingênuo RLA_all; azul, experiente RLA_cop vs. naïve RLA_cop) (ANOVAs de duas vias feitas nos dados mostrados após sua transformação logarítmica, conforme explicado em “Materiais e Métodos”, Subseção “Estatísticas”, seguido pelos testes HSD de Tukey).
Consistentes
Os ratos RHA e RLA mostram um número diferente de ereções sem contato e padrões diferentes de comportamento copulatório
Em consonância com estudos anteriores (Sanna et al., 2014a,b, 2015), um número diferente de ratos RHA e RLA sexualmente ingênuos se envolveu em atividade sexual quando colocados juntos com uma fêmea receptiva durante a coleta de alíquotas de dialisato do mPFC por microdiálise intracerebral. Resumidamente, neste estudo 10 de 12 ratos RHA sexualmente ingênuos (83%) copularam para ejaculação em seu primeiro teste copulatório, contra somente 6 de ratos RNX sem 12 (50%) durante o experimento de microdiálise. Esta diferença foi também encontrada em ratos RHA e RLA machos sexualmente experientes (após cinco testes copulatórios), embora atenuados, com todos os ratos 12 RHA a atingir a ejaculação contra 9 em ratos 12 RLA durante a microdiálise. Sempre de acordo com estudos anteriores, também neste estudo as duas linhas de ratos romanos exibiram diferentes latências e freqüências de ereções penianas sem contato quando expostas a uma fêmea receptiva inacessível, bem como diferentes padrões de comportamento copulatório durante a interação sexual. Essas diferenças foram encontradas quando considerados os dados de todos os animais experimentais de um determinado grupo, independentemente de terem copulado para a ejaculação ou não (por exemplo, atribuindo pontuações completas aos animais que não mostraram o comportamento, ou seja, ratos que não copularam para ejaculação) ou quando se considera os dados dos animais que mostraram o comportamento de um determinado grupo apenas (por exemplo, animais que copularam para a ejaculação; 1). Assim, análises estatísticas dos valores das ereções sem contato (NCPE) e dos parâmetros copulatórios medidos na primeira série de atividade copulatória por ANOVAs de duas vias revelaram diferenças significativas entre as duas linhagens de ratos quando consideramos todos os ratos experimentais ou copularam apenas ratos nos quatro grupos experimentais (ver Tabela 2). De fato, quando os dados obtidos de todos os animais experimentais foram considerados, o número de ereções não contato foi maior e NCPEL, ML, IL e PEI foram significativamente menores em ratos RHA em comparação com ratos RLA. Além disso, o EF e o CE foram maiores, enquanto o III foi significativamente mais curto, em ratos RHA em comparação com ratos RLA. Algumas das diferenças acima entre as duas linhas romanas de ratos tendem a diminuir ou desaparecer com repetidos testes copulatórios. No entanto, algumas dessas diferenças ainda estavam presentes após a estabilização do comportamento sexual pela experiência sexual repetida. Resultados semelhantes foram obtidos quando considerados apenas os valores dos animais dos quatro grupos experimentais que copularam para ejaculação, exceto para a FE (ver Tabelas). 1, 2).
Concentrações basais de dopamina extracelular, DOPAC e noradrenalina em dialisados mPFC de ratos RHA e RLA sexualmente ingênuos e experientes
Nas presentes condições experimentais, as quantidades de dopamina, DOPAC e NA nos dialisados obtidos do mPFC de todos (por exemplo, independentemente de terem copulado para ejaculação ou não) ratos sexualmente ingênuos RHA e RLA, foram 2.32 pg e 2.11 pg para dopamina, 322.56 pg e 124.42 pg para DOPAC e 6.05 pg e 2.24 pg para NA, respectivamente, em 20 μl de dialisado. Quantidades semelhantes foram medidas nos dialisados obtidos do mPFC de ratos RHA e RLA sexualmente experientes (dopamina: 3.73 pg e 3.33 pg, DOPAC: 353.62 pg e 133.32 pg, NA: 7.17 pg e 2.78 pg em ratos RHA e RLA, respectivamente) . Estes valores indicam uma concentração de ≅0.8 – 1.2 nM e ≅95 – 100 nM para dopamina extracelular e DOPAC, respectivamente, e ≅1.8 – 2.2 nM para NA extracelular, no mPFC de ratos RHA, e uma concentração ≅0.8 – 1.2 nM e ≅35 – 40 nM para dopamina extracelular e DOPAC, respectivamente, e ≅0.6 – 0.9 nM para NA extracelular, no mPFC de ratos RLA (Tabela 3). Os valores acima foram obtidos após um período de equilíbrio 2 h do tampão de diálise com o fluido extracelular mPFC. Como a recuperação de dopamina autêntica, DOPAC e NA das sondas de diálise foi estimada em cerca de 20%, as concentrações extracelulares de dopamina, DOPAC e NA podem ser estimadas próximas a ≅4 – 5 nM em ambas as linhas para dopamina, ≅500 e ≅ 200 nM para DOPAC e ≅10 e ≅4 nM para NA no mPFC de ratos RHA e RLA, respectivamente. ANOVA fatorial para medidas repetidas feitas nos valores acima após a transformação do logaritmo, revelou diferenças significativas nos níveis basais de dopamina entre ratos sexualmente ingênuos e sexualmente experientes (F(1,44,132) = 26.05, P <0.001), e nos níveis basais de DOPAC (F(1,44,132) = 63.36, P <0.001) e de NA (F(1,44,132) = 42.14, P <0.001) entre ratos RHA e RLA (últimas quatro amostras coletadas antes da introdução da fêmea na gaiola pequena). Valores semelhantes foram encontrados ao considerar apenas os ratos que copularam para a ejaculação durante o experimento de microdiálise (Tabela 3). Também neste caso os ANOVA fatorial para medidas repetidas feitas nos valores após a transformação do logaritmo revelaram diferenças significativas nos níveis basais de dopamina entre ratos sexualmente ingênuos e sexualmente experientes (F(1,33,99) = 26.97, P <0.001), e nos níveis basais de DOPAC (F(1,33,99) = 42.95, P <0.001) e de NA (F(1,33,99) = 27.63, P <0.001) entre ratos RHA e RLA.
As concentrações de dopamina extracelular, DOPAC e noradrenalina nos dialisados mPFC de ratos sexualmente ingênuos e experientes de RHA e RLA mudam diferencialmente durante a atividade sexual
A presença da fêmea receptiva inacessível na pequena gaiola interna e subsequente interação sexual direta aumentou as concentrações de dopamina extracelular, DOPAC e NA nos dialisados mPFC obtidos de ratos machos RHA e RLA sexualmente ingênuos e experientes, embora com diferenças significativas entre os grupos. duas linhagens de ratos e condições de nível de experiência, quando se consideram todos os animais experimentais ou apenas aqueles que copularam para a ejaculação (Figura 1 2). De facto, ANOVAs de duas vias das AUCs das concentrações de dopamina, DOPAC e NA, mostraram que o conteúdo global de dopamina, DOPAC e NA de todo o teste, foram significativamente mais elevados em ratos RHA em comparação com ratos RLA, e em sexualmente em comparação com ratos ingênuos (ver Tabela 4 para valores e tabelas de AUCs 5 pela F valores e nível de significância). Estas diferenças foram confirmadas por ANOVA fatorial ponto-a-ponto para medidas repetidas de dopamina, DOPAC e NA das duas linhagens de rato sexualmente ingênuas e experimentadas ao longo do experimento, que revelaram efeitos principais significativos de Linha, Experiência, Tempo e Linha Significativa × Tempo e Experiência × Interações de primeira ordem no tempo (consulte a Tabela 6 pela F valores e nível de significância).
As concentrações de dopamina extracelular, DOPAC e noradrenalina nos dialisados mPFC obtidos de ratos sexualmente ingênuos RHA e RLA mudam diferencialmente durante a atividade sexual
Em ratos sexualmente ingênuos, RHA e RLA, a dopamina extracelular, mas não a DOPAC, aumentou primeiro quando colocada na presença da fêmea inacessível, enquanto a NA aumentou apenas em ratos RHA. Em ratos virgens de ratos RHA, a dopamina extracelular aumentou ainda mais no primeiro 15 min de interação sexual como encontrado com DOPAC e NA. Os incrementos em dopamina extracelular, DOPAC e NA duraram por todo o tempo de cópula, com dopamina, DOPAC e NA atingindo um valor máximo em 60 min, 75 min e 75 min, respectivamente, depois dopamina extracelular e, em menor extensão, DOPAC e NA , diminuiu para valores semelhantes aos basais após a remoção da fêmea.
Por outro lado, o primeiro aumento significativo, e o valor de pico, na dopamina extracelular, DOPAC e NA ocorreu em ratos RLA naïve durante o primeiro 15 min de cópula. No entanto, em variância de ratos RHA, em ratos RLA dopamina extracelular, DOPAC e NA tenderam a retornar a valores semelhantes aos basais dentro do primeiro 30 min de interação direta com a fêmea (ver Figuras 2A, C, E).
Finalmente, durante a cópula, os incrementos na dopamina extracelular foram maiores e mais persistentes em ratos com RHA sexualmente ingênua do que em ratos RLA (ver 2A) na ausência da fêmea, as suas concentrações foram semelhantes, embora não idênticas, nas duas linhas de ratos (ver acima e Tabela 3). Curiosamente, embora nenhuma diferença tenha sido encontrada entre as duas linhas romanas no aumento percentual do DOPAC e, em menor grau, nas concentrações de NA durante a presença da fêmea receptiva, uma diferença de mais de duas vezes nos valores absolutos das concentrações de DOPAC e NA foi observada entre as duas linhagens de ratos, tanto em condições basais 3) e durante todo o teste com a fêmea receptiva (ver Figuras 2C, E).
As concentrações de dopamina extracelular, DOPAC e noradrenalina nos dialisados mPFC obtidos de ratos sexualmente experientes, RHA e RLA, mudam diferencialmente durante a atividade sexual
Tal como encontrado em ratos sexualmente ingénuos RHA e RLA, a presença da fêmea inacessível e subsequente interacção sexual directa levou a um aumento de dopamina extracelular e DOPAC no dialisado mPFC obtido de ratos RHA e RLA machos sexualmente experientes. No entanto, também neste caso, foram encontradas diferenças relacionadas à linha de rato, considerando todos os animais experimentais ou, em menor medida, apenas aqueles que copularam para a ejaculação (Figura 1). 2). Em ratos sexualmente experientes, RHA e RLA, o primeiro aumento na dopamina extracelular ocorreu com a fêmea inacessível. Posteriormente, as concentrações de dopamina aumentaram durante a cópula em ambas as linhagens de ratos. As concentrações de dopamina atingiram seus valores de pico após 45-60 min de copulação e diminuíram lentamente em direção aos valores basais no final do teste copulatório (ver figura 2B). Similarmente aos ratos sexualmente ingênuos, as concentrações de dopamina extracelular foram maiores nos ratos RHA sexualmente experientes em comparação aos ratos RLA, principalmente nas alíquotas de dialisato coletadas durante a parte central da fase de copulação (após 30-45 min de cópula). As concentrações de DOPAC aumentaram durante a presença da fêmea inacessível apenas em ratos RHA, enquanto aumentaram de forma semelhante em ambas as linhagens de ratos durante a cópula (valores de pico em 75-90 min), e tenderam a retornar aos valores basais no final do estudo. teste copulatório (ver figura 2D). As concentrações de NA também aumentaram em ambas as linhagens de ratos sexualmente experientes após a introdução da fêmea atingindo os valores máximos em 90 min em ratos RHA e em 45 min em ratos RLA (ver figura 2F). Diferindo da dopamina (ver acima), diferenças semelhantes foram encontradas nos aumentos das concentrações de DOPAC e NA em ratos sexualmente experientes de ambas as linhagens, considerando todos os animais versus animais que atingiram apenas a ejaculação. Entretanto, como encontrado em ratos sexualmente ingênuos, embora os aumentos percentuais das concentrações de DOPAC e NA fossem semelhantes, diferenças significativas nos valores absolutos de DOPAC e NA foram encontradas entre ratos RHA e RLA sexualmente experientes durante todo o teste (ie, antes, durante e depois a presença da fêmea receptiva), com ratos RHA apresentando valores mais elevados do que os ratos RLA (ver Tabela 3 para valores basais e figuras 2D, F para os valores ao longo de todo o teste).
Experiência sexual influencia as mudanças nas concentrações de dopamina e noradrenalina nos dialisados mPFC obtidos de ratos RHA e RLA durante a atividade sexual
Uma comparação das concentrações de dopamina extracelular e NA em ratos RHA e RLA sexualmente ingênuos e experientes mostrou que a experiência sexual mudou a concentração de dopamina extracelular e, em menor escala, de NA em ambas as linhagens de ratos, considerando todos os animais experimentais ou aqueles que copularam apenas para ejaculação (Figuras 2A, B, E, F). Consequentemente, ocorreram diferenças significativas nos valores basais da dopamina entre ratos sexualmente ingênuos e experientes de ambas as linhas (ver Tabela 3) e os valores de dopamina durante a atividade copulatória foram, em geral, mais altos em ratos RHA e RLA sexualmente experientes, em comparação com seus pares ingênuos (ver Figura 2B). Além disso, em ratos experimentados sexualmente, embora o aumento de dopamina encontrado durante a presença da fêmea inacessível seguisse o mesmo padrão temporal observado em ratos sexualmente ingênuos, diferenças no padrão temporal foram encontradas durante a cópula. De fato, neste caso, diferentemente do observado em ratos sexualmente ingênuos RHA (um pico principal em 60 min), dois valores de pico principais foram encontrados, o primeiro após 15 min e o segundo após 45 min de cópula. Após este último aumento, os valores de dopamina tenderam a retornar aos valores basais (ver 2B).
Um quadro semelhante foi encontrado em ratos RLA sexualmente ingênuos versus experientes. De fato, similarmente ao encontrado em ratos RHA, também em ratos RLA experientes foi encontrado um padrão temporal caracterizado por três aumentos principais na dopamina extracelular: o primeiro com a fêmea inacessível e os outros dois durante a cópula, ou seja, após 15 min e 60 min de copulação (ver Figuras 2A, B). Em geral, a experiência sexual parece produzir mudanças mais duradouras no padrão temporal de liberação de dopamina em ambas as linhagens de ratos, quando passam da condição sexualmente ingênua para a condição vivenciada.
Como mostrado nas figuras 2C – F, apenas pequenas diferenças foram detectadas na concentração de DOPAC e NA entre ratos ingênuos e experientes de ambas as linhagens. No entanto, uma tendência não significativa na NA extracelular para valores mais altos em ratos experientes em comparação com ratos ingênuos foi observada na fase basal (antes da introdução da fêmea receptiva, ver Tabela 3) e nas quantidades totais de NA, conforme revelado pelos valores das AUCs (ver 4) calculado em concentrações extracelulares obtidas a partir de todo o teste (antes e durante a presença da fêmea receptiva). Finalmente, o padrão temporal das concentrações de NA foi muito semelhante ao comparar ratos experientes de ambas as linhagens de ratos com os seus congêneres ingênuos, indicando que a experiência sexual pode levar a um aumento geral no NA extracelular, em vez de mudanças no padrão de seu aumento. 2E, F).
Alterações nas Concentrações de Dopamina Extracelular e Noradrenalina nos Diálise de mPFC Obtidas a partir de RHAs Sexualmente Ingênuos e Experientes e RLA Ocorrem Concomitantemente com Alterações no Comportamento Sexual
As diferenças nas concentrações de dopamina extracelular, DOPAC e NA nos dialisados obtidos a partir do mPFC encontrados em ambos os ratos RHA e RLA sexualmente ingênuos e experientes durante o teste (Figura 2) ocorreu juntamente com modificações nos diferentes parâmetros sexuais medidos durante as duas fases principais dos experimentos (ie, inacessíveis e disponíveis ao homem, respectivamente), que incluem, entre outras, ereções penianas sem contato (Figuras 3A, B), montagens (Figuras 3C, D), intromissões (Figuras 3E, F) e ejaculações (Figuras 3G, H). Essas diferenças foram encontradas quando se considera todos os animais experimentais ou aqueles que copularam apenas para a ejaculação. De fato, uma análise preliminar dos valores das AUCs desses parâmetros por ANOVAs bidirecionais (ver Tabela 4 para valores e tabelas de AUCs 5 pela F valores e nível de significância) seguido por uma análise ponto a ponto dos valores desses parâmetros ao longo do experimento por ANOVA fatorial para medidas repetidas revelou efeitos principais significativos de Linha, Experiência, Tempo, Interações significativas de primeira ordem Linha × Tempo e Experiência × Tempo, e uma interação significativa de segunda ordem para montagens (veja Tabela 6 pela F valores e nível de significância). Por conseguinte, de acordo com estudos anteriores (Sanna et al., 2015): (i) ratos sexualmente ingênuos RHA mostraram mais ereções, montes, intromissões e ejaculações penianas do que suas contrapartes de RLA; (ii) ratos RHA sexualmente experientes mostraram um número maior de ereções penianas sem contato, intromissões e ejaculações do que suas contrapartes RLA; e (iii) ratos sexualmente experientes de ambas as linhas, e em particular ratos RLA, mostraram mais ereções penianas sem contato, intromissões e ejaculações do que suas contrapartes sexualmente ingênuas. De fato, enquanto em ratos RHA a maior parte das mudanças produzidas pela experiência sexual ocorreu no curso do tempo e não na freqüência dos parâmetros comportamentais (por exemplo, ratos experientes copularam principalmente na primeira metade do teste, enquanto ratos virgens mostraram uma tendência copulatória semelhante ao longo do teste), em ratos RLA, uma melhora geral mais marcante do comportamento sexual, principalmente no desempenho sexual, foi produzida pela experiência sexual, revelada pelas mudanças nos valores dos parâmetros copulatórios em ratos RLA experientes contrapartes ingênuas.
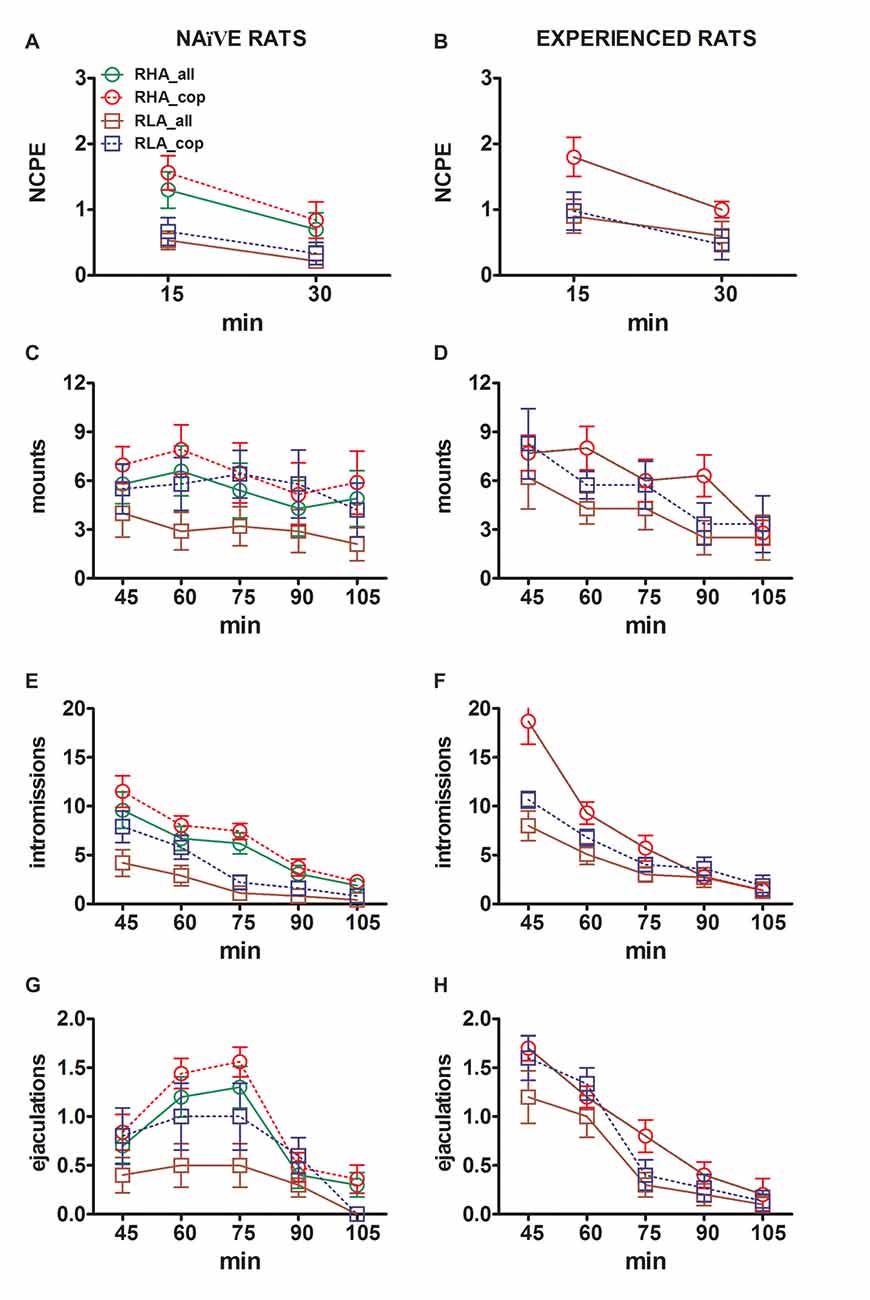 Figura 3. Alterações no número de ereções penianas sem contato (NCPE), montagens, intromissões e ejaculações registradas de crianças sexualmente ingênuas (A, C, E, G) e experientes ratos RHA e RLA (B, D, F, H) durante os experimentos de microdiálise relatados na Figura 2. Todas as condições experimentais foram idênticas às descritas na legenda da Figura 2. Os valores são médias ± DPM dos valores obtidos por todos os ratos 12 por grupo (incluindo aqueles que foram atribuídos pontuações a tempo inteiro; RHA = linhas verdes sólidas, RLA = linhas castanhas sólidas) ou aqueles que copularam apenas para ejaculação (RHA = linhas vermelhas tracejadas ; RLA = linhas azuis tracejadas). Em ratos RHA experientes, todos os animais atingiram a cópula e os valores são idênticos. Como não houve interações significativas entre o fator de linha de rato (RHA vs. RLA), o fator de nível de experiência sexual (ingênuo versus experiente) e o tempo foram encontrados analisando os dados com ANOVAs bidirecionais como mostrado na Tabela 6, post hoc comparações não foram relatadas, conforme explicado na subseção “Materiais e métodos”, “Estatística”.
Figura 3. Alterações no número de ereções penianas sem contato (NCPE), montagens, intromissões e ejaculações registradas de crianças sexualmente ingênuas (A, C, E, G) e experientes ratos RHA e RLA (B, D, F, H) durante os experimentos de microdiálise relatados na Figura 2. Todas as condições experimentais foram idênticas às descritas na legenda da Figura 2. Os valores são médias ± DPM dos valores obtidos por todos os ratos 12 por grupo (incluindo aqueles que foram atribuídos pontuações a tempo inteiro; RHA = linhas verdes sólidas, RLA = linhas castanhas sólidas) ou aqueles que copularam apenas para ejaculação (RHA = linhas vermelhas tracejadas ; RLA = linhas azuis tracejadas). Em ratos RHA experientes, todos os animais atingiram a cópula e os valores são idênticos. Como não houve interações significativas entre o fator de linha de rato (RHA vs. RLA), o fator de nível de experiência sexual (ingênuo versus experiente) e o tempo foram encontrados analisando os dados com ANOVAs bidirecionais como mostrado na Tabela 6, post hoc comparações não foram relatadas, conforme explicado na subseção “Materiais e métodos”, “Estatística”.
Discussão
Este estudo confirma e amplia os achados de estudos anteriores mostrando que ratos RHA e RLA, que apresentam estilos de coping marcadamente diferentes em resposta a condições aversivas (Driscoll e Bättig, 1982; Giorgi et al., 2003a; Steimer e Driscoll, 2003), apresentam padrões comportamentais distintos na presença de uma mulher receptiva inacessível e durante testes copulatórios clássicos (Sanna et al., 2014a,b, 2015). Assim, também este estudo mostra que ratos sexualmente ingênuos RHA (nunca expostos a uma fêmea receptiva antes) exibiram maior motivação sexual e melhor desempenho sexual em comparação com ratos sexualmente ingênuos RLA, por exemplo, ratos RHA mostram maior número de ereções penianas sem contato (feromônio). ereções penianas induzidas consideradas um índice de excitação sexual; Sachs et al., 1994; Sachs, 2000) e diferentes mudanças pró-sexuais em vários parâmetros copulatórios como encontrado em estudos prévios (Sanna et al., 2014a,b, 2015). Essas diferenças ainda persistiram em ambas as linhas de ratos romanos após a aquisição da experiência sexual, ou seja, após a estabilização do comportamento sexual com cinco exposições e testes de cópula com uma fêmea receptiva. Este estudo também mostra pela primeira vez que nas linhagens de ratos RHA e RLA, na condição de ingênua ou experiente, ereções penianas sem contato e interação sexual, ocorreram concomitantemente a um aumento nas concentrações de dopamina extracelular (e seu principal metabólito DOPAC). ) e NA nos dialisados obtidos por microdiálise intracerebral a partir do mPFC de ambas as linhas de ratos romanas. Curiosamente, estes aumentos na dopamina extracelular e NA no mPFC parecem estar relacionados com diferenças quer no fenótipo da ratazana quer no nível da experiência sexual. Consequentemente, como mostrado extensivamente na secção Resultados, tanto os ratos RHA sexualmente ingénuos como os experientes apresentaram aumentos superiores e prolongados da dopamina extracelular e NA, quando comparados com os seus homólogos de RLA, tanto na fase apetitiva como na fase consumatória do comportamento sexual. Além disso, os ratos sexualmente experientes de ambas as linhas romanas apresentaram aumentos mais altos e duradouros da dopamina extracelular e NA no mPFC em ambas as fases do comportamento sexual em comparação com suas contrapartes ingênuas. Finalmente, todas as diferenças acima em dopamina extracelular e NA no mPFC foram aparentemente relacionadas a diferenças nos parâmetros comportamentais do comportamento sexual, por exemplo, ratos sexualmente experientes de ambas as linhas romanas apresentaram maior motivação sexual e melhor desempenho copulatório quando comparados com seus pares ingênuos. e ratos RHA sexualmente ingênuos e experientes demonstraram maior motivação sexual e melhores desempenhos copulatórios quando comparados com suas contrapartes de RLA sexualmente ingênuas e experientes.
Quanto aos aumentos na dopamina extracelular encontrados no dialisado mPFC durante a atividade sexual, o primeiro aumento ocorreu no primeiro 15 min após a introdução da fêmea sexualmente receptiva no aparelho de acasalamento, quando a fêmea era inacessível ao macho, o que geralmente mostra nesta condição não contato ereções penianas. Um segundo e maior aumento na dopamina extracelular ocorreu durante o primeiro 15 min depois que a fêmea ficou acessível ao macho para a cópula. Curiosamente, enquanto na condição ingênua a dopamina extracelular aumentou principalmente na primeira parte do teste com a mulher acessível com tendência a retornar aos valores basais na segunda parte, na condição sexualmente experiente a dopamina permaneceu elevada durante todo o teste atingindo valores máximos após 45-60 min de copulação em ambas as linhas de rato romano.
Além das diferenças acima na dopamina extracelular durante as diferentes fases da atividade sexual entre ratos RHA e RLA na condição sexualmente ingênua e experiente, este estudo também mostra diferenças inesperadas importantes e altamente significativas na concentração de DOPAC extracelular entre as duas linhas romanas de ratos antes (ie , valores basais) e após a introdução da fêmea receptiva no aparato de acasalamento. Consequentemente, os valores basais de DOPAC extracelular no dialisado mPFC foram encontrados em ratos RHA em cerca de 2.6 vezes superiores aos encontrados em ratos RLA. Esta diferença entre ratos RHA e RLA ocorreu e persistiu nas condições sexualmente ingênuas e experientes, apesar de diferenças muito modestas nos valores basais de dopamina extracelular nos dialisados mPFC obtidos a partir das duas linhas de ratos romanas nestas condições experimentais. Em conjunto, essas diferenças podem indicar um turnover de dopamina diferente no nível do mPFC entre as duas linhas de ratos romanas. Consequentemente, as concentrações de DOPAC geralmente refletem a quantidade de dopamina liberada e depois recapturada pelos terminais nervosos dopaminérgicos e convertida em DOPAC pela monoamino-oxidase (MAO; Carlsson, 1975). Assim, as maiores concentrações basais de DOPAC encontradas em ratos RHA sugerem que quantidades maiores de dopamina são liberadas e recapturadas por terminações nervosas dopaminérgicas no mPFC de ratos RHA, por exemplo, uma maior atividade basal do sistema dopaminérgico mesocortical de ratos RHA quando comparado ao RLA. ratos. Um tom dopaminérgico mesocortical mais elevado em ratos RHA do que em ratos RLA é também suportado pelas maiores concentrações de dopamina extracelular encontradas durante a actividade sexual no dialisado mPFC de ratos RHA sexualmente ingénuos e experientes em relação às suas contrapartes de RLA. A este respeito, é pertinente recordar que um tom dopaminérgico mais elevado é considerado responsável, pelo menos em parte, de muitas das características comportamentais diferentes e mesmo opostas presentes nos ratos RHA e RLA. Assim, os ratos RHA são copers ativos, altamente impulsivos, novatos e que gostam de sensações e são propensos a ingerir e abusar de várias classes de drogas de vício, enquanto os RLA são reativos, hiperemocionais e propensos a desenvolver sintomas depressivos (Zeier et al., 1978; Giorgi et al., 1994, 2003b, 2007; Corda e outros, 1997, 2014; Escorihuela et al., 1999; Steimer e Driscoll, 2003; Lecca et al., 2004; Giménez-Llort e outros, 2005; Carrasco e outros, 2008; Fattore e outros, 2009; Moreno e outros, 2010; Coppens e outros, 2012; Díaz-Morán e outros, 2012; Sabariego e outros, 2013; Manzo et al., 2014a,b; Oliveras et al., 2015), incluindo a maior motivação sexual e melhor desempenho copulatório de ratos RHA sexualmente ingênuos e experientes vs. ratos RHA (Sanna et al., 2015). O maior aumento nas concentrações de dopamina extracelular encontradas no dialisato do mPFC de ratos RHA sexualmente ingênuos e experientes se assemelha às maiores concentrações extracelulares de dopamina encontradas no dialisato do núcleo accumbens de ratos RHA sexualmente ingênuos e experientes em relação às suas contrapartes de RLA (Sanna et al., 2015). No entanto, em relação ao mPFC, os níveis basais de dopamina extracelular e DOPAC no dialisado do nucleus accumbens de ratos RHA e RLA sexualmente ingênuos e experientes não diferiram significativamente (Sanna et al., 2015). Assim, o maior tônus dopaminérgico mesolímbico no nucleus accumbens de ratos RHA sexualmente ingênuos e experientes pode ser inferido apenas pelas maiores concentrações de dopamina extracelular encontradas nos dialisados do núcleo accumbens de ratos RHA quando comparados aos homólogos de RLA na presença do receptor receptivo. feminino, e não por uma atividade dopaminérgica basal diferente, revelada pelos níveis basais superior e inferior da concentração de DOPAC em ratos RHA e ratos RLA, respectivamente, como ocorrido no mPFC neste estudo. Isto também está de acordo com os resultados de relatórios anteriores mostrando que os níveis extracelulares de dopamina no dialisato do mPFC de ratos RHA foram encontrados mais elevados do que os ratos RLA, enquanto níveis semelhantes de dopamina foram encontrados no dialisato da casca do núcleo accumbens de ratos RHA e RLA sob diferentes condições experimentais (D'Angio et al., 1988; Scatton et al., 1988; Willig et al., 1991; Giorgi et al., 2003a, 2007). Apesar das diferenças acima entre o CPFm e o nucleus accumbens, a experiência sexual parece produzir no CPFM aumentos mais duradouros no padrão temporal de liberação de dopamina em ambas as linhagens de ratos romanos ao passar da condição sexualmente ingênua para a experimentada, embora tais aumentos eram geralmente mais elevados em ratos RHA do que em ratos RLA. Isto é um pouco diferente do encontrado no nucleus accumbens, onde a experiência sexual tende a alterar os aumentos de dopamina extracelular para a primeira parte do teste copulatório em ratos RHA, enquanto aumenta ao longo de todo o teste de cópula em ratos RLA (Sanna et al., 2015).
Até onde sabemos, este estudo também mostra pela primeira vez que não apenas a dopamina extracelular, mas também o NA extracelular aumenta durante a atividade sexual no dialisato do mPFC de ambos os ratos RHA e RLA, como encontrado com a dopamina, mas também neste caso com diferenças relacionadas à linha romana do rato e à experiência sexual. Em particular, em variância dos níveis de dopamina basal que foram encontrados muito semelhantes no dialisado do mPFC de ambos os ratos RHA e RLA, os níveis basais de NA extracelular no dialisado do mPFC de ratos RHA foram significativamente maiores (cerca de 2,5 vezes ) do que os ratos RLA. As razões para estas diferenças são desconhecidas, mas como discutido acima para dopamina e DOPAC, estes resultados podem indicar que existe um tom noradrenérgico basal superior pelo menos no córtex mPFC de ratos RHA quando comparado com ratos RLA. Um tal tónus noradrenérgico superior pode também estar envolvido nas características comportamentais diferentes e muitas vezes opostas presentes em ratos RHA e RLA, como discutido acima para a dopamina. Suporte adicional para um tom noradrenérgico mais alto em ratos RHA que em ratos RLA no mPFC vem dos resultados que mostram que as diferenças nos níveis extracelulares basais de NA persistiram também durante a atividade sexual em condições sexualmente ingênuas e experientes, por exemplo, níveis extracelulares de NA aumentados em o dialisado mPFC em ratos RHA quando expostos à fêmea inacessível e durante todo o período de copulação muito mais do que em ratos RLA, em que os níveis de NA aumentaram apenas no primeiro 15 do período de cópula, após o qual os níveis de NA retornaram aos valores basais . Essas mudanças nos níveis extracelulares de NA foram observadas tanto na condição sexualmente ingênua quanto na experiência, mas os aumentos nos níveis de NA foram mais altos nos ratos com experiência em RHA e RLA sexualmente, em comparação com seus pares sexualmente ingênuos. Assim, a experiência sexual aparentemente induz uma potenciação adicional da atividade noradrenérgica no mPFC de ratos RHA e, em menor grau, de ratos RLA, como já discutido para a dopamina. Os concomitantes tons noradrenérgicos e dopaminérgicos superiores no mPFC de ratos RHA quando comparados com ratos RLA merecem algum comentário. De fato, foi relatado que a atividade noradrenérgica no mPFC pode afetar a liberação de dopamina através de mecanismos diferentes (por exemplo, ver Carboni et al., 1990, 2006; Gresch e outros, 1995; Westernik et al., 1998 e referências nele). Uma delas está relacionada à atividade da NET, que mostrou ligar a dopamina com uma afinidade ainda maior àquela para NA em sinaptossomas obtidos a partir de homogenatos do PFC (Horn, 1973). Se isso ocorrer no mPFC in vivo, a dopamina liberada no mPFC poderia ser recapturada não apenas por terminais dopaminérgicos, mas também por nervos noradrenérgicos, a uma velocidade ainda maior que a do NA. Isso poderia causar, por sua vez, um aumento na liberação de dopamina dos terminais dopaminérgicos (por exemplo, um aumento do tônus dopaminérgico) para compensar a quantidade de dopamina removida da fenda sináptica; assim, o maior tónus noradrenérgico encontrado em ratos RHA poderia contribuir, pelo menos em parte, para o maior tónus dopaminérgico encontrado no mPFC de ratos RHA quando comparado com ratos RLA. Mais estudos são necessários para verificar essa possibilidade. Notavelmente, a diferença na atividade noradrenérgica identificada neste estudo no mPFC entre ratos RHA e RLA também pode desempenhar um papel em sua resposta diferente ao tratamento com antidepressivos. De fato, os ratos RHA são considerados um modelo de resiliência e os ratos RLA um modelo de vulnerabilidade à depressão. Assim, o tratamento agudo ou crônico com antidepressivos como a desipramina, fluoxetina e clorimipramina, melhorou significativamente as respostas comportamentais no teste de natação forçada em ratos RLA sem afetar as respostas de ratos RHA (Piras et al., 2010, 2014).
Finalmente, este estudo confirma e amplia descobertas anteriores mostrando que o mPFC está envolvido no controle do comportamento sexual, embora o papel exato dessa área do cérebro no comportamento sexual esteja longe de ser claro (Fernández-Guasti e outros, 1994; Agmo e Villalpando, 1995; Agmo et al., 1995; Hernández-Gonzáles e outros, 1998, 2007; Kakeyama et al., 2003; Balfour e outros, 2006; Afonso et al., 2007; Davis et al., 2010; Febo, 2011). Recentemente, a queima seletiva de células no mPFC durante a aproximação de comportamentos de um rato macho em direção a uma fêmea sexualmente receptiva inacessível foi mostrada usando um paradigma de queima de célula única (Febo, 2011). Curiosamente, os neurônios que não responderam durante a primeira exposição do macho à mulher receptiva inacessível, tornam-se ativos durante a segunda exposição, como se a experiência anterior fosse capaz de induzir mudanças plásticas levando à variação nas respostas comportamentais. No entanto, as lesões com ácido ibotênico do mPFC foram encontradas incapazes de alterar o comportamento sexual de ratos machos com uma mulher sexualmente receptiva, nem a expressão de preferência condicionada pelo lugar para a recompensa sexual. No entanto, a capacidade de formar aversão condicionada à atividade sexual quando emparelhada com estímulos aversivos foi encontrada completamente abolida nestes animais lesionados (Davis et al., 2010). Isso sugere que a ativação do mPFC durante o comportamento sexual desempenha um papel na integração de informações externas e internas para a execução e controle de comportamentos direcionados por objetivos, e não na expressão de respostas inatas a reforçadores naturais (ver Goto e Grace, 2005). Mais importante para este trabalho, vários estudos mostraram que as áreas cerebrais envolvidas na motivação sexual e comportamento sexual, como núcleo accumbens, área tegmentar ventral, área pré-óptica medial, núcleo leito da estria terminal, amígdala basolateral e núcleo talâmico parafocular sub-facascular recebem insumos. do mPFC durante a atividade sexual (ver Balfour e outros, 2006). Muitas dessas áreas do cérebro participam de um complexo circuito neural envolvido no controle do comportamento sexual, da motivação sexual e recompensa ao desempenho sexual. Este circuito inclui neurônios oxitocinérgicos que se originam no PVN do hipotálamo (PVN) e se projetam para a área ventral tegmentar, núcleo accumbens, hipocampo, amígdala, PFC, núcleo leito da estria terminal, medula oblonga e medula espinhal mesolímbica / dopaminérgica mesocortical neurônios que se projetam da área tegmentar ventral para o núcleo accumbens e PFC, neurônios dopaminérgicos incertohipotalâmicos e neurônios glutamatérgicos que participam no nível local e do sistema em diversas áreas do circuito Melis e Argiolas, 1995, 2011; Melis et al., 2003, 2007, 2009, 2010; Succu et al., 2007, 2008, 2011). Assim, o aumento da dopamina extracelular (mas também de NA) nos dialisados do CPFM observado durante a atividade sexual pode indicar que o CPFm é uma parte ativa deste circuito, ou seja, os neurônios dopaminérgicos mesocorticais participam do controle da atividade sexual em conjunto com neurônios noradrenérgicos, dopaminérgicos mesolímbicos, oxitocinérgicos e glutamatérgicos, embora não pareça estritamente necessário para a expressão do comportamento sexual (Davis et al., 2010). De acordo com esta hipótese, este estudo mostra que as diferenças na liberação de dopamina e NA no CPFM durante a atividade sexual ocorreram concomitantemente com diferenças no número de ereções penianas sem contato e mudanças nos parâmetros copulatórios, com maior liberação de dopamina e NA durante as fases apetitiva e consumatória relacionadas à maior motivação sexual e melhor desempenho copulatório tanto nas linhas romanas quanto nas condições sexualmente ingênuas e experientes.
A discussão acima é baseada principalmente no aumento bem conhecido da dopamina extracelular que ocorre no núcleo accumbens de ratos machos durante a atividade sexual (Pfaus et al., 1990; Pleim et al., 1990; Pfaus e Phillips, 1991; Damsma et al., 1992; Wenkstern e outros, 1993). No entanto, a dopamina e outros neurotransmissores podem ser liberados no mPFC (e no nucleus accumbens) em outras condições experimentais, como durante o estresse ou lidar com ele (Thierry e outros, 1976; Fadda et al., 1978), que não estão estritamente relacionados ao comportamento sexual. Assim, pode-se argumentar que o aumento de dopamina encontrado na dopamina extracelular no mPFC (este estudo) ou no nucleus accumbens (Sanna et al., 2015), está mais relacionada ao estresse secundário, ou ao enfrentamento da presença e / ou interação com a mulher sexualmente receptiva do que à atividade sexual. Isso pode ter alguma relevância em ratos RHA e RLA, que mostram marcadamente diferentes estilos de enfrentamento e diferentes ativação do eixo hipotálamo-hipófise-adrenal em resposta a condições aversivas (ratos RLA mostram uma ativação deste eixo muito maior que a de ratos RHA; Carrasco e outros, 2008; Díaz-Morán e outros, 2012). Isso também aumenta a possibilidade de que os hormônios do estresse (isto é, corticosteróides) possam influenciar a atividade dopaminérgica no mPFC e em outras áreas cerebrais também. De fato, os ratos RLA são copers reativos e apresentam comportamento hiperemocional caracterizado por hipomotilidade e congelamento, enquanto os ratos RHA mostram um comportamento proativo de enfrentamento visando obter controle sobre o estressor (Driscoll e Bättig, 1982; Willig et al., 1991; Escorihuela et al., 1999; Steimer e Driscoll, 2003; Giorgi et al., 2007) Embora seja impossível excluir completamente que isso possa ocorrer em ratos sexualmente virgens (que nunca interagiram com uma fêmea receptiva antes) e em ratos RLA sexualmente virgens em particular (veja acima), isso é improvável. Em nossas condições experimentais, todo o possível é feito para evitar todo tipo de estresse (ver seção “Materiais e Métodos”) e os experimentos são organizados de forma que os machos não interajam imediatamente com a fêmea, mas somente após 30 min de exposição a ela , durante o qual o aumento medido na dopamina é muito menor do que o que ocorre durante a interação sexual. Se o aumento de dopamina encontrado no mPFC (e no nucleus accumbens) de ratos romanos (ratos RHA e RLA sexualmente virgens têm níveis basais de dopamina extracelular no dialisado do mPFC e no nucleus accumbens, muito semelhantes) eram realmente secundários aos estresse devido a, ou o enfrentamento da presença / interação com a fêmea: (i) o aumento de dopamina teria sido encontrado muito maior no primeiro período de tempo quando os machos são colocados na presença da fêmea e não mais tarde, como encontrado durante a cópula; e (ii) seria esperado que o aumento de dopamina desapareça ou seja fortemente reduzido após a aquisição de uma atividade sexual estável, isto é, quando os processos de aprendizagem (incluindo lidar com a novidade) foram concluídos e desempenham apenas um papel menor na execução do sexo atividade, enquanto em contraste continua a ocorrer quando a atividade sexual é repetida. Em outras palavras, a dopamina extracelular no mPFC (e no nucleus accumbens) sempre aumenta durante a cópula, mesmo depois que o comportamento sexual foi aprendido. Mais estudos são necessários para esclarecer o papel funcional desse aumento na atividade da dopamina no mPFC (e no nucleus accumbens; consulte a seção “Introdução e referências”).
Em conclusão, este estudo mostra pela primeira vez que a presença de, e ainda mais, a interação com uma mulher sexualmente receptiva leva a um aumento de dopamina extracelular e NA no dialisato da CPFM de sexualmente ingênua (nunca exposta a estímulos sexuais ) e sexualmente experientes (que foram submetidas a cinco testes preliminares de cópula e apresentam desempenhos sexuais estáveis) RHA e RLA, que apresentam estilos de coping marcadamente diferentes em resposta a condições aversivas e diferentes padrões de comportamento copulatório com uma fêmea sexualmente receptiva também. Consequentemente, as concentrações de dopamina e NA foram encontradas mais elevadas no dialisado de ratos RHA sexualmente ingénuos e experientes, quando comparados com os seus homólogos de RLA durante ambas as fases antecipatória e consumatória da actividade sexual. Estes resultados podem ser devidos em parte a um tônus dopaminérgico e noradrenérgico no mPFC de ratos RHA mais alto que o de ratos RLA, como relatado para a dopamina no nucleus accumbens (Sanna et al., 2015). Além disso, mPFC dopamina extracelular e NA também foram encontrados em ratos com RHA experimentada sexualmente e, em menor grau, RLA comparados com seus pares sexualmente ingênuos em condições basais, bem como durante as fases apetitiva e consumatória do comportamento sexual. Isso sugere que a experiência sexual induz processos plásticos que potencializam ainda mais a dopamina e a neurotransmissão de NA no mPFC, como encontrado para a dopamina no nucleus accumbens de ambas as linhas de ratos romanas (Sanna et al., 2015). As diferenças acima na neurotransmissão dopaminérgica e noradrenérgica no mPFC das duas linhas romanas podem desempenhar um papel não apenas nos diferentes padrões de comportamento sexual, mas também nos diferentes e muitas vezes opostos traços comportamentais que caracterizam essas duas linhas de ratos.
Contribuições do autor
FS, AA, MRM, OG e MGC projetaram o projeto. FS e JB projetaram, executaram e analisaram os dados de comportamento sexual e experimentos de microdiálise. MAP, OG e MGC selecionaram e criaram ratos romanos. FS, AA, MRM, OG e MGC supervisionaram o estudo. FS, AA, MRM, OG e MGC escreveram o manuscrito. Todos os autores discutiram os resultados e comentaram o manuscrito.
Financiamento
Este trabalho foi parcialmente apoiado por doações do Ministério Italiano da Universidade e Pesquisa (MIUR) (Independência Científica de Jovens Pesquisadores, SIR 2014, Código nº. RBSI14IUX7) para FS, da Região Autônoma da Sardenha (ARS) (LR 7 / 2007 , “Promoção da Pesquisa Científica e Inovação Tecnológica na Sardenha”, Fundo Social Europeu, 2007 – 2013, Projeto Código No. CRP-59842) para OG e da Universidade de Cagliari para AA, MRM, FS, OG e MGC.
Declaração de conflito de interesse
Os autores declaram que a pesquisa foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.
Agradecimentos
Dr. Barbara Tuveri, da Universidade de Cagliari, para alojamento e cuidados com animais.
Referências
Afonso, VM, Sison, M., Lovic, V. e Fleming, AS (2007). Lesões do córtex pré-frontal medial no rato feminino afetam o comportamento sexual e materno e sua organização sequencial. Behav. Neurosci. 121, 515 – 526. doi: 10.1037 / 0735-7044.121.3.515
Agmo, A. e Villalpando, A. (1995). Estimulantes do sistema nervoso central facilitam o comportamento sexual em ratos machos com lesões do córtex pré-frontal medial. Cérebro Res. 696, 187–193. doi: 10.1016/0006-8993(95)00853-i
Agmo, A., Villalpando, A., Picker, Z. e Fernández, H. (1995). Lesões do córtex pré-frontal medial e comportamento sexual no rato macho. Cérebro Res. 696, 177–186. doi: 10.1016/0006-8993(95)00852-h
Argiolas, A. e Melis, MR (1995). Neuromodulação da ereção peniana: uma visão geral do papel dos neurotransmissores e neuropeptídeos. Prog. Neurobiol. 47, 235–255. doi: 10.1016/0301-0082(95)00020-8
Argiolas, A. e Melis, MR (2005). Controle central da ereção peniana: papel do núcleo paraventricular do hipotálamo. Prog. Neurobiol. 76, 1 – 21. doi: 10.1016 / j.pneurobio.2005.06.002
Argiolas, A. e Melis, MR (2013). Neuropeptídeos e controle central do comportamento sexual do passado para o presente: uma revisão. Prog. Neurobiol. 108, 80 – 107. doi: 10.1016 / j.pneurobio.2013.06.006
Balfour, ME, Brown, JL, Yu, L. e Coolen, LM (2006). Potenciais contribuições de eferentes do córtex pré-frontal medial para ativação neural após comportamento sexual em ratos machos. Neuroscience 137, 1259 – 1276. doi: 10.1016 / j.neuroscience.2005.11.013
Balfour, ME, Yu, L. e Coolen, LM (2004). Comportamento sexual e sinais ambientais associados ao sexo ativam o sistema mesolímbico em ratos machos. Neuropsychopharmacology 29, 718 – 730. doi: 10.1038 / sj.npp.1300350
Belo, LN, Omrani, A., Adan, RA, Webb, IC e Coolen, LM (2016). A ativação da célula dopaminérgica da área tegmentar ventral durante o comportamento sexual de ratos machos regula a neuroplasticidade e a sensibilização cruzada da d-anfetamina após a abstinência sexual. J. Neurosci. 36, 9949 – 9961. doi: 10.1523 / JNEUROSCI.0937-16.2016
Berridge, K. e Robinson, T. (1998). Qual é o papel da dopamina na recompensa: impacto hedônico, aprendizagem de recompensa ou saliência de incentivo? Cérebro Res. Rev. 28, 309–369. doi: 10.1016/s0165-0173(98)00019-8
Bignami, G. (1965). Seleção para altas taxas e baixas taxas de condicionamento de evitação no rato. Anim Behav. 13, 221–227. doi: 10.1016/0003-3472(65)90038-2
Broadhurst, PL e Bignami, G. (1965). Efeitos correlativos da seleção psicogenética: um estudo das cepas de alta e baixa evasão romana de ratos. Behav. Res. Ther. 3, 273–280. doi: 10.1016/0005-7967(64)90033-6
Carboni, E., Silvagni, A., Vacca, C. e Di Chiara, G. (2006). Efeito cumulativo da norepinefrina e do bloqueio do portador de dopamina no aumento da dopamina extracelular na camada do núcleo accumbens, no núcleo do leito da estria terminal e no córtex pré-frontal. J. Neurochem. 96, 473 – 481. doi: 10.1111 / j.1471-4159.2005.03556.x
Carboni, E., Tanda, GL, Frau, R. e Di Chiara, G. (1990). Bloqueio do portador de noradrenalina aumenta as concentrações de dopamina extracelular no córtex pré-frontal: evidência de que a dopamina é absorvida in vivo por terminais noradrenérgicos. J. Neurochem. 55, 1067–1070. doi: 10.1111/j.1471-4159.1990.tb04599.x
Carlsson, A. (1975). “Controle mediado por receptores do metabolismo da dopamina”, em Receptores pré e pós-sinápticos, Eds E. Usdin e WE Bunney (Nova York, NY: Marcel Deker), 49-65.
Carrasco, J., Márquez, C., Nadal, R., Tobeña, A., Fernández-Teruel, A. e Armario, A. (2008). Caracterização dos componentes centrais e periféricos do eixo hipotálamo-hipófise-adrenal nas linhagens de ratos romanos endogâmicos. Psiconeuroendocrinologia 33, 437 – 445. doi: 10.1016 / j.psyneuen.2008.01.001
Coppens, CM, de Boer, SF, Steimer, T. e Koolhaas, JM (2012). Impulsividade e comportamento agressivo em ratos romanos de alto e baixo risco: diferenças de linha de base e alterações induzidas pelo estresse social do adolescente. Physiol. Behav. 105, 1156 – 1160. doi: 10.1016 / j.physbeh.2011.12.013
Corda, MG, Lecca, D., Piras, G., Di Chiara, G. e Giorgi, O. (1997). Parâmetros bioquímicos da neurotransmissão dopaminérgica e gabaérgica no SNC de ratos romanos de alta evitação e de baixa evitação. Behav. Genet 27, 527 – 536. doi: 10.1023 / A: 1021452814574
Corda, MG, Piras, G., Piludu, MA e Giorgi, O. (2014). Efeitos diferenciais do consumo voluntário de etanol sobre a produção de dopamina na carapaça nucleus accumbens de ratos romanos de alto e de baixo risco: um estudo comportamental e de microdiálise cerebral. Mundo J. Neurosci. 4, 279 – 292. doi: 10.4236 / wjns.2014.43031
Dahlstrom, A. e Fuxe, K. (1964). Evidência da existência de neurônios contendo monoamina no sistema nervoso central. I. Demonstração de monoaminas nos corpos celulares dos neurónios do tronco cerebral. Acta Physiol. Scand. Supl. 62, 1-54.
Damsma, G., Pfaus, J., Wenkstern, D., Phillips, A. e Fibiger, H. (1992). O comportamento sexual aumenta a transmissão de dopamina no núcleo accumbens e estriado de ratos machos: comparação com novidade e locomoção. Behav. Neurosci. 106, 181 – 191. doi: 10.1037 / 0735-7044.106.1.181
D'Angio, M., Serrano, A., Driscoll, P. e Scatton, B. (1988). Estímulos ambientais estressantes aumentam os níveis extracelulares de DOPAC no córtex pré-frontal de ratos hipoemotais (alta evitação romana), mas não hiperemocionais (baixos índices de evitação romana). a in vivo estudo voltamétrico. Cérebro Res. 451, 237–247. doi: 10.1016/0006-8993(88)90768-8
Davis, JF, Loos, M. Di Sebastiano, AR, Brown, JL, Lehman, MN e Coolen, LM (2010). Lesões do córtex pré-frontal medial causam comportamento sexual mal-adaptativo em ratos machos. Biol. Psiquiatria 67, 1199 – 1204. doi: 10.1016 / j.biopsych.2009.12.029
Díaz-Morán, S., Palência, M., Mont-Cardona, C., Cañete, T., Blázquez, G., Martínez-Membrives, E., et ai. (2012) Respostas ao estilo de enfrentamento e hormônio do estresse em ratos geneticamente heterogêneos: comparação com as linhagens de ratos romanos. Behav. Cérebro Res. 228, 203 – 210. doi: 10.1016 / j.bbr.2011.12.002
Driscoll, P. e Bättig, K. (1982). “Perfis comportamentais, emocionais e neuroquímicos de ratos selecionados para diferenças extremas no desempenho ativo de fuga bidirecional”, Genética do Cérebroed. I. Lieblich (Amsterdã: Elsevier), 95-123.
Dunlop, BW e Nemeroff, CB (2007). O papel da dopamina na fisiopatologia da depressão. Arco. Gen. Psiquiatria 64, 327 – 337. doi: 10.1001 / archpsyc.64.3.327
Escorihuela, RM, Fernández-Teruel, A., Gil, L., Aguilar, R., Tobeña, A. e Driscoll, P. (1999). Ratos romanos de alta e baixa evitação: diferenças na ansiedade, na busca de novidades e nos comportamentos de transporte. Physiol. Behav. 67, 19–26. doi: 10.1016/S0031-9384(99)00064-5
Everitt, BJ (1990). Motivação sexual: uma análise neural e comportamental dos mecanismos subjacentes às respostas apetitivas e copulatórias de ratos machos. Neurosci. Biobehav. Rev. 14, 217–232. doi: 10.1016/s0149-7634(05)80222-2
Everitt, BJ e Robbins, TW (2005). Sistemas neurais de reforço para dependência de drogas: das ações aos hábitos à compulsão. Nat. Neurosci. 8, 1481 – 1489. doi: 10.1038 / nn1579
Fadda, F., Argiolas, A., Melis, MR, Tissari, AH, Onali, PL e Gessa, GL (1978). Aumento induzido pelo estresse nos níveis de ácido 3,4-dihidroxifenilacético (DOPAC) no córtex cerebral e n.accumbens: reversão pelo diazepam. Life Sci. 23, 2219–2224. doi: 10.1016/b978-0-08-023768-8.51934-4
Fattore, L., Piras, G., Corda, MG e Giorgi, O. (2009). As linhas de ratos de alta e baixa evasão romanas diferem na aquisição, manutenção, extinção e reintegração da auto-administração intravenosa de cocaína. Neuropsychopharmacology 34, 1091 – 1101. doi: 10.1038 / npp.2008.43
Febo, M. (2011). Disparo de células pré-frontais em ratos machos durante a abordagem de uma fêmea sexualmente receptiva: interações com a cocaína. Sinapse 65, 271 – 277. doi: 10.1002 / syn.20843
Fernández-Guasti, A., Omaña-Zapata, I., Luján, M. e Condés-Lara, M. (1994). Ações da ligadura do nervo ciático no comportamento sexual de ratos machos sexualmente experientes e inexperientes: efeitos da decorticação do pólo frontal. Physiol. Behav. 55, 577–581. doi: 10.1016/0031-9384(94)90119-8
Fernández-Teruel, A., Driscoll, P., Gil, L., Aguilar, R., Tobeña, A. e Escorihuela, RM (2002). Efeitos duradouros do enriquecimento ambiental na busca de novidade, sacarina e consumo de etanol em duas linhagens de ratos (RHA / Verh e RLA / Verh) diferindo no comportamento de busca por incentivo. Pharmacol. Biochem. Behav. 73, 225–231. doi: 10.1016/s0091-3057(02)00784-0
Giménez-Llort, L., Cañete, T., Guitart-Masip, M., Fernández-Teruel, A. e Tobeña, A. (2005). Dois fenótipos distintos induzidos por apomorfina nos ratos de alta e baixa evitação romana. Physiol. Behav. 86, 458 – 466. doi: 10.1016 / j.physbeh.2005.07.021
Giorgi, O., Lecca, D., Piras, G. e Corda, MG (2005). Ativação diferencial da liberação de dopamina no núcleo e no invólucro do nucleus accumbens após injeções agudas ou repetidas de anfetaminas: um estudo comparativo nas linhagens de ratos de alta e baixa evasão romanas. Neuroscience 135, 987 – 998. doi: 10.1016 / j.neuroscience.2005.06.075
Giorgi, O., Lecca, D., Piras, G., Driscoll, P. e Corda, MG (2003a). Dissociação entre liberação de dopamina mesocortical e comportamentos relacionados ao medo em duas linhagens de ratos psicogeneticamente selecionadas que diferem nas estratégias de enfrentamento das condições aversivas. EUR. J. Neurosci. 17, 2716 – 2726. doi: 10.1046 / j.1460-9568.2003.02689.x
Giorgi, O., Piras, G., Lecca, D., Hansson, S., Driscoll, P. e Corda, MG (2003b). Propriedades neuroquímicas diferenciais da transmissão serotoninérgica central em ratos de alto e baixo risco. J. Neurochem. 86, 422 – 431. doi: 10.1046 / j.1471-4159.2003.01845.x
Giorgi, O., Orlandi, M., Escorihuela, RM, Driscoll, P., Lecca, D. e Corda, MG (1994). Transmissão GABAérgica e dopaminérgica no cérebro de ratos romanos de alta evitação e ratos de baixa evitação. Cérebro Res. 638, 133–138. doi: 10.1016/0006-8993(94)90642-4
Giorgi, O., Piras, G. e Corda, MG (2007). As linhas de ratazana romanas de alta e baixa evasão psicogeneticamente selecionadas: um modelo para estudar a vulnerabilidade individual à dependência de drogas. Neurosci. Biobehav. Rev. 31, 148 – 163. doi: 10.1016 / j.neubiorev.2006.07.008
Goto, Y. e Grace, AA (2005). Modulação dopaminérgica do drive límbico e cortical do nucleus accumbens de comportamento dirigido por metas. Nat. Neurosci. 8, 805 – 812. doi: 10.1038 / nn1471
Gresch, PJ, Sved, AF, Zigmond, MJ e Finlay, JM (1995). Influência local da norepinefrina endógena na dopamina extracelular no córtex pré-frontal medial de ratos. J. Neurochem. 65, 111 – 116. doi: 10.1046 / j.1471-4159.1995.65010111.x
Hernández-Gonzáles, M., Guevara, MA, Cervantes, M., Morali, G. e Corsi-Cabrera, M. (1998). Bandas de frequência características do EEG cortico-frontal durante a interação sexual do rato macho como resultado de análise fatorial. J. fisiologia 92, 43–50. doi: 10.1016/s0928-4257(98)80022-3
Hernández-Gonzáles, M., Prieto-Beracoechea, CA, Arteaga-Silva, M. e Guevara, MA (2007). Diferentes funcionalidades do córtex pré-frontal medial e orbital durante uma tarefa sexualmente motivada em ratos. Physiol. Behav. 90, 450 – 458. doi: 10.1016 / j.physbeh.2006.10.006
Horn, AS (1973). Relações estrutura-atividade para a inibição da captação de catecolaminas em sinaptossomas de neurônios noradrenérgicos e dopaminérgicos em homogeneizados cerebrais de ratos. Fr. J. Pharmacol. 47, 332–338. doi: 10.1111/j.1476-5381.1973.tb08331.x
Casca, EM, Du, J., Lorrain, DS e Matuszewich, L. (1995). Dopamina extracelular na região pré-óptica medial: implicações para a motivação sexual e controle hormonal da cópula. J. Neurosci. 15, 7465-7471.
Casco, EM, Lorrain, DS, Du, J., Matuszewich, L., Lumley, LA, Putnam, SK, et al. (1999) Interações hormonais-neurotransmissores no controle do comportamento sexual. Behav. Cérebro Res. 105, 105–116. doi: 10.1016/s0166-4328(99)00086-8
Casco, EM, Weber, MS, Eaton, RC, Dua, R., Markowski, VP, Lumley, L., et al. (1991) Receptores dopaminérgicos na área tegmentar ventral afetam os componentes motores, mas não motivacionais ou reflexivos, da cópula em ratos machos. Cérebro Res. 554, 72–76. doi: 10.1016/0006-8993(91)90173-S
Ikemoto, S. e Panksepp, J. (1999). O papel do nucleus accumbens dopamine no comportamento motivado: uma interpretação unificadora com referência especial à busca de recompensas. Cérebro Res. Rev. 31, 6–41. doi: 10.1016/S0165-0173(99)00023-5
Kakeyama, M., Sone, H., Miyabara, Y. e Tohyama, C. (2003). A exposição perinatal ao 2,3,7,8-tetrachlorodibenzo-p-dioxina altera a expressão dependente da atividade do mRNA do BDNF no neocórtex e no comportamento sexual de ratos machos na vida adulta. Neurotoxicologia 24, 207–217. doi: 10.1016/S0161-813x(02)00214-0
Lecca, D., Piras, G., Driscoll, P., Giorgi, O. e Corda, MG (2004). Uma ativação diferencial da produção de dopamina na casca e no núcleo do nucleus accumbens está associada às respostas motoras a drogas aditivas: um estudo de diálise cerebral em ratos de alta e baixa evasão. Neurofarmacologia 46, 688 – 699. doi: 10.1016 / j.neuropharm.2003.11.011
Manzo, L., Gómez, MJ, Callejas-Aguilera, JE, Donaire, R., Sabariego, M., Fernández-Teruel, A., et ai. (2014a). Relação entre preferência por etanol e busca por sensação / novidade. Physiol. Behav. 133, 53 – 60. doi: 10.1016 / j.physbeh.2014.05.003
Manzo, L., Gómez, MJ, Callejas-Aguilera, JE, Fernández-Teruel, A., Papini, MR e Torres, C. (2014b). Auto-medicação anti-ansiedade induzida por perda de incentivo em ratos. Physiol. Behav. 123, 86 – 92. doi: 10.1016 / j.physbeh.2013.10.002
Masana, M., Bortolozzi, A. e Artigas, F. (2011). Enchimento seletivo da transmissão dopaminérgica mesocortical por drogas noradrenérgicas: oportunidades terapêuticas na esquizofrenia. Int. J. Neuropsychopharmacol. 14, 53 – 68. doi: 10.1017 / s1461145710000908
Meisel, RL e Sachs, BD (1994). "A fisiologia do comportamento sexual masculino", em A fisiologia da reproduçãoVol. 2, 2nd Edn., Eds E. Knobil e J. Neil (Nova York, NY: Raven Press), 3-96.
Melis, MR e Argiolas, A. (1995). Dopamina e comportamento sexual. Neurosci. Biobehav. Rev. 19, 19–38. doi: 10.1016/0149-7634(94)00020-2
Melis, MR e Argiolas, A. (2011). Controle central da ereção peniana: uma revitalização do papel da ocitocina e sua interação com a dopamina e o ácido glutâmico em ratos machos. Neurosci. Biobehav. Rev. 35, 939 – 955. doi: 10.1016 / j.neubiorev.2010.10.014
Melis, MR, Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., et al. (2007) A oxitocina injetada na área tegmentar ventral induz a ereção peniana e aumenta a dopamina extracelular no núcleo accumbens e no núcleo paraventricular do hipotálamo de ratos machos. EUR. J. Neurosci. 26, 1026 – 1035. doi: 10.1111 / j.1460-9568.2007.05721.x
Melis, MR, Succu, S., Cocco, C., Caboni, E., Sanna, F., Boi, A., et al. (2010) A ocitocina induz ereção peniana quando injetada no subículo valvar: papel do óxido nítrico e do ácido glutâmico. Neurofarmacologia 58, 1153 – 1160. doi: 10.1016 / j.neuropharm.2010.02.008
Melis, MR, Succu, S., Mascia, MS, Cortis, L. e Argiolas, A. (2003). A dopamina extracelular aumenta no núcleo paraventricular de ratos machos durante a atividade sexual. EUR. J. Neurosci. 17, 1266 – 1272. doi: 10.1046 / j.1460-9568.2003.02558.x
Melis, MR, Succu, S., Sanna, F., Boi, A. e Argiolas, A. (2009). A oxitocina injetada no subículo ventral ou no núcleo cortical posteromedial da amígdala induz a ereção peniana e aumenta os níveis extracelulares de dopamina no núcleo accumbens de ratos machos. EUR. J. Neurosci. 30, 1349 – 1357. doi: 10.1111 / j.1460-9568.2009.06912.x
Moreno, M., Cardona, D., Gómez, MJ, Sánchez-Santed, F., Tobeña, A., Fernández-Teruel, A., et ai. (2010) Caracterização da impulsividade nas cepas de ratos de alta e baixa evitação: diferenças comportamentais e neuroquímicas. Neuropsychopharmacology 35, 1198 – 1208. doi: 10.1038 / npp.2009.224
Oliveras, I., Río-Álamos, C., Cañete, T., Blázquez, G., Martínez-Membrives, E., Giorgi, O., et ai. (2015) A inibição de Prepulse prediz o desempenho da memória espacial de trabalho nos ratos de alta e baixa evitação romanos endogâmicos e em ratos NIH-HS geneticamente heterogêneos: relevância para o estudo de anomalias pré-atencionais e cognitivas na esquizofrenia. Frente. Behav. Neurosci. 9: 213. doi: 10.3389 / fnbeh.2015.00213
Paxinos, G. e Watson, C. (2004). O cérebro de rato em coordenadas estereotáxica, 4th Edn, Nova York: Academic Press.
Pfaus, JG (2010). Dopamina: ajudando homens a copular por pelo menos 200 milhões de anos: comentário teórico sobre Kleitz-Nelson et al. (2010) Behav. Neurosci. 124, 877 – 880. doi: 10.1037 / a0021823
Pfaus, JG, Damsma, G., Nomikos, GG, Wenkstern, DG, Blaha, CD, Phillips, AG, et ai. (1990) O comportamento sexual aumenta a transmissão central de dopamina no rato macho. Cérebro Res. 530, 345–348. doi: 10.1016/0006-8993(90)91309-5
Pfaus, JG e Everitt, BJ (1995). “A psicofarmacologia do comportamento sexual”, em Psicofarmacologia: a quarta geração de progressoEds FE Bloom e DJ Kupfer (Nova York, NY: Raven Press), 742-758.
Pfaus, JG e Phillips, AG (1991). Papel da dopamina nos aspectos antecipatórios e consumatórios do comportamento sexual em ratos machos. Behav. Neurosci. 105, 727 – 747. doi: 10.1037 / 0735-7044.105.5.727
Piras, G., Giorgi, O. e Corda, MG (2010). Efeitos dos antidepressivos sobre o desempenho no teste de natação forçada de duas linhas de ratos psicogeneticamente selecionados que diferem nas estratégias de enfrentamento das condições aversivas. Psychopharmacology 211, 403–414. doi: 10.1007/s00213-010-1904-x
Piras, G., Piludu, MA, Giorgi, O. e Corda, MG (2014). Efeitos dos tratamentos antidepressivos crônicos em um modelo genético putativo de vulnerabilidade (ratos de baixa evitação romana) e resistência (ratos de alta evitação romanos) à depressão induzida por estresse. Psychopharmacology 231, 43–53. doi: 10.1007/s00213-013-3205-7
Pitchers, KK, Frohmader, KS, Vialou, V., Mouzon, K., Nestler, EJ, Lehman, MN, et al. (2010) ΔFosB no núcleo accumbens é fundamental para reforçar os efeitos da recompensa sexual. Genes Brain Behav. 9, 831–840. doi: 10.1111/j.1601-183X.2010.00621.x
Pitchers, KK, Coppens, CM, Beloate, LN, Fuller, J., Van, S., Frohmader, KS, et al. (2014) A neuroplasticidade endógena induzida por opióides dos neurônios dopaminérgicos na área tegmentar ventral influencia a recompensa natural e opiácea. J. Neurosci. 34, 8825 – 8836. doi: 10.1523 / JNEUROSCI.0133-14.2014
Jarras, KK, Vialou, V., Nestler, EJ, Laviolette, SR, Lehman, MN e Coolen, LM (2013). As recompensas naturais e de drogas atuam nos mecanismos comuns de plasticidade neural com ΔFosB como um mediador chave. J. Neurosci. 33, 3434 – 3442. doi: 10.1523 / JNEUROSCI.4881-12.2013
Pleim, E., Matochik, J., Barfield, R. e Auerbach, S. (1990). Correlação da liberação de dopamina no nucleus accumbens com o comportamento sexual masculino em ratos. Cérebro Res. 524, 160–163. doi: 10.1016/0006-8993(90)90507-8
Robbins, TW e Arnsten, AFT (2009). A neuropsicofarmacologia da função fronto-executiva: modulação monoaminérgica. Ann. Rev. Neurosci. 32, 267 – 287. doi: 10.1146 / annurev.neuro.051508.135535
Sabariego, M., Morón, I., Gómez, MJ, Donaire, R., Tobeña, A., Fernández-Teruel, A., et ai. (2013) Perda de incentivo e expressão gênica do hipocampo em ratos de alto risco (RHA-I) e romanos baixos (RLA-I). Behav. Cérebro Res. 257, 62 – 70. doi: 10.1016 / j.bbr.2013.09.025
Sachs, BD (2000). Abordagens contextuais da fisiologia e classificação da função erétil, disfunção erétil e excitação sexual. Neurosci. Biobehav. Rev. 24, 541–560. doi: 10.1016/S0149-7634(00)00022-1
Sachs, BD, Akasofu, K., Citro, JK, Daniels, SB e Natoli, JH (1994). Estimulação sem contato de fêmeas de cio evoca ereção peniana em ratos. Physiol. Behav. 55, 1073–1079. doi: 10.1016/0031-9384(94)90390-5
Sachs, BD e Barfield, RJ (1976). Análise funcional do comportamento copulatório masculino no rato. Adv. Viga. Behav. 7, 91–154. doi: 10.1016/s0065-3454(08)60166-7
Salamone, JD, Pardo, M., Yohn, SE, López-Cruz, L., San Miguel, N. e Correa, M. (2016). A dopamina mesolímbica e a regulação do comportamento motivado. Curr. Topo. Behav. Neurosci. 27, 231–257. doi: 10.1007/7854_2015_383
Sanna, F., Corda, MG, Melis, MR, Piludu, MA, Giorgi, O. e Argiolas, A. (2014a). Ratos romanos machos de alta e baixa evitação mostram diferentes padrões de comportamento copulatório: comparação com ratos dawley sprague. Physiol. Behav. 127, 27 – 36. doi: 10.1016 / j.physbeh.2014.01.002
Sanna, F., Piludu, MA, Corda, MG, Argiolas, A., Giorgi, O. e Melis, MR (2014b). A dopamina está envolvida nos diferentes padrões de comportamento copulatório de ratos romanos de alta e baixa evitação: estudos com apomorfina e haloperidol. Pharmacol. Biochem. Behav. 124, 211 – 219. doi: 10.1016 / j.pbb.2014.06.012
Sanna, F., Piludu, MA, Corda, MG, Melis, MR, Giorgi, O. e Argiolas, A. (2015). Envolvimento da dopamina nas diferenças de comportamento sexual entre ratos romanos de alto e baixo risco de evitação: um estudo de microdiálise intracerebral. Behav. Cérebro Res. 281, 177 – 186. doi: 10.1016 / j.bbr.2014.12.009
Scatton, B., D'Angio, M., Driscoll, P. e Serrano, A. (1988). A in vivo Estudo voltamétrico da resposta de neurônios dopaminérgicos mesocorticais e mesoacústicos a estímulos ambientais em cepas de ratos com diferentes níveis de emocionalidade. Ann. NY Acad. Sci. 537, 124–137. doi: 10.1111/j.1749-6632.1988.tb42101.x
Steimer, T. e Driscoll, P. (2003). Respostas de estresse divergentes e estilos de enfrentamento em ratos romanos de alto risco (RHA) e baixo (RLA) psicogeneticamente selecionados: aspectos comportamentais, neuroendócrinos e de desenvolvimento. Estresse 6, 87 – 100. doi: 10.1080 / 1025389031000111320
Succu, S., Sanna, F., Argiolas, A. e Melis, MR (2011). A oxitocina injetada no subículo do hipocampo ventral induz a ereção peniana em ratos machos, aumentando a neurotransmissão glutamatérgica na área tegmentar ventral. Neurofarmacologia 61, 181 – 188. doi: 10.1016 / j.neuropharm.2011.03.026
Succu, S., Sanna, F., Cocco, C., Melis, T., Boi, A., Ferri, GL, et ai. (2008) A ocitocina induz ereção peniana quando injetada na área tegmentar ventral de ratos machos: papel do óxido nítrico e GMP cíclico. EUR. J. Neurosci. 28, 813 – 821. doi: 10.1111 / j.1460-9568.2008.06385.x
Succu, S., Sanna, F., Melis, T., Boi, A., Argiolas, A. e Melis, MR (2007). A estimulação de receptores de dopamina no núcleo paraventricular do hipotálamo de ratos machos induz a ereção peniana e aumenta a dopamina extracelular no núcleo accumbens: envolvimento da ocitocina central. Neurofarmacologia 52, 1034 – 1043. doi: 10.1016 / j.neuropharm.2006.10.019
Thierry, AM, Tassin, JP, Blanc, G. e Glowinski, J. (1976). Ativação seletiva do sistema DA mesocortical por estresse. Natureza 263, 242 – 244. doi: 10.1038 / 263242a0
Wenkstern, D., Pfaus, J. e Fibiger, H. (1993). A transmissão de dopamina aumenta no núcleo accumbens de ratos machos durante sua primeira exposição a ratas sexualmente receptivas. Cérebro Res. 618, 41–46. doi: 10.1016/0006-8993(93)90426-N
Westernik, BHC, De Boer, P., De Vries, JB e Long, S. (1998). As drogas antipsicóticas induzem efeitos similares na liberação de dopamina e noradrenalina no córtex pré-frontal medial do cérebro de ratos. EUR. J. Pharmacol. 361, 27–33. doi: 10.1016/s0014-2999(98)00711-0
Willig, F., M'Harzi, M., Bardelay, C., Viet, D. e Delacour, J. (1991). Estirpes romanas como modelo psicogenético para o estudo da memória de trabalho: dados comportamentais e bioquímicos. Pharmacol. Biochem. Behav. 40, 7–16. doi: 10.1016/0091-3057(91)90313-q
Palavras-chave: comportamento sexual, dopamina, noradrenalina, mPFC, microdiálise, ratos RHA e RLA
Citação: Sanna F, Bratzu J, Piludu MA, Corda MG, Melis MR, Giorgi O e Argiolas A (2017) Dopamina, Noradrenalina e Diferenças no Comportamento Sexual entre Ratos Masculinos de Evitação Romana Alta e Baixa: Um Estudo de Microdiálise no Córtex Pré-frontal Medial . Frente. Behav. Neurosci. 11: 108. doi: 10.3389 / fnbeh.2017.00108
Recebido: 22 Dezembro 2016; Aceito: 19 May 2017;
Publicado: 07 June 2017.