KOMMENTARER: En av få studier som täcker hur avhållsamhet påverkar halterna av dopaminreceptorer i primater.
- D2-receptorer studsar tillbaka ganska snabbt - mindre än en månad
- D1-receptorer är alldeles för höga på en månad, men studsar tillbaka inom 90 dagar.
- Höga eller låga D1-receptorer kan vara nycklar till akut abstinens och sug
Thomas JR Beveridge1, Hilary R Smith1, Michael A Nader1 och Linda J Porrino1
1Institutionen för fysiologi och farmakologi, Centrum för neurobiologisk undersökning av drogmissbruk, Wake Forest University School of Medicine, Winston-Salem, NC, USA
Korrespondens: Dr LJ Porrino, Institutionen för fysiologi och farmakologi, Centrum för neurobiologisk undersökning av drogmissbruk, Wake Forest University School of Medicine, Medical Center Boulevard, Winston-Salem, NC 27157-1083, USA. Tel: + 1 336 716 8575; Fax: + 1 336 716 8501; E-post: [e-postskyddad]
Mottagen 29 april 2008; Reviderad 25 juli 2008; Godkänd 30 juli 2008; Publicerad online den 3 september 2008.
Abstrakt
Även om dysreglering inom dopaminsystemet (DA) är ett kännetecken för kronisk exponering för kokain, är frågan om dessa förändringar kvarstår i abstinens i stort sett obesvarad. Icke-mänskliga primater representerar en idealisk modell för att bedöma effekterna av avhållsamhet på DA-systemet efter kronisk exponering för kokain. I den här studien har man själv administrerat kokain (0.3) mg/kg per injektion, 30-förstärkare per session) under ett fast intervall 3-min sschema för 100 dagar följt av antingen 30 eller 90 dagar avhållsamhet. Denna varaktighet av självadministrering av kokain har tidigare visats minska DA D2-liknande receptortätheter och öka nivåerna av D1-liknande receptorer och DA-transportörer (DAT). Svar av kontrollapor upprätthölls genom matpresentation under ett identiskt protokoll och samma avhållsamhetsperioder. [3H]SCH 23390 bindande till DA D1-receptorer efter 30 dagars avhållsamhet var signifikant högre i alla delar av striatum, jämfört med kontrolldjur, medan [3H]raclopridbindning till DA D2-receptorer skilde sig inte mellan grupperna. [3H]WIN 35 428-bindning till DAT var också signifikant högre i praktiskt taget alla delar av rygg- och ventralt striatum efter 30 dagars avhållsamhet. Efter 90 dagars avhållsamhet var nivåerna av DA D1-receptorer och DAT emellertid inte olika från kontrollvärden. Även om dessa resultat indikerar att det finns en eventuell återhämtning av de separata elementen i DA-systemet, belyser de också dessa komponenters dynamiska karaktär under de initiala faserna för avhållsamhet från kronisk kokain självadministrering.
Nyckelord:
kokain, dopamin, autoradiografi, avhållsamhet, striatum
INLEDNING
Kronisk kokainanvändning bland mänskliga missbrukare har associerats med neuroadaptationer i dopamin (DA) -systemet (Malison et al,, 1998; Volkow et al,, 1993, 1997). Dessa inkluderar ökning av tätheten hos DA-transportörer (DAT) och minskningar i koncentrationerna av DA D2-liknande receptorer (Liten et al,, 1999; Mash et al,, 2002; Volkow et al,, 1993). Dessutom har förändringar i frisläppande av DA också observerats. Till exempel undersökare som använder positronemissionstomografi (PET) -studier med [11C]racloprid och metylfenidat har visat minskningar i DA-frisättning i striatumet för kroniska kokainanvändare (Volkow et al,, 1997; Wong et al,, 2006). Ett problem är emellertid att det ofta är svårt att utesluta påverkan av andra faktorer som användningen av andra olagliga och lagliga droger, skillnader i tidigare drogintag och användningsmönster och skillnader i livsstil. Dessa skillnader, liksom förekomsten av tillstånd som kan föregripa läkemedelsanvändning, kan begränsa tolkningen av studier på mänskliga patienter.
Icke-mänskliga primatmodeller, i vilka variabler systematiskt kan manipuleras, representerar en alternativ metod för studien av konsekvenserna av kronisk kokain självadministrering och efterföljande avhållsamhet. Tidigare studier har visat att kronisk exponering för kokain åtföljs av signifikanta minskningar i koncentrationerna av DA D2-receptorer såväl som förhöjningar i nivåerna av D1-receptorer och densiteten för DAT (Letchworth et al,, 2001; Moore et al,, 1998a, 1998b; Nader et al,, 2002, 2006). Dessa effekter speglar de som ses hos människor, vilket underbygger användbarheten för dessa modeller av läkemedelseksponering.
Även om det finns betydande bevis för dysreglering av DA-system har det visat sig vara svårare att utvärdera om det finns några bevis för återhämtning efter upphörande av droganvändning (Malison et al,, 1998; Jacobsen et al,, 2000; Volkow et al,, 1993) eller om dessa förändringar kvarstår utanför tidsramen för fortsatt exponering för kokain. Återigen kan icke-mänskliga primatmodeller ge insikt i denna faser av beroende. Farfel et al, (1992) rapporterade minskade koncentrationer av DAT- och D1-liknande receptorer i striatumet av apor efter avhållsamhet från kronisk icke-kontingent exponering för kokain. Den specifika rollen för avhållsamhet var emellertid svår att fastställa på grund av bristen på mätningar i en grupp utan avhållsamhetsperiod. På liknande sätt Melega et al, (2008) rapporterade signifikant minskade nivåer av DAT i striatumet av vervet apor efter 3 veckor av avhållsamhet från en eskalerande metamfetaminregim. Administrering av stimulantia i båda studierna var emellertid icke kontingent. Administrationsvägen (kontingent vs noncontingent) har visat sig påverka hjärnan differentiellt med avseende på både DA-frisläppande (Hemby et al,, 1997) och glukosmetabolism (Graham och Porrino, 1995; Porrino et al,, 2002). Således kringgår användningen av självadministrering i den nuvarande studien denna fråga. Dessutom har effekterna av långvarig självadministrering av kokain på hjärn-DA-system studerats i stor utsträckning med användning av denna modell för självadministrering i rhesus-apor, och därigenom tillhandahåller en baslinje för att bedöma neuroadaptationerna som uppstår under avhållsamhet.
Syftet med dessa studier var därför att bestämma om förändringarna i DAT och DA D1 och D2-receptorkoncentrationer som tidigare har visats hos djur exponerade för självadministrering av kokain (Letchworth et al,, 2001; Moore et al,, 1998a, 1998b; Nader et al,, 2002) skulle vändas efter utökade perioder med avhållsamhet. På grundval av studier på mänskliga läkemedelsanvändare (jfr Volkow et al,, 1993), vi ansåg att dessa förändringar i DA-systemet skulle fortsätta även efter 3 månader av avhållsamhet. För detta ändamål apar självadministrerad kokain för 100-sessioner, med totala intag av 900 mg/kg, följt av antingen 30 eller 90 dagar avhållsamhet från läkemedlet. DA D1- och D2-receptorer, liksom DAT, mättes med kvantitativt vitro receptor autoradiografi.
METODER
Ämnen
Totalt 17 experimentellt naiva rhesus-apor för vuxna (Macaca mulatta) som väger mellan 7.7 och 13 kg (medelvärde ± SD, 10.2 ± 1.32) i början av studien fungerade som försökspersoner. Alla procedurer utfördes i enlighet med fastställd praxis som beskrivs i National Institute of Health Guide for care and Use of Laboratory Animals. Dessutom granskades och godkändes alla förfaranden av Animal Care and Use Committee vid Wake Forest University. Apor hölls individuellt i burar i rostfritt stål med vatten AD libitum; djur hade fysisk och visuell kontakt med varandra. Deras kroppsvikt hölls på ungefär 90 – 95% av fritt matande vikter av banan-smaksatta pellets som tjänades under försökssessionerna och genom kompletterande utfodring av Lab Diet Monkey Chow, tillhandahölls inte förr än 30 min efter sessionen. Dessutom fick de färsk frukt eller jordnötter minst tre gånger per vecka. Varje apa vägdes en gång i veckan, och vid behov justerades deras dieter för att upprätthålla stabila vikter.
Beteendeapparat
Experimentella sessioner genomfördes i ventilerade och ljuddämpade operakammare (1.5 × 0.74 × 0.76 m; Med Associates Inc., East Fairfield, VT) utformad för att rymma en primatstol (Model R001; Primate Products, Redwood City, CA). Kammaren innehöll en underrättelsespanel (48 × 69 cm), som bestod av två utdragbara hävarmar (5 cm bred) och tre stimulansljus. Spakarna placerades inom räckhåll för apan som satt i primitstolen. Banan-smaksatt matpellets (1 g; Bio-Serv, Frenchtown, NJ) levererades från en matare belägen ovanpå kammaren. En peristaltisk infusionspump (7531-10; Cole-Parmer Co., Chicago, IL) användes för att leverera läkemedelsinjektioner med en hastighet av ungefär 1 ml per 10 s till dessa djur som själv administrerar kokain. Driften av kamrarna och datainsamlingen genomfördes med ett Power Macintosh-datorsystem med ett gränssnitt (Med Associates Inc.).
Kirurgiska ingrepp
Alla apor, inklusive kontroller, bereddes kirurgiskt under sterila förhållanden med inneboende intravenösa katetrar och vaskulära åtkomstportar (Model GPV; Access Technologies, Skokie, IL). Apor bedövades med en kombination av ketamin (15 mg/kg, im) och butorfanol (0.03 mg/kg, im) och ett snitt gjordes nära lårbenen. Efter trubbig dissektion och isolering av venen infördes den proximala änden av katetern i venen under ett avstånd beräknat att avsluta i den underordnade vena cava. Den distala änden av katetern gängades subkutant till ett snitt som gjordes något från ryggens mittlinje. Den vaskulära åtkomstporten placerades i en ficka som bildades genom trubbig dissektion nära detta snitt. Apor fick 24 – 48 h återhämtningstid innan han återvänder till matförstärkt svar. Ungefär 5 dagar före terminalförfarandet implanterades varje apa med en kronisk inbyggnadskateter i den intilliggande femoral artären för insamling av tidsbestämda arteriella blodprover. De kirurgiska ingrepparna var identiska med de som beskrivits för venekatetrarna. Dagen för den sista sessionen genomfördes en terminal cerebral glukosmetabolismundersökning där apor injicerades med 2-[14C]deoxyglukos (2-DG) ungefär 2 min efter sessionens slut och blodprov erhölls genom arteriell kateter över en 45 min period (se Beveridge et al,, 2006 för detaljer). Metabolismdata från dessa studier presenteras inte här.
Självadministrationsförfaranden
Apor utbildades ursprungligen för att svara på en av två spakar genom att förstärka varje svar på rätt spak med en matpellet. Under ungefär en 3-veckors period ökades intervallet mellan tillgängligheten av matpellets gradvis tills ett 3-min-intervall uppnåddes (dvs. fixerat intervall 3-min-armeringsschema; FI 3-min). Under de slutliga schemaläggningsförhållandena, det första svaret på spaken efter 3 min resulterade i leveransen av en matpellets; sessioner avslutades efter 30 matpresentationer. I slutet av varje session drogs svarsspakarna in, husbelysningen och stimulansljusen släcktes och djur förblev i den mörka kammaren under ungefär 30 min innan de återvände till sina hemburar. Alla apor svarade under FI 3-min schema för matpresentation under minst 20 sessioner och tills stabila prestanda uppnåddes (± 20% av medelvärdet under tre på varandra följande sessioner, utan trender i svarsfrekvensen). När svaret med livsmedlet upprätthöll var stabilt kopplades mataren ut och effekterna av utrotning på att svara undersöktes under fem sammanhängande sessioner, varefter svaret återupprättades och upprätthölls genom matpresentation.
Efter att baslinprestanda hade fastställts, bereddes alla apor kirurgiskt med venekatetrar, såsom beskrivits ovan, och tilldelades slumpmässigt till en av tre grupper. En grupp apor fungerade som kontroller och fortsatte att svara under FI 3-min schema för matpresentation i totalt 100 sessioner (N=6). De återstående 11-aporna tilldelades kokainens självadministreringsgrupper (0.3 mg/kg per injektion). Eftersom 0.3 mg/kg kokain per injektion ansågs vara en hög dos för tidigare kokainnaiva apor, för de flesta djur uppnåddes denna dos inom två sessioner genom att först låta apan själv administrera 0.1 mg/kg kokain. Livsmedelsbibehållen prestanda tilläts stabiliseras efter operationen (ungefär 4 – 6 dagar) innan kokainens självadministrationssessioner påbörjades. Före varje försökssession rensades djurets baksida med 95% etanol- och betadinskrubba och en 22-mätare Huber Point Needle (modell PG20-125) infördes i porten som ledde till den venösa katetern och anslöt en infusionspump innehållande kokainlösningen till katetern. Innan sessionen började kördes pumpen i ungefär 3 s, fylla hamnen med den dos kokain som var tillgänglig under försökssessionen. Sessioner slutade efter 30-injektioner; som under kontrollförhållanden förblev apor i den mörka kammaren under ungefär 30 min. I slutet av varje session fylldes porten med hepariniserad saltlösning (100 U/ml) för att förhindra koagulering.
Experimentella sessioner genomfördes vid ungefär samma tid varje dag och fortsatte under totalt 100 sessioner. Efter avslutandet av 100-sessionerna infördes en avhållsamhetsperiod på 30 eller 90 dagar under vilken tid katetrar spolades dagligen med hepariniserad saltlösning, men inga kokain- eller livsmedelsadministrationssessioner genomfördes. För kontrollgruppen infördes abstinensperioder på 30 dagar i fyra djur och 90 dagar i de två andra. För kokaingruppen belastades åtta djur med avhållsamhetsperioder på 30 dagar och 90 dagar på tre djur. I slutet av abstinensperioden genomfördes en slutlig självadministrationssession (livsmedelskontroll eller kokain) och 2-DG-förfarandet inleddes omedelbart efter sessionen. I två kontroller och fyra självadministreringsdjur av kokain i 30-dagars avhållsamhetsgrupp mottogs ingen kokain vid den sista sessionen. Djur dödades mänskligt med en överdos av pentobarbital (100) mg/kg, iv) i slutet av 45 min spårupptagningsperiod.
Vävnadsbehandling
Efter avlivningen avlägsnades hjärnan omedelbart, blockerades och frystes i isopentan vid −35 till −55 ° C och lagrades sedan vid −80 ° C. Vävnadsblocken innehållande striatum skars sedan i en kryostat vid -20 ° C i koronalplanet till 20 μm-sektioner, samlade på elektrostatiskt laddade objektglas, torkade under vakuum över natt vid 4 ° C, lagras sedan vid −80 ° C tills de behandlats för autoradiografi. Hjärnsektioner samlades upp från delarna av caudatkärnan, putamen och nucleus accumbens som ligger rostral till den främre kommissuren. Denna region kallas det förkommissurala striatum. Vidare betecknades rostrala och caudala nivåer av det förkommissurala striatum med hänvisning till nucleus accumbens. Det rostrala förkommissurala striatumet är det område där nucleus accumbens inte är differentierade till distinkta skal- och kärnunderdelar Det caudala förkommissurala striatumet är det område som överensstämmer med utseendet på skalet och kärnan i nucleus accumbens, vilket är bakom uppkomsten av luktknölen. För var och en av bindningsstudierna togs två angränsande sektioner vid var och en av fem nivåer (två rostral och tre caudal) genom det föregångsliga striatum för totalt 10-sektioner per djur.
D1 Receptor Binding
DA D1-receptorbindningsställets densitet bestämdes med [3H]SCH 23390 (specifik aktivitet — 85 Ci/mmol; PerkinElmer, Boston, MA) genom kvantitativ vitro receptor autoradiografi enligt förfaranden anpassade från Lidow et al, (1991) och Nader et al, (2002). Sektioner förinkuberades för 20 min i buffert (50 mM Tris, 120 mM NaCl, 5 mM KCl, 2 mM CaCl2, 1 mM MgCl2, pH 7.4, 25 ° C) för att avlägsna endogent DA, kokain och [14C] från 2-DG-proceduren. Avsnitt inkuberades sedan för 30 min i samma buffert, pH 7.4, 25 ° C, innehållande 1 mM askorbinsyra, 40 nM ketanserin och 1 nM [3H]SCH 23390. Efter inkubation sköljdes sektioner två gånger för 20 s i buffert innehållande 1 mM askorbinsyra vid pH 7.4, 4 ° C, doppades sedan i destillerat vatten vid 4 ° C och torkades under en ström av sval luft. Ospecifik bindning definierades genom inkubering av intilliggande sektioner i inkubationslösningen i närvaro av 5 μM (+) -Butaclamol. Avsnitt, tillsammans med kalibrerade [3H] autoradiografiska standarder (Amersham, Piscataway, NJ) överlämnades till Kodak Biomax MR-film (Fisher Scientific, Pittsburgh, PA) under 6 veckor.
D2 Receptor Binding
Densiteten och fördelningen av DA D2-receptorbindningsställen bestämdes med [3H]racloprid (specifik aktivitet, 87 Ci/mmol; PerkinElmer) enligt förfaranden anpassade från Lidow et al, (1991) och Nader et al, (2002). Sektioner förinkuberades för 20 min i buffert (50 mM Tris, 120 mM NaCl, 5 mM KCl, pH 7.4, 25 ° C) för att avlägsna endogent DA, kokain och [14C] från 2-DG-proceduren. Objektglas inkuberades sedan för 30 min i samma buffert, innehållande 5 mM askorbinsyra och 2 nM [3H]rakloprid. Avsnitt sköljdes 3 × 2 min i buffert vid pH 7.4, 4 ° C, doppades sedan i destillerat vatten vid 4 ° C och torkades under en ström av sval luft. Ospecifik bindning definierades genom inkubering av intilliggande sektioner i inkubationslösningen i närvaro av 1 μM (+) -Butaclamol. Avsnitt, tillsammans med kalibrerade [3H] autoradiografiska standarder applicerades på Kodak Biomax MR-film under 8 veckor.
Dopamin Transporter Binding
Densiteten för DAT-bindningsställen bestämdes med användning av [3H]VINNA 35,428 (specifik aktivitet, 87 Ci/mmol; PerkinElmer) autoradiografi enligt förfaranden anpassade från Canfield et al, (1990) och Letchworth et al, (2001). Vävnadssektioner förinkuberades i buffert (50 mM Tris, 100 mM NaCl, pH 7.4, 4 ° C) för 20 min för att ta bort eventuella rester av DA, kokain och [14C] från 2-DG-proceduren. Avsnitt inkuberades sedan för 2 h vid 4 ° C i samma buffert innehållande 5 nM [3H]VINNA 35 428. Avsnitt sköljdes under totalt 2 min i buffert vid 4 ° C, doppades därefter i destillerat vatten vid 4 ° C och torkades under en ström av kall luft. Ospecifik bindning definierades genom inkubering av intilliggande sektioner i inkubationslösningen i närvaro av 30 μM kokain. Avsnitt, tillsammans med kalibrerade [3H] autoradiografiska standarder applicerades på Kodak Biomax MR-film under 6 veckor.
Densitometri och dataanalys
Filmer utvecklades med Kodak GBX-utvecklare, stoppbad och Rapid Fixer (VWR, West Chester, PA) och sköljdes sedan. Analys av autoradiogram utfördes med kvantitativ densitometri med ett datoriserat bildbehandlingssystem (MCID, Imaging Research; InterFocus Imaging Ltd, Cambridge, UK). Optiska densitetsvärden omvandlades till fmol/mg (av våtviktsvävnad) med referens till den kalibrerade [3H] standarder. Specifik bindning bestämdes genom att digitala subtrahera bilder av icke-specifik bindning från överlagrade intilliggande bilder av total bindning. Strukturer identifierades genom Nissl-färgning av sektioner intill de som analyserades för receptorbindning. Data från varje analys analyserades oberoende med hjälp av en enkelriktad variansanalys följt av skillnader i minst kvadrat post hoc tester för flera jämförelser. Varje region bestod av en separat analys. Eftersom bindningsdata erhållna från kontrolldjur som avhåller sig under 30 och 90 dagar inte skilde sig signifikant från varandra, liknande tidigare studier (Nader et al,, 2002) kombinerades data från kontrollgrupperna. Dessutom fanns det inga signifikanta skillnader mellan data från de djur som fick kokain och de som inte gjorde vid deras sista session, så data från dessa grupper kombinerades också.
RESULTAT
Effekter av avhållsamhet från kronisk kokain självadministrering under 30 dagar
Koncentrationer av [3H]SCH 23390-bindning till DA D1-receptorer i det förekommissionella striatum visas i Tabell 1. Speciell bindning av [3H]SCH 23390 stod för mer än 90% av total bindning. I överensstämmelse med tidigare rapporter (Moore et al,, 1998a; Nader et al,, 2002), bindningen av [3H]SCH 23390 till D1-receptorer i icke-läkemedelseksponerade kontrolldjur var heterogena med märkbara skillnader i graden av bindning mellan subregioner av striatum. Märkningen var tätare i de mer rostrala och mediala delarna i hela striatum.
![Tabell 1 - Effekt av 30 eller 90 dagar för avhållsamhet efter kronisk kokain självadministrering på täthet av [lsqb] 3H [rsqb] SCH 23390 Bindning till D1-receptorer i precommissural Striatum of Rhesus Monkeys - Tyvärr kan vi inte tillhandahålla tillgänglig alternativ text för detta. Om du behöver hjälp för att komma åt den här bilden, vänligen kontakta help@nature.com eller författaren](https://yourbrainonporn.com/wp-content/uploads/2020/01/table_thumb.gif)
Efter 30 dagar med avhållsamhet från exponering av kokain kännetecknades bindning till D1-receptorer av utbredda förhöjningar över hela rostral-caudal omfattning av det föregångsliga striatum, jämfört med bindningsdensiteter hos icke-läkemedelseksponerade kontrolldjur (Tabell 1; Figur 1). I det mer rostrala striatumet var koncentrationerna signifikant högre i caudatkärnan inklusive den dorsolaterala (+27%), central (+27%), dorsomedial (+27%) och ventromedial (+23%) delar, såväl som i rygg (+17%), central (+22%) och ventral (+23%) delar av putamen, jämfört med densiteterna i icke-läkemedelseksponerade kontrolldjur. Betydande höjningar var också tydliga i nucleus accumbens (+23%) på denna nivå. På nivån av striatum där kärnan och skalet i nucleus accumbens är mest differentierade var densiteterna av D1-liknande receptorer också signifikant högre genom hela dorsolaterala (+31%), central (+29%), dorsomedial (+30%) och ventromedial (+18%) caudatkärnan, såväl som rygg (+23%), central (+29%) och ventral (+28%) putamen, jämfört med densiteterna för icke-läkemedelseksponerade kontroller. Inom det ventrala striatumet på denna nivå koncentrationen av D1 receptorbindningsställen var högre i nucleus accumbens-kärnan (+45%) och skal (+20%), såväl som i luktknölen (+26%) jämfört med densiteterna i kontrollerna.
Figur 1.
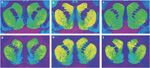 Representativa autoradiogram av [3H] SCH 23390-bindning till D1-receptorer (övre panelen) och [3H]VINNA 35428-bindning till dopamintransportörer (bottenpanelen) i koronalsektioner av rhesus monkey striatum. (a, d) Kontrolldjur som svarar på förstärkning av mat (b, e) Djur med självadministrering av kokain med 30 dagar avhållsamhet. (c, f) Kokain självadministrationsdjur med 90 dagar avhållsamhet.
Representativa autoradiogram av [3H] SCH 23390-bindning till D1-receptorer (övre panelen) och [3H]VINNA 35428-bindning till dopamintransportörer (bottenpanelen) i koronalsektioner av rhesus monkey striatum. (a, d) Kontrolldjur som svarar på förstärkning av mat (b, e) Djur med självadministrering av kokain med 30 dagar avhållsamhet. (c, f) Kokain självadministrationsdjur med 90 dagar avhållsamhet.
Koncentrationer av [3H]raclopridbindning till DA D2-receptorer i det föregångsliga striatum visas i Tabell 2. Speciell bindning med [3H]raclopride stod för mer än 90% av total bindning. Distributionen av [3H]raclopridbindning till D2-receptorer var också heterogen över subregioner av dorsalt och ventralt striatum som i tidigare rapporter (Moore et al,, 1998b; Nader et al,, 2002). I de icke-läkemedelseksponerade kontrollerna var högre koncentrationer av D2-bindningsställen närvarande i rygg jämfört med det ventrala striatum. Dessutom fanns bevis på en medial till lateral gradient med de högre koncentrationerna av bindningsställen närvarande i de mer laterala delarna av striatum.
Efter 30 dagars avhållsamhet skilde sig nivåerna av D2-receptorbindning i kokaineksponerade och matförstärkta djur inte signifikant från varandra i de flesta områden i striatum. Högre koncentrationer av bindningsställen observerades i den ventrale putamen (+10%) och anterior nucleus accumbens (+12%) i vävnad från apor som exponerats för kokain jämfört med kontroller. Inga andra signifikanta skillnader noterades.
Koncentrationer av [3H]VINN 35428-bindning till DAT i det föregångsliga striatum visas i Tabell 3. I överensstämmelse med tidigare rapporter (Letchworth et al,, 2001), bindning till DAT-ställen i icke-läkemedelseksponerade djur var högre i rygg jämfört med ventral striatum. Inom nucleus accumbens observerades högre densiteter i kärnan jämfört med skalavdelningarna. Slutligen stod icke-specifik bindning för mindre än 10% av totalen.
Efter 30 dagar med avhållsamhet från exponering för kokain var bindningen till DAT signifikant högre i majoriteten av regionerna i striatum rostral till den främre upprepningen jämfört med bindning i icke-läkemedelseksponerade kontrolldjur (Figur 1). Specifikt var koncentrationerna av DAT-bindningsställen vid rostrala nivåer signifikant högre i centrala (+22%), dorsomedial (+25%) och ventromedial (+28%) caudatkärnan och i rygg (+16%) och centrala (+23%) putamen, jämfört med livsmedelsförstärkta kontroller. Dessutom fanns det också signifikant högre bindning till DAT i anterior nucleus accumbens (+37%) i kokain - jämfört med matförstärkta apor. Inom de mer caudala delarna av det förkommissurala striatumet var tätheter av DAT-bindningsställen signifikant högre i den centrala caudatkärnan (+21%) och putamen, central (+20%) och ventral (+19%; Figur 1). Inom det ventrala striatumet på denna nivå var bindning till DAT signifikant högre i nucleus accumbens kärna (+20%) och olfaktorisk tuberkel (+24%) i vävnad från kokain- vs matförstärkta apor.
Effekter av avhållsamhet från kronisk kokain självadministrering under 90 dagar
I motsats till de utbredda skillnaderna i densiteten hos D1-receptorbindningsställen observerade hos kokain exponerade djur efter 30 dagars avhållsamhet, efter 90 dagar av avhållsamhet, fanns det inga signifikanta skillnader jämfört med livsmedelsförstärkta kontroller i någon del av det föregångsliga striatum (Tabell 1; Figur 1). På liknande sätt koncentrationer av [3H]raclopridbindning till DA D2-receptorer i det föregångsliga striatumet efter 90 dagars avhållsamhet skilde sig inte signifikant från de hos icke-läkemedelseksponerade djur (Tabell 2).
Koncentrationerna av [3H]WIN 35428-bindning till DAT uppvisade ett liknande mönster som observerats med D1- och D2-receptorer. Det fanns inga signifikanta skillnader i tätheten av DAT mellan apor som exponerats för kokain efter 90 dagars avhållsamhet jämfört med nivåer i icke-läkemedelseksponerade kontroller (Tabell 3; Figur 1) även om det bör noteras att det fanns en trend mot högre bindningsnivåer i anterior nucleus accumbens.
DISKUSSION
Tidigare studier från vår grupp har visat att kronisk exponering för självadministrering av kokain åtföljs av betydande dysreglering av DA-systemet för icke-mänskliga primater (Letchworth et al,, 2001; Moore et al,, 1998a, 1998b; Nader et al,, 2002, 2006). Resultaten från den aktuella studien visar att denna dysreglering förblir uppenbar efter upphörande av exponering för kokain. Efter 30 dagars avhållsamhet, var koncentrationerna av DA D1-receptorer och DAT signifikant förhöjda i hela striatumet av apor med historia om kronisk självkadinering av kokain jämfört med livsmedelsförstärkta kontroller. Emellertid ger den aktuella studien också tydliga bevis för återhämtning inom DA-systemet efter mer förlängda perioder med avhållsamhet (90 dagar), vilket framgår av avsaknaden av betydande skillnader mellan kokain exponerade och kontrollerar djur vid denna tidpunkt. Dessa uppgifter tyder på att exponering för kokain inte kan leda till permanenta förändringar i DA-systemet, men att återhämtning kan inträffa med långvarig avhållsamhet från narkotikamissbruk.
Dysreguleringen av DAT-koncentrationer som visas här efter avhållsamhet överensstämmer med tidigare rapporter från icke-mänskliga primater (Letchworth et al,, 2001), som visade signifikanta förhöjningar i tätheten av DAT-bindningsställen i både ventral och ryggstratum. Även om det inte uttryckligen testats, verkar det som om upphöjningen av DAT-bindningsställets täthet efter upphörande av läkemedelseksponeringen är minst lika stor i storlek och mer utbredd över striatumets regioner än de som rapporterats utan någon återkallningsperiod (Letchworth et al,, 2001). På liknande sätt är de förhöjda koncentrationerna av D1-receptorbindningsställen observerade här efter 30 dagars avhållsamhet också förenliga med tidigare studier som visade ökade D1-liknande receptorbindningsdensiteter i striatumet av icke-mänskliga primater exponerade för en identisk administrering av kokain självadministrering (Nader et al,, 2002). I motsats härtill observerades inga signifikanta skillnader mellan nivåerna av D2-liknande receptorbindningsdensiteter i striatumet för kokain exponerade och kontrolldjur. Denna brist på dysregulation var närvarande, trots de stora minskningarna i koncentrationerna av D2-receptorer som har rapporterats hos båda mänskliga missbrukare (Volkow et al,, 1993) och djurmodeller för självadministrering av kokain (Moore et al,, 1998a, 1998b; Nader et al,, 2002, 2006). Föreliggande data föreslår då en snabbare normalisering av kontrollnivåer i detta system jämfört med D1-liknande receptorer och DAT. Sammantaget visar förändringarna i DA-receptorer och DAT tydligt att perioden omedelbart efter upphörande av självadministrering av kokain är mycket labil med betydande förändringar i regleringen av DA-systemet som äger rum, men att detta följs av omreglering av systemet närmar sig en mer normal fördelning av DA-receptorer och DAT efter mer långvarig avhållsamhet.
D1 mottagarändringar
De utbredda förhöjningarna i D1-receptorer som observerats här efter upphörandet av kokainanvändning överensstämmer med rapporter som visar en ökad känslighet för D1-receptorer under uttag. Mätning av D1-känslighet genomfördes i en studie av Henry och vit (1991), i vilka enstaka enhetsregistrering av neuroner i nucleus accumbens befanns vara mer känsliga för D1-receptoragonisten SKF 38393 efter kroniska dagliga injektioner av kokain jämfört med saltbehandlade kontroller. Denna effekt var beständig, eftersom den ökade känsligheten förblev uppenbar upp till en månad från uttag. Författarna ansåg att D1-receptorsensibiliseringen berodde på D2 autoreceptors underkänslighet i det somatodendritiska A10-området och därigenom minskade det hämmande impulsflödet genom mesoaccumbens DA-system (Henry och White, 1991). Data från den aktuella studien antyder ökade D1-receptorbindningsställen i avhållsamhet, vilket kan förklara denna ökade känslighet hos dopaminerge neuroner för en direktverkande D1-receptoragonist. Dessutom var den förbättrade effekten av SKF 38393 på nucleus accumbens neuroner inte uppenbar två månader efter tillbakadragande, vilket tyder på att det var återhämtning av D1-receptorkänsligheten (Henry och White, 1991); ett resultat som överensstämmer med återhämtningen av D1-receptortätheter noterade i den aktuella studien efter 90 dagars avhållsamhet. Andra rapporter stödjer också en viktig roll för D1-receptorer vid återfall. Direktstimulering av D1-receptorer i skalet hos kärnkrabborna kan återställa kokain som söker i abstenta gnagare (Schmidt et al,, 2006). Litteraturen är emellertid något inkonsekvent genom att både D1-agonister och antagonister kan dämpa läkemedelssökande framkallade av kokainprim eller kokainrelaterade stimuli (Alleweireldt et al,, 2002; De Vries et al,, 1999; Khroyan et al,, 2000; Själv et al,, 1996; Vit et al,, 2001). Nyligen, Khroyan et al, (2003) rapporterade att D1-agonister och antagonister minskar återfallet i en icke-mänsklig primatmodell av kokain-sökande. Dessa författare föreslog att det kan finnas ett kritiskt intervall av D1-receptoraktivitet som är nödvändig för att söka kokain, och att både antagonister och agonister kan flytta aktivitet ut ur detta fönster. De ökade koncentrationerna av D1-receptorer som åtföljer avhållsamhet kan modifiera detta intervall, vilket resulterar i en förändring av känsligheten i detta system. En annan övervägande är att D1-aktivitet kan verka för att modulera aktivitet vid D2-receptorer (Nolan et al,, 2007; Ruskin et al,, 1999; Walters et al,, 1987). Föreliggande data antyder att förhållandet mellan D1 och D2-receptorer förändras under avhållsamhetsförloppet och därigenom kan förändra effektiviteten hos denna modulering.
Även om det, i motsats till nuvarande data, har rapporterats om minskade nivåer av D1-receptorer efter kronisk kokain självadministrering (Moore et al,, 1998a), det finns betydande skillnader mellan dessa studier, såsom dos och längd på exponering för kokain, totalt intag och jämförande kontrollgrupper. Sammantaget tyder därför konvergerande bevis starkt på att D1-systemet befinner sig i betydande flöde efter borttagande från kronisk kokainadministration.
Dopamine Transporter Change
Resultaten av förhöjda koncentrationer av DAT över striatumet för kokain exponerade djur efter upphörande av läkemedelsanvändning utvidgar de från våra tidigare studier som visade ökade nivåer av DAT-bindningsställen som åtföljer kokain självadministrering i icke-mänskliga primater. Föreliggande data visar att denna dysreglering kvarstår under de inledande faserna för avhållsamhet. Vidare föreslår de att återhämtning till kontrollnivåer följer en relativt lång tid (upp till 90 dagar i den aktuella studien). I våra tidigare studier visade vi att även om de ursprungligen var begränsade till i stort sett ventrala striatalregioner, förändrades förändringar i tätheten av DAT-bindningsställen för att omfatta fler dorsala och rostrala delar av striatum med längre perioder med exponering för kokain (Letchworth et al,, 2001; Porrino et al,, 2004). I den aktuella studien verkade återgången till kontrollnivåer av DAT-koncentrationer under avhållsamhet vara större och snabbare i ryggstriatum än i ventralt striatum, och såg ut sålunda att följa en omvänd anatomisk bana till mönstret av effekter inducerade av kronisk kokaineksponering .
Föreliggande data överensstämmer också med rapporter från mänskliga kokainanvändare (Liten et al,, 1999; Malison et al,, 1998; Mash et al,, 2002; Staley et al,, 1994) som har visat förhöjda nivåer av bindning till DAT-ställen i striatum jämfört med kontroller, med de mest markanta ökningarna lokaliserade i det ventrale striatum. Nyligen har dessa förhöjningar visat sig åtföljas av signifikanta minskningar av vesikulär monoamintransportör 2 (VMAT2) -bindning (Liten et al,, 2003), vilket tyder på en faktisk förlust av DA-nervceller. Författarna drog slutsatsen att den förhöjda DAT sannolikt direkt berodde på ett kompensationssvar på den farmakologiska blockaden av kokain, medan minskningarna i VMAT2 mer sannolikt återspeglade de totala förändringarna i DA-metabolism, vilket resulterade i hypodopaminerg funktion.
Mänskliga kokainberoende har rapporterats ha minskat DA-koncentrationer i det ventrala striatum, mätt med PET, som svar på metylfenidatutmaning jämfört med friska kontroller (Volkow et al,, 1997). Nyligen, Martinez et al, (2007) rapporterade att kokainanvändare hade ett trubbigt svar på en amfetaminutmaning i det ventrala striatum och putamen. Dessutom var denna minskning av amfetamininducerad DA-frisättning korrelerad med valet för kokain i separata självadministrationssessioner, så att de användare med den lägsta graden av DA-frisättning som svar på amfetamin mest troligtvis skulle välja kokain framför en alternativ förstärkare (Martinez et al,, 2007). Nya studier i gnagarmodeller av kokainanvändning stöder också denna idé. Mateo et al, (2005)rapporterade till exempel att exponering för självadministrering av kronisk kokain är förknippad med förändringar i DAT-funktion. Dessa utredare visade att baslinjen av DA-upptag ökades, vilket resulterade i snabbare clearance av synaptisk DA, och därför minskade basnivåerna av extracellulärt DA eller ett hypodopaminergt tillstånd. Således är det troligt att de ökade DAT-koncentrationerna efter borttagning från kronisk kokain-självadministrering som observerats i den aktuella studien representerar ett kompensationssvar, vilket resulterar i lägre baslinjenivåer av extracellulär DA.
D2 mottagarändringar
Ett resultat från den aktuella studien var att koncentrationerna av D2-receptornivåer hade återgått till kontrollvärden efter 30 dagars avhållsamhet, jämfört med de signifikanta minskningar som observerades hos djur utan uttagningsperiod (Nader et al,, 2002). Till skillnad från den aktuella undersökningen har studier av mänskliga avbildning generellt sett funnit att D2-receptornivåer är lägre än kontrollerna efter långvarig abstinens från kronisk exponering för kokain (Martinez et al,, 2004; Volkow et al,, 1993). Potentiella förklaringar av skillnaderna mellan dessa mänskliga studier och den nuvarande icke-mänskliga primatundersökningen inkluderar skillnader i mönstret och varaktigheten av kokainintaget, liksom möjligheten att föregripa lägre nivåer av D2-receptorer hos mänskliga missbrukare.
I överensstämmelse med den senare idén finns det bevis som antyder att lägre basnivåer av D2-receptorer hos friska människor förutspår ökad förstärkande effekt av stimulanter såsom metylfenidatoch (Volkow et al,, 1999), och på liknande sätt hos apor, förutspådde baslinjenivåerna av D2-receptorer benägenheten att själv administrera kokain (Morgan et al,, 2002; Nader et al,, 2006). I överensstämmelse med den parallella karaktären av dessa fynd över arter, båda människor (Volkow et al,, 1993) och icke-mänskligt primat (Nader et al,, 2006) avbildningstudier har visat lägre nivåer av tillgänglighet av D2-receptor efter avhållsamhet från exponering av kokain. Speciellt schemat för självadministrering av kokain i det senare experimentet (Nader et al,, 2006) liknade schema för förstärkning som användes i denna studie. Således är det osannolikt att de olika resultaten från dessa två studier berodde på metodologiska skillnader, såsom schema för förstärkning eller kumulativt intag under självadministrering av kokain.
En mer trolig förklaring involverar den funktionella dynamiken i DA-systemet. Mätningar av tillgänglighet av D2-receptor med PET har beskrivits som "funktionell" eftersom signalen är relaterad till mängden protein (i detta fall densiteter av D2-receptorer) och nivåerna av cirkulerande neurotransmitter (Se Laruelle, 2000; Nader och Czoty, 2008 för vidare diskussion). jagI motsats härtill är receptorautoradiografi förorenad av cirkulerande nivåer av DA. Således antyder den aktuella studien, tillsammans med vårt tidigare arbete, att D2-receptortätheter minskas genom kokain självadministrering, men receptornivåerna verkar återhämta sig under avhållsamhet. I en liknande genomförd PET-studie (Nader et al,, 2006), återhämtning noterades i tre av fem apor. Föreliggande resultat tyder på att dessa apor förmodligen inte skilde sig åt i D2-receptortätheter, men kanske var responsen hos DA-systemet (dvs nivåer av cirkulerande DA under avhållsamhet) skilde mellan "återhämtade" och "icke återhämtade" individer.
Begränsningar
En viktig begränsning av de aktuella studierna är att våra studier inte kan behandla funktionaliteten hos D1 och D2 receptorer eller DAT. Snarare undersökte vi bara förändringarna i densiteten hos receptorproteiner. Även om resultaten har konsekvenser för de systemens potentiella roller, kommer ytterligare studier att krävas för att belysa beteendekonsekvenserna av förändringarna som visas här. En annan begränsning av den aktuella studien är att autoradiografiska ligander ofta misslyckas med att skilja mellan intracellulära och cytoplasmiska platser för deras mål, eftersom radiomärkta antagonister ofta är membranpermeabla. Liten et al, (2002) visade att kronisk exponering för kokain leder till en betydande uppreglering av DAT vid membranytan, samtidigt med en minskning av den intracellulära DAT-koncentrationen, i transfekterade celler. Nyligen, Samuvel et al, (2008) rapporterade om en liknande upptäckt i striatal synaptosomala preparat från råttor. Dessa resultat antyder att förändringarna i DAT-distributionen som observerats i den aktuella studien kan representera förändringar vid membranytan snarare än intracellulära ställen.
Slutligen bör en viss försiktighet ges vid tolkningen av studierna av förlängd (90 dagar) avhållsamhet, eftersom dessa fynd baserades på en relativt liten grupp av djur (N=3). Trots det lilla antalet försökspersoner var data som erhölls från denna grupp ganska konsekvent, vilket kan ses i en spridningsdiagram som visas i Figur 2. Koncentrationen av D1-receptorbindningsställen över striatum visade liten variation inom grupper, vilket antyder tillförlitligheten hos dessa fynd. Liknande konsistens var också tydligt i data från D2-receptor- och DAT-bindningsanalyser. Även om försiktighet bör iakttas, tyder dessa data starkt på att återställande av koncentrationer av DAT- och DA-receptorer inom striatum kan ske med långvarig abstinens.
Figur 2.
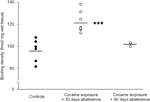 Bindningsdensiteter av D1-receptorer för enskilda djur i genomsnitt i hela striatum efter antingen livsmedelsförstärkning (kontroller) eller 30 eller 90 dagar avhållsamhet från kronisk kokain självadministrering. Organ för grupper visas med svarta staplar, ***p<0.001 jämfört med livsmedelsförstärkta kontroller.
Bindningsdensiteter av D1-receptorer för enskilda djur i genomsnitt i hela striatum efter antingen livsmedelsförstärkning (kontroller) eller 30 eller 90 dagar avhållsamhet från kronisk kokain självadministrering. Organ för grupper visas med svarta staplar, ***p<0.001 jämfört med livsmedelsförstärkta kontroller.
SLUTSATSER
Sammanfattningsvis orsakade exponering för självadministrering av kokain betydande förändringar i regleringen av DA-system som varade under de tidiga faserna (första 30 dagar) av avhållsamhet. Detta var mest tydligt vid regleringen av koncentrationen av D1-receptorer och DAT, både vad gäller deras förändringsstorlek och deras topografiska omfattning. Däremot fanns bevis för normalisering med längre avhållsamhetstider från exponering för kokain, eftersom koncentrationerna av DAT-, D1- och D2-receptorer efter 90 dagar av avhållsamhet inte skilde sig från koncentrationerna från icke-läkemedelseksponerade kontroller. These-system följer emellertid inte nödvändigtvis samma tidsmässiga återhämtningsförlopp, vilket antyder att det troligtvis finns en viss instabilitet i regleringen av DA-nivåer särskilt tidigt i avhållsamhet. Denna dopaminerge dysreglering kan påverka effektiviteten för eventuell farmakoterapi som ges till abstinenta kokainberoende, särskilt om medicinen är beroende av DA-systemet för dess verkningsmekanism.
Anmärkningar
AVSLÖJANDE/INTRESSEKONFLIKT
Författarna har ingen intressekonflikt att avslöja.
Referensprojekt
- Alleweireldt AT, Weber SM, Kirschner KF, Bullock BL, Neisewander JL (2002). Blockering eller stimulering av D1 dopaminreceptorer dämpar cue återinförande av släckt kokain-sökande beteende hos råttor. Psykofarmakologi (Berl) 159: 284–293. | Artikeln | PubMed | ChemPort |
- Beveridge TJ, Smith HR, Daunais JB, Nader MA, Porrino LJ (2006). Kronisk självadministrering av kokain är förknippad med förändrad funktionell aktivitet i temporala lober hos icke-humana primater. Eur J Neurosci 23: 3109–3118. | Artikeln | PubMed |
- Canfield DR, Spealman RD, Kaufman MJ, Madras BK (1990). Autoradiografisk lokalisering av kokainbindande platser med [3H] CFT ([3H] WIN 35428) i apahjärnan. Synapsen 6: 189–195. | Artikeln | PubMed | ChemPort |
- De Vries TJ, Schoffelmeer AN, Binnekade R, Vanderschuren LJ (1999). Dopaminergiska mekanismer som medierar incitamentet att söka kokain och heroin efter långvarigt tillbakadragande av självadministrering av IV-läkemedel. Psykofarmakologi (Berl) 143: 254–260. | Artikeln | PubMed | ChemPort |
- Farfel GM, Kleven MS, Woolverton WL, Seiden LS, Perry BD (1992). Effekter av upprepade injektioner av kokain på katekolaminreceptorbindningsställen, dopamintransportörs bindningsställen och beteende hos rhesus apa. Brain Res 578: 235–243. | Artikeln | PubMed | ChemPort |
- Graham J, Porrino LJ (1995). Neuroanatomiska underlag av kokain självförvaltning. CRC: Boca Raton, FL.
- Hemby SE, Co C, Koves TR, Smith JE, Dworkin SI (1997). Skillnader i extracellulära dopaminkoncentrationer i kärnan uppsamlas under responsberoende och svarsoberoende kokainadministrering i råttan. Psykofarmakologi (Berl) 133: 7–16. | Artikeln | PubMed | ChemPort |
- Henry DJ, White FJ (1991). Upprepad kokainadministration orsakar en kontinuerlig förbättring av D1-dopaminreceptorkänslighet inom råttkärnan. J Pharmacol Exp Ther 258: 882–890. | PubMed | ChemPort |
- Jacobsen LK, Staley JK, Malison RT, Zoghbi SS, Seibyl JP, Kosten TR et al, (2000). Förhöjd tillgänglighet av central serotonintransportör hos akut abstenta kokainberoende patienter. Am J Psykiatri 157: 1134–1140. | Artikeln | PubMed | ISI | ChemPort |
- Khroyan TV, Barrett-Larimore RL, Rowlett JK, Spealman RD (2000). Dopamin D1- och D2-liknande receptormekanismer i återfall till kokain-sökande beteende: effekter av selektiva antagonister och agonister. J Pharmacol Exp Ther 294: 680–687. | PubMed | ISI | ChemPort |
- Khroyan TV, Platt DM, Rowlett JK, Spealman RD (2003). Dämpning av återfall till kokain som söker av dopamin D1-receptoragonister och antagonister i icke-mänskliga primater. Psykofarmakologi (Berl) 168: 124–131. | Artikeln | PubMed | ChemPort |
- Laruelle M (2000). Imaging synaptisk neurotransmission med in vivo- bindande tävlingstekniker: en kritisk granskning. J Cereb Blood Flow Metab 20: 423–451. | Artikeln | PubMed | ISI | ChemPort |
- Letchworth SR, Nader MA, Smith HR, Friedman DP, Porrino LJ (2001). Progression av förändringar i dopamintransportörens bindningsställets densitet till följd av självadministrering av kokain hos rhesusapor. J Neurosci 21: 2799–2807. | PubMed | ISI | ChemPort |
- Lidow MS, Goldman-Rakic PS, Gallager DW, Rakic P (1991). Distribution av dopaminerga receptorer i primärt hjärnbark: kvantitativ autoradiografisk analys med användning av [3H] racloprid, [3H] spiperon och [3H] SCH 23390. Neuroscience 40: 657–671. | Artikeln | PubMed | ChemPort |
- Little KY, Elmer LW, Zhong H, Scheys JO, Zhang L (2002). Kokaininduktion av handel med dopamintransportör till plasmamembranet. Mol Pharmacol 61: 436–445. | Artikeln | PubMed | ISI | ChemPort |
- Little KY, Krolewski DM, Zhang L, Cassin BJ (2003). Förlust av striatal vesikulärt monoamintransporterprotein (VMAT2) hos humana kokainanvändare. Am J Psykiatri 160: 47–55. | Artikeln | PubMed |
- Little KY, Zhang L, Desmond T, Frey KA, Dalack GW, Cassin BJ (1999). Striatal dopaminerge avvikelser hos mänskliga kokainanvändare. Am J Psykiatri 156: 238–245. | PubMed | ChemPort |
- Malison RT, Best SE, van Dyck CH, McCance EF, Wallace EA, Laruelle M et al, (1998). Förhöjda striatal dopamintransportörer under akut kokainavhållsamhet mätt med [123I] beta-CIT SPECT. Am J Psykiatri 155: 832–834. | PubMed | ChemPort |
- Martinez D, Broft A, Foltin RW, Slifstein M, Hwang DR, Huang Y et al, (2004). Kokainberoende och tillgänglighet av d2-receptor i de funktionella underavdelningarna i striatum: förhållande till kokain-sökande beteende. Neuropsychopharmacology 29: 1190–1202. | Artikeln | PubMed | ISI | ChemPort |
- Martinez D, Narendran R, Foltin RW, Slifstein M, Hwang DR, Broft A et al, (2007). Amfetamininducerad dopaminfrisättning: markant avstannat i kokainberoende och förutsäga valet att själv administrera kokain. Am J Psykiatri 164: 622–629. | Artikeln | PubMed | ISI |
- Mash DC, Pablo J, Ouyang Q, Hearn WL, Izenwasser S (2002). Dopamintransportfunktionen är förhöjd hos kokainanvändare. J Neurochem 81: 292–300. | Artikeln | PubMed | ChemPort |
- Mateo Y, Brist CM, Morgan D, Roberts DC, Jones SR (2005). Nedsatt dopaminterminal funktion och okänslighet för kokain efter kokain beror på självadministrering och berövning. Neuropsychopharmacology 30: 1455–1463. | Artikeln | PubMed | ISI | ChemPort |
- Melega WP, Jorgensen MJ, Lacan G, Way BM, Pham J, Morton G et al, (2008). Långtidsadministrering av metamfetamin i vervet apan modellerar aspekter av människors exponering: neurotoxicitet i hjärnan och beteendeprofiler. Neuropsychopharmacology 33: 1441–1452. | Artikeln | PubMed | ChemPort |
- Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP (1998a). Effekt av självadministrering av kokain på striatal dopamin D1-receptorer i rhesusapor. Synapsen 28: 1–9. | Artikeln | PubMed | ChemPort |
- Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP (1998b). Effekt av självadministrering av kokain på dopamin D2-receptorer hos rhesusapor. Synapsen 30: 88–96. | Artikeln | PubMed | ChemPort |
- Morgan D, Grant KA, Gage HD, Mach RH, Kaplan JR, Prioleau O et al, (2002). Social dominans hos apor: Dopamin D2-receptorer och självadministrering av kokain. Nat Neurosci 5: 169–174. | Artikeln | PubMed | ISI | ChemPort |
- Nader MA, Czoty PW (2008). Hjärnavbildning i icke-mänskliga primater: insikter om narkotikamissbruk. ILAR J 49: 89–102. | PubMed | ChemPort |
- Nader MA, Daunais JB, Moore T, Nader SH, Moore RJ, Smith HR et al, (2002). Effekter av självadministrering av kokain på striatal dopaminsystem hos rhesusapor: initial och kronisk exponering. Neuropsychopharmacology 27: 35–46. | Artikeln | PubMed | ISI | ChemPort |
- Nader MA, Morgan D, Gage HD, Nader SH, Calhoun TL, Buchheimer N et al, (2006). PET-avbildning av dopamin D2-receptorer under självadministrering av kronisk kokain hos apor. Nat Neurosci 9: 1050–1056. | Artikeln | PubMed | ChemPort |
- Nolan EB, Harrison LM, Lahoste GJ, Ruskin DN (2007). Beteende synergism mellan D (1) och D (2) dopaminreceptorer hos möss beror inte på gapskorsningar. Synapsen 61: 279–287. | Artikeln | PubMed | ChemPort |
- Porrino LJ, Lyons D, Miller MD, Smith HR, Friedman DP, Daunais JB et al, (2002). Metabolisk kartläggning av effekterna av kokain under de initiala faserna av självadministrering i den icke-mänskliga primaten. J Neurosci 22: 7687–7694. | PubMed | ChemPort |
- Porrino LJ, Lyons D, Smith HR, Daunais JB, Nader MA (2004). Självadministrering av kokain producerar ett progressivt engagemang av limbiska, associerande och sensorimotoriska streatal domäner. J Neurosci 24: 3554–3562. | Artikeln | PubMed | ISI | ChemPort |
- Ruskin DN, Bergstrom DA, Walters JR (1999). Multisekundsvängningar i avfyrningshastighet i globus pallidus: synergistisk modulering av D1 och D2 dopaminreceptorer. J Pharmacol Exp Ther 290: 1493–1501. | PubMed | ISI | ChemPort |
- Samuvel DJ, Jayanthi LD, Manohar S, Kaliyaperumal K, Se RE, Ramamoorthy S (2008). Dysregulering av handel med dopamintransportör och funktion efter avhållsamhet från kokain självadministrering hos råttor: bevis för differentiell reglering i caudate putamen och nucleus accumbens. J Pharmacol Exp Ther 325: 293–301. | Artikeln | PubMed | ChemPort |
- Schmidt HD, Anderson SM, Pierce RC (2006). Stimulering av D1-liknande eller D2-dopaminreceptorer i skalet, men inte kärnan, i nucleus accumbens återställer kokain-sökande beteende hos råtta. Eur J Neurosci 23: 219–228. | Artikeln | PubMed |
- Self DW, Barnhart WJ, Lehman DA, Nestler EJ (1996). Motsatt modulation av kokain-sökande beteende av D1- och D2-liknande dopaminreceptoragonister. Vetenskap 271: 1586–1589. | Artikeln | PubMed | ISI | ChemPort |
- Staley JK, Hearn WL, Ruttenber AJ, Wetli CV, Mash DC (1994). Kokainigenkänningsplatser med hög affinitet på dopamintransportören är förhöjda hos dödliga kokainöverdosoffer. J Pharmacol Exp Ther 271: 1678–1685. | PubMed | ChemPort |
- Volkow ND, Fowler JS, Wang GJ, Hitzemann R, Logan J, Schlyer DJ et al, (1993). Minskad tillgänglighet av dopamin D2-receptorer är associerad med minskad frontal metabolism hos kokainmisbrukare. Synapsen 14: 169–177. | Artikeln | PubMed | ISI | ChemPort |
- Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Gifford A et al, (1999). Förutsägelse av förstärkande respons på psykostimulantia hos människor genom D2-receptornivåer i hjärnan. Am J Psykiatri 156: 1440–1443. | PubMed | ISI | ChemPort |
- Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Hitzemann R et al, (1997). Minskad striatal dopaminergisk respons hos avgiftade kokainberoende personer. Natur 386: 830–833. | Artikeln | PubMed | ISI | ChemPort |
- Walters JR, Bergstrom DA, Carlson JH, Chase TN, Braun AR (1987). D1 dopaminreceptoraktivering krävs för postsynaptisk expression av D2-agonisteffekter. Vetenskap 236: 719–722. | Artikeln | PubMed | ChemPort |
- Weiss F, Martin-Fardon R, Ciccocioppo R, Kerr TM, Smith DL, Ben-Shahar O (2001). Varaktig motståndskraft mot utrotning av kokain-sökande beteende inducerat av drogerelaterade signaler. Neuropsychopharmacology 25: 361–372. | Artikeln | PubMed | ISI | ChemPort |
- Wong DF, Kuwabara H, Schretlen DJ, Bonson KR, Zhou Y, Nandi A et al, (2006). Ökad beläggning av dopaminreceptorer i mänskligt striatum under cue-framkallade kokaintrang. Neuropsychopharmacology 31: 2716–2727. | Artikeln | PubMed | ChemPort |
