Behaviour Front Neurosci. 2015 pode 19; 9: 128. doi: 10.3389 / fnbeh.2015.00128. eCollection 2015.
Andreatta M1, Pauli P1.
Sumário
No condicionamento clássico, um estímulo inicialmente neutro (estímulo condicionado, CS) torna-se associado a um evento biologicamente saliente (estímulo não condicionado, US), que pode ser dor (condicionamento aversivo) ou comida (condicionamento apetitivo). Após algumas associações, o CS é capaz de iniciar respostas defensivas ou consumatórias, respectivamente. Ao contrário do condicionamento aversivo, o condicionamento apetitivo raramente é investigado em humanos, embora sua importância para comportamentos normais e patológicos (por exemplo, obesidade, vício) seja inegável. O presente estudo pretende traduzir as descobertas em animais sobre o condicionamento apetitivo para humanos usando alimentos como um US. Trinta e três participantes foram investigados entre 8 e 10 horas, sem café da manhã, para garantir que sentiam fome. Durante duas fases de aquisição, uma forma geométrica (avCS +) previu um US aversivo (choque elétrico doloroso), outra forma (appCS +) previu um US apetitivo (chocolate ou pretzel salgado de acordo com a preferência dos participantes) e uma terceira forma (CS- ) não previu nenhum dos EUA. Em uma fase de extinção, essas três formas mais uma nova forma (NOVA) foram apresentadas novamente sem entrega nos Estados Unidos. Avaliações de valência e excitação, bem como respostas de sobressalto e condutância da pele (SCR) foram coletadas como índices de aprendizagem. Encontramos condicionamento aversivo e apetitivo bem-sucedido. Por um lado, o avCS + foi classificado como mais negativo e mais estimulante do que o CS– e a potenciação do sobressalto induzida e o SCR aprimorado. Por outro lado, o appCS + foi classificado como mais positivo do que o CS– e induziu atenuação de sobressalto e maior SCR. Em resumo, confirmamos com sucesso achados em animais em humanos (famintos), demonstrando aprendizado apetitivo e aprendizado aversivo normal.
Introdução
Prever ameaças e alimentos é de importância crucial para a sobrevivência de qualquer organismo. No condicionamento clássico (Pavlov, 1927), uma sugestão precede um evento aversivo, como um choque elétrico leve e doloroso (estímulo aversivo não-condicionado, US), ou um evento apetitivo, como um pellet de alimento (US apetitivo), várias vezes. Posteriormente, este estímulo sozinho (agora rotulado CS, CS +) é capaz de extrair respostas defensivas ou consumativas, respectivamente. O primeiro tipo de aprendizagem associativa é chamado de condicionamento aversivo, enquanto o segundo é chamado de condicionamento apetitivo.
Embora a predição de eventos apetitivos seja tão importante para a sobrevivência quanto a predição de eventos aversivos, o condicionamento apetitivo é notavelmente menos investigado em animais (Bouton e Peck, 1989; Koch et al., 1996; McDannald et al., 2011, para uma revisão, veja Martin-Soelch e outros, 2007), bem como em humanos (Klucken et al., 2009, 2013; Austin e Duka, 2010; Delgado et al., 2011; Levy e Glimcher, 2011). Essa falta de pesquisa pode ser devida à complexidade do paradigma apetitivo em relação ao aversivo. Por exemplo, a comida como um reforçador primário deve ser entregue quando o organismo está com fome, a fim de ser gratificante (para uma revisão recente, ver Dickinson e Balleine, 1994; Clark et al., 2012). Na pesquisa humana, essa dificuldade foi superada com o uso do dinheiro (Austin e Duka, 2010; Delgado et al., 2011; Levy e Glimcher, 2011) ou imagens eróticas (Klucken et al., 2009, 2013). No entanto, estudos de neuroimagem apontaram que os reforços primários (ou seja, lanches ou bebidas) e secundários (ou seja, o dinheiro) ativam algumas regiões cerebrais comuns (por exemplo, estriado), mas também implicam padrões distintos de ativação (Delgado et al., 2011; Levy e Glimcher, 2011).
Para nosso conhecimento, apenas alguns estudos de condicionamento humano examinaram os efeitos de reforçadores primitivos como o odor (Gottfried e outros, 2002), agua (Kumar et al., 2008) ou comida (Prévost et al., 2012). No primeiro estudo, Gottfried et al. (2002) rostos neutros associados (CSs) com um odor desagradável, agradável ou neutro. Curiosamente, eles encontraram uma maior ativação no córtex orbitofrontal (OFC) e no estriado ventral em resposta ao CS + apetitivo versus o CS + aversivo. Eles concluíram que o OFC processa o valor do odor e está envolvido na transferência do valor afetivo do sistema olfativo (US) para o visual (CS). A ativação do estriado (ventro) tem sido interpretada como refletindo a CR apetitiva desencadeada pelo CS + apetitivo. No segundo estudo, Kumar et al. (2008) Convidou seus participantes (controles saudáveis e pacientes com depressão maior) para o laboratório no início da manhã e pediu-lhes para se abster de beber durante a noite, a fim de garantir que eles estavam com sede antes do exame. Fotos fractais foram CSs e 0.1 ml de água nos EUA. Curiosamente, entre outras ativações, os participantes saudáveis (mas não os pacientes com depressão) mostraram maior ativação no estriado ventral para o SC + apetitivo, sugerindo que este estímulo foi processado como recompensador. No terceiro estudo, Prévost et al. (2012) também apresentou imagens fractais como CSs e lanches doces ou salgados como US de acordo com a preferência dos participantes. O CS foi apresentado por 6 se, no último segundo, uma imagem de comida foi apresentada adicionalmente. Cada vez que a imagem da comida aparecia, o experimentador colocava um pedaço de comida nas mãos dos participantes, que podiam consumir o lanche imediatamente. Infelizmente, os autores não relataram a ativação do cérebro para o CS + apetitivo durante a fase de condicionamento clássico; no entanto, eles observaram menos desaceleração cardíaca para o CS + recompensado em comparação com o CS não recompensado–, indicando efeitos de condicionamento diferenciais.
A falta de estudos usando a resposta de sobressalto como um índice para condicionamento apetitivo é surpreendente, especialmente considerando seu amplo uso no condicionamento aversivo. Resposta de sobressalto é uma resposta defensiva ancestral e automática em relação a eventos aversivos súbitos, inesperados e fortes (Koch, 1999). Esta resposta defensiva é mediada por uma via neuronal relativamente simples envolvendo os neurônios da raiz coclear, o núcleo pontino caudal da formação reticular (PnC) e os motoneurônios espinhais (Fendt e Fanselow, 1999; Koch, 1999). Estudos em animais revelaram que a potenciação de sobressalto depende das projeções da amígdala para o PnC (Fendt e Fanselow, 1999; Koch, 1999), enquanto a atenuação de sobressalto depende de um nucleus accumbens intacto (NAcc, Koch, 1999). Tal modulação da resposta de sobressalto é uma medida implícita útil para a valência do estímulo de primeiro plano apresentado. Assim, a potenciação indica valência negativa, enquanto a atenuação indica valência positiva, ambos sem ser grandemente influenciados por processos cognitivos (Hamm e Weike, 2005; Andreatta et al., 2010). Até onde sabemos, apenas um estudo em animais investigou o condicionamento apetitivo medindo as respostas de sobressalto como uma medida dependente. Curiosamente, os ratos examinados apresentaram atenuação de sobressalto para um CS + (isto é, luz) associado à entrega de uma solução de sacarose. Notavelmente, tal atenuação foi prejudicada naqueles animais com lesões de NAcc, mas não naqueles com lesões de amígdala, sugerindo que o NAcc desempenha um papel específico em induzir CRs apetitivos e atenuar as respostas de sobressalto (Koch et al., 1996).
O presente estudo é para o nosso conhecimento o primeiro a traduzir este paradigma de condicionamento apetitivo para os seres humanos usando reforçadores primários como EUA, ou seja, doce (chocolate Smarties®) ou comida salgada (pretzel salgado) e modulação de sobressalto como medida de CRs. Esperávamos que o appCS + acionasse CRs apetitivos fortes, como refletido na atenuação de sobressalto, SCR aprimorado e classificação de valência positiva, em comparação com os outros estímulos, ou seja, o avCS + e o CS–.
Materiais e Métodos
Participantes
Quarenta e dois voluntários aceitaram participar do estudo e receberam os créditos do curso. Nove participantes foram excluídos da análise, dois devido a problemas técnicos, três porque foram codificados como não respondedores (amplitude média de sobressalto <5 μV) e quatro porque não exibiram respostas de sobressalto suficientes por condição (mínimo = 2; para detalhes, consulte Materiais e Método). Ao final, 33 participantes foram considerados na análise (16 homens; idade média: 22.09 anos, SD: 2.84; intervalo: 18 – 29 anos). Quatro participantes não eram alemães nativos e seis eram canhotos. Um participante permaneceu inconsciente das associações CSs-US durante o experimento (consulte Procedimento), mas decidimos não excluir esse participante porque suas respostas eram normais e não afetavam os resultados.
Materiais
Estímulos Incondicionais (EUA)
Dois tipos de EUA foram usados. Como EUA aversivos, aplicamos um choque elétrico levemente doloroso no antebraço não dominante dos participantes. O choque elétrico foi realizado por meio de dois eletrodos com diâmetro 9 mm e espaçamento 30 cm. O estímulo elétrico consistiu de um estímulo de pulso com frequência de 50 Hz e duração de 200 ms, gerado por um estimulador de corrente (Digitimer DS7A, Digitimer Ltd, Welwyn Garden City, Reino Unido, 400 V, máximo de 9.99 mA). A intensidade do choque elétrico foi determinada individualmente através de um procedimento limiar descrito anteriormente (Andreatta et al., 2010). Resumidamente, os participantes foram submetidos a duas séries de intensidades ascendente e descendente em intervalos de 0.5 mA. Eles tiveram que avaliar cada estímulo em uma escala visual variando de 0 (sentindo absolutamente nada) a 10 (dor realmente intensa) com 4 como uma âncora para o limiar (apenas dor perceptível). A intensidade média do estímulo elétrico foi 2.12 mA (SD = 0.56) e foi classificado como doloroso (M = 6.45, SD = 1.73). Os apetitosos EUA consistiam em chocolate (Smarties®) ou um pequeno pretzel salgado. A escolha do US apetitivo dependeu da preferência individual do participante relatada durante a entrevista preliminar. Ou seja, os participantes tiveram que relatar se normalmente comem alimentos salgados ou doces durante o café da manhã. Eles também podiam escolher livremente se preferiam o chocolate ou o salgado pretzel durante o próprio experimento. No final, 22 participantes escolheram o chocolate e 11 o pretzel salgado.
Estímulos Condicionados (CS)
Formas geométricas (quadrado azul, círculo amarelo, triângulo verde, hexágono vermelho) com uma diagonal de 8 cm foram apresentadas como CSs. As formas foram apresentadas no meio de uma tela de computador preta para 8 s. Uma forma (avCS +) sempre esteve associada à US aversiva (choque elétrico doloroso), uma forma (appCS +) sempre esteve associada à US apetitiva (pretzel de chocolate ou salgado), uma forma (CS–) nunca esteve associada à aversão ou os USs apetitosos, e a quarta forma (NEW) foi apresentada na fase de extinção, mas não durante a fase de aquisição, a fim de assegurar sua neutralidade.
Sonda de Surto
O ruído branco de 103 dB com duração de 50 ms foi usado como uma prova de alarme. Os estímulos acústicos foram apresentados binauralmente em fones de ouvido e ocorreram aleatoriamente 4 a 6 s após o início da forma.
Questionários
Antes e depois do experimento, os participantes tiveram que preencher as versões alemãs do Inventário de Ansiedade Traço-Estado (STAI, Laux et al., 1981) e o Cronograma Positivo Negativo Afectado (PANAS, Krohne e outros, 1996) O STAI é um inventário para avaliar o traço e / ou ansiedade do estado dos participantes e consiste em 20 itens para as versões do traço e do estado. Nível de ansiedade dos participantes antes de (M = 37.06, SD = 7.80) e depois (M = 39.33, SD = 9.16) o experimento não mudou significativamente [t(32) = 1.61, p = 0.117]. Os escores de ansiedade do traço na amostra atual variaram entre 20 e 58 (M = 36.6, SD = 8.98), que é comparável ao intervalo normal publicado de adultos (Laux et al., 1981). O PANAS (Krohne e outros, 1996) é um índice de humor positivo e negativo. Indivíduos com pontuações altas na escala de afetos positivos (PAS) são propensos a emoções como entusiasmo, enquanto indivíduos com pontuações altas na escala de afetos negativos (NAS) são propensos a emoções como angústia. Cada item é composto por um adjetivo, e os participantes indicam em uma escala que varia de 1 (muito levemente) a 5 (extremamente) em que medida o adjetivo reflete seus sentimentos naquele momento particular. Nenhuma diferença significativa foi encontrada no efeito negativo dos participantes como resultado do experimento [começar: M = 11.67, SD = 2.29; fim: M = 12.88, SD = 4.69; t(32) = 1.55, p = 0.130]. De alguma forma, os participantes mudaram significativamente seu humor positivo desde o início (M = 26.72, SD = 4.70) até o final (M = 23.66, SD = 7.02) da experiência [t(31) = 3.11, p = 0.004]. Essa diminuição do humor positivo dos participantes pode estar relacionada ao desagrado do paradigma (foram apresentados choques elétricos dolorosos e também ruído branco aversivo).
Procedimento
Ao chegar ao laboratório, os participantes leram e assinaram um termo de consentimento informado, aprovado pelo comitê de ética do Departamento de Psicologia da Universidade de Würzburg. Eles não foram informados sobre a contingência entre CSs e USs. Depois de preencher os questionários, os eletrodos foram fixados e o procedimento do limiar de dor foi realizado como descrito acima.
Durante a fase de habituação, as quatro formas geométricas foram apresentadas duas vezes com um intervalo entre ensaios (ITI) variando entre 18 e 25 s (média: 21.5 s). Nenhum US ou startle probes foram entregues durante esta fase.
Antes da fase de aquisição, sete surtos de ruído branco eram entregues a cada 7-15 s para diminuir a reatividade inicial do sobressalto.
Os dois seguintes fases de aquisição eram idênticos (Figura 1). Cada fase de aquisição consistia em testes 24: testes 8 CS, testes 8 avCS + e testes 8 appCS +. A sequência CS foi pseudo-aleatória, com a única restrição sendo que o mesmo estímulo não poderia ser apresentado mais do que duas vezes seguidas. Notavelmente, o avCS + foi apresentado juntamente com um relâmpago como símbolo do choque elétrico e o doloroso US foi entregue no deslocamento. O appCS + foi apresentado em combinação com uma imagem de Smarties ou um pretzel salgado, e o participante poderia escolher um Smarties ou um pretzel de uma jarra. O CS - foi apresentado juntamente com um símbolo de proibição e nenhum dos EUA foi entregue. Durante três das apresentações 8 CS de cada tipo, uma sonda de sobressalto foi entregue entre 4 e 6 após o início do estímulo. Três sondas startle adicionais foram apresentadas durante os ITIs para assegurar sua imprevisibilidade e reduzir a habituação do sobressalto. O ITI, consistindo de uma tela preta, variava entre 18 e 25 s com uma média de 21.5 s.
FIGURA 1
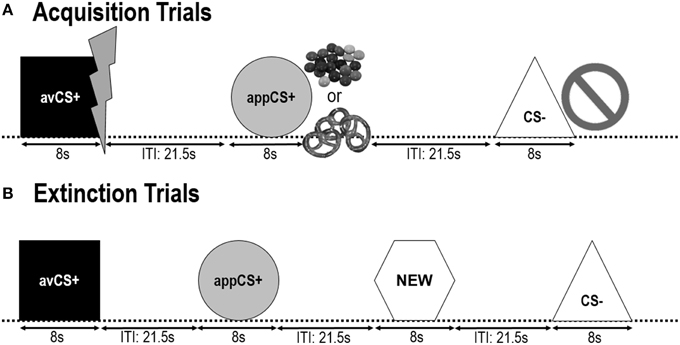
Figura 1. Ensaios durante as duas fases de aquisição (A) e a fase de extinção (B). Os participantes aprenderam que uma forma (avCS +) previa um choque elétrico leve e doloroso, uma forma (appCS +) previa um pedaço de chocolate ou um pretzel salgado pequeno (de acordo com sua preferência) e uma terceira forma (CS–) não predizia nenhum evento biologicamente saliente. Cada forma foi apresentada em conjunto com uma imagem representando um choque elétrico, smarties / pretzel salgado, ou nada dependendo da associação com os EUA. Durante a fase de extinção, as três formas geométricas foram apresentadas mais uma vez, mas nenhum US foi entregue. Além disso, uma quarta forma geométrica (NEW) foi apresentada como um controle neutro.
Durante a fase de extinção, os participantes viram as três formas geométricas (ou seja, o avCS +, o appCS + e o CS–) novamente, além de uma nova forma neutra (NEW). Nenhum dos EUA foi entregue e as formas não foram apresentadas em conjunto com imagens de flashes, pretzels de chocolate / salgado ou proibições. Cada estímulo foi apresentado oito vezes em uma ordem pseudo-aleatória (ou seja, o mesmo estímulo não foi apresentado mais do que duas vezes seguidas), criando testes 32. Estímulos de sonda de sobressalto foram apresentados durante 4 de apresentações de estímulo 8 de cada tipo CS. Como na fase de aquisição e de habituação, os ITIs variaram entre 18 e 25 s, e as sondas de sobressalto adicionais 4 foram entregues imprevisivelmente durante os ITIs.
Após cada fase, os participantes avaliaram a valência (agradabilidade) e a excitação (excitatória) dos CSs com escalas visuais analógicas (VAS) variando de 1 a 9. A escala de valência variou de “1” indicando “muito desagradável” a “9” indicando “muito agradável”; a escala de excitação variou de “1” indicando “calmo” a “9” indicando “emocionante”. Além disso, os valores de contingência foram avaliada após as duas fases de aquisição e a fase de extinção. Os participantes viram uma forma geométrica para os 1s e depois precisaram indicar se essa forma estava associada ao choque elétrico, com o pretzel de chocolate / salgado, sem nada, ou se não eram capazes de fazer qualquer associação. Notavelmente, todos os participantes (exceto um) estavam cientes da contingência após a Aquisição 2.
Redução de dados
As respostas fisiológicas foram gravadas com um amplificador 16 V-Amp e um software Vision Recorder V-Amp Edition (Versão 1.03.0004, BrainProducts Inc., Munique, Alemanha). Uma taxa de amostragem de 1000 Hz e um filtro de entalhe 50 Hz foram aplicados. As análises offline foram realizadas com o Brain Vision Analyzer (Versão 2.0; BrainProducts Inc., Munique, Alemanha).
Resposta de Surto
A resposta do sobressalto foi medida por eletromiografia (EMG) à esquerda orbicularis oculi músculo com dois eletrodos 5 mm Ag / AgCl. De acordo com as diretrizes (Blumenthal et al., 2005), um eletrodo foi posicionado sob a pupila e o segundo 1 cm lateralmente. Os eletrodos terra e referência foram colocados nas mastoides direita e esquerda, respectivamente. Antes de ligar os eletrodos, a pele foi levemente desgastada e limpa com álcool para manter a impedância abaixo de 10 kΩ. O sinal eletromiográfico foi filtrado off-line com um filtro de corte baixo 28 Hz e um filtro de corte alto 500 Hz. Então, o sinal EMG foi retificado e uma média móvel de 50 ms foi aplicada. Usamos o 50 ms antes do início do probe startle como linha de base (Grillon e outros, 2006). As respostas às sondas de sobressalto foram classificadas manualmente, e os ensaios com excessivos desvios da linha de base (± 5 μV) ou artefatos de movimento foram excluídos da análise posterior. Respostas de sobressalto inferiores a 5 μV foram codificadas como zero e consideradas para o cálculo da magnitude do sobressalto (Blumenthal et al., 2005). No total, 10.4% dos ensaios foram rejeitados, e um mínimo de 2 de respostas de sobressalto 3 nas fases de aquisição e 4 de respostas de sobressalto 8 na fase de extinção para cada condição foram necessários para manter o participante no pool de análise. Por esse motivo, quatro participantes foram excluídos. A amplitude do pico foi definida como o pico máximo em relação à linha de base durante a janela de tempo 20-120 ms após o início da sonda startle. Os dados brutos foram então normalizados dentro dos sujeitos usando z-escores e, em seguida, T-scores, a fim de reduzir a influência da variabilidade individual e para melhor detectar processos psicológicos. Os escores T foram calculados para cada condição (avCS +, appCS +, CS–, NEW e ITI). Para investigar a potenciação de sobressalto ou atenuação de sobressalto, as pontuações para as respostas de sobressalto ITI foram subtraídas das respostas de sobressalto para cada condição.
Resposta de Condutância da Pele (SCR)
A resposta de condutância da pele (SCR) foi registrada usando dois eletrodos 5 mm Ag / AgCl colocados na palma da mão não dominante. A resposta galvânica foi filtrada offline com um filtro de corte alto 1 Hz. O SCR foi definido como a diferença (em μS) entre o início da resposta (1-3 s após o início do estímulo) e o pico de resposta (Tranel e Damasio, 1994; Delgado et al., 2011). Ensaios contendo sondas de sobressalto não foram considerados na análise do SCR. As respostas abaixo de 0.02 μS foram codificadas como zero. Cinco outros participantes foram excluídos da análise SCR porque tinham um SCR médio inferior a 0.02 μS. Os dados de condutância da pele crua foram transformados em raiz quadrada para normalizar a distribuição e as pontuações foram calculadas para cada condição separadamente para as duas fases de aquisição (avCS +, appCS +, CS–) e a fase de extinção (avCS +, appCS +, CS– e NEW ).
Análise Estatística
Todos os dados foram analisados com o SPSS for Windows (versão 20.0, SPSS Inc.). Para as respostas fisiológicas, análises de variância (ANOVA) multivariadas separadas foram calculadas para as duas fases de aquisição e a fase de extinção. A ANOVA para as fases de aquisição teve estímulo (avCS +, appCS +, CS–) e fase (Aquisição 1, Aquisição 2) como fatores dentro dos sujeitos. A ANOVA para a fase de extinção tinha apenas um estímulo (avCS +, appCS +, CS–, NEW) como um fator dentro dos sujeitos. As classificações de valência, excitação e contingência foram analisadas com ANOVAs separados, contendo os estímulos dos fatores dentro dos sujeitos (avCS +, appCS +, CS– e NEW) e fase. Este fator teve quatro níveis para as classificações de valência e excitação (T1: após a fase de habituação, T2: após a primeira fase de aquisição, T3: após a segunda fase de aquisição, T4: após a fase de extinção), mas três níveis para as classificações de contingência (T1: primeira fase de aquisição, T2: após a segunda fase de aquisição, T3: após a fase de extinção).
O nível alfa (α) foi estabelecido em 0.05 para todas as análises. O tamanho do efeito é reportado como parcial η2.
Resultados
As classificações de valência e ativação para cada fase são mostradas na Figura 2; as respostas de susto e os SCRs são representados na Figura 3.
FIGURA 2
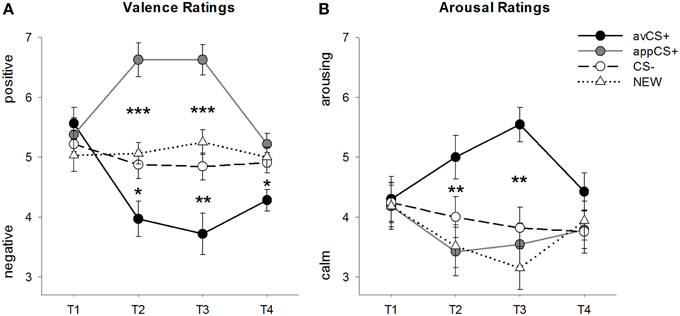
Figura 2. Classificações para valência (A) e excitação (B). As linhas (com erros padrão) representam as classificações após a fase de habituação (T1), Aquisição 1 (T2), Aquisição 2 (T3) e a fase de extinção (T4). O CS + aversivo (linha sólida preta) adquiriu valência negativa e excitação alta após as duas fases de aquisição em comparação com o CS- (linha tracejada preta) e o NOVO (linha pontilhada preta). É importante ressaltar que o apetitivo CS + (linha sólida cinza) adquiriu valência positiva em relação ao CS– e ao NOVO. *p <0.05 **p > 0.01, ***p <0.001.
FIGURA 3
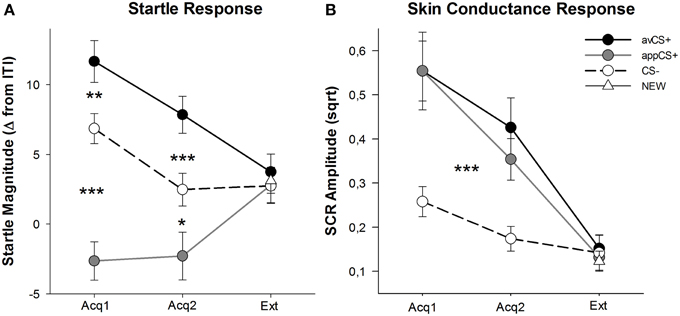
Figura 3. Respostas de sobressalto (A) e condutância da pele (B) (com erros padrão) durante a primeira fase de aquisição (Acq1), a segunda fase de aquisição (Acq2) e a fase de extinção (Ext). As respostas de sobressalto foram significativamente potenciadas para o CS + aversivo (linha sólida preta) e significativamente atenuadas para o CS + apetitivo (linha sólida cinza) em comparação com o CS– (linha tracejada preta) durante as fases de aquisição. O SCR foi significativamente maior para o avCS + e o appCS + em comparação com o CS–. Nenhuma diferença foi revelada durante a fase de extinção. *p <0.05 **p > 0.01, ***p <0.001.
classificações
A ANOVA para o classificações de valência durante a aquisição revelou efeitos principais significativos do estímulo [F(3, 93) = 17.26, GG-ε = 0.801, p <0.001, η parcial2 = 0.358] e fase [F(3, 93) = 3.30, GG-ε = 0.731, p = 0.039, parcial η2 = 0.096], bem como uma interação significativa entre estímulo e fase [F(9, 279) = 9.54, GG-ε = 0.463, p <0.001, η parcial2 = 0.235]. Acompanhamento t-testes indicam que as valências das quatro formas geométricas no início da experiência eram idênticas (ps> 0.19), enquanto após a Aquisição 1 e a Aquisição 2 o avCS + foi classificado como especialmente negativo e o appCS + como especialmente positivo. Especificamente, o avCS + foi classificado como mais negativamente valenciado em comparação com o CS– [Acq1: t(31) = 2.34, p = 0.026; Acq2: t(31) = 3.07, p = 0.004], o NOVO [Acq1: t(31) = 2.70, p = 0.011; Acq2: t(31) = 3.89, p <0.001], e o appCS + [Acq1: t(31) = 5.41, p <0.001; Acq2: t(31) = 6.11, p <0.001]. O appCS + foi classificado como significativamente mais positivo do que o CS– [Acq1: t(31) = 4.99, p <0.001; Acq2: t(31) = 5.31, p <0.001] e o NOVO [Acq1: t(31) = 4.92, p <0.001; Acq2: t(31) = 4.14, p <0.001]. As diferenças entre o CS– e o NOVO nunca foram significativas (ps> 0.18).
É importante ressaltar que não foram encontradas diferenças significativas ao comparar as classificações de valência para o appCS + associado ao chocolate versus o pretzel salgado [Acq1: t(30) = 0.03, p = 0.477; Acq2: t(30) = 0.29, p = 0.775].
Após a fase de extinção, o avCS + ainda foi classificado como mais negativo em comparação ao CS- [t(31) = 2.40, p = 0.023], o NOVO [t(31) = 3.40, p = 0.002] e o appCS + [t(31) = 3.35, p = 0.002]. Pelo contrário, a valência do appCS + não diferiu mais do CS- [t(31) = 1.77, p = 0.086] ou o NOVO [t(31) = 1.07, p = 0.293].
A ANOVA para o classificações de excitação durante a aquisição revelou um efeito principal significativo do estímulo [F(3, 96) = 7.07, GG-ε = 0.737, p = 0.001, parcial η2 = 0.181], mas não de fase [F(3, 96) = 1.27, GG-ε = 0.805, p = 0.289, parcial η2 = 0.038], e uma interação significativa entre estímulo e fase [F(9, 288) = 4.53, GG-ε = 0.582, p = 0.001, parcial η2 = 0.124]. Acompanhamento t-testes não indicaram diferenças significativas entre os estímulos em relação ao despertar inicial (ps> 0.74). No entanto, após as duas fases de aquisição, o avCS + foi classificado como mais excitante do que o NOVO [Acq1: t(32) = 2.99, p = 0.005; Acq2: t(32) = 5.97, p <0.001] e o appCS + [Acq1: t(32) = 2.62, p = 0.013; Acq2: t(32) = 4.42, p <0.001], e marginalmente mais excitante do que o CS - após a primeira fase de aquisição [t(32) = 1.96, p = 0.058], mas significativamente mais excitante após a segunda fase de aquisiçãot(32) = 3.65, p = 0.001]. Em contraste com as classificações de valência, o despertar do appCS + não diferiu do despertar do CS– e do NEW (ps > 0.13) após as fases de aquisição.
Da mesma forma que com as classificações de valência, CS– e NEW não diferiram em sua excitação (ps> 0.07).
Como foi o caso da valência, não foram reveladas diferenças para o appCS + associado ao chocolate e o appCS + associado ao pretzel salgado [Acq1: t(31) = 0.26, p = 0.797; Acq2: t(31) = 0.33, p = 0.724].
Após a fase de extinção, não foram reveladas diferenças significativas nas classificações de excitação (ps> 0.08).
Para adicional post-hoc t-testes comparando os ratings após a fase de habituação, a primeira e a segunda fases de aquisição e a fase de extinção, consulte Materiais Suplementares.
Resposta de Surto
A ANOVA para as fases de aquisição retornou um efeito principal de estímulo [F(2, 64) = 49.92, GG-ε = 0.964, p <0.001, η parcial2 = 0.609], mas não fase [F(1, 32) = 3.16, p = 0.085, parcial η2 = 0.090] e uma interação significativa Estímulo × Fase [F(2, 64) = 3.37, GG-ε = 0.875, p = 0.048, parcial η2 = 0.095]. Acompanhamento t-testes revelaram potenciação de sobressalto significativa para o avCS + em comparação com o CS - durante ambos os primeiros [t(32) = 3.27, p = 0.003] e o segundo [t(32) = 4.00, p <0.001] fases de aquisição. Respostas de sobressalto ao avCS + também foram significativamente potencializadas em comparação com as do appCS +, novamente após a Aquisição 1 [t(32) = 8.20, p <0.001] e Aquisição 2 [t(32) = 5.74, p <0.001]. É importante ressaltar que a magnitude do sobressalto para o appCS + foi significativamente atenuada em comparação com o CS– durante a Aquisição 1 [t(32) = 6.34, p <0.001] e Aquisição 2 [t(32) = 2.91, p = 0.007]. Novamente e de acordo com as classificações, nenhuma resposta diferencial de sobressalto foi revelada para o appCS + quando associado ao chocolate ou ao pretzel salgado [Acq1: t(31) = 1.04, p = 0.309; Acq2: t(31) = 0.07, p = 0.947]. Durante a fase de extinção, nenhum efeito significativo foi encontrado [F(3, 96) = 0.26, GG-ε = 0.906, p = 0.833, parcial η2 = 0.008].
Resposta de Condutância da Pele (SCR)
A partir das ANOVAs para o SCR durante as duas fases de aquisição, os principais estímulos de efeitos [F(2, 54) = 18.04, GG-ε = 0.908, p <0.001, η parcial2 = 0.401] e fase [F(1, 27) = 20.91, p <0.001, η parcial2 = 0.436] acabou por ser significativo, mas não a sua interação [F(2, 54) = 0.68, GG-ε = 0.637, p = 0.451, parcial η2 = 0.024]. Post-hoc t-testes indicaram SCR significativamente maior para o avCS + [t(27) = 6.46, p <0.001] e para o appCS + [t(27) = 4.84, p <0.001] em comparação com o CS–, enquanto os participantes mostraram SCR comparável ao avCS + e ao appCS + [t(27) = 0.64, p = 0.527]. Notavelmente, nenhuma diferença em SCRs para o appCS + de chocolate e para o appCS + salgado foi encontrada [Acq1: t(26) = 2.55, p = 0.120; Acq2: t(26) = 1.29, p = 0.210]. Como foi verdade para as classificações e a resposta de sobressalto, não foram encontrados efeitos significativos para a fase de extinção [F(3, 81) = 0.28, GG-ε = 0.634, p = 0.743, parcial η2 = 0.010].
Discussão
O objetivo deste estudo foi traduzir as descobertas de animais para humanos usando um paradigma de condicionamento apetitivo clássico com um reforçador primário como estímulo incondicionado (US). Para isso, os participantes vinham ao laboratório de manhã cedo, sem café da manhã, para garantir que estivessem com fome e, de acordo com sua preferência, pedaços de chocolate ou pretzels salgados eram usados como USs apetitosos. Durante a fase de aquisição, uma forma geométrica (avCS +) foi associada a um choque elétrico levemente doloroso (aversive US), outra forma (appCS +) com o US apetitivo e uma terceira forma (CS–) sem o US apetitivo nem com o EUA aversivos. Os resultados indicam um condicionamento aversivo e apetitivo bem-sucedido no nível verbal explícito (ou seja, classificações), no nível comportamental implícito (isto é, resposta de sobressalto) e no nível fisiológico (isto é, SCR). Especificamente, o avCS + em comparação com o CS– gerou mais classificações negativas de valência, maiores classificações de despertar, potencialização de sobressalto e maior SCR. Mais importante ainda, o appCS + comparado ao CS - acionou classificações de valência mais positivas, atenuação de sobressalto e maior SCR. Nossas descobertas sobre o condicionamento aversivo estavam de acordo com as expectativas, uma vez que estudos anteriores descobriram que um estímulo que prevê ameaça (avCS +) é classificado como aversivo, provoca maiores respostas de medo e aumenta a excitação fisiológica (Fendt e Fanselow, 1999; Hamm e Weike, 2005; Andreatta et al., 2010, 2013). Nossos resultados sobre condicionamento apetitivo também estavam de acordo com estudos anteriores em humanos e animais mostrando que um estímulo preditor de recompensa (appCS +) é classificado como positivo, inibe respostas de medo e aumenta a excitação fisiológica (Koch et al., 1996; Gottfried e outros, 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Austin e Duka, 2010; Prévost et al., 2012). Para nosso conhecimento, este é o primeiro estudo demonstrando atenuação de sobressalto condicionada em humanos para um estímulo que prevê uma recompensa primária. É importante ressaltar que conseguimos transferir e confirmar os resultados de um estudo em animais (Koch et al., 1996). Este estudo em animais demonstrou que a atenuação de sobressalto em ratos depende de projeções do NAcc (parte do estriado ventral) para o PnC. Portanto, a atenuação do sobressalto em nosso estudo poderia implicar NAcc atividade, que também está em consonância com os achados fMRI (Gottfried e outros, 2002; Kumar et al., 2008; Klucken et al., 2009, 2013; Delgado et al., 2011; Levy e Glimcher, 2011). Assim, concluímos que nosso paradigma de condicionamento apetitivo foi bem-sucedido, como indicaram as valências positivas explícitas (classificações) e implícitas (atenuação de sobressalto).
Além desta nova descoberta, mas previsível, dois outros resultados interessantes merecem ser mencionados. Primeiro, as respostas de excitação verbal e fisiológica ao appCS + dissociaram-se. Em segundo lugar, encontramos uma extinção mais rápida dos CRs apetitivos em comparação com os CRs aversivos.
O SCR indica ativação simpática, que é aumentada tanto para o estímulo associado à recompensa (appCS +) quanto para o estímulo associado à ameaça (avCS +). Este resultado está de acordo com um estudo de condicionamento anterior, em que imagens eróticas foram usadas como US apetitivo (Klucken et al., 2013), e um estudo revelando SCR comparável para uma sugestão de previsão de dinheiro e para uma sugestão prevendo um ruído aversivo (Austin e Duka, 2010). Notavelmente, o SCR é uma resposta orientadora relacionada à ativação do sistema simpático. Esta resposta foi sugerida para refletir a preparação de uma reação comportamental a eventos motivacionalmente salientes (Bradley, 2009). Com base nisso, acreditamos que tanto os estímulos associados à ameaça quanto os associados à recompensa provocaram uma resposta preparatória para respostas comportamentais sucessivas. Em outras palavras, os sinais de comida e dor iniciaram a preparação para o comportamento de aproximação e evitação, respectivamente. Em contraste com a alta estimulação fisiológica, as respostas verbais indicaram baixa ativação do estímulo associado à recompensa. É possível que a excitação verbal seja mais influenciada pela natureza excitante dos EUA do que pela ativação fisiológica. per se. De fato, achamos que o appCS + vs. o avCS + foi classificado como humilde vs. altamente excitante porque estava ligado a Estados Unidos modestos e altamente instigantes, respectivamente. Infelizmente, não coletamos classificações de alerta para os EUA e, portanto, dependeremos de estudos futuros para testar essa hipótese explicitamente.
Durante a fase de extinção, nenhum dos EUA foi entregue. Isto pode ter induzido uma nova aprendizagem inibitória chamada extinção (para uma revisão ver Milad e Quirk, 2012), com os CRs aversivo e apetitivo diminuindo como conseqüência. Evidentemente, nos níveis comportamental (isto é, resposta ao susto) e fisiológico (isto é, SCR), não foram detectadas reações diferenciais ao avCS +, ao appCS + e ao CS–. Da mesma forma, a estimulação explícita dos estímulos condicionados avaliados após a fase de extinção equalizou em um nível baixo, sugerindo o sucesso da aprendizagem da extinção. No entanto, o avCS + ainda foi avaliado significativamente mais negativo que o CS–, enquanto o appCS + valência não diferiu mais do CS-valence. A extinção mais lenta da resposta explícita e aversiva pode ser devida a um conservadorismo evolucionário, o que significa que os sinais de ameaça são especialmente difíceis de esquecer, porque uma não resposta a um sinal de ameaça pode ser fatal.
Uma questão ainda permanece: por que então a resposta de sobressalto (isto é, a valência implícita) se extingue completamente durante a fase de extinção? Primeiro, essas respostas foram calculadas durante todo o curso da fase. Portanto, é concebível que RCs discriminativos ainda sejam detectáveis durante os primeiros testes da fase de extinção. De uma maneira exploratória, seguimos essa hipótese e examinamos as respostas de sobressalto e a SCR ao longo da fase de extinção (ver Material Suplementar). Embora não tenhamos encontrado diferenças significativas, observamos uma magnitude de sobressalto ligeiramente maior para o avCS + quando comparado ao CS– e ao novo estímulo de controle. Observamos também um pouco mais de atenuação de sobressaltos para o appCS + em comparação com o CS - e com o novo estímulo de controle no início da fase de extinção, que, no entanto, desapareceu em algumas tentativas. SCR para o appCS + caiu depois do 2nd extinction trial, enquanto o SCR para o avCS + permaneceu mais alto para quase todas as extinções em comparação com o SCR para o NEW. Embora devêssemos interpretar esses resultados com grande cautela, as respostas de sobressalto pareciam paralelas às classificações de valência. Além disso, tanto as respostas de sobressalto como os SCRs apoiam ainda mais a ideia de um conservadorismo evolucionário em relação aos estímulos de ameaça.
Finalmente, temos que reconhecer algumas limitações deste estudo. Primeiro, devido a um problema técnico, não pudemos reportar classificações de agradabilidade (e excitação) para o chocolate e o pretzel salgado. No entanto, os CRs apetitivos sugerem que os participantes realmente sentiram os dois EUA como apetitivos. Em segundo lugar, a duração dos EUA aversivos e apetitosos diferiu bastante. Assim, o doloroso choque elétrico foi entregue no offset do avCS + e durou exatamente 200 ms, enquanto o chocolate e o pretzel salgado foram apresentados aos participantes por volta do 2 após o aparecimento do appCS +, e a duração era indefinível porque dependia da rapidez com que o indivíduo os comeu. A escolha de fornecer os USs apetitivos dessa maneira foi baseada em um estudo anterior em humanos (Prévost et al., 2012). No entanto, seria metodologicamente mais elegante entregar um US apetitivo mais comparável aos EUA aversivos, como um gole de suco ou ginger ale no appCS + offset. Para compensar essa grande diferença perceptiva, apresentamos formas geométricas em conjunto com um estímulo visual simbolizando os EUA. Terceiro, não podemos definitivamente excluir a possibilidade de que as respostas fisiológicas rapidamente extintas na fase de extinção se devam a um aspecto metodológico. De fato, os estímulos visuais nessa fase não foram apresentados em conjunto com o símbolo dos EUA, como foi o caso nas fases de aquisição. É possível que a apresentação dos CSs possa ter influenciado os CRs e, por essa razão, não houve diferenças significativas durante os primeiros ensaios de extinção. No entanto, nossas respostas rápidas e extinguidas estão de acordo com as respostas extintas em um estudo anterior em que o CS + (face) foi apresentado em conjunto com os EUA (grito) durante a aquisição, mas não durante a extinção (Lissek et al., 2008).
Em conclusão, encontramos respostas condicionadas aversivas e apetitivas bem-sucedidas a um estímulo associado à ameaça e a um estímulo associado à recompensa, respectivamente. Curiosamente, o nível de respostas explícito (classificações) e implícito (reflexo de sobressalto) funcionou de maneira sinérgica, em que o avCS + foi relatado como potenciação de sobressalto negativa e induzida e o appCS + foi relatado como atenuação de sobressalto positiva e induzida. Além disso, a excitação explícita (classificação) e fisiológica (SCR) do appCS + dissociou-se, refletindo dois processos distintos.
Declaração de conflito de interesse
Os autores declaram que a pesquisa foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.
Agradecimentos
Este trabalho foi apoiado pelo Centro de Pesquisa Colaborativa “Medo, Ansiedade e Transtornos de Ansiedade”, SFB-TRR 58, projeto B1.
Material suplementar
O Material Complementar deste artigo pode ser encontrado online em: http://journal.frontiersin.org/article/10.3389/fnbeh.2015.00128/abstract
Referências
Andreatta, M., Mühlberger, A., Glotzbach-Schoon, E. e Pauli, P. (2013). A previsibilidade da dor reverte as classificações de valência de um estímulo associado ao alívio. Frente. Syst. Neurosci. 7: 53. doi: 10.3389 / fnsys.2013.00053
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Andreatta, M., Mühlberger, A., Yarali, A., Gerber, B. e Pauli, P. (2010). Uma brecha entre a valência condicionada implícita e explícita após o aprendizado do alívio da dor em humanos. Proc. Biol. Sci. 277, 2411 – 2416. doi: 10.1098 / rspb.2010.0103
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Austin, AJ e Duka, T. (2010). Mecanismos de atenção para resultados apetitivos e aversivos no condicionamento pavloviano. Behav. Res do cérebro. 213, 19 – 26. doi: 10.1016 / j.bbr.2010.04.019
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Blumenthal, TD, Cuthbert, BN, Filion, DL, Hackley, S., Lipp, OV e van Boxtel, A. (2005). Relatório do comitê: diretrizes para estudos eletromiográficos de sobressalto do olho humano. Psicofisiologia 42, 1 – 15. doi: 10.1111 / j.1469-8986.2005.00271.x
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Bouton, ME e Peck, CA (1989). Efeitos de contexto no condicionamento, extinção e reintegração em uma preparação de condicionamento apetitivo. Anim Aprender. Comportamento. 17, 188 – 198. doi: 10.3758 / BF03207634
Texto Completo de CrossRef | Google Scholar
Bradley, MM (2009). Atenção seletiva natural: orientação e emoção. Psicofisiologia 46, 1 – 11. doi: 10.1111 / j.1469-8986.2008.00702.x
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Clark, JJ, Hollon, NG e Phillips, PEM (2012). Sistemas de avaliação pavloviana na aprendizagem e tomada de decisão. Curr. Opin. Neurobiol. 22, 1054 – 1061. doi: 10.1016 / j.conb.2012.06.004
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Delgado, MR, Jou, RL e Phelps, EA (2011). Sistemas neurais subjacentes ao condicionamento aversivo em humanos com reforços primários e secundários. Frente. Neurosci. 5: 71. doi: 10.3389 / fnins.2011.00071
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Dickinson, A. e Balleine, B. (1994). Controle motivacional de ação direcionada por objetivos. Anim Aprender. Comportamento. 22, 1 – 18. doi: 10.3758 / BF03199951
Texto Completo de CrossRef | Google Scholar
Fendt, M. e Fanselow, MS (1999). A base neuroanatômica e neuroquímica do medo condicionado. Neurosci. Biobehav. Rev. 23, 743 – 760. doi: 10.1016 / j.ijpsycho.2012.09.006
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Gottfried, JA, O'Doherty, J. e Dolan, RJ (2002). Aprendizagem olfativa apetitiva e aversiva em humanos estudados por meio de ressonância magnética funcional relacionada a eventos. J. Neurosci. 22, 10829 – 10837. Disponível online em: http://www.jneurosci.org/content/22/24/10829.full
Grillon, C., Baas, JM, Cornwell, B. e Johnson, L. (2006). Condicionamento contextual e evitação comportamental em um ambiente de realidade virtual: efeito da previsibilidade. Biol. Psiquiatria 60, 752 – 759. doi: 10.1016 / j.biopsych.2006.03.072
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Hamm, AO e Weike, AI (2005). A neuropsicologia da aprendizagem do medo e regulação do medo. Int. J. Psychophysiol. 57, 5 – 14. doi: 10.1016 / j.ijpsycho.2005.01.006
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Klucken, T., Schweckendiek, J., Merz, CJ, Tabbert, K., Walter, B., Kagerer, S., et al. (2009) Ativações neurais da aquisição da excitação sexual condicionada: efeitos da consciência de contingência e do sexo. J. Sex. Med. 6, 3071 – 3085. doi: 10.1111 / j.1743-6109.2009.01405.x
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Klucken, T., Wehrum, S., Schweckendiek, J., Merz, CJ, Hennig, J., Vaitl, D., et ai. (2013) O polimorfismo 5-HTTLPR está associado a respostas hemodinâmicas alteradas durante o condicionamento apetitivo. Cantarolar. Mapp do cérebro. 34, 2549 – 2560. doi: 10.1002 / hbm.22085
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Koch, M. (1999). A neurobiologia do sobressalto. Prog. Neurobiol. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Koch, M., Schmid, A. e Schnitzler, H.-U. (1996) A atenuação do prazer do sobressalto é interrompida por lesões do núcleo accumbens. Neuroreport 7, 1442–1446. doi: 10.1097/00001756-199605310-00024
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Krohne, HW, Egloff, B., Kohmann, C.-W. e Tausch, A. (1996). Untersuchungen mit einer deutschen versão do “Programa de Afectação Positiva e Negativa” (PANAS). Diagnostica 42, 139-156.
Kumar, P., Waiter, G., Ahearn, T., Milders, M., Reid, I. e Steele, JD (2008). Diferença temporal anormal recompensa-aprendizagem sinais na depressão maior. Cérebro 131, 2084 – 2093. doi: 10.1093 / brain / awn136
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Laux, L., Glanzmann, P., Schaffner, P. e Spielberger, CD (1981). O Estado-Traço Angstinventar. Weinheim: Teste de Beltz.
Levy, DJ e Glimcher, PW (2011). Comparando maçãs e laranjas: usando representação de valor subjetivo específico de recompensa e recompensa geral no cérebro. J. Neurosci. 31, 14693 – 14707. doi: 10.1523 / JNEUROSCI.2218-11.2011
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Lissek, S., Levenson, J., Biggs, AL, Johnson, LL, Ameli, R., Pine, DS, et al. (2008) Condicionamento elevado do medo a estímulos incondicionados socialmente relevantes no transtorno de ansiedade social. Sou. J. Psiquiatria 165, 124 – 132. doi: 10.1176 / appi.ajp.2007.06091513
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Martin-Soelch, C., Linthicum, J. e Ernst, M. (2007). Condicionamento apetitivo: bases neurais e implicações para a psicopatologia. Neurosci. Biobehav. Rev. 31, 426 – 440. doi: 10.1016 / j.neubiorev.2006.11.002
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
McDannald, MA, Lucantonio, F., Burke, KA, Niv, Y. e Schoenbaum, G. (2011). O estriado ventral e o córtex orbitofrontal são ambos necessários para o aprendizado por reforço baseado em modelo, mas não em modelo livre. J. Neurosci. 31, 2700 – 2705. doi: 10.1523 / JNEUROSCI.5499-10.2011
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Milad, MR e Quirk, G. (2012). Extinção do medo como modelo para a neurociência translacional: dez anos de progresso. Annu Rev. Psychol. 63, 129 – 111. doi: 10.1146 / annurev.psych.121208.131631
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Pavlov, IP (1927). Reflexos Condicionados: uma Investigação da Atividade Fisiológica do Córtex Cerebral. Londres: Oxford University Press.
Prévost, C., Liljeholm, M., Tyszka, JM e O'Doherty, JP (2012). Correlatos neurais de transferência pavloviana-para-instrumental específica e geral dentro das sub-regiões amigdalares humanas: um estudo de fMRI de alta resolução. J. Neurosci. 32, 8383 – 8390. doi: 10.1523 / jneurosci.6237-11.2012
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Tranel, D. e Damasio, H. (1994). Correlatos neuroanatômicos das respostas da condutância da pele eletrodérmica. Psicofisiologia 31, 427–438. doi: 10.1111/j.1469-8986.1994.tb01046.x
Resumo PubMed | Texto Completo de CrossRef | Google Scholar
Palavras-chave: condicionamento clássico, recompensa, punição, reflexo de sobressalto, resposta de condutância da pele
Citação: Andreatta M e Pauli P (2015) Apetitoso vs. Condicionamento aversivo em humanos. Frente. Behav. Neurosci. 9: 128. doi: 10.3389 / fnbeh.2015.00128
Recebido: 11 February 2015; Aceito: 05 May 2015;
Publicado: 19 May 2015.
Editado por:
Niels BirbaumerUniversidade de Tuebingen, Alemanha
Revisados pela:
Tim KluckenJustus Liebig University Giessen, Alemanha
Alessandro AngrilliUniversidade de Pádua, Itália
Copyright © 2015 Andreatta e Pauli. Este é um artigo de acesso aberto distribuído sob os termos do Licença Creative Commons Attribution (CC BY). O uso, distribuição ou reprodução em outros fóruns é permitido, desde que o (s) autor (es) original (is) ou licenciador (s) sejam creditados e que a publicação original desta revista seja citada, de acordo com a prática acadêmica aceita. Não é permitida a utilização, distribuição ou reprodução que não esteja em conformidade com estes termos.
* Correspondência: Marta Andreatta, Departamento de Psicologia (Psicologia Biológica, Psicologia Clínica e Psicoterapia), Universidade de Würzburg, Marcusstraße 9-11, D-XUMUM Würzburg, Alemanha, [email protegido]