KOMMENTARER: Kontroversavsnittet i detta dokument innehåller följande:
Det finns ett antal elektrofysiologiska baserade studier av Prauses grupp som ger bevis på att sexuell lust, inte hypersexualitet, förutsäger självreglering av sexuell upphetsning [92]. Dessa utredare föreslår i annat arbete att försökspersoner som rapporterade problem som reglerar deras visning av visuella sexuella stimuli (VSS) som också rapporterade högre sexuell lust visade lägre sena positiva potentialer (LPP) som svar på VSS. Författarna föreslår att detta mönster verkar annorlunda från ämnesberoende modeller [93].
De två EEG-artiklarna från "Prause's group" har kritiserats i stor utsträckning i peer-reviewed tidskrifter och av andra (inklusive YBOP). I verkligheten överensstämmer resultaten i båda artiklarna med missbruksmodellen, trots deras författares tvivelaktiga slutsatser. För EEG-studien 2013 se:
- Ingenting korrelerar med ingenting i SPAN Labs nya porrstudie (2013)
- Granskad kritik: "High Desire" eller "bara" ett missbruk? Ett svar till Steele et al. (2014) av Donald L. Hilton, Jr., MD,
- Peer-granskad kritik: “Neurovetenskap för internetpornografiberoende: En recension och uppdatering” (2015)
- Peer-granskad analys: ”Neurala korrelationer av sexuell köreaktivitet hos individer med och utan tvångsmässiga sexuella beteenden” (2014)
För 2015 EEG-studien se:
- Kritik av "Modulering av sena positiva potentialer av sexuella bilder hos problemanvändare och kontroller som är oförenliga med" Porn Addiction "(2015)"
- Peer-granskad kritik: Neuroscience of Internet Pornography Addiction: A Review and Update (2015)
Skulle det inte vara bra om forskare faktiskt läser de studier de citerar snarare än att bara papegoja huvudförfattarens rubriker?
LÄNK TILL PUBLIKATION
Publicerad: Oktober 12, 2015 (se historia)
DOI: 10.7759 / cureus.348
Citera denna artikel som: Blum K, Badgaiyan RD, Gold MS (October 12, 2015) Hypersexualitetsberoende och tillbakadragande: fenomenologi, neurogenetik och epigenetik. Cureus 7 (10): e348. doi: 10.7759 / cureus.348
Abstrakt
Hypersexualitet har definierats som onormalt ökad sexuell aktivitet. Epidemiologiska och kliniska studier har visat att detta icke-parafila tillstånd består av ”överdrivna” sexuella beteenden och störningar åtföljda av personlig nöd och social och medicinsk sjuklighet. Det är ett mycket kontroversiellt och politiskt ämne när det gäller hur man bäst kan kategorisera det som likartat eller inte lik beroendeframkallande beteende inklusive missbruk. Hypersexuell sjukdom är begreppsmässig som en icke-parafil sexuell luststörning med impulsivitet. Patofysiologiska perspektiv inkluderar dysreglering av sexuell upphetsning och lust, sexuell impulsivitet och sexuell tvångsmakt. Kärnan accumbens, belägen i det ventrala striatumet, förmedlar de förstärkande effekterna av missbruksdroger, såsom kokain, alkohol, nikotin och mat samt musik. Det antas faktiskt att denna struktur mandat beteenden framkallas av incitament stimuli. Dessa beteenden inkluderar naturliga belöningar som utfodring, dricka, sexuellt beteende och utforskande rörelse. En viktig regel för positiv förstärkning är att motoriska svar kommer att öka i storlek och kraft om de följs av en givande händelse. Här antar vi att det finns en gemensam verkningsmekanism (MOA) för de kraftfulla effekter som läkemedel, musik, mat och sex har på mänsklig motivation. Den mänskliga strävan efter de tre nödvändiga motiveringsbeteenden "hunger, törst och kön" kan alla ha vanliga molekylära genetiska antecedenter som, om de försämras, leder till avvikande beteenden. Vi antar att baserat på en mängd vetenskapligt stöd hypersexuell aktivitet verkligen är som droger, mat och musik som aktiverar hjärnans mesolimbiska belöningskretsar. Dessutom påverkar dopaminerg gen och eventuellt andra kandidat neurotransmittor-genpolymorfier både hedoniska och anhedoniska beteendemässiga resultat. Det finns lite känt om både genetik och epigenetik av hypersexualitet i den aktuella litteraturen. Vi förväntar oss dock att framtida studier baserade på bedömningar med kliniska instrument i kombination med genotypning av sexmissbrukare kommer att ge bevis för specifika kluster av sexuella typologier med polymorfa föreningar. Det har gjorts några studier med elektrofysiologiska tekniker som inte stöder uppfattningen att hypersexualitet verkligen liknar missbruk och andra beteendemissbruk. Författarna uppmuntrar också både kliniska och akademiska forskare att inleda forskning med hjälp av neuroimaging-verktyg för att undersöka naturliga dopaminerga agonistiska medel riktade mot specifika genpolymorfismer för att "normalisera" hypersexuellt beteende.
Introduktion & bakgrund
Visst har hypersexuellt beteende dokumenterats inom kliniska miljöer och forskningsinställningar under det senaste decenniet [1]. Benjamin Rush, en läkare, en av USA: s grundande far, dokumenterade kliniskt överdrivet sexuellt beteende [2] Richard von Krafft-Ebing, en västeuropeisk pionjärsexolog från 19-talet och Hirshfeld i 1948 fortsatte båda arbetet [3-4]. De grundläggande hyresgästerna för dessa utredare föreslog att hypersexualitet utgjorde en ihållande socialt avvikande sexuellt beteende hos både män och kvinnor, med överdriven sexuell aptit som dålig anpassning. I 1975 karakteriserade Barnvagn tillståndet som Don Juanism [5]. I 1969 föreslog Allen satyriasis för män och nymfomani hos kvinnor som stöds av Ellis och Sagarin [6-7]. Även om hypersexualitet inte ingår som en psykiatrisk diagnos i DSM, kan arbete av ett antal samtida utredare inklusive Kafka, Reid, Bancroft, deras kollegor och lutningen från Världshälsoorganisationen leda till att detta tillstånd inkluderas som en separat diagnostisk enhet [8-11].
översyn
Litteraturmetodik
Medline-databasen, per den 12 juli 2015, användes för att utföra en internetbaserad litteratursökning. Följande termer inkluderades: hypersexuell (170), hypersexualitet (479), sexuell missbruk (1,652), sexmissbrukare (1,842), sexuell impulsivitet (989), tvångsmässig sexuell (946), tvångsmässig sex (1,512 782), sexuell tvång (234) ), parafili-relaterad sjukdom (857) och överdriven sexuell (17). Eftersom den här artikeln är en kort recension snarare än en metaanalys baseras den på ett representativt urval av dessa studier som har relevans för de ämnen som omfattas. Att inte inkludera någon särskild studie förnekar inte dess betydelse för fältet. Det är förståeligt att det finns de som inte håller med konceptet att sexmissbruk verkligen är en verklig störning och till och med kan ha bevis för att de är på fast mark. Vi hävdar dock att det finns gott om bevis för att föreslå att sexuell missbruk existerar och studier som involverar neuroimaging, neurogenetics och till och med epigenetics stöder uppfattningen att tvångsmässig sexmissbruk såväl som hypersexualitet kan betraktas som en beroendeframkallande störning. Även om vi är medvetna om denna avvikelse lämnar vi in vår synpunkt för att generera ytterligare vetenskaplig granskning och inte för att skingra vetenskaplig retorik om detta viktiga ämne. För att belysa ämnet ytterligare har vi sökt i litteraturen för en viss konsensus. Den 2015 augusti 46 sökte vi PUBMED Central med följande term - “Är sex ett verkligt missbruk?” och hämtat XNUMX artiklar.
Definition av sexuellt beroende
Sexuellt beroende definieras som alla tvångsmässiga sexuella beteenden som stör det normala livet och orsakar allvarlig stress på familjen, vänner, nära och kära och ens arbetsmiljö. Sexuellt beroende har kallats sexuellt beroende, hypersexualitet och sexuell tvångsmakt. Under alla namn är det ett tvångsmässigt beteende som helt dominerar missbrukarens liv. Sexmissbrukare gör sex till en prioritet viktigare än familj, vänner och arbete. Sex blir den organiserande principen för missbrukarnas liv. De är villiga att offra det de värdesätter mest för att bevara och fortsätta sitt ohälsosamma beteende [12]. Hypersexuell lust har avgränsats som lust baserat på en livslängdsbedömning av frekvensen av sexuellt beteende och tid tillhörande sexuella fantasier. Hos män definierades en utvärdering av hypersexuell lust av Kafka och Hannen som den högsta varaktiga perioden (minst sex månaders lägsta varaktighet) av ihållande antagen sexuellt beteende (total sexuellt utlopp / vecka efter ålder 15). I själva verket identifierades en longitudinell historia av hypersexuell önskan, operativt definierad som ovan, hos 72-80% av män som söker behandling för paraphilias och paraphilia-relaterade störningar [13].
Hypersexualitet och könsskillnader
Det är väl etablerat att inom den mänskliga sexuella gemenskapen och litteraturen är sexuell lust kapitulerad som närvaron av sexuella fantasier, aktiviteter eller uppmaningar och motivation av människan att engagera sig i sexuella beteenden. Det finns både interna och externa relevanta ledtrådar [14]. Förespråkare av evolutionsteorin har hävdat att män och kvinnor har olika dagordningar när det gäller sexuell aktivitet [15]. Många studier visar tydliga skillnader mellan män och kvinnor. Män har ökat sexuell fantasi [16], ökad frekvens av onani [17], ökad benägenhet för externt genererad visuell sexuell upphetsning [18] tillåtna attityder till casual sex [19], lätthet av upphetsning [20]och iboende motivation [21]. Däremot visar kvinnor ett annat sexuellt landskap med sexuell motivation, sexuell upphetsning och sexuellt beteende som formas av evolutionära faktorer [22] och större biologiska, emotionella och temporära investeringar i reproduktion och uppfödning av barn [23]. Kvinnor är mindre sårbara för hypersexualitet [24] och anpassad för att främja anknytande relationer och långsiktigt partnerskap [25]. Medan sexuellt beroende uppskattas drabbas upp till 3% till 6% av befolkningen, är den tydliga förståelsen för de neurobiologiska antecedentema begränsad [26] såväl som kliniska bedömningar [27]. Vi uppmuntrar vidare att läsa om sexuell tvång, vidhäftning och sexuell läggning [28]och könsskillnader i svar på sexuella stimuli [29-30].
Det är anmärkningsvärt att Kafka och Hennen [13], fann att medelåldern för början av ihållande hypersexuellt beteende var 18.7 ± 7.2 år hos sexuellt aktiva män och åldersintervallet för början av hypersexuellt beteende var ålder 7-46. Den genomsnittliga varaktigheten för denna högsta konsekvent bibehöll frekvens av sexuellt aptitligt beteende var 12.3 ± 10.1 år. Men medelåldern för dessa aktiva sexuella män som var hypersexuella som sökte behandling var 37 ± 9 år. Hanson et al. utvärderade också hypersexualitet hos gärningsmän och fann att låga gärningsmän hade lägre recidivism än högriskbrottslingar [31].
Hypersexualitet och missbruk av dödlig substans
Det finns en hög morbiditet mellan hypersexuell störning och andra missbruk, som t.ex. [32-33]. I synnerhet föreslog Garcia och Thibaut att fenomenologin för överdriven icke-parafilisk sexuell störning skulle klassificeras som ett beroendeframkallande beteende snarare än en tvångssyndrom eller en impulskontrollstörning [34]. De påpekar med rätta att kriterierna är ganska nära de för beroendeframkallande sjukdomar som också föreslagits av andra [35]. Dessa utredare har gett drivkraft för fortsatt forskning inom detta område och möjlig framtida inkludering av hypersexuell störning i DSM-6. Drogmissbruk, rock 'n' roll och sex förekommer samtidigt, och hela festivaler har byggts kring dessa kombinationer från Woodstock till nutid.
Litteraturen avslöjar att användare av metamfetamin rapporterar att detta stimulerande läkemedel ökar sexuell lust, särskilt riskabelt beteende. Amfetamin har emellertid visat sig minska den sexuella aktiviteten hos kvinnliga råttor. Med detta i åtanke håller Holder et al. utvärderade metamfetaminens roll i honråttor [36]. De fann att tvärtom, metamfetamin underlättade kvinnligt sexuellt beteende, och denna effekt beror på förbättring av dopaminerg överföring och till och med möjlig neurotransmission på grund av kombinationen av äggstockshormoner och metamfetamin. Specifikt fann de en förbättring av sexuell motivation i kombination med aktivering av neuronal aktivitet i den mediala amygdala och ventromediala kärnan i hypotalamus.
Dessutom har forskare från Nederländerna studerat ämnets missbruk hos självidentifierade swingers [37]. I denna studie har Spauwen et al. drog slutsatsen att 79% av swingers rapporterade droganvändning (inklusive alkohol och användning av droger med erektil dysfunktion); 46% av dem rapporterade multipel läkemedelsanvändning. I själva verket var rekreationsdroganvändning (exklusive alkohol och droger med erektil dysfunktion) signifikant kopplad till sexuellt beteende hos män och kvinnor. Även droganvändning var oberoende förknippad med sexuellt överförda infektioner (STI) hos kvinnliga swingers, särskilt de som deltar i gruppsex.
Castelo-Branco et al. rapporterade att unga vuxna kvinnor uppfattar att sexualitet är en viktig del av deras liv men inte en primär oro (77.6%) [38]. De rapporterade också att alkohol tar bort hindren för att ha sex (62.3%). Viktigare fann de också att alkoholmissbruk var en förutsägbar variabel för att förbättra riskabelt beteende oberoende av kvinnans ålder.
Det är anmärkningsvärt att Jia et al. rapporterade farligt sexuellt beteende bland missbrukare av psykostimulant och heroin, inklusive flera sexuella samlag, avslappnade sexuella partners, homosexuella partners och aldrig eller ibland öva på säkert sex [39].
Vår viktigaste hyresgäst är att droger, såsom metamfetamin, kokain, heroin och alkohol, kan stimulera sexuell lust hos icke-missbrukare. Hos missbrukare är det helt annorlunda; samma läkemedel kan orsaka anhedoni på kronisk basis. Emellertid har eftermissbruk under återhämtningsfasen i många fall observerats afrodisiakalt beteende.
Hypersexualitet och tillbakadragande
En PubMed-sökning (7-19-15) med termen "hypersexualitet och abstinenssymptom" resulterade i endast fem artiklar, varav ingen beskrev "abstinenssymtomatologi." En alternativ sökning med termerna ”abstinenssymptom med hög sexuell aktivitet” resulterade dock i 25 listade artiklar.
Narkomaner i återhämtning rapporterar ökningar av ätande och en aptitdrivning för vissa livsmedel och missbruk av cigaretter under långvarig avhållsamhet. Viktökning har också visats och dokumenterats hos nyligen och långvariga abstinenta djur och människor [39]. Bruijnzeel gjorde den intressanta iakttagelsen att akut opiatavlägsnande kan resultera i spontana orgasmer [40]. I artikeln föreslog det viktigt att Bruijnzeel föreslog att abstinenssymtomatologi från läkemedel och eventuellt kronisk intensiv sexuell aktivitet kan bero på den opåverkade funktionen av kappa opioidreceptorsignaler som hämmar frisättningen av dopamin samtidigt som norepinefrin ökar i hjärnans belöningskretsar.
Artiklarna i litteraturen led av bristande noggrannhet angående akut och långvarig tillbakadragande och avhållsamhet bekräftad genom urinprovning. Kliniska behandlingsprogram har i allt högre grad gått från att behandla båda könen till att isolera de två könen, tillhandahålla könsspecifika behandlingsprogram. De erbjuder också utbildning för att ta itu med de tidiga och långvariga ökningarna av sexuellt intresse och aktivitet, och förhållandet mellan återhämtning till mat och överätande.
Hypersexualitetsinducerade abstinenssymtom har rapporterats av vissa utredare med varierande grad av svårighetsgrad och samtidigt sjukligt missbruk [41-45]. Som ett resultat av denna sökning hittade vi inte en enda uppsats som beskrev faktiska abstinenssymtom förknippade med avhållsamhet från aktiva sexuella möten. De flesta av de citerade handlingarna involverade effekterna av borttagande från missbruk av droger, såsom opioider, nikotin, amfetamin och kokain, vilket kan försämra sexuell aktivitet.
Hypersexualitet och neurogenetik
En PubMed-sökning (7-19-15) avslöjade endast sex listade artiklar som använde termen ”gener och hypersexualitet” som mest fokuserade på artiklar relaterade till Kleine-Levins syndrom (KLS), en mycket sällsynt sjukdom där hypersexualitet kan vara upp till 27 år. I en studie visade det sig att ett immunsvarigt HLA-DQBl, DQBl * 0602 detekterades i signifikanta mängder hos patienter med KLS och kunde höja risken för KLS [46-47].
Men när vi använde termerna ”sexuell aktivitet och gener” listades 2,826 artiklar och vi ger en kort sammanfattning av några viktiga neurogenetiska aspekter. Det är vår hypotes att både hedoniskt och anhedoniskt beteende är resultat i en del av en individs riskalleler för dessa beteenden och att behandlingen består av att rikta in sig på dessa identifierade polymorfismer. Dessutom beror behandlingssvar också på dessa riskalleler och ger en viktig grund för farmakogenetisk testning och farmakogenomiska / nutrigenomiska lösningar.
Efter det kontroversiella initiala konstaterandet från Blum et al. i 1990 av det första beviset för en koppling mellan DRD2 Al-allelen och allvarlig alkoholism har det förekommit 3,938-artiklar i PubMed (7-19-15) [48]. Studierna täcker den psykiatriska genpolymorfismen, DRD2 Al-allelen och tillhörande beteenden och fysiologi. Det finns emellertid ett brist på data som kopplar sexuell aktivitet till denna och andra relaterade gener trots de överväldigande bevisen för mesolimbisk aktivering, särskilt i dopaminergiska vägar och neuronalokaler relaterade till sexuella stimuli och aktivitet. Det är anmärkningsvärt att Blum och Noble korrekt klassificerade DRD2-genen som en generaliserad belönningsgen som är ansvarig för allt belöningsbristsyndrom (RDS) beteende. Faktum är att använda Bayesianska teoremanalysbärare av Taq Al-allelen under deras livstid har 74% chans att de kommer att möta ett eller flera belöningsunderskottssyndrom (RDS) beteende [49].
Den första föreningen av någon genpolymorfism och sexuell aktivitet inträffade inte förrän 1999 när Miller et al. utvärderade några dopaminerge gener [50]. Den grundläggande upptäckten är att det dopaminergiska systemet i hjärnan verkar spela en viktig roll i regleringen av sexuellt beteende. Förhållandet mellan generna för Dl-, D2- och D4-dopaminreceptorerna och ålder vid första samlag (AFSI) undersöktes i ett prov av 414 icke-spansktalande, europeisk-amerikanska män och kvinnor. En signifikant association observerades mellan en DRD2-allel och AFSI och en ännu starkare associering när DRD2-allelen interagerades med en DRDl-allel. En begränsad regressionsmodell konstruerades för att förutsäga AFSI med användning av kön och en grupp av nio psykosociala variabler som prediktorer. Att lägga till DRD2- och DRD2-för-DRD1-prediktorerna till denna modell ökade den förklarade variansen med 23% respektive 55%. Det faktum att dessa fynd tyder på en starkare förening bland män än hos kvinnor är i överensstämmelse med andras nyligen uppvisade högre sexuella stimulansrespons hos män än hos kvinnor [51]. Så kanske "män är från Mars och kvinnor från Venus" och detta kan till och med vara sant för kokainmissbruk [52].
Specifikt har både prekliniska och kliniska studier visat sexuellt dimorfa mönster i beteendeansvar på kokain i alla faser av kokainberoende (induktion, underhåll och återfall). Därför framträder en tydlig bild som antyder att det finns en biologisk grund för könsspecifika skillnader i kokainberoende. Dessa skillnader härrör från den olika regleringen av CNS med manliga och kvinnliga gonadala hormoner och kan förutsägas av närvaron av DRD2-genpolymorfismer [53]. Dessutom är det känt att genetiska samband mellan COMT och olika psykiatriska fenotyper ofta visar skillnader mellan män och kvinnor. Dessa inkluderar den funktionella Val (158) Met polymorfism i COMT förknippas med tvångssyndrom hos män och med ångestfenotyper hos kvinnor. Val (158) Met polymorfism i COMT har dessutom en större inverkan på kognitiv funktion hos pojkar än flickor [54].
Miller et al. hittade inte en associering av polymorfismerna kopplade till DRD4-genen och ålder för första samlag [50]. Andra fann dock en betydande förening i vissa etniska grupper. Specifikt indikerar deras analys av polymorfismerna i DRD4 att de med någon - 3R-genotyp upplevde en risk för första samlag högre än de med någon annan (eller någon - 4R) genotyp i alla etniciteter (n = 2,552). Intressant nog skiljer sig inte risken för första kön mellan de två genotyperna i det afroamerikanska urvalet, vilket väcker frågan om kulturell uppfostran [55].
Den sexuella upplevelsen, liksom upprepad droganvändning, ger långvariga förändringar, inklusive sensibilisering i nucleus accumbens (NAc) och ryggstriatum. Bradley et al. med hjälp av mikroarray-analys för att studera hamstrar som för första gången fann att den sexuella upplevelsen hos antingen manliga eller kvinnliga djur differentiellt upp eller ner reglerar mRNA-uttryck för en serie gener i NAc [56]. De fann att i jämförelse med sexuellt naiva djur uppvisade sexuellt erfarna hamstrar som fick en stimulanshane på vecka 7 en ökning av ett stort antal gener. Omvänt uppvisade sexuellt erfarna kvinnliga hamstrar som inte fick en stimulanshane på vecka 7 en minskning av uttrycket för många gener. Enligt författarna kan denna första genprofilering någonsin i kvinnliga hamstrar ge en inblick i mekanismerna genom vilka både beteende (kön) och missbruksläkemedel inducerar långvariga förändringar i de mesolimbiska och nigrostriatala dopaminvägarna.
Bipolära elektroder, implanterade bilateralt i lateral hypothalamus och substantia nigraventral tegmental område, användes stereotaxiskt för att ge kronisk självstimulerande belöningsupplevelser som liknar sexuellt beteende. Denna typ av stimulering har visat sig inducera en signifikant ökning i antalet synapser i CA3-regionen i hippocampus och det molekylära lagret av motorcortex hos råttor. I huvudsak inducerade kronisk hjärnstimulering långvarig potentiering (LTP), vilket är känt för att öka nya synaptiska anslutningar [57]. En enda exponering för kokain hos naiva djur är tillräcklig för att utlösa långvariga förändringar i glutamatergiska synapser i ventral tegmental area (VTA) som liknar aktivitetsberoende LTP i andra hjärnregioner. Denna kokaininducerade LTP verkar medieras via dopamin D5-receptoraktivering av N-metyl-D-aspartat (NMDA) receptorer och kräver proteinsyntes [58], som återigen stöder vårt förslag som här föreslagits att läkemedel och kön kan ha vanliga neurokemiska underlag.
Empirisk forskning har avslöjat ett positivt samband mellan antalet sexpartners och engagemang i antisociala beteenden [59]. De flesta försök att förklara denna förening har tagit ett evolutionärt perspektiv. Ur evolutionsperspektivet är samma egenskaper, till exempel impulsivitet, kortsiktighet och aggressivitet, som är relaterade till ett stort antal sexpartner, också relaterade till kriminellt engagemang. Det finns emellertid också anledning att tro att samvariationen mellan könspartners och brottsbeteenden delvis kan förklaras med en gemensam genetisk väg, där gener som är relaterade till könspartnerantal också är relaterade till antisocialt beteende. Med användning av ovan beskrivna skäl, Beaver et al. hittade ett starkt positivt samband mellan könspartners och antisocialt beteende och polymorfism av dopamintransportgenen (DAT1) förklarar variation i både antal sexpartner och kriminellt beteende för män [59]. Den polymorfiska effekten av DATl-genen och antalet sexuella partners kan bero på en associering som har hittats mellan vissa polymorfismer och för tidig utlösning av manlig penil. Bärare av 1OR / 1OR-genotypen hade poäng som indikerade en lägre tröskel att ejakulera på var och en av indikatorerna jämfört med den kombinerade bärargruppen 9R9R / 9R10R (9R högre aktivitet lägre dopamin) [60]. Polymorfismer av DATl-genen, specifikt 10R / 10R-genotypen, har hittats i ungdomliga brottslingar som deltar i Brown School (San Marcos, Texas) för patologiskt aggressivt beteende, inklusive antisocialt beteende [61]. En positiv korrelation mellan både DRD2 och DATl-polymorfismerna observerades med patologiskt våld hos ungdomar i en blindad klinisk prövning. Även om de ursprungligen konceptualiserats som en följd av peer-imitation av barndom eller livslängd-persistent ungdom, finns det alltmer bevis från tvillingstudier om att ungdomar-start eller ungdomsbegränsat antisocialt beteende också kan påverkas genetiskt. Burt och Mikolajewski bekräftade inte bara dessa fynd med DATl-genen utan utvidgade dessa fynd till att omfatta His452Tyr-varianten av genen som kodar för 5-HT2A-receptorn också [62]Mer nyligen har Jozkow et al. rapporterade en koppling mellan den sexuella dimensionen av åldrande hannes symptom (AMS) och genetiska varianter av 5-HTRlB G861C [63]. Dessutom har Sales et al. hittades genom multivariabel logistisk regressionsanalys, en interaktion mellan missbruk och 5-HTTLPR-gruppen, varigenom det fanns en signifikant förening med status som inte förändrades, tillsammans med partnerkommunikationsfrekvensresultat vid uppföljning [64] Att ha en historia av missbruk var signifikant förknippad med större odds för icke-förändring i kondomanvändning efter intervention för endast de med s-allelen.
Det är känt att polymorfismer i icke-kodande regioner i vasopressin la receptorgen (Avpr la) är kopplade till socioemotionella egenskaper hos människor, schimpanser och voles, och kan bero på en platsspecifik variation i genuttryck. Enligt Barrett m.fl. erbjuder den socialt monogama prärievollen en unik möjlighet att studera neurobiologin i monogami [65]. Faktum är att vasopressin la receptor (VlaR) signalering är nödvändig för bildandet av parbindningen hos män. Intressant nog uppvisar sociala prärievolter större VlaR-bindning i belöningsbearbetningen ventral pallidum än asociala volader av samma släkte. Barrett et al. fann att nedreglering av pallidal VlaR-densitet resulterade i en signifikant försämring av preferensen för en parad kvinnlig partner och en minskning av ångestliknande beteende i vuxen ålder [65]. Annat arbete av Garcia et al. avslöjade att individer med minst en 7-repeterande allel (7R +) av DRD4 rapporterar en större kategorisk frekvens av promiskuöst sexuellt beteende, inklusive att någonsin ha haft en "one-night stand", och rapporterar en mer än 50% ökning av sexuell otrohet [66].
Det är viktigt att Daw och Guo rapporterade att individer som bär genotyperna DRD2 * Al / A2, DRD2 * A2 / A2, DATl * 9R / 10R och MAOA * 2R / är förknippade med högre odds på oskyddade samlag än andra genotyper på dessa [67]. DRD2-föreningarna gäller både män och kvinnor, medan de andra länkarna endast gäller kvinnor. Slutligen, Emanuele, et al. rapporterade en signifikant koppling mellan DRD2 TaqI A-genotyper och ”Eros · (en kärleksfull stil som kännetecknas av en tendens att utveckla intensiva känslomässiga upplevelser baserat på den fysiska attraktionen till partnern), liksom mellan C516T 5HT2A-polymorfism och” mani ”( en besittande och beroende romantisk anknytning, kännetecknad av självdödande känslor) [68].
Epigenetik och sexuell aktivitet
En genomgång av litteraturen avslöjar att ett antal nya artiklar påpekar vikten av epigenetiska effekter på sexuell aktivitet. Till exempel granskade Matsuda de epigenetiska förändringarna av östrogenreceptorn a (ERalpha) och påverkan på sociosexuellt beteende [69]. Faktum är att förändring av ER-alfa-genaktivitet förmedlad av epigenetiska mekanismer, såsom histonmodifikationer och DNA-metylering, förändrar ens sexuella beteenden. När det gäller homosexualitet, Rice, et al. utvecklat en modell som kan förklara kanalisering (konvertering) sexuell utveckling av homosexualitet [70]. De förklarar att denna modell är baserad på epigenetiska märken som fastställts som svar på XX mot XY-karyotyp i embryonala stamceller. Följaktligen förbättrar dessa märken känsligheten för testosteron i XY-fostrar och sänker det i XX-fostrar, varigenom den sexuella utvecklingen kanaliseras. Det har antagits att en delmängd av dessa konverterande epigenetiska märken kvantitativt kan överföra transgenetiskt, och kan leda till mosaik för sexuell utveckling hos avkommor av motsatt kön - den homoseksuella fenotypen.
I den socialt monogama prärievollen (Microtus ochrogaster) inducerar parning varaktiga parbindningar som initieras av partnerpreferensbildning och regleras av olika neurotransmittorer, inklusive oxytocin, vasopressin och dopamin. Arbet av Gundersen [71]och Wang et al. [72] antyder att histondeacetylas kan underlätta bildning av partner i kvinnliga prärievolter som kan ha relevans för människor. Specifikt Wang et al. fann att histonedeacetylas-hämmare-natriumbutyrat och trikostatin A (TSA) förbättrade bildningsförmågan hos kvinnliga prärievolter [72]. Denna partnerpreferensbildning var associerad med en uppreglering av oxytocinreceptor (OTR, oxtr) och vasopressin V la-receptor (VlaR, avprla) i NAc genom en ökning av histonacetylering vid deras respektive promotorer.
Det finns intresseväxande bevis som tyder på att kvinnor aktivt engagerar sig i polyandry antingen för att undvika genetisk inkompatibilitet eller för att fördärma faderskap till förmån för genetiskt överlägsna män. Det finns möjlighet att val av överlägsen manlig kondition kan bero på epigenetiska effekter. Enligt Zeh och Zeh, till skillnad från DNA-sekvensbaserad variation, kan epigenetisk variation starkt påverkas av miljömässiga och stokastiska effekter som upplevs under en individs livstid [73]. De antyder att epigenetisk variation kan vara viktig för den sexuella selektionen efter samställningen och kan redogöra för fynd som kopplar konkurrensförmåga till spermier till avkommans kondition.
Genetisk och meme evolution: mänsklig förökning
Eysenck föreslog en positiv korrelation mellan extraversion och intensifierat sexuellt beteende och mellan neuroticism och problem i sexuellt beteende (antisocialt beteende). En tidigare studie med gifta personer visade inte någon av dessa samband. Det antogs att denna anslutning endast finns för ogifta personer som inte är engagerade i långvariga relationer eftersom kvaliteten på förhållandet avgör den sexuella interaktionen. Inom ett urval av unga ogifta män fanns det ett positivt samband mellan extraversion och föremål där personen beskrev tidigare sexuell aktivitet med fler individer och i högre frekvens. Ingen korrelation hittades med neuroticism. Det fanns också små korrelationer med andra personlighets- och sociala attitydskalor. På grund av korrelationen med en fungerande personlighetsskala tolkades resultaten ur ett socialpsykologiskt perspektiv. I dagens samhälle förväntas den unga mannen ta initiativ till en sexuell interaktion som en extravert ung man kan förstå bättre än en introvert. [74]. Detta perspektiv överensstämmer direkt med Richard Brodies idé om själviska gener i sinnet [75]. Ur DNA: s synvinkel är naturligtvis antropologer överens om att "vi är fortfarande här bara av en anledning; att gå ut och föröka sig. ” Medan evolutionära framsteg går långsamt, ett steg var 20: e år eller så, jämfört med "meme-evolution, muterar en idé den tiden det tar att läsa en mening." Våra hjärnor har inget att göra med genetisk utveckling förutom eftersom det är relaterat till smarta människor som har färre barn. Faktum är att om det finns gener som ger människor en tendens att ta på sig memes som begränsar deras antal avkommor, kommer de att dö ut inom några generationer till förmån för gener som ger människor en tendens att förvärva barn. Även om det är något kontroversiellt tyder ett antal studier tyvärr på att Homo sapiens under de senaste 42,000 XNUMX åren har sänkt sina IQ på grund av selektiv parning [76].
Extraordinärt visar det sig att eftersom extraversion är kopplad till ökad sexuell aktivitet, särskilt hos män, uppskattar kvantitativ genetiker arvbarheten hos den extraverterade personligheten till omkring 40-60%. Smillie och medarbetare studerade och fann att en kopia av DRD2-genen Al-allelen var associerad med signifikant högre extraversion [77]. Denna förening väcker en intressant fråga när det gäller mänsklig fortplantning. Comings föreslog att på grund av deras markanta effekt på reproduktionsbeteende, inlärningssjukdomar och andra impulsiva, tvångsmässiga, aggressiva och beroendeframkallande störningar har bärarna av DRD2 Al potential att orsaka progressiva och permanenta förändringar i frekvensen av DRD2Al-allelen som " den genetiska smältningen av arten ” [78].
I sin bok ger Comings bevis på att personer med beroendeframkallande beteende har barn tidigare, och detta påverkar valet av missbrukgener som DRD2 Al-allelen [79]. Han föreslår att individer som bär denna störande riskallel kommer att ha barn, låt oss säga vid 20 års ålder och individer utan denna allel kommer att få barn vid 25 år. Som ett resultat kommer den mutanta genen att reproducera snabbare, nämligen vart 20: e år medan den normala formen av genen kommer att reproducera var 25: e år. Förhållandet 25/20 är 1.25. Således kommer hastigheten med vilken en gen som har en 1.25-faldig selektiv fördel kommer att öka i frekvens från generation till generation. En skillnad på fem år i åldern hos mödrar eller fäder när de får sina första barn är tillräcklig för att resultera i ett signifikant och relativt snabbt urval för gener som bärs av gruppen som initierar fertilitet vid en tidigare ålder. Ökningar av vissa RDS-beteenden har dokumenterats från 1955 till nu. Dessa ökningar inkluderar ungdomars beteendesyndrom (droger, kön, tonårsgraviditeter och kriminella beteenden, rökning), beteendestörning, brottslighet, drogmissbruk, alkoholism, oskyddat sexuellt beteende, ogiftade mödrar, välfärd, utvisad skola och skolavbrott, samt en samtidig minskning av IQ [80]. Dessa resultat är baserade på Berkeley-studien som använder longitudinella data från barnhälso- och utvecklingsstudier och National Longitudinal Surveys of Youth eller NLYS [81]. Med hjälp av denna information förutspådde Comings att från 1955 till 2015 kommer det att bli en fördubbling av frekvensen av till exempel DRD2 Al-allelen, vilket därför ökar förekomsten av RDS-beteenden, inklusive äldre sexuellt samlag [50]. Vi uppmuntrar till en uppföljning av denna intressanta förutsägelse.
Trots viss oenighet föreslår vi hypersexualitetsstörning som en subtyp av RDS som delar egenskaper med substans- och icke-substansberoendeframkallande beteenden, eftersom dess kliniska uttryck delvis påverkas av både genetik och epigenetik. Även om det inte är testat för närvarande föreslår vi också kortvariga FDA-godkända medicineringsassisterade behandlingar (MAT) som gynnar blockering av dopaminfunktion följt av försiktig aktivering av dopaminerga vägar som leder till långvarig dopaminhomeostas. Det senare skulle kunna åstadkommas med vissa sätt som kan hjälpa till att återhämta sig.
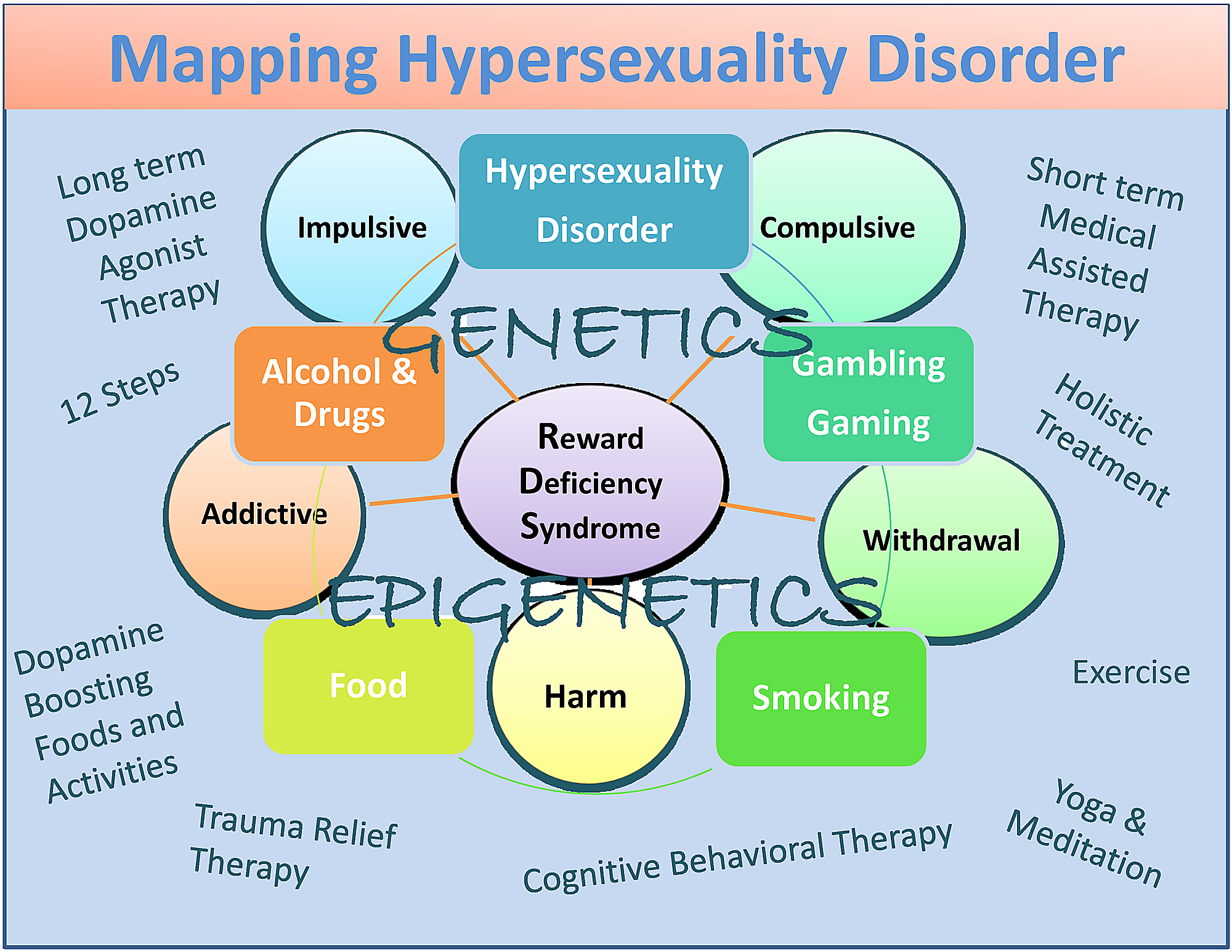
Även om potentiell förspänning inkluderar de dopamin-agonistterapi-nutraceuticals (KB220), 12 Step-programmet och traditionen, holistisk behandling, kognitiv beteendeterapi (CBT), och trauma lättnadsterapi (TRT) samt dopamin-boosting aktiviteter och livsmedel (figur 1) [82].
Kontroversen
Medan vi är övertygade om att hypersexualitetsstörning bör inkluderas i framtida utgåvor av DSM, är vi något förvirrade att så lite är känt om denna störning när det gäller neurogenetik och epigenetik och till och med abstinenssymtomatologi och övergripande fenomenologi [83]. Det främsta hemmeddelandet är att vi nu uppmuntrar det vetenskapliga samhället att genomföra experiment, särskilt inom området neuroimaging och neurogenetics, inklusive epigenetics specifika för gener, såsom oxytocin-vasopressin-orexin-dopamin samt andra belöningsgener. Eventuellt kan detta tillstånd dra nytta av behandling som riktar belöning av genpolymorfismer för att hjälpa till att främja dopaminhomeostas [84-89]. Ett antal recensioner av Joranby et al. och Edge and Gold stöder vanliga behandlingsmöjligheter relaterade till delade neurokemiska mekanismer i hjärnbelöningskretsar, som framförts tidigare i RDS-konceptet [90-91].
Historiskt inkluderades "sexmissbruk" i DSM - III, men det togs bort från DSM-1V eftersom konsensusen från författarna till DSM-1V trodde att det inte fanns tillräckliga bevis för att förtjäna dess slutsats. Detta beslut fylldes av enorma känslor av ledare inom området. Efter denna händelse beslutade ett antal forskare inklusive Kafka, Reid, Prause och andra att inrama ”Hypersexualitet” inte som ett sexberoende utan en fristående psykisk störning och inte som en beroende i sig. Medan deras tidigare arbete under 2010 föreslog som det hänvisas häri att "hypersexualitet" liknade sexberoende och eventuellt andra missbruk, inklusive missbruk, stöder deras senaste arbete från detta påstående. Det senaste arbetet inom detta område avslöjar fortsatt kontrovers. Det finns ett antal elektrofysiologiskt baserade studier av Prauses grupp som ger bevis för att sexuell lust, inte hypersexualitet, förutsäger självreglering av sexuell upphetsning [92]. Dessa utredare föreslår i annat arbete att försökspersoner som rapporterade problem som reglerar deras visning av visuella sexuella stimuli (VSS) som också rapporterade högre sexuell lust visade lägre sena positiva potentialer (LPP) som svar på VSS. Författarna föreslår att detta mönster verkar annorlunda från ämnesberoende modeller [93]. Även om de inte involverade personer med hypersexuell störning har arbete av Voon's grupp visat att i tvångsmässigt beteendeobjekt, exponerade för sexuellt explicita videoklipp, en större aktivitet i nervnätverket liknar det som observerades i läkemedels-cue-reaktivitetsstudier [94]. Större önskan eller önskan snarare än att gilla var ytterligare förknippad med aktivitet i detta neurala nätverk. Detta arbete överensstämmer med teorier om stimulansmotivation [95].
Vi, författarna till den aktuella artikeln, erkänner att vi inte har varit medvetna om alla viktiga interaktioner som har inträffat mellan förespråkarna för “Hypersexuality Disorder” och deras ärliga avsikt att få denna störning inkluderad i den nuvarande DSM-5. Även om det misslyckades med det så kallade "acid-test", finns det all anledning att tro att det kommer att ingå i framtida utgåvor av DSM. Det är anmärkningsvärt att Steven Hyman, den nuvarande NIH-regissören, korrekt hävdade att ”DSM är en dålig spegel av kliniska och biologiska verkligheter; ett grundläggande nytt tillvägagångssätt för diagnostisk klassificering behövs eftersom forskare avslöjar nya sätt att studera och förstå psykisk sjukdom ” [96]. Dessutom har Casey et al. föreslog att även om DSM betraktar olika störningar som distinkta enheter, "är gränserna mellan störningar ofta inte så stränga som DSM antyder" [97].
I 2014, Karila et al. föreslog att sexuellt beroende, även känd som hypersexuell störning, har ignorerats av vissa psykiatriker, trots att tillståndet orsakade allvarliga psykosociala problem. Enligt dessa författare föreslår de att sexuellt beroende eller hypersexuell störning representerar olika termer för samma problem. De påpekar att prevalensgraden för sexuella missbrukrelaterade störningar varierar från 3% till 6%. Följaktligen visar konstruktionen av sexuellt missbruk / hypersexuell störning problematiska beteenden inklusive: överdriven onani, cybersex, användning av pornografi, avvikande sexuellt beteende med samtyckande vuxna, telefonsex, stripklubbbesök och annat beroendeframkallande beteende [98]. Vi håller helt klart med om att det kan finnas distinkta skillnader mellan sexuellt beroende och hypersexualitet som noterats av Carvalho et al. [99], Rettenberger, et al. [100]Kor, et al. [1], Reid, et al. [9], Kafka och Hennen [13]och Prause et al. [93-94] bland andra.
Sammanfattningsvis har vi föreslagit att även om det finns vissa skillnader mellan hypersexualitet och könsberoende, krävs mer forskning för att lämpligt kategorisera dessa mycket viktiga villkor. Vi håller med om Walters et al. [101] som antydde att individuella skillnader i hypersexualitet är kvantitativa snarare än kvalitativa till sin natur. De föreslår också att hypersexualitet är organiserad längs ett kontinuum som faller i den övre änden av kontinuummet (figur 1).

Bild 1: En beskrivande karta över hypersexualitetsstörning som en subtyp av RDS
Figuren illustrerar interaktiva neurogenetiska och epigenetiska effekter. Både kortvariga dopaminblockerande och långvariga "dopaminerga-homeostas" -baserade behandlingar och dopaminförstärkande terapier och dagliga aktiviteter listas. Cirklar anger RDS-egenskaper och rutor anger RDS-beteenden.
Slutsatser
Samtidigt som vi känner igen kontroversen föreslår vi att möjliga skillnader och likheter mellan hypersexualitetsstörning och sexberoende bör undersökas på lämpligt sätt med hjälp av neuroimaging (fMRI, PET, SPECT), optogenetik, kandidat- och mikroarrayanalys och epigenetiska tekniker. Vi tror att dessa undersökningar kommer att utgöra grunden för att inkludera hypersexualitet som en störning i framtida utgåvor av DSM.
Referensprojekt
- Kor A, Fogel Y, Reid RC, Potenza MN: Bör hypersexuell störning klassificeras som ett missbruk?. Sex Addict Compulsivity. 2013, 20:http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3836191/. 10.1080/10720162.2013.768132
- Skynda till B: Medicinska undersökningar och iakttagelser om sinnets sjukdom. Rush B (ed): Gryphon Editions Ltd., Birmingham, AL; (Originalverk publicerat 1812) granskat i 1979.
- von Krafft-Ebing R: Psychopathia Sexualis. Klaf FS (ed): Stein och Day, New York; (originalverk publicerat 1886) granskat i 1965.
- Hirshfeld M: Sexuella anomalier: Ursprung, art och behandling av sexuella störningar. Hirshfeld M (ed): Emerson Books, New York; 1948.
- Barnvagn RJ: Perversion: Den erotiska formen av hat. Pantheon Books, New York; 1975.
- Allen CA: En lärobok om psykoseksuella störningar. Oxford University Press, London; 1962.
- Ellis A, Sagarin E: Nymfomani: En studie av den överexkas kvinnan. Gilbert Press, New York; 1964.
- Kafka MP: Vad hände med hypersexuell störning?. Arch Sex Behav. 2014, 43: 1259-1261. 10.1007 / s10508-014-0326-y
- Reid RC, Temko J, Moghaddam JF, Fong TW: Skam, idissling och självkänsla hos män bedömdes för hypersexuell störning. J Psychiatr Practice. 2014, 20: 260 – 268. 10.1097 / 01.pra.0000452562.98286.c5
- Bancroft J: Mänsklig sexualitet och dess problem. Tredje upplagan. Elsevier, Oxford, England; 2009.
- Internationell statistisk klassificering av sjukdomar och relaterade hälsoproblem, 10th Revision . (2007). Åtkomst: juli 23, 2015: http://apps.who.int/classifications/apps/icd/icd10online2007/.
- Sångare B, Toates FM: Sexuell motivation. J Sexforskning. 1987, 23: 481 – 501. 10.1080/00224498709551386
- Kafka MP, Hennen J: Hyperseksuell lust hos män: skiljer sig män med parafili från män med parafili-relaterade störningar?. Sexmisbruk. 2003, 15: 307 – 321. 10.1023 / A: 1025000227956
- Buss DM, Schmitt DP: Teori för sexuella strategier: ett evolutionärt perspektiv på mänsklig parning. Psychol Rev. 1993, 100: 204 – 232. 10.1037 / 0033-295X.100.2.204
- Corbett-Detig RB, Hartl DL, Sackton TB: Naturligt urval begränsar neutral mångfald över ett brett sortiment av arter. Psychol Bull. 1995, 13: e1002112.- 469-496. doi: 10.1371 / journal.pbio.1002112
- Laumann EO, Michael RT, Gagnon JH: En politisk historia av den nationella könsundersökningen av vuxna. Fam Plann Perspect. 1994, 26: 34 – 38. 10.2307/2136095
- Jones JC, Barlow DH: Självrapporterad frekvens av sexuella behov, fantasier och onani-fantasier hos heteroseksuella män och kvinnor. Arch Sex Behav. 1990, 19: 269 – 79. 10.1007 / BF01541552
- Oliver MB, Hyde JS: Könsskillnader i sexualitet: en metaanalys. Psychol Bull. 1993, 114: 29 – 51. 10.1037 / 0033-2909.114.1.29
- Okami P, Shackelford TK: Mänskliga könsskillnader i sexuell psykologi och beteende. Annu Rev Sex Res. 2001, 12: 186–241. 10.1080/10532528.2001.10559798
- Leitenberg H, Detzer MJ, Srebnik D: Könsskillnader i onani och förhållandet mellan onani erfarenhet av preadolescens och / eller tidig tonåring till sexuellt beteende och sexuell anpassning i ung vuxen ålder. Arch Sex Behav. 1993, 22: 87 – 98. 10.1007 / BF01542359
- Fetterman AK, Kruger NN, Robinson MD: Sexkopplade parningsstrategier avviker från en manipulation av könsförmågan. Motiv Emot. 2015, 39: 99 – 103. 10.1007/s11031-014-9420-7
- Basson R: Använda en annan modell för kvinnligt sexuellt svar för att ta itu med kvinnors problematiska låga sexuella lust. J Sex Marital Ther. 2001, 27: 395–403. 10.1080/713846827
- Andersen BL, Cyranowski JM, Aarestad S: Utöver konstgjorda, könsbundna distinktioner för att föreställa kvinnlig sexualitet: kommentera Baumeister. Psychol Bull. 2000, 126: 380 – 389. 10.1037 // 0033-2909.126.3.380
- Knight RA, Sims-Knight JE: Utvecklingens föregångare av sexuell tvång mot kvinnor: testa alternativa hypoteser med strukturell ekvationsmodellering. Ann NY Acad Sci. 2003, 989: 72 – 85. 10.1111 / j.1749-6632.2003.tb07294.x
- Blum K, Werner T, Carnes S, Carnes P, Bowirrat A, Giordano J, Oscar-Berman M, Gold M: Sex, droger och rock 'n' roll: hypotes om vanlig mesolimbisk aktivering som en funktion av belöningsgenpolymorfismer. J Psykoaktiva droger. 2012, 44: 38 – 55. 10.1080/02791072.2012.662112
- Carnes PJ, Green BA, Merlo LJ, Polles A, Carnes S, Gold MS: PATHOS: en kort screeningapplikation för bedömning av sexuellt beroende. J Addict Med. 2012, 6: 29 – 34. 10.1097/ADM.0b013e3182251a28
- Hanson RK: Förutspår Static-99 recidivism bland äldre sexuella gärningsmän?. Sexmissbruk. 2006, 18: 343–355. 10.1007 / s11194-006-9027-y
- Weinstein A, Katz L, Eberhardt H, Cohen K, Lejoyeux M: Sexuell tvång - Förhållande till kön, bilaga och sexuell läggning. J Behav Addict. 2015, 4: 22 – 26. 10.1556 / JBA.4.2015.1.6
- Chung WS, Lim SM, Yoo JH, Yoon H: Könsskillnad i hjärnaktivering till audiovisuell sexuell stimulering; upplever kvinnor och män samma nivå av upphetsning som svar på samma videoklipp?. Int J Impot Res. 2013, 25: 138-142. 10.1038 / ijir.2012.47
- Rupp HA, Wallen K: Könsskillnader som svar på visuella sexuella stimuli: en recension. Arch Sex Behav. 2008, 37: 206 – 218. 10.1007/s10508-007-9217-9
- Hanson RK, Harris AJ, Helmus L, Thornton D: Könsbrottsliga med hög risk kanske inte är högrisk för alltid. J Interpers våld. 2014, 29: 2792-813. 10.1177/0886260514526062
- Reid RC, Cyders MA, Moghaddam JF Fong TW: Psykometriska egenskaper hos Barratt Impulsivity Scale hos patienter med spelstörningar, hypersexualitet och metamfetaminberoende. Addict Behav. 2014, 39: 1640 – 1645. 10.1016 / j.addbeh.2013.11.008
- Reid RC, Carpenter BN, Hook JN, Garos S, Manning JC, Gilliland R, Cooper EB, McKittrick H, Davtian M, Fong T: Rapport om resultat i en DSM-5-fältstudie för hypersexuell störning. J Sex Med. 2012, 9: 2868 – 2877. 10.1111 / j.1743-6109.2012.02936.x
- Garcia FD, Thibaut F: Sexuella missbruk. Am J Drug Alkoholmissbruk. 2010, 36: 254-260. 10.3109/00952990.2010.503823
- Reid RC, Bramen JE, Anderson A, Cohen MS: Mindfulness, emotionell dysregulation, impulsivitet och stressbenägenhet hos hypersexuella patienter. J Clin Psychol. 2014, 70: 313 – 321. 10.1002 / jclp.22027
- Hållare MK, Hadjimarkou MM, Zup SL, Blutstein T, Benham RS, McCarthy MM, Mong JA: Metamfetamin underlättar kvinnligt sexuellt beteende och förbättrar neuronal aktivering i den mediala amygdala och ventromediala kärnan i hypotalamus. Psychon. 2010, 35: 197 – 208. 10.1016 / j.psyneuen.2009.06.005
- Spauwen LW, Niekamp AM, Hoebe CJ, Dukers-Muijrers NH: Droganvändning, sexuellt riskbeteende och sexuellt överförda infektioner bland swingers: en tvärsnittsstudie i Nederländerna. Sex Transm Infect. 2015, 91: 31 – 36. 10.1136 / sextrans-2014-051626
- Castelo-Branco C, Parera N, Mendoza N, Pérez-Campos E, Lete I, CEA-gruppen: Alkohol- och drogmissbruk och riskabelt sexuellt beteende hos unga vuxna kvinnor. Gynekol Endokrinol. 2014, 30: 581 – 586. 10.3109/09513590.2014.910190
- Jia ZJ, Yan SY, Bao YP, Lian Z, Zhang HR, Liu ZM: Sexuella beteendeskillnader mellan användare av stimulanter av amfetamin-typ och heroinanvändare. J Addict Med. 2013, 7: 422 – 427. 10.1097/ADM.0b013e3182a952b2
- Bruijnzeel AW: kappa-opioidreceptorsignalering och hjärnbelöningsfunktion. Brain Res Rev. 2009, 62: 127 – 146. 10.1016 / j.brainresrev.2009.09.008
- Orsini CA, Ginton G, Shimp KG, Avena NM, Gold MS, Setlow B: Livsmedelsförbrukning och viktökning efter upphörande av kronisk amfetaminadministration. Aptit. 2014, 78: 76 – 80. 10.1016 / j.appet.2014.03.013
- Ibrahim C, Reynaert C: Hypersexualitet vid neurokognitiva störningar hos äldre människor - en omfattande genomgång av litteraturen och fallstudien. Psykiater Danub. 2014, 26: 36 – 40.
- Vallejo-Medina P, Sierra JC: Effekten av narkotikamissbruk och abstinens påverkan på sexuell funktion i ett spanskt drogberoende prov för man: en flersidig studie. J Sex Med. 2013, 10: 333 – 341. 10.1111 / j.1743-6109.2012.02977.x
- Baskerville TA, Douglas AJ: Dopamin- och oxytocininteraktioner som ligger bakom beteenden: potentiella bidrag till beteendestörningar. CNS Neurosci Ther. 2010, 16: e92-123. 10.1111 / j.1755-5949.2010.00154.x
- Riebe CJ, Lee TT, Hill MN, Gorzalka BBB: Utfällt uttag räknar de negativa effekterna av subkronisk cannabinoidadministration på sexuellt beteende hos råttor hos män. Neurosci Lett. 2010, 472: 171 – 174. 10.1016 / j.neulet.2010.01.079
- Korda JB, Pfaus JG, Kellner CH, Goldstein I: Persistent genital arousal disorder (PGAD): fallrapport av långvarig symptomatisk hantering med elektrokonvulsiv terapi. J Sex Med. 2009, 6: 2901 – 2909. 10.1111 / j.1743-6109.2009.01421.x
- Huang CJ, Liao HT, Yeh GC, Hung KL: Distribution av HLA-DQB1-alleler hos patienter med Kleine-Levin-syndrom. J Clin Neurosci. 2012, 19: 628 – 630. 10.1016 / j.jocn.2011.08.020
- Blum K, Noble EP, Sheridan PJ, Montgomery A, Ritchie T, Jagadeeswaran P, Nogami H, Briggs AH, Cohn JB: Allelisk förening av human dopamin D2-receptorgen i alkoholism. JAMA. 1990, 263: 2055 – 2060. 10.1001 / jama.263.15.2055
- Blum K, Sheridan PJ, Wood RC, Braverman ER, Chen TJ, Cull JG, Comings DE: D2-dopaminreceptorgenen som en avgörande faktor för belöningsbristsyndrom. JR Soc Med. 1996, 89: 396-400. 10.1177/014107689608900711
- Miller WB, Pasta DJ, MacMurray J, Chiu C, Wu H, Comings DE: Dopaminreceptorgener förknippas med ålder vid första samlag. J Biosoc Sci. 1999, 31: 43 – 54. 10.1017 / S0021932099000437
- Hamann S, Herman RA, Nolan CL, Wallen K: Män och kvinnor skiljer sig åt i amygdala-svaret på visuella sexuella stimuli. Nat Neurosci. 2004, 7: 411 – 416. 10.1038 / nn1208
- Quiñones-Jenab V: Varför är kvinnor från Venus och män från Mars när de missbrukar kokain?. Brain Res. 2006, 1126: 200 – 203. 10.1016 / j.brainres.2006.08.109
- Noble EP, Blum K, Khalsa ME, Ritchie T, Montgomery, A Wood RC, Fitch RJ, Ozkaragoz T, Sheridan PJ, Anglin MD, Paredes A, Treiman LJ, Sparkes RS: Allelisk associering av D2-dopaminreceptorgenen med kokainberoende. Drogalkohol beror. 1993, 33: 271 – 285. 10.1016/0376-8716(93)90113-5
- Harrison PJ, Tunbridge EM: Catechol-O-metyltransferas (COMT): en gen som bidrar till könsskillnader i hjärnfunktion och till sexuell dimorfism i predisposition för psykiatriska störningar. Neuropsychopharmacology. 2008, 33: 3037 – 3045. 10.1038 / sj.npp.1301543
- Guo G, Tong Y: Ålder vid första samlag, gener och social kontext: bevis från tvillingar och Dopamin D4-receptorn. Demografi. 2006, 43: 747 – 769. 10.1353 / dem.2006.0029
- Bradley KC, Boulware MB, Jiang H, Doerge RW, Meisel RL, Mermelstein PG: Förändringar i genuttryck i nucleus accumbens och striatum efter sexuell upplevelse. Gener Brain Behav. 2005, 4: 31 – 44. 10.1111 / j.1601-183X.2004.00093.x
- Rao BS, Raju TR, Meti BL: Ökad numerisk densitet av synapser i CA3-regionen i hippocampus och molekylskikt av motorisk cortex efter självstimulering som givande erfarenhet. Neuroscience. 1999, 91: 799 – 803. 10.1016/S0306-4522(99)00083-4
- Heshmati M: Kokaininducerad LTP i det ventrale tegmentalområdet: ny insikt i mekanism och tidsförlopp belyser beroendets cellulära underlag. J Neurophysiol. 2009, 101: 2735 – 2737. 10.1152 / jn.00127.2009
- Beaver KM, Wright JP, Walsh A: En genbaserad evolutionär förklaring för sambandet mellan kriminellt engagemang och antal sexpartners. Biodemografi Soc Biol. 2008, 54: 47 – 55. 10.1080/19485565.2008.9989131
- Santtila P, Jern P, Westberg L, Walum H, Pedersen CT, Eriksson E, Sandnabba N: Dopamintransportörgenen (DAT1) polymorfism är associerad med för tidig utlösning. J Sex Med. 2010, 7: 1538 – 1546. 10.1111 / j.1743-6109.2009.01696.x
- Chen TJH, Blum K, Mathews D, Fisher L, Schnautz N, Braverman Er, Schoolfield J, Downs W, Blum SH, Mengucci J, Meshkin B, Arcuri V, Bajaj A, Waite RL, Comings DE: Preliminär associering av både Dopamine D2-receptorn (DRD2) [Taq1 A1 Allele] och Dopamine Transporter (DAT1) [480 bp Allele] -gener med patologiskt aggressivt beteende, en klinisk subtyp av belöningsunderskottssyndrom (RDS) hos ungdomar. Gene Ther Mol Biol. 2007, 1: 93-112. Åtkomst: juli 23, 2015: http://gtmb.org/pages/Vol11A/HTML/11._Chen_et_al,_93-102.htm.
- Burt SA, Mikolajewski AJ: Preliminära bevis för att specifika kandidatgener är associerade med antisocialt beteende hos ungdomar. Aggress Behav. 2008, 34: 437 – 445. 10.1002 / ab.20251
- Jóźków P, Słowińska-Lisowska M, Łaczmański Ł, Mędraś M: Polymorfa varianter av neurotransmittorreceptorgener kan påverka sexuell funktion hos åldrande män: data från HALS-studien. Neuroendokrinologi. 2013, 98: 51 – 59. 10.1159/000350324
- Försäljning JM, DiClemente RJ, Brody GH, Philibert RA, Rose E: Interaktion mellan 5-HTTLPR-polymorfism och missbrukshistoria på tonåriga afroamerikanska kvinnors kondomanvändningsbeteende efter deltagande i en HIV-förebyggande intervention. Föregående Sci. 2014, 15: 257 – 267. 10.1007/s11121-013-0378-6
- Barrett CE, Keebaugh AC, Ahern TH, Bass CE, Terwilliger EF, Young LJ: Variation i vasopressinreceptor (Avpr1a) uttryck skapar mångfald i beteenden relaterade till monogami i prärievolum. Horm Behav. 2013, 63: 518 – 526. 10.1016 / j.yhbeh.2013.01.005
- Garcia JR, MacKillop J, Aller EL, Merriwether AM, Wilson DS, Lum JK: Föreningar mellan dopamin D4-receptorgenvariation med både otrohet och sexuell promiskuitet. PLoS One. 2010, 5: e14162. 10.1371 / journal.pone.0014162
- Daw J, Guo G: Påverkan av tre gener på hur ungdomar använder preventivmedel, USA 1994-2002. Popul Stud (Camb). 2011, 65: 253 – 271. 10.1080/00324728.2011.598942
- Emanuele E, Brondino N, Pesenti S, Re S, Geroldi D: Genetisk belastning på mänskliga kärleksfulla stilar. Neuro Endocrinol Lett. 2007, 28: 815 – 821.
- Matsuda KI: Epigenetiska förändringar i östrogenreceptorn a genpromotor: implikationer i sociosexuella beteenden. Främre Neurosci. 2014, 8: 344. 10.3389 / fnins.2014.00344
- Rice WR, Friberg U, Gavrilets S: Homosexualitet via kanaliserad sexuell utveckling: ett testprotokoll för en ny epigenetisk modell. BioEssays. 2013, 35: 764 – 770. 10.1002 / bies.201300033
- Gundersen B: Parbindning genom epigenetik. Nat Neurosci. 2013, 16: 779. 10.1038 / nn0713-779
- Wang H, Duclot F, Liu Y, Wang Z, Kabbaj M: Histondeacetylasinhibitorer underlättar bildning av partnerpreferenser i prärievolym hos kvinnor. Nat Neurosci. 2013, 16: 919 – 924. 10.1038 / nn.3420
- Zeh JA, Zeh DW: Moderns arv, epigenetik och utvecklingen av polyandri. Genetica. 2008, 134: 45 – 54. 10.1007 / s10709-007-9192-z
- Addad M, Lesiau A: Extraversion, neurotism, omoralisk bedömning och kriminellt beteende. Med Law. 1989, 8: 611 – 622.
- Brodie R: Virus of the Mind: The New Science of the Meme. Hay House, Inc, New York, NY; 1996, pp 66.
- Hernnstein R, Murray C: Klockkurvan: intelligens och klassstruktur i det amerikanska livet. The Free Press, New York, NY; 1994.
- Smillie LD, Cooper AJ, Proitsi P, Powell JF, Pickering AD: Variation i DRD2 dopamingen förutspår extraverted personlighet. Neurosci Lett. 2010, 468: 234 – 327. 10.1016 / j.neulet.2009.10.095
- Kommer DE: Genbomben. Snabbar högre utbildning och avancerad teknologi urvalet av gener för inlärningssvårigheter, ADHD, beroendeframkallande och störande beteenden?. Hope Press, Duarte CA; 1996.
- Kommer DE: Problemet beteende syndrom. Genbomben. Accelererar högre utbildning och avancerad teknik urvalet av gener för inlärningssvårigheter, ADHD, beroendeframkallande och störande beteenden ?. Hope Press, Duarte CA; 1996. pp 91-94.
- Kommer DE: Genval. Genbomben. Accelererar högre utbildning och avancerad teknik urvalet av gener för inlärningssvårigheter, ADHD, beroendeframkallande och störande beteenden ?. Hope Press, Duarte CA; 1996. pp 89-90.
- Kommer DE: Berkeley-studien. Genbomben. Accelererar högre utbildning och avancerad teknik urvalet av gener för inlärningssvårigheter, ADHD, beroendeframkallande och störande beteenden ?. Hope Press, Duarte Kalifornien; 1996. sid 105 – 210.
- Borsten J: Malibu Beach Recovery Diet Cookbook. Vidov Publishing Inc, Malibu, CA; 2015.
- Derbyshire KL, Grant JE: Tvångsmässigt sexuellt beteende: En genomgång av litteraturen. J Behav Addict. 2015, 4: 37 – 43. 10.1556/2006.4.2015.003
- Nirenberg MJ: Dopamin-abstinenssyndrom: konsekvenser för patientvård. Drugs Aging. 2013, 30: 587 – 592. 10.1007 / s40266-013-0090-z
- Grant JE, Brewer JA, Potenza MN: Neurobiologi för substans- och beteendemissbruk. CNS Spectr. 2006, 11: 924 – 930.
- Sakata K, Duke SM: Brist på BDNF-uttryck genom promotor IV stör uttryck av monoamingener i frontal cortex och hippocampus. Neuroscience. 2014, 260: 265 – 75. 10.1016 / j.neuroscience.2013.12.013
- Blum K, Liu Y, Wang W, Wang Y, Zhang Y, Oscar-Berman M, Smolen A, Febo M, Han D, Simpatico T, Cronjé FJ, Demetrovics Z, Gold MS: rsfMRI-effekter av KB220Z ™ på neurala vägar i belöningskretsar av abstinenta genotypade heroinberoende. Postgrad Med. 2015, 127: 232 – 241.
- McLaughlin T, Blum K, Oscar-Berman M, Febo M, Demetrovics Z, Agan G, Fratantonio J, Gold MS: Använda Neuroadaptagen KB200z ™ för att förbättra skrämmande, klara mardrömmar hos RDS-patienter: rollen som förbättrad, hjärnbelöning, funktionell anslutning och dopaminerg homeostas. J Reward Defic Syndr. 2015, 1: 24-35. 10.17756 / jrds.2015-006
- Blum K, Thanos PK, Badgaiyan RD, Febo M, Oscar-Berman M, Fratantonio J, Demotrovics Z, Gold MS: Neurogenetics och genterapi för belöningsbrist syndrom: ska vi till det lovade landet?. Expert Opin Biol Ther. 2015, 5: 973 – 985. 10.1517/14712598.2015.1045871
- Joranby L, Pineda-Frost KY, Gold MS: Beroende av belöningssystem för mat och hjärnor. Sexuell beroende och kompulsivitet. 2005, 12: 201–217. 10.1080/10720160500203765
- Edge PJ, Gold MS: Läkemedelsabstinens och hyperfagi: lektioner från tobak och andra droger. Curr Pharm Des. 2011, 17: 1173 – 1179. 10.2174/138161211795656738
- Moholy M, Prause N, Proudfit, GH, Rahman A, Fong T: Sexuell lust, inte hypersexualitet, förutsäger självreglering av sexuell upphetsning. Cogn Emot. 2015, 6: 1012.
- Prause N, Steele, VR, Staley C, Sabatinelli, D, Hajcak G: Modulation av sena positiva potentialer genom sexuella bilder hos problemanvändare och kontroller som inte överensstämmer med "pornoberoende". Biol Psychol. 2015, 109: 192-199. 10.1016 / j.biopsycho.2015.06.005
- Mechelmans DJ, Irvine M, Banca P, et al: Förbättrad uppmärksamhetsförskjutning mot sexuellt uttryckliga ledtrådar hos individer med och utan tvångsmässigt sexuella beteenden. PLoS One. 2014, 25, 9 (8): e105476. 10.1371 / journal.pone.0105476
- Blum K, Gardner E, Oscar-Berman M, Guld M: "Gilla" och "vilja" kopplat till belöningsbristsyndrom (RDS): hypotes om differentiell respons i hjärnbelöningskretsar. Curr Pharm Des. 2012, 18 (1): 113-118.
- Hyman SE: Diagnos av DSM: Diagnostisk klassificering behöver grundläggande reformer. Stora hjärnan. 2011, 2011: 6. Åtkomst: 2011 apr 26: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3574782/.
- Casey BJ, Craddock N, Cuthbert BN, Hyman SE, Lee FS, Ressler KJ: DSM-5 och RDoC: framsteg inom psykiatrisk forskning?. Nat Rev Neurosci. 2013, 14: 810-14. 10.1038 / nrn3621
- Karila L, Wéry A, Weinstein A, Cottencin O, Petit A, Reynaud M, Billieux J: Sexuellt beroende eller hypersexuell störning: olika termer för samma problem? En genomgång av litteraturen. Curr Pharm Des. 2014, 20: 4012-20. 10.2174/13816128113199990619
- Carvalho J, Štulhofer A, Vieira AL, Jurin T: Hypersexualitet och hög sexuell lust: utforska strukturen för problematisk sexualitet. J Sex Med. 2015, 12: 1356-67. Åtkomst: 2015 Mar 23: http://onlinelibrary.wiley.com/doi/10.1111/jsm.12865/abstract;jsessionid=6F416CCBB66B7F0EA28E428D4993EBD5.f01t04. 10.1111 / jsm.12865
- Rettenberger M, Klein V, Briken P: Förhållandet mellan hypersexuellt beteende, sexuell upphetsning, sexuell hämning och personlighetsdrag. Arch Sex Behav. 2015, Jan 6: (Epub före tryck). Åtkomst: 2015 Jan 6: 10.1007/s10508-014-0399-7
- Walters GD, Knight RA, Långström N: Är hypersexualitet dimensionell? Bevis för DSM-5 från allmän population och kliniska prover. Arch Sex Behav. 2011, 40: 1309-21. Åtkomst: 2011 feb 3: 10.1007/s10508-010-9719-8

{kind=link}