评论:本文的争议部分包含以下内容:
Prause的研究小组进行了许多基于电生理学的研究,这些研究提供了一些证据表明性欲,而不是性欲过度,可预测性唤起的自我调节。 [92]。 这些研究者在其他研究中表明,报告调节视力性刺激(VSS)观察问题的受试者也报告了较高的性欲,并且对VSS的反应显示出较低的晚期正电位(LPP)。 作者提出这种模式与物质成瘾模型不同 [93].
“ Prause小组”撰写的两篇EEG论文在同行评审期刊和其他期刊(包括YBOP)中受到广泛批评。 实际上,尽管他们的结论令人怀疑,但两篇论文的发现都与成瘾模型相符。 有关2013年脑电图的研究,请参阅:
- SPAN Lab的新色情研究(2013)中没有任何关联
- 同行评审批评: '高欲望',或'仅仅'成瘾? 对Steele等人的回应。 (2014),Donald L. Hilton,Jr.,MD,
- 同行评审批评:“互联网色情成瘾神经科学:回顾与更新”(2015年)
- 同行评审分析: “有或没有强迫性行为的人的性暗示反应的神经相关性”(2014年)
对于2015 EEG研究,请参阅:
如果研究人员真正阅读他们引用的研究内容,而不是简单地模仿主要作者的头条新闻,那不是很好吗?
关于出版物的链接
出版日期: 2015 年 10 月 12 日 (见历史)
作者: 10.7759 / cureus.348
引用本文为: Blum K,Badgaiyan RD,Gold MS(10月12,2015)Hypersexuality Addiction and Withdrawal:Phenomenology,Neurogenetics and Epigenetics。 Cureus 7(10):e348。 DOI:10.7759 / cureus.348
抽象
性欲过高被定义为性活动异常增加。 流行病学和临床研究表明,这种非寄生性疾病包括“过度”的性行为和障碍,并伴有个人困扰,社会和医学发病率。 就如何最好地将其归类为包括药物滥用在内的成瘾行为而言,这是一个极富争议性和政治性的话题。 将性欲障碍概念化为具有冲动性的非副性性欲障碍。 病理生理学观点包括性唤起和性欲失调,性冲动和性强迫性。 位于腹侧纹状体内的伏伏核介导滥用药物(例如可卡因,酒精,尼古丁,食物和音乐)的增强作用。 确实,据信这种结构要求由激励刺激引起的行为。 这些行为包括自然的奖励,例如进食,饮水,性行为和探索性运动。 积极强化的基本规则是,如果随后发生有益的事件,运动反应的强度和活力也会增强。 在这里,我们假设药物,音乐,食物和性行为对人类动机具有强大的作用,并且存在一个共同的作用机制(MOA)。 人类对三种必要的动机行为“饥饿,口渴和性行为”的驱动力可能都具有共同的分子遗传前因,如果受损,则会导致异常行为。 我们假设,基于大量的科学支持,性欲过剩的行为确实就像激活大脑中脑边缘奖励电路的药物,食物和音乐一样。 此外,多巴胺能基因以及可能的其他候选神经递质相关基因多态性会影响享乐和快感行为结果。 在当前文献中,关于性欲的遗传学和表观遗传学知之甚少。 但是,我们预计,基于临床工具评估并结合成瘾者基因分型的未来研究将为具有多态性关联的性类型的特定聚类提供证据。 已经有一些使用电生理技术的研究不支持以下观点,即性欲过剩确实类似于药物滥用和其他行为成瘾。 作者还鼓励临床和学术科学家开始使用神经影像工具来研究针对特定基因多态性的天然多巴胺能激动剂,以“正常化”性行为。
简介与背景
当然,在过去的十年中,在临床和研究环境中已经记录了过度性行为 [1]。 Benjamin Rush是美国创始人之一的医生,临床上记录了过度的性行为 [2] 19世纪西欧先驱性学家Richard von Krafft-Ebing和1948的Hirshfeld都继续这项工作 [3-4]。 这些调查人员的基本承租人表明,性欲过度在男性和女性中构成了持续存在社会偏离性的性行为,性欲过度是适应不良的。 在1975中,Stroller将这种状况描述为Don Juanism [5]。 在1969中,艾伦建议在Ellis和Sagarin支持的女性中使用男性和女性躁狂症进行satyriasis治疗。 [6-7]。 即使在DSM中未将性欲诊断作为精神病学诊断,但包括卡夫卡,里德,班克罗夫特在内的许多当代研究人员的工作以及世界卫生组织的意愿也可能导致将这种情况作为独立的诊断实体纳入 [8-11].
评价
文学方法论
截至12年2015月170日的Medline数据库用于执行基于互联网的文献搜索。 以下术语包括:性欲(479),性欲(1,652),性成瘾(1,842),性成瘾者(989),性冲动(946),强迫性(1,512),强迫性(782),性强迫(234) ),与恋爱症相关的疾病(857)和过度性行为(17)。 由于本文是一篇简短的评论,而不是荟萃分析,因此,本文基于与所涵盖子主题相关的这些研究的代表性选择。 不包括任何特定研究并不否认其在该领域的重要性。 可以理解的是,有些人不同意性成瘾确实是一种真正的疾病,甚至可能有证据表明性成瘾是有根据的。 但是,我们认为,有充分的证据表明性成瘾确实存在,并且涉及神经影像,神经遗传学甚至表观遗传学的研究都支持强迫性成瘾以及性欲亢进可以被视为成瘾性疾病的观点。 在我们认识到这种差异的同时,我们分别提出了自己的观点,以引起进一步的科学审查,而不是消除关于这一重要主题的科学言论。 为了进一步阐明该主题,我们在文献中进行了搜索以获得共识。 2015年46月XNUMX日,我们使用以下字词搜索了PUBMED Central:“性行为是真正的瘾吗?” 并检索到XNUMX条文章。
性瘾的定义
性成瘾被定义为会干扰正常生活并给家人,朋友,亲人和工作环境造成严重压力的任何强迫性行为。 性成瘾被称为性依赖,性欲亢进和性强迫症。 不管怎么说,这是一种强迫行为,完全控制着吸毒者的生活。 性上瘾者将性作为重中之重,而不是家人,朋友和工作。 性成为成瘾者生活的组织原则。 他们愿意牺牲自己最珍惜的东西来保存并继续其不健康的行为 [12]。 基于对性行为频率和相关性幻想所花费时间的终生评估,将性欲欲望描绘为欲望。 在男性中,Kafka和Hannen将性欲异常的评估定义为持续性行为的最高持续期(至少6个月最短持续时间)(总性出口/年龄15后)。 事实上,72-80%的男性寻求治疗副伤寒和副伤寒相关疾病,确定了上述操作定义的性欲欲望的纵向病史。 [13].
性欲亢进和性别差异
众所周知,在人类性社区和文学中,性欲被投降为性幻想,活动或冲动的存在,以及人类从事性行为的动机。 有内部和外部相关线索 [14]。 进化理论的支持者认为,男性和女性在性活动方面有不同的议程 [15]。 大量研究揭示了男性和女性之间的明显差异。 男性增加了性幻想 [16],手淫的频率增加 [17],增加了外部产生的视觉性唤起的倾向 [18] 对随意性行为的宽容态度 [19],轻松的唤醒 [20]和内在动机 [21]。 相比之下,女性表现出不同的性别景观,性动机,性唤起和性行为受到进化因素的影响 [22] 在生殖和育儿方面进行更大的生物,情感和时间投资 [23]。 女性不太容易受到性欲亢进的影响 [24] 并适应促进联盟关系和长期合作伙伴承诺 [25]。 虽然估计性成瘾在3%至6%的人口中受到影响,但对神经生物学前因的清晰认识有限。 [26] 以及临床评估 [27]。 我们鼓励进一步阅读性强迫,依恋和性取向 [28],以及对性刺激的反应中的性别差异 [29-30].
值得注意的是卡夫卡和亨嫩 [13],发现持续性性欲行为的平均发病年龄为性活跃男性的18.7±7.2年,且性欲行为发生的年龄范围为年龄7-46。 这种最高持续保持的性欲行为频率的平均持续时间为12.3±10.1年。 然而,这些活跃的性男性寻求治疗的性欲亢进的平均年龄为37±9年。 汉森等人。 还评估了罪犯的过度性行为,发现低犯罪者的再犯率低于高风险罪犯 [31].
性欲亢进和共病物质滥用
在性欲亢进与其他成瘾之间存在高度共病,例如物质使用障碍 [32-33]。 具体而言,Garcia和Thibaut提出过度非嗜性性别障碍的现象学应归类为成瘾行为,而不是强迫性或冲动性控制障碍。 [34]。 他们正确地指出,这些标准与其他人提出的成瘾性疾病非常接近 [35]。 这些研究人员为该领域的继续研究提供了动力,并为今后将性欲障碍纳入DSM-6提供了动力。 滥用毒品,摇滚乐和性行为是同时发生的,从伍德斯托克到现在,围绕这些组合的整个节日都已建立。
文献显示甲基苯丙胺的使用者报告说这种刺激性药物会增加性欲,特别是危险行为。 然而,苯丙胺已被证明可降低雌性大鼠的性活动。 考虑到这一点,Holder,et al。 评估甲基苯丙胺在雌性大鼠中的作用 [36]。 他们发现,恰恰相反,甲基苯丙胺促进了女性的性行为,这种效应是由于多巴胺能传播的增强,甚至由于卵巢激素和甲基苯丙胺的结合可能导致的神经传递。 具体而言,他们发现性动机的增强伴随着下丘脑内侧杏仁核和腹内侧核的神经元活动的激活。
此外,荷兰的科学家研究了自我识别的浪荡公子中药物滥用的共病 [37]。 在这项研究中,Spauwen等人。 结论是,79%的浪荡者报告使用娱乐性药物(包括饮酒和使用勃起功能障碍药物); 其中46%报告多种药物使用。 事实上,娱乐性药物使用(不包括酒精和勃起功能障碍药物)与男性和女性的高风险性行为显着相关。 此外,药物使用与女性浪荡公子中的性传播感染(STI)独立相关,特别是那些参与群体性行为的人。
Castelo-Branco,et al。 据报道,年轻的成年女性认为性行为是她们生活的重要组成部分,但不是主要关注点(77.6%) [38]。 他们还报告说,酒精消除了发生性行为的障碍(62.3%)。 重要的是,他们还发现酒精滥用是增强风险行为的预测变量,与女性的年龄无关。
值得注意的是贾,等。 据报告,精神兴奋剂和海洛因滥用者中存在危险的性行为,包括多次性交,偶然的性伴侣,同性恋伴侣以及从不或偶尔实行安全性行为 [39].
我们的主要承租人是甲基苯丙胺,可卡因,海洛因和酒精等毒品可以刺激非成瘾者的性欲。 在成瘾者中,它是完全不同的; 同样的药物可以长期引起快感缺失。 然而,在许多情况下,在恢复阶段后成瘾已经观察到类似壮阳药的行为。
性欲亢进和退缩
在PubMed中使用“同性恋和戒断症状”进行搜索(7-19-15)仅得到五篇文章,其中没有一篇描述“戒断症状”。 但是,使用“高性活动量戒断症状”一词进行的替代搜索导致列出25篇文章。
恢复报告中的成瘾者在长期禁欲期间饮食增加,食欲增加,某些食物和香烟滥用。 在最近和长期的禁食动物和人类中也已经证实并记录了体重增加 [39]。 Bruijnzeel做了有趣的观察,急性阿片戒断可导致自发性高潮 [40]。 重要的是,在文章中,Bruijnzeel提出,药物和可能的慢性强烈性活动的戒断症状可能是由于κ阿片受体信号的无阻碍功能,其抑制多巴胺的释放,同时增加脑回馈回路中的去甲肾上腺素。
文献中的论文缺乏关于尿液检测证实的急性和长期戒断和禁欲的严格性。 临床治疗计划越来越多地从治疗性别到分离两性,从而提供性别特异性治疗方案。 他们还提供教育,以解决性兴趣和活动的早期和长期增加,以及恢复与食物和暴饮暴食之间的关系。
一些研究者报告了过度性行为引起的戒断症状,其严重程度不同,并且是同时滥用药物 [41-45]。 作为这种搜索的结果,我们没有找到一篇描述与禁欲积极性接触相关的实际戒断症状的论文。 所引用的大多数论文都涉及戒除滥用药物的影响,例如阿片类药物,尼古丁,安非他明和可卡因,这些都可能损害性活动。
性欲亢进和神经遗传学
PubMed的搜索结果(7月19日至15日)仅列出了六篇使用“基因和性欲过剩”一词的论文,这些论文主要关注与克莱因-莱文综合症(KLS)有关的文章,这种疾病极少发生,导致性欲过剩可能长达27年。 在一项研究中,发现在KLS患者中检测到大量的免疫应答HLA-DQB0602,DQBXNUMX * XNUMX,这可能会增加KLS的风险 [46-47].
但是,当我们使用“性活动和基因”一词时,列出了2,826篇文章,并且提供了一些重要的神经遗传学方面的简要提要。 我们的假设是享乐和无享乐行为都是这些行为的个人风险等位基因一部分的结局,并且治疗包括适当地针对这些已识别的多态性。 此外,治疗反应还取决于这些风险等位基因,并为药物遗传学测试和药物基因组/营养学解决方案提供了重要的依据。
继Blum等人的有争议的初步调查结果之后。 在1990中DRD2 Al等位基因与严重酒精中毒之间关联的第一个证据,PubMed中有3,938文章(7-19-15) [48]。 这些研究涵盖了精神病学基因多态性,DRD2 A1等位基因以及相关的行为和生理学。 然而,尽管有大量的中脑边缘激活证据,特别是在多巴胺能通路和与性刺激和活动相关的神经元位点,但仍然缺乏将性活动与这种和其他相关基因联系起来的数据。 值得注意的是,Blum和Noble正确地将DRD2基因分类为负责所有奖赏缺陷综合症(RDS)行为的广义奖励基因。 事实上,使用Taq Al等位基因的贝叶斯定理分析载体,在其一生中,将有一个74%的机会,它们会与一个或多个奖励缺陷综合症(RDS)行为会合 [49].
任何基因多态性和性活动的第一个关联直到1999才发生,而米勒等人。 评估了一些多巴胺能基因 [50]。 基本发现是大脑中的多巴胺能系统似乎在性行为的调节中起主要作用。 在2非西班牙裔,欧裔美国男性和女性的样本中检查了D1,D4和D414多巴胺受体的基因与第一次性交时的年龄(AFSI)之间的关系。 当DRD2等位基因与DRD1等位基因相互作用时,在DRD2等位基因和AFSI之间观察到显着关联,并且甚至更强的关联。 构建了约束回归模型,使用性别和一组九个心理社会变量作为预测因子预测AFSI。 将DRD2和DRD2-by-DRD1预测变量添加到此模型分别将解释的方差增加了23%和55%。 事实上,这些研究结果表明男性之间的关联性强于女性,这与其他人的近期工作一致,即男性的性刺激反应高于女性。 [51]。 因此,也许“男人来自火星,女人来自金星”,甚至对可卡因滥用也是如此 [52].
具体而言,临床前和临床研究都表明,在可卡因成瘾过程的所有阶段(诱导,维持和复发)中,对可卡因的行为反应具有性别二态模式。 因此,出现了清晰的图景,表明可卡因成瘾的性别差异存在生物学基础。 这些差异是由于雄性和雌性性腺激素对中枢神经系统的不同调节所致,可能是由于DRD2基因多态性的存在所预测的。 [53]。 此外,众所周知,COMT与各种精神病学表型之间的遗传关联经常显示出男女之间的差异。 这些包括COMT中的功能性Val(158)Met多态性与男性强迫症和女性焦虑症表型相关。 此外,COMT中的Val(158)Met多态性对男孩的认知功能的影响大于女孩 [54].
米勒等人。 未发现与DRD4基因相关的多态性与第一次性交年龄的关联 [50]。 但是,其他人在某些种族中发现了重要的联系。 具体来说,他们对DRD4中多态性的分析表明,在所有种族中,具有-3R基因型的人发生第一次性行为的风险高于具有其他(或任何-4R)基因型的人(n = 2,552)。 有趣的是,非裔美国人样本中两种基因型的初次性交风险没有差异,这引发了文化养育的问题。 [55].
性体验,如反复吸毒,会产生长期变化,包括伏隔核(NAc)和纹状体背侧的致敏。 布拉德利等人。 使用微阵列分析研究仓鼠首次发现雄性或雌性动物的性经历差异上调或下调NAc中一系列基因的mRNA表达 [56]。 他们发现,与性天真的动物相比,在7周接受刺激雄性的性经验的仓鼠表现出大量基因的增加。 相反,在第7周未接受刺激雄性的性经验的雌性仓鼠表现出许多基因表达的减少。 这组作者说,这首关于雌性仓鼠的基因谱分析可以深入了解这些行为(性别)和滥用药物诱导中脑边缘和黑质纹状体多巴胺途径长期变化的机制。
双侧电极,双侧植入下丘脑外侧和黑质外侧被盖区,立体定位用于提供类似于性行为的慢性自我刺激奖励体验。 已发现这种类型的刺激诱导大鼠海马的CA3区域和运动皮层的分子层中突触数量的显着增加。 本质上,慢性脑刺激诱导长时程增强(LTP),已知增加新的突触连接 [57]。 在幼稚动物中单次暴露于可卡因足以引发腹侧被盖区(VTA)谷氨酸能突触的持续变化,其类似于其他大脑区域中的活动依赖性LTP。 这种可卡因诱导的LTP似乎是通过多巴胺D5受体激活N-甲基-D-天冬氨酸(NMDA)受体介导的,并且需要蛋白质合成。 [58],再次支持我们在这里提出的前提,药物和性别可能有共同的神经化学底物。
实证研究表明,性伴侣的数量与参与反社会行为之间存在正相关关系 [59]。 解释这种关联的大多数尝试都采用了进化的观点。 从进化的角度来看,与大量性伴侣有关的相同特征,例如冲动,短视和攻击性,也与犯罪参与有关。 然而,也有理由相信性伴侣和犯罪行为之间的共同变异可以通过共同的遗传途径来部分解释,其中与性伴侣数量相关的基因也与反社会行为有关。 使用上述基本原理,Beaver等人。 发现性伴侣和反社会行为之间存在强烈的正相关关系,多巴胺转运基因(DAT1)的多态性解释了性伴侣的数量和男性犯罪行为的变异 [59]。 DAT1基因的多态性效应和性伴侣的数量可能是由于某些多态性与男性早产性阴茎射精之间的关联。 与组合的1R1R / 9R9R(9R活性较低的多巴胺可用性)载体组相比,10OR / 9OR基因型的携带者得分表明射精在每个指标上的阈值较低。 [60]。 DAT1基因的多态性,特别是10R / 10R基因型,已在参加布朗学校(德克萨斯州圣马科斯)的青少年犯罪中被发现用于病理性攻击行为,包括反社会行为 [61]。 在盲法临床试验中,观察到DRD2和DAT1多态性与青少年病理性暴力的正相关。 此外,虽然最初概念化是由同伴模仿儿童发病或生命过程中持续存在的青年造成的,但双胞胎研究中有越来越多的证据表明,青少年发病或青少年限制的反社会行为也可能受遗传影响。 Burt和Mikolajewski不仅用DAT1基因证实了这些发现,而且将这些发现扩展到包括编码452-HT5A受体的基因的His2Tyr变体。 [62]最近,Jozkow等。 报告了衰老男性症状(AMS)的性别与5-HTR861B GXNUMXC遗传变异之间的关联 [63]。 此外,Sales,et al。 通过多变量逻辑回归分析发现滥用与5-HTTLPR组之间的相互作用,其中与非变化状态存在显着关联,并且随访时伴随通信频率得分 [64] 有干预史与干预后安全套使用不变的几率显着相关,只有那些有s等位基因的人。
已知加压素1a受体基因(Avpr 1a)的非编码区中的多态性与人,黑猩猩和田鼠的社会情绪特征相关,并且可能由于基因表达的位点特异性变异。 根据Barrett等人的说法,社会一夫一妻制的草原田鼠为研究一夫一妻制的神经生物学提供了独特的机会。 [65]。 事实上,血管加压素α受体(VlaR)信号传导对于雄性中双键的形成是必需的。 有趣的是,社会草原田鼠在奖赏处理腹侧苍白球中表现出比同一属的社会田鼠更大的VlaR结合。 巴雷特等人。 发现轻微的苍白球VlaR密度下调导致对交配女性伴侣的偏好显着受损,并且成年后焦虑样行为减少 [65]。 Garcia等的其他工作。 揭示具有DRD7的至少7个重复4个等位基因(50R +)的人报告说,其混杂性行为的分类率更高,包括曾经有过“一夜情”,并且报告的情况下,此类行为增加了XNUMX%以上性不忠 [66].
重要的是,Daw和Guo报道携带基因型DRD2 * A1 / A2,DRD2 * A2 / A2,DAT1 * 9R / 10R和MAOA * 2R /的个体与这些基因座上的其他基因型相比,无保护性交的几率更高。 [67]。 DRD2协会适用于男性和女性,而其他链接仅适用于女性。 最后,Emanuele等。 报道了DRD2 TaqI A基因型与“爱神”(一种爱心风格,其特征在于基于对伴侣的身体吸引力而发展出强烈的情感体验)之间以及C516T 5HT2A多态性与“躁狂症”之间存在显着关联。占有欲和依赖性的浪漫依恋,特征是自我挫败的情绪) [68].
表观遗传学和性活动
对文献的回顾表明,最近的一些文章指出了表观遗传效应对性活动的重要性。 例如,Matsuda回顾了雌激素受体a(ERalpha)的表观遗传变化以及对社会性行为的影响 [69]。 实际上,由表观遗传机制介导的ERα基因活性的改变,例如组蛋白修饰和DNA甲基化,会改变一个人的性行为。 在同性恋方面,赖斯等人。 开发了一个模型,该模型可以解释运河化(转化)同性恋性行为的发展 [70]。 他们解释说,该模型基于对胚胎干细胞中XX与XY核型的反应而设定的表观遗传标记。 因此,这些标记提高了对XY胎儿中睾酮的敏感性,并降低了XX胎儿的睾酮,从而消除了性发育。 据推测,这些转化表观遗传标记的子集可能在数量上具有跨基因遗传性,并可能导致异性后代的性发育镶嵌 - 同性恋表型。
在社会一夫一妻制的草原田鼠(Microtus ochrogaster)中,交配诱导由伴侣偏好形成引发并由多种神经递质调节的持久对键,包括催产素,加压素和多巴胺。 Gundersen的工作 [71]和王等人。 [72] 表明组蛋白去乙酰化酶可以促进可能与人类有关的雌性草原田鼠中的伴侣形成。 具体来说,王,等。 发现组蛋白去乙酰化酶抑制剂 - 丁酸钠和曲古抑菌素A(TSA)增强了雌性草原田鼠的伴侣偏好形成 [72]。 这种伴侣偏好形成与NAc中催产素受体(OTR,oxtr)和血管加压素V 1a受体(VlaR,avprla)的上调有关,通过其各自启动子处组蛋白乙酰化的增加。
有越来越多的兴趣证据表明,女性积极参与一妻多夫制,以避免遗传不相容或偏向父权,转而支持遗传上优越的男性。 选择优越的男性健康可能是由于表观遗传效应。 根据Zeh和Zeh的观点,与基于DNA序列的变异不同,表观遗传变异可能受到个体生命周期中经历的环境和随机效应的强烈影响。 [73]。 他们认为表观遗传变异可能对后交配性选择很重要,并可能解释将精子竞争能力与后代适应性联系起来的结果。
遗传和模因演变:人类生殖
艾森克提出外向性与加剧性行为之间,神经质与性行为问题(反社会行为)之间存在正相关。 一项与已婚人士进行的较早研究并未显示任何这些相关性。 据推测,这种联系仅存在于未建立长期关系的未婚者,因为关系的质量决定了性交。 在一个年轻未婚男子的样本中,外向性与项目之间存在正相关关系,在该项目中,该人描述了更多人,更频繁地描述了较早的性行为。 没有发现与神经质相关。 与其他人格和社交态度量表也有轻微的相关性。 由于与行动型人格量表的相关性,因此从社会心理学的角度解释了这些发现。 在当今社会中,期望年轻男性主动进行性交,使外向的年轻男性比内向的男性更能实现 [74]。 这种观点与理查德·布罗迪(Richard Brodie)关于心灵自私的基因的观点直接相符。 [75]。 当然,从脱氧核糖核酸的角度来看,人类学家会同意“我们仍然在这里只是出于一个原因; 继续繁殖。” 尽管进化的进展很慢,但与“模因进化相比,一个想法在阅读句子的时间上却发生了变化”,每20年左右迈出一步。 我们的大脑与遗传进化无关,因为它与聪明的人有更少的婴儿有关。 实际上,如果有一些基因使人们倾向于接受限制其后代数量的模因,那么它们将在几代后消失,而有利于使人们倾向于生育孩子的基因。 尽管有些争议,但不幸的是,许多研究表明,由于选择性交配,过去的42,000年智人智商降低了。 [76].
另外,事实证明,由于外向性与性活动增加有关,特别是在男性中,定量遗传学家估计外向性格的遗传性在40-60%左右。 Smillie及其同事研究并发现DRD2基因Al等位基因的一个拷贝与显着更高的外向性相关 [77]。 这种关联在人类繁殖方面提出了一个有趣的问题。 Comess认为,由于DRD2 A2携带者对生殖行为,学习障碍和其他冲动,强迫,攻击和成瘾性疾病具有显着影响,因此它们有可能导致DRDXNUMXAXNUMX等位基因频率的进行性和永久性改变,“导致该物种的遗传崩溃” [78].
在他的书中,Comings提供的证据表明,具有成瘾性破坏性行为的人早先生育孩子,这会影响成瘾基因的选择,如DRD2 Al等位基因 [79]。 他建议携带这种破坏性风险等位基因的个体将在20岁时生孩子,而没有这种等位基因的个体将在25岁时生孩子。 结果,突变基因将更快地繁殖,即每20年繁殖一次,而正常形式的基因每25年繁殖一次。 25/20的比率是1.25。 因此,具有1.25倍选择优势的基因的发生频率将随着世代的增加而增加。 母亲或父亲育有第一个孩子时,年龄相差五年就足以导致对年龄较大的群体发起生育的基因进行重要且相对快速的选择。 从1955年至今,已经记录了一些RDS行为的增加。 这些增加包括青少年行为综合症(毒品,性,青少年怀孕和犯罪行为,吸烟),品行障碍,犯罪,滥用药物,酗酒,无保护的性行为,未婚母亲,福利,开学和辍学,以及智商随之下降 [80]。 这些结果基于伯克利研究,该研究利用儿童健康与发育研究和全国青年纵向调查或NLYS的纵向数据 [81]。 利用这些信息,Comings预测,从1955到2015,例如DRD2 Al等位基因的频率会翻倍,从而增加RDS行为的流行,包括性早熟性行为 [50]。 我们鼓励对这个有趣的预测进行跟进。
尽管存在一些分歧,我们提出性功能亢进症作为RDS共享特征的亚型与物质和非物质成瘾行为,其临床表达部分受遗传学和表观遗传学的影响。 虽然目前尚未经过测试,但我们还建议短期FDA批准的药物辅助治疗(MAT),有利于阻断多巴胺功能,然后温和激活多巴胺能通路,导致长期多巴胺稳态。 后者可以通过一些可能有助于恢复的方式来实现。
尽管存在潜在偏倚,但它们包括多巴胺激动剂治疗 - 营养保健品(KB220),12步骤计划和传统,整体治疗,认知行为疗法(CBT)和创伤缓解疗法(TRT)以及多巴胺促进活动和食物(图 1) [82].
争议
虽然我们坚信在DSM的未来版本中应该包含性欲亢进症,但我们有点困惑的是,在神经遗传学和表观遗传学甚至戒断症状学和整体现象学方面对这种疾病了解甚少。 [83]。 最重要的信息是,我们现在鼓励科学界进行实验,特别是在神经成像和神经遗传学领域,包括基因特异性的表观遗传学,如催产素 - 血管加压素 - 食欲素 - 多巴胺以及其他奖励基因。 可能这种情况可能受益于针对奖励基因多态性以帮助促进多巴胺稳态的治疗 [84-89]。 Joranby等人的一些评论。 Edge和Gold支持与大脑奖励电路中共享的神经化学机制相关的常见治疗机会,如RDS概念中早先所述 [90-91].
从历史上讲,“性成瘾”被包括在DSM – III中,但是,由于DSM-1V作者的共识认为没有足够的证据值得得出结论,因此将其从DSM-1V中删除。 该决定引起了该领域领导者的极大热情。 此事件之后,包括卡夫卡,里德,普劳斯在内的许多科学家决定将“同性恋”定性为不是性成瘾而是独立的精神障碍而不是成瘾本身。 虽然他们在2010年的早期工作暗示此处提到的“同性恋”与性成瘾以及可能的其他成瘾(包括滥用药物)相似,但他们最近的工作却摆脱了这一争论。 在这一领域的最新工作揭示了持续的争议。 Prause的小组进行了许多基于电生理的研究,这些研究提供了一些证据表明,性欲而非性欲过高可以预测性唤起的自我调节 [92]。 这些研究者在其他研究中表明,报告调节视力性刺激(VSS)观察问题的受试者也报告了较高的性欲,并且对VSS的反应显示出较低的晚期正电位(LPP)。 作者提出这种模式与物质成瘾模型不同 [93]。 然而,虽然不涉及患有性欲紊乱的受试者,但Voon小组的工作表明,在暴露于性暴露视频的强迫性行为受试者中,神经网络中的活动类似于药物 - 线索 - 反应性研究中观察到的活动。 [94]。 更大的欲望或想要而不是喜欢进一步与这个神经网络中的活动相关联。 这项工作与激励动机理论相吻合 [95].
我们,即当前文章的作者,我们承认我们并不了解“同性恋症”的拥护者与他们希望将这种疾病纳入当前DSM-5的诚实意图之间发生的所有重要相互作用。 尽管它没有通过所谓的“酸测试”,但我们有充分的理由相信它将被包含在DSM的未来版本中。 值得注意的是,现任国立卫生研究院主任史蒂文·海曼(Steven Hyman)正确地指出,“帝斯曼(DSM)不能很好地反映临床和生物学的现实; 由于研究人员发现了研究和理解精神疾病的新颖方法,因此需要一种根本上用于诊断分类的新方法。” [96]。 此外,凯西,等。 提议虽然帝斯曼将不同的疾病视为不同的实体,但“疾病之间的界限通常不像帝斯曼所暗示的那样严格” [97].
在2014中,Karila等人。 一些精神病学家认为,性成瘾,也称为性欲紊乱,已被忽视,尽管这种情况会导致严重的社会心理问题。 根据这些作者的观点,他们提出性成瘾或性功能紊乱代表了同一问题的不同术语。 他们指出,性成瘾相关疾病的患病率从3%到6%不等。 因此,性成瘾/性功能紊乱的构造表现出有问题的行为,包括:过度手淫,网络化,色情使用,与成年人同意的异常性行为,电话性行为,脱衣舞俱乐部访问和其他成瘾行为 [98]。 当然,我们同意Carvalho等人指出,性成瘾和性欲亢进之间可能存在明显差异。 [99],Rettenberger,et al。 [100],Kor,et al。 [1],里德,等。 [9],卡夫卡和亨嫩 [13]和Prause等人。 [93-94] 在其他人中。
总之,我们已经提出,虽然性欲亢进与性成瘾之间存在一些差异,但还需要更多的研究来对这些非常重要的条件进行适当的分类。 我们同意Walters等人的工作。 [101] 这表明性欲亢进的个体差异在本质上是定量的而不是定性的。 他们还提出,性欲过度是在连续统一体的上端组织起来的(图 1).

图1:作为RDS子类型的Hypersexuality Disorder的描述性地图
该图说明了相互作用的神经遗传和表观遗传作用。 同时列出了短期多巴胺阻断和长期基于“多巴胺能体内稳态”的治疗方法以及多巴胺促进疗法和日常活动。 圆圈表示RDS特性,方框表示RDS行为。
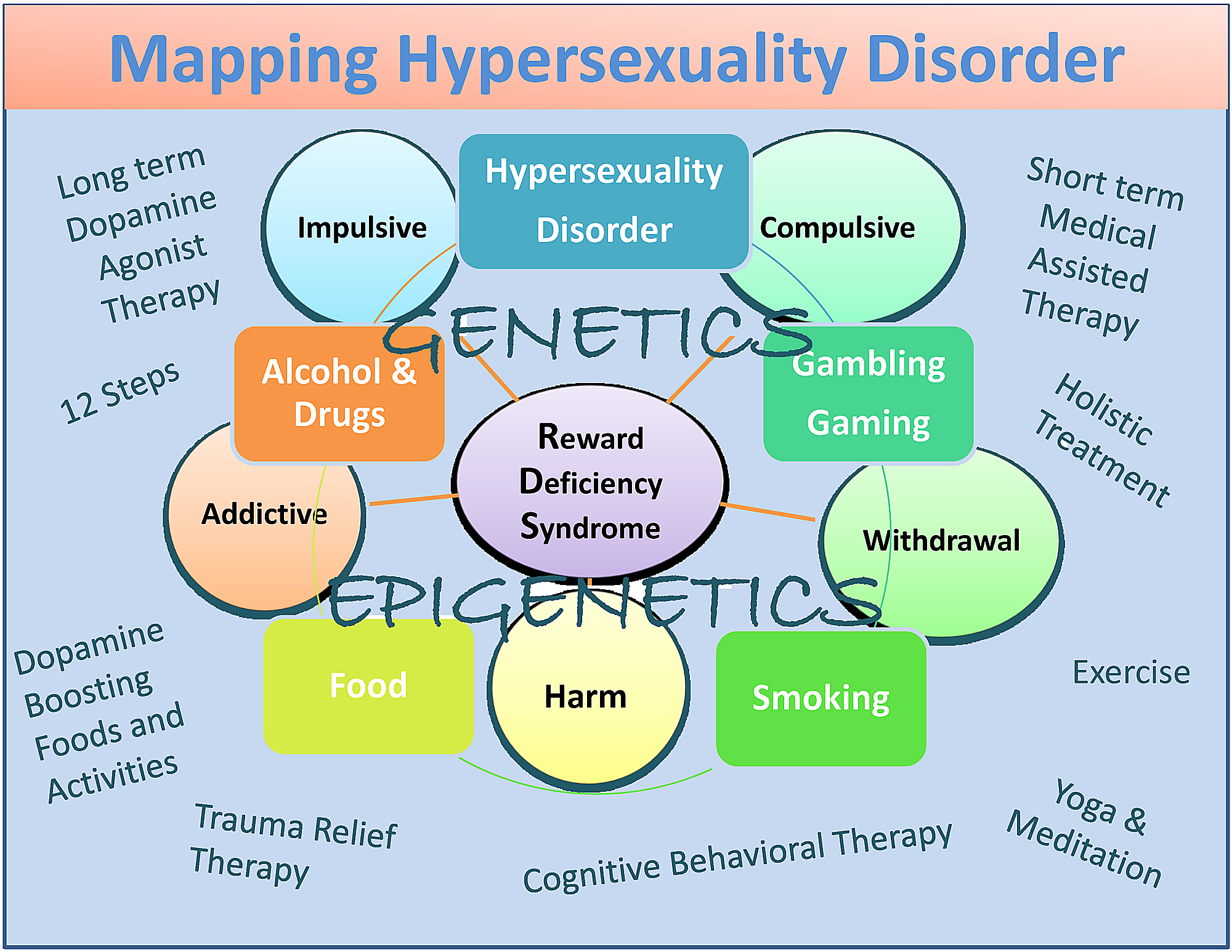{kind=link}
结论
在认识到争议的同时,我们建议应使用神经影像学(fMRI,PET,SPECT),光遗传学,候选物和微阵列分析以及表观遗传学技术来充分研究性欲障碍和性成瘾之间可能存在的异同。 我们相信,这些研究将为在以后的DSM版本中将性欲过剩纳入疾病提供依据。
案例
- Kor A,Fogel Y,Reid RC,Potenza MN: 性爱紊乱应该归类为成瘾吗?。 性瘾者强迫症。 2013,20:http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3836191/. 10.1080/10720162.2013.768132
- 拉什B: 心理疾病的医学调查和观察。 Rush B(编辑):Gryphon Editions Ltd.,Birmingham,AL; (原创作品1812)在1979中进行了审核。
- von Krafft-Ebing R: Psychopathia Sexualis。 克拉夫FS(编辑):纽约斯坦和日; (原创作品1886)在1965中进行了审核。
- Hirshfeld M: 性异常:性功能障碍的起源,性质和治疗。 Hirshfeld M(编辑):纽约艾默生书籍; 1948。
- 婴儿车RJ: 变态:仇恨的色情形式。 纽约万神殿图书; 1975。
- Allen CA: 一本关于性心理障碍的教科书。 牛津大学出版社,伦敦; 1962。
- Ellis A,Sagarin E: Nymphomania:对过度劳累女性的研究。 吉尔伯特出版社,纽约; 1964。
- 卡夫卡议员: 性欲紊乱发生了什么?。 Arch Sex Behav。 2014,43:1259-1261。 10.1007 / s10508-014 - 0326-Y
- Reid RC,Temko J,Moghaddam JF,Fong TW: 男性的羞耻,反刍和自我同情被评估为性欲亢进。 J Psychiatr Pract。 2014,20:260-268。 10.1097 / 01.pra.0000452562.98286.c5
- 班克罗夫特J: 人类性行为及其问题。 第三版。 Elsevier,英国牛津; 2009。
- 疾病和相关健康问题的国际统计分类,10th修订版 。 (2007)。 访问:7月23,2015: http://apps.who.int/classifications/apps/icd/icd10online2007/.
- 歌手B,Toates FM: 性动机。 J性研究。 1987,23:481-501。 10.1080/00224498709551386
- 卡夫卡议员,亨嫩·J: 男性的性欲欲望:男性患有与患有与瘫痪有关的疾病的男性不同的副作用?。 性虐待。 2003,15:307-321。 10.1023 / A:1025000227956
- DM巴士,Schmitt DP巴士: 性策略理论:人类交配的进化观点。 Psychol Rev. 1993,100:204-232。 10.1037 / 0033-295X.100.2.204
- Corbett-Detig RB,Hartl DL,Sackton TB: 自然选择限制了各种物种的中性多样性。 Psychol公牛。 1995,13:e1002112.-469-496。 doi:10.1371 / journal.pbio.1002112
- Laumann EO,Michael RT,Gagnon JH: 成人全国性调查的政治史。 Fam Plann Perspect。 1994,26:34-38。 10.2307/2136095
- 琼斯·J·C,巴洛·DH 自我报告的异性恋男性和女性的性冲动,幻想和自慰幻想的频率。 Arch Sex Behav。 1990,19:269-79。 10.1007 / BF01541552
- Oliver MB,海德JS: 性别中的性别差异:荟萃分析。 心理公牛。 1993,114:29-51。 10.1037 / 0033 - 2909.114.1.29
- Okami P,Shackelford TK: 性心理和行为的人类性别差异。 Annu Rev Sex Res。 2001,12:186–241。 10.1080/10532528.2001.10559798
- Leitenberg H,Detzer MJ,Srebnik D: 手淫中的性别差异以及青春期前和/或青春期早期的手淫经历与青年期性行为和性行为的关系。 Arch Sex Behav。 1993,22:87-98。 10.1007 / BF01542359
- Fetterman AK,Kruger NN,Robinson MD: 与性别相关的交配策略与生殖器显着性的操纵不同。 Motiv Emot。 2015,39:99-103。 10.1007/s11031-014-9420-7
- Basson R: 对女性性反应使用不同的模型来解决女性的性欲低下问题。 J性爱婚姻疗法。 2001,27:395–403。 10.1080/713846827
- Andersen BL,Cyranowski JM,Aarestad S: 除了人为的,与性别相关的区别,以概念化女性的性行为:对鲍迈斯特的评论。 心理公牛。 2000,126:380-389。 10.1037 0033 // - 2909.126.3.380
- Knight RA,Sims-Knight JE: 针对女性的性胁迫的发展前因:用结构方程模型测试替代假设。 Ann NY Acad Sci。 2003,989:72-85。 10.1111 / j.1749-6632.2003.tb07294.x
- Blum K,Werner T,Carnes S,Carnes P,Bowirrat A,Giordano J,Oscar-Berman M,Gold M: 性,毒品和摇滚乐:假设常见的中脑边缘激活是奖励基因多态性的函数。 J精神药物。 2012,44:38-55。 10.1080/02791072.2012.662112
- Carnes PJ,Green BA,Merlo LJ,A Polles A,Carnes S,MS金: PATHOS:用于评估性成瘾的简短筛查应用程序。 J Addict Med。 2012,6:29-34。 10.1097/ADM.0b013e3182251a28
- Hanson RK: Static-99是否预测老年性犯罪者的再犯?。 性虐待。 2006,18:343–355。 10.1007 / s11194-006 - 9027-Y
- Weinstein A,Katz L,Eberhardt H,Cohen K,Lejoyeux M: 性强迫 - 与性,依恋和性取向的关系。 J Behav Addict。 2015,4:22-26。 10.1556 / JBA.4.2015.1.6
- Chung WS,Lim SM,Yoo JH,Yoon H: 大脑激活与视听性刺激的性别差异; 为了响应同一个视频片段,女性和男性是否会经历相同程度的唤醒?。 Int J Impot Res。 2013,25:138-142。 10.1038 / ijir.2012.47
- Rupp HA,Wallen K: 对视觉性刺激的反应性别差异:综述。 Arch Sex Behav。 2008,37:206-218。 10.1007/s10508-007-9217-9
- Hanson RK,Harris AJ,Helmus L,Thornton D: 高危性犯罪者可能不会永远存在高风险。 J Interpers Violence。 2014,29:2792-813。 10.1177/0886260514526062
- Reid RC,Cyders MA,Moghaddam JF Fong TW: 赌博障碍,性欲亢进和甲基苯丙胺依赖患者的Barratt冲动量表的心理测量特性。 Addict Behav。 2014,39:1640-1645。 10.1016 / j.addbeh.2013.11.008
- Reid RC,Carpenter BN,Hook JN,Garos S,Manning JC,Gilliland R,Cooper EB,McKittrick H,Davtian M,Fong T: 针对性欲亢进症的DSM-5田间试验的研究结果报告。 J性别医学。 2012,9:2868-2877。 10.1111 / j.XNUMX 1743.x
- Garcia FD,Thibaut F: 性瘾。 Am J Drug Alcohol Abuse。 2010,36:254-260。 10.3109/00952990.2010.503823
- Reid RC,Bramen JE,Anderson A,Cohen MS: 在性欲亢进的患者中正念,情绪失调,冲动和压力倾向。 J Clin Psychol。 2014,70:313-321。 10.1002 / jclp.22027
- 持有人MK,Hadjimarkou MM,Zup SL,Blutstein T,Benham RS,McCarthy MM,Mong JA: 甲基苯丙胺促进女性性行为并增强下丘脑内侧杏仁核和腹内侧核的神经元激活。 心理神经。 2010,35:197-208。 10.1016 / j.psyneuen.2009.06.005
- Spauwen LW,Niekamp AM,Hoebe CJ,Dukers-Muijrers NH: 浪荡公子中的药物使用,性风险行为和性传播感染:荷兰的一项横断面研究。 性传染感染。 2015,91:31-36。 10.1136 / sextrans-2014,051626
- Castelo-Branco C,Parera N,Mendoza N,Pérez-Campos E,Lete I,CEA group: 年轻成年女性中的酒精和药物滥用以及危险的性行为。 Gynecol Endocrinol。 2014,30:581-586。 10.3109/09513590.2014.910190
- 贾志杰,严SY,包玉萍,连泽,张力,刘志敏: 苯丙胺类兴奋剂使用者和海洛因使用者之间的性行为差异。 J Addict Med。 2013,7:422-427。 10.1097/ADM.0b013e3182a952b2
- Bruijnzeel AW: κ-阿片受体信号传导和脑回报功能。 Brain Res Rev. 2009,62:127-146。 10.1016 / j.brainresrev.2009.09.008
- Orsini CA,Ginton G,Shimp KG,Avena NM,Gold MS,Setlow B: 停止慢性安非他明给药后的食物消耗和体重增加。 食欲。 2014,78:76-80。 10.1016 / j.appet.2014.03.013
- 易卜拉欣C,雷纳特C: 老年人神经认知障碍中的性欲过剩–文献综述和案例研究。 Psychiatr Danub。 2014,26:36-40。
- Vallejo-Medina P,Sierra JC: 西班牙男性药物依赖性样本中药物使用的影响和禁欲对性功能的影响:多地点研究。 J性别医学。 2013,10:333-341。 10.1111 / j.XNUMX 1743.x
- Baskerville TA,道格拉斯AJ: 多巴胺和催产素相互作用的潜在行为:对行为障碍的潜在贡献。 CNS Neurosci Ther。 2010,16:e92-123。 10.1111 / j.XNUMX 1755.x
- Riebe CJ,Lee TT,Hill MN,Gorzalka BBB: 沉淀戒断可抵消亚慢性大麻素给药对雄性大鼠性行为的不利影响。 Neurosci Lett。 2010,472:171-174。 10.1016 / j.neulet.2010.01.079
- Korda JB,Pfaus JG,Kellner CH,Goldstein I: 持续性生殖器性唤起障碍(PGAD):电痉挛治疗长期对症治疗的病例报告。 J性别医学。 2009,6:2901-2909。 10.1111 / j.XNUMX 1743.x
- Huang CJ,Liao HT,Yeh GC,Hung KL: Kleine-Levin综合征患者HLA-DQB1等位基因的分布。 J Clin Neurosci。 2012,19:628-630。 10.1016 / j.jocn.2011.08.020
- Blum K,Noble EP,Sheridan PJ,Montgomery A,Ritchie T,Jagadeeswaran P,Nogami H,Briggs AH,Cohn JB: 人体多巴胺D2受体基因在酒精中毒的等位基因关联。 JAMA。 1990,263:2055-2060。 10.1001 / jama.263.15.2055
- Blum K,Sheridan PJ,Wood RC,Braverman ER,Chen TJ,Cull JG,Comings DE: D2多巴胺受体基因作为奖赏缺乏综合征的决定因素。 JR Soc Med。 1996,89:396-400。 10.1177/014107689608900711
- Miller WB,Pasta DJ,MacMurray J,Chiu C,Wu H,Comings DE: 多巴胺受体基因与第一次性交时的年龄相关。 J Biosoc Sci。 1999,31:43-54。 10.1017 / S0021932099000437
- Hamann S,Herman RA,Nolan CL,Wallen K: 男性和女性对杏仁核对视觉性刺激的反应不同。 Nat Neurosci。 2004,7:411-416。 10.1038 / nn1208
- Quiñones-Jenab V: 为什么来自金星的女性和来自火星的男性滥用可卡因?。 Brain Res。 2006,1126:200-203。 10.1016 / j.brainres.2006.08.109
- Noble EP,Blum K,Khalsa ME,Ritchie T,Montgomery,A Wood RC,Fitch RJ,Ozkaragoz T,Sheridan PJ,Anglin MD,Paredes A,Treiman LJ,Sparkes RS: D2多巴胺受体基因与可卡因依赖的等位基因关联。 药物酒精依赖。 1993,33:271-285。 10.1016/0376-8716(93)90113-5
- Harrison PJ,Tunbridge EM: 儿茶酚-O-甲基转移酶(COMT):一种导致脑功能性别差异的基因,以及导致精神疾病倾向的性别二态性。 神经精神药理学。 2008,33:3037-3045。 10.1038 / sj.npp.1301543
- 郭庚,佟烨: 首次性交,基因和社会背景的年龄:来自双胞胎和多巴胺D4受体基因的证据。 人口。 2006,43:747-769。 10.1353 / dem.2006.0029
- Bradley KC,Boulware MB,Jiang H,Doerge RW,Meisel RL,Mermelstein PG: 性经验后伏隔核和纹状体内基因表达的变化。 基因脑行为。 2005,4:31-44。 10.1111 / j.1601-183X.2004.00093.x
- Rao BS,Raju TR,Meti BL: 自我刺激奖励经验后海马CA3区和运动皮层分子层突触的数值密度增加。 神经科学。 1999,91:799-803。 10.1016/S0306-4522(99)00083-4
- Heshmati M: 腹侧被盖区可卡因诱导的LTP:对机制和时间过程的新见解阐明了成瘾的细胞基质。 J神经生理学。 2009,101:2735-2737。 10.1152 / jn.00127.2009
- Beaver KM,Wright JP,Walsh A: 基于基因的进化解释,用于确定犯罪参与与性伴侣数量之间的关系。 Biodemography Soc Biol。 2008,54:47-55。 10.1080/19485565.2008.9989131
- Santtila P,Jern P,Westberg L,Walum H,Pedersen CT,Eriksson E,Sandnabba N: 多巴胺转运蛋白基因(DAT1)多态性与早泄有关。 J性别医学。 2010,7:1538-1546。 10.1111 / j.XNUMX 1743.x
- Chen TJH,Blum K,Mathews D,Fisher L,Schnautz N,Braverman Er,Schoolfield J,Downs W,Blum SH,Mengucci J,Meshkin B,Arcuri V,Bajaj A,Waite RL,Comings DE: 多巴胺D2受体(DRD2)[Taq1 A1等位基因]和多巴胺转运蛋白(DAT1)[480 bp Allele]基因与病理性攻击行为的初步关联,这是青少年奖赏缺陷综合症(RDS)的临床亚型。 Gene Ther Mol Biol。 2007,1:93-112。 访问:7月23,2015: http://gtmb.org/pages/Vol11A/HTML/11._Chen_et_al,_93-102.htm.
- Burt SA,Mikolajewski AJ: 初步证据表明特定候选基因与青少年发病的反社会行为有关。 侵略行为。 2008,34:437-445。 10.1002 / ab.20251
- JóźkówP,Słowińska-LisowskaM,ŁaczmańskiŁ,MędraśM: 神经递质受体基因的多态变异可能影响衰老男性的性功能:HALS研究的数据。 神经内分泌学。 2013,98:51-59。 10.1159/000350324
- 销售JM,DiClemente RJ,Brody GH,Philibert RA,Rose E: 参与艾滋病毒预防干预措施后的5-HTTLPR多态性与青少年非裔美国女性避孕套使用行为的相互作用。 Prev Sci。 2014,15:257-267。 10.1007/s11121-013-0378-6
- 巴雷特(Barrett)CE,基博(Keebaugh)AC,艾伦(Ahern),巴斯(Bass CE),特威利格(Terwilliger)EF,年轻LJ: 血管加压素受体(Avpr1a)表达的变异在草原田鼠的一夫一妻制行为中产生多样性。 Horm Behav。 2013,63:518-526。 10.1016 / j.yhbeh.2013.01.005
- Garcia JR,MacKillop J,Aller EL,Merriwether AM,Wilson DS,Lum JK: 多巴胺D4受体基因变异与不忠和性滥交之间的关联。 PLoS One。 2010,5:e14162。 10.1371 / journal.pone.0014162
- 道J,郭G: 三种基因对青少年是否使用避孕的影响,美国,1994-2002年。 Popul Stud(Camb)。 2011,65:253-271。 10.1080/00324728.2011.598942
- Emanuele E,Brondino N,Pesenti S,Re S,Geroldi D: 对人类爱情风格的遗传加载。 Neuro Endocrinol Lett。 2007,28:815-821。
- Matsuda KI: 雌激素受体α基因启动子的表观遗传变化:社会性行为的影响。 前神经科。 2014,8:344。 10.3389 / fnins.2014.00344
- 赖斯WR,弗里贝格U,加夫里莱特S: 通过管性性发育进行同性恋:一种新表观遗传模型的测试方案。 BIOESSAYS。 2013,35:764-770。 10.1002 / bies.201300033
- Gundersen B: 通过表观遗传学进行配对。 Nat Neurosci。 2013,16:779。 10.1038 / nn0713-779
- Wang H,Duclot F,Liu Y,Wang Z,Kabbaj M: 组蛋白去乙酰化酶抑制剂促进雌性草原田鼠中的伴侣偏好形成。 Nat Neurosci。 2013,16:919-924。 10.1038 / nn.3420
- Zeh JA,Zeh DW: 母体遗传,表观遗传学和一妻多夫制的演变。 遗传学。 2008,134:45-54。 10.1007 / s10709-007-9192-Z
- Addad M,Lesiau A: 外向性,神经质,不道德的判断和犯罪行为。 医学法。 1989,8:611-622。
- Brodie R: 心灵的病毒:模因的新科学。 Hay House,Inc,纽约,纽约; 1996,第66页。
- Hernnstein R,Murray C: 钟形曲线:美国生活中的智慧与阶级结构。 自由新闻,纽约,纽约; 1994。
- Smillie LD,Cooper AJ,Proitsi P,Powell JF,Pickering AD: DRD2多巴胺基因的变异预示着外向性格。 Neurosci Lett。 2010,468:234-327。 10.1016 / j.neulet.2009.10.095
- 来自DE: 基因炸弹。 高等教育和先进技术是否加速了学习障碍,注意力缺陷障碍,成瘾性和破坏性行为的基因选择?。 Hope Press,Duarte CA; 1996。
- 来自DE: 青少年问题行为综合症。 基因炸弹。 高等教育和先进技术是否加速了学习障碍,注意力缺陷障碍,成瘾性和破坏性行为的基因选择? Hope Press,Duarte CA; 1996。 pp 91-94。
- 来自DE: 基因选择。 基因炸弹。 高等教育和先进技术是否加速了学习障碍,注意力缺陷障碍,成瘾性和破坏性行为的基因选择? Hope Press,Duarte CA; 1996。 pp 89-90。
- 来自DE: 伯克利研究。 基因炸弹。 高等教育和先进技术是否加速了学习障碍,注意力缺陷障碍,成瘾性和破坏性行为的基因选择? Hope Press,Duarte California; 1996。 pp 105-210。
- Borsten J: 马里布海滩恢复饮食食谱。 Vidov Publishing Inc,Malibu,CA; 2015。
- Derbyshire KL,Grant JE: 强迫性行为:文献综述。 J Behav Addict。 2015,4:37-43。 10.1556/2006.4.2015.003
- Nirenberg MJ: 多巴胺激动剂戒断综合征:对患者护理的影响。 药物老化。 2013,30:587-592。 10.1007 / s40266-013-0090-Z
- 明尼苏达州波坦察市Brewer JA的Grant JE: 物质和行为成瘾的神经生物学。 CNS光谱。 2006,11:924-930。
- 坂田K,公爵SM: 通过启动子IV缺乏BDNF表达会干扰额叶皮层和海马中单胺基因的表达。 神经科学。 2014,260:265-75。 10.1016 / j.neuroscience.2013.12.013
- Blum K,Liu Y,Wang W,Wang Y,Zhang Y,Oscar-Berman M,Smolen A,Febo M,Han D,Simpatico T,CronjéFJ,Demetrovics Z,Gold MS: KB220Z™对戒断基因型海洛因成瘾者奖赏回路中神经通路的rsfMRI作用。 Postgrad Med。 2015,127:232-241。
- McLaughlin T,Blum K,Oscar-Berman M,Febo M,Demetrovics Z,Agan G,Fratantonio J,Gold MS: 使用Neuroadaptagen KB200z™改善RDS患者的恐怖,清醒噩梦:增强,脑奖励,功能连接和多巴胺能稳态的作用。 J Reward Defic Syndr。 2015,1:24-35。 10.17756 / jrds.2015-006
- Blum K,Thanos PK,Badgaiyan RD,Febo M,Oscar-Berman M,Fratantonio J,Demotrovics Z,Gold MS: 奖励缺陷综合症的神经遗传学和基因治疗:我们要去应许之地吗?。 专家Opin Biol Ther。 2015,5:973-985。 10.1517/14712598.2015.1045871
- Joranby L,Pineda-Frost KY,Gold MS: 对食物和大脑奖励系统的成瘾。 性成瘾和强迫性。 2005,12:201–217。 10.1080/10720160500203765
- Edge PJ,金牌MS: 戒毒和饮食过多:吸烟和其他药物的教训。 Curr Pharm Des。 2011,17:1173-1179。 10.2174/138161211795656738
- Moholy M,Prause N,Proudfit,GH,Rahman A,Fong T: 性欲,而不是性欲过度,预测性唤起的自我调节。 Cogn Emot。 2015,6:1012。
- Prause N,Steele,VR,Staley C,Sabatinelli,D,Hajcak G: 在问题使用者和控制中通过性图像调制晚期正电位与“色情成瘾”不一致。 Biol Psychol。 2015,109:192-199。 10.1016 / j.biopsycho.2015.06.005
- Mechelmans DJ,Irvine M,Banca P,等: 在有和没有强迫性行为的个体中,对性暗示线索的注意偏向增强。 PLoS One。 2014,25,9(8):e105476。 10.1371 / journal.pone.0105476
- Blum K,Gardner E,Oscar-Berman M,Gold M: 与奖励缺乏综合症(RDS)相关的“喜欢”和“想要”:假设大脑奖励电路中的差异反应性。 Curr Pharm Des。 2012,18(1):113-118。
- 海曼SE: 诊断DSM:诊断分类需要基本改革。 大脑。 2011,2011:6。 访问:2011 Apr 26: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3574782/.
- Casey BJ,Craddock N,Cuthbert BN,Hyman SE,Lee FS,Ressler KJ: DSM-5和RDoC:精神病学研究的进展?。 Nat Rev Neurosci。 2013,14:810-14。 10.1038 / nrn3621
- Karila L,WéryA,Weinstein A,Cottencin O,Petit A,Reynaud M,Billieux J: 性成瘾或性欲紊乱:同一问题的不同术语? 文献综述。 Curr Pharm Des。 2014,20:4012-20。 10.2174/13816128113199990619
- Carvalho J,ŠtulhoferA,Vieira AL,Jurin T: 性欲过度和性欲高:探索有问题的性行为的结构。 J性别医学。 2015,12:1356-67。 访问:2015 Mar 23: http://onlinelibrary.wiley.com/doi/10.1111/jsm.12865/abstract;jsessionid=6F416CCBB66B7F0EA28E428D4993EBD5.f01t04. 10.1111 / jsm.12865
- Rettenberger M,Klein V,Briken P: 性功能,性兴奋,性抑制和人格特征的关系。 Arch Sex Behav。 2015,Jan 6 :(电子版提前印刷)。 访问:2015 Jan 6: 10.1007/s10508-014-0399-7
- Walters GD,Knight RA,LångströmN: 性欲亢奋吗? 来自一般人群和临床样本的DSM-5的证据。 Arch Sex Behav。 2011,40:1309-21。 访问:2011 Feb 3: 10.1007/s10508-010-9719-8