前神经科。 2015 Mar 18; 9:86。 doi:10.3389 / fnins.2015.00086。 eCollection 2015。
特贝克S.1, Akkus F.2, 切斯特曼LP3, 哈斯勒G.2.
抽象
在本文中,我们概述了代谢型谷氨酸受体5(mGluR5)的活性和密度与病理性焦虑,情绪障碍和成瘾的关系。 具体来说,我们将描述采用正电子发射断层扫描(PET)并将其发现与临床前动物研究相结合的人类mGluR5研究。 从基本的神经生物学方法到人体研究,这种不同方法学方法的综合观点可能比仅基于临床前数据的观点更全面和临床相关地提供了mGluR5在心理健康中的作用的观点。 我们还将根据研究领域标准(RDoC)审查有关mGluR5的当前研究数据。 首先,我们发现了与正价系统和负价系统有关的异常谷氨酸活性的证据,这表明拮抗mGluR5干预具有显着的抗上瘾,抗抑郁和抗焦虑作用。 其次,有证据表明,mGluR5在社会功能系统和对社会压力的反应中起着重要作用。 最后,mGluR5在睡眠稳态中的重要作用表明该谷氨酸受体可能在RDoC的唤醒和调节系统域中发挥重要作用。 谷氨酸以前主要是在非人类研究中进行的,但是,最初的人类临床PET研究现在也支持这样的假说,即通过介导大脑兴奋性,神经可塑性和社会认知,代谢型谷氨酸的异常活动可能使个体容易出现广泛的精神病问题。
介绍
谷氨酸是大脑中主要的兴奋性神经递质,许多研究人员认为它在各种心理健康和医疗条件中起着重要作用。 实际上,研究人员经常提到以激动剂或拮抗剂的形式开发基于离子型或代谢型谷氨酸盐的药理学治疗多种精神疾病的潜力。 关于谷氨酸能系统的知识在过去十年中已经取得了很大进展,这是由于人类受体和发射器成像技术的进步所致。 尽管有大量关于谷氨酸盐干预各种精神疾病的实验研究,但缺乏系统评价,重点是将临床前动物研究结果与其他神经科学方法相结合,如人体正电子发射断层扫描(PET) 。 因此,与之前的评论相比(Swanson等,2005; Pittenger等,2006; Kalivas,2009; Brennan等,2012; Luykx等,2012; Riaza Bermudo-Soriano等,2012),这篇综述研究人体研究,主要是PET研究,并将这些结果与临床前研究结果相结合。
正电子发射断层扫描(PET)
PET是一种核,敏感和非侵入性医学成像技术,用于成像受体分布,浓度和功能。 为了鉴定人脑受体,需要开发放射性标记的受体配体(示踪剂)。 扫描仪设备检测由示踪剂发射的伽马射线,该伽马射线被引入体内。 适用于人体研究的mGluR5 PET示踪剂在瑞士Villigen的Paul Scherrer研究所(PSI)和瑞士苏黎世的瑞士联邦理工学院(ETH)成功开发。 ABP688是一种非竞争性和高选择性拮抗剂,可与mGluR5的变构位点结合。 11C-ABP688显示出对mGluR5的高选择性和富含受体的大脑区域的高摄取。 11C-ABP688在动物中的这些特征的首次描述已经发表(Ametamey等,2006)。 我们已经看到在使用β-探针估计该示踪剂的动力学的大鼠中进行的研究的有希望的结果。 此外,当我们在人类中进行第一项PET研究以估计人类的动力学时(Ametamey等,2006)我们发现结果与大鼠研究中发现的结果相当(Soares和Law,2009)。 我们现已成功使用开发的示踪剂对健康志愿者和精神病患者的mGluR5进行研究。 PET技术提供有关所检查的脑区域内受体相对密度的信息。 没有关于神经递质浓度的信息(即,高受体密度是否可能导致或是神经递质作用增加或减少的结果)。 PET研究提供了有关活体人类患者受体水平异常的信息,因此有助于评估应该通过药物治疗靶向哪些受体。 相反,磁共振波谱(MRS)提供 体内 检查组织的生化信息(Hasler等,2009并因此提供关于患者组中谷氨酸代谢物的相对量的信息。 PET和MRS研究的结果可为临床前动物研究提供额外的证据。 动物研究是最先进的临床前研究方法。 已经建立了各种精神疾病的动物模型。 动物研究允许测试机械模型,并在这些模型中测试新药物以减少对人类的潜在危害。 然而,精神病学中动物研究的一个缺点可能在于为不同的精神病症开发合适的动物模型,具体而言,涉及内部或独特的人类认知方面的病症可能导致问题。 因此,显示动物行为改善的新药物可能无助于精神疾病的人类内部心理方面。
因此,本文将研究动物临床前研究和人类精神病学PET研究的结果是否形成了mGluR5参与精神障碍的连贯观点。 本综述将仅讨论mGluR5活性,因为该受体类型可获得广泛的PET人类数据。 mGluR5为药物开发提供了一个有前途的目标,因为PET示踪剂可以测量人体中mGluR5的结合,从而进一步了解其在人体中的功能。 事实上,研究表明,针对代谢型谷氨酸I类受体的药物是目前正在开发用于治疗精神疾病的最有希望的药物之一(Krystal等,2010).
在本综述中,我们首先简要介绍谷氨酸系统和mGluR5,然后通过比较先前的临床前试验和最近的PET研究,评估mGluR5参与焦虑和情绪障碍以及成瘾。 然后,我们将描述一种方法,即基于mGluR5的干预措施如何通过促进学习和社会功能的变化以及降低各脑区域内的兴奋性来提高效率。
谷氨酸能系统和mGluR5
谷氨酸通过离子型和代谢型受体的作用调节中枢神经系统功能。 已经深入研究了谷氨酸在各种精神病和医学病症中的参与。 然而,早期的工作主要集中在离子型谷氨酸受体上。 与离子型受体的快速和直接作用相反,三组代谢型(mGlu)受体通过G蛋白偶联信号传导改变神经元活性。 mGluR组以其药理学和细胞内信号传导特性而着称。 mGluR5,首先在1992中的动物中克隆,几年后在人类中克隆,属于I组代谢型受体(Olive,2005)。 它的行为主要是兴奋的(Meldrum,2000). Cleva和Olive(2011) 描述了mGluR5和NMDA受体之间的强烈联系和受体相互作用,表明mGluR5也可能广泛涉及介导神经可塑性以及学习和记忆过程。 此外,有证据表明mGluR5激活可增强GABA,特别是在伏隔核中(Hoffpauir和Gleason,2002)。 因此,提示代谢型谷氨酸受体活性可以调节兴奋性和抑制性(GABA)信号传导途径。 高mGluR5受体密度主要在前脑区域,纹状体和边缘区域(包括杏仁核和海马区)中被识别(Swanson等,2005)。 利用先进的分子生物学技术确定啮齿动物脑中mGluR5 mRNA的表达,研究确定嗅球,背侧纹状体,伏隔核,侧隔和海马区域的mGluR5表达水平最高(Abe等,1992)(见图 1).
图1
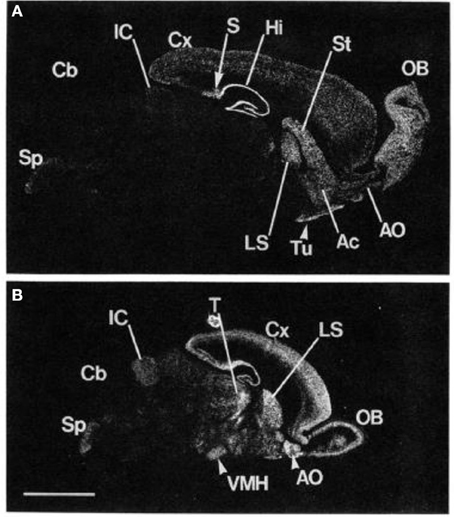
图1。 mGluR5 mRNA在成人和6天大鼠脑中的定位 原位 杂交。 成年大鼠脑的矢状切面的负片图像 (A) 和6天大鼠脑 (B) 显示。 OB,主要嗅核; Ac伏隔核; Tu,嗅结节; St,纹状体; Hi,海马; S,下; Cx,大脑皮质; LS,侧间隔核; IC,下丘; Cb,小脑皮质; Sp,脊柱三叉神经核; T,丘脑; VMH,腹内侧下丘脑核。 比例尺,4 mm。 从 Abe等。 (1992)。 注意:图形及其图例经许可后复制。
发现突触前mGluR5受体参与突触可塑性的调节和神经元兴奋性的变化以维持体内平衡(Schoepp,2001)。 重要的是,在产前5妊娠周(GW)已经可以确定显着的mGluR9表达(Boer等,2010)。 此外,发现年轻动物的mGluR5表达比成人高得多(Romano等,1996),提示针对mGluR5的早期干预可能对神经发育障碍有预防作用。
由于其在不同神经元过程中的功能,谷氨酸系统中的严重异常导致严重的神经损伤(例如,癫痫症),而较小的变化可导致各种精神疾病(Yüksel和Öngür,2010)。 开发有选择性靶向mGluR5的药物的努力始于90后期。 迄今为止,已开发出各种mGluR5激动剂和拮抗剂(参见 Lea和Faden,2006 有关特定药物开发的更多信息)。 例如,3- [2-甲基-1,3-噻唑-4yl]乙炔基]吡啶(MTEP)是一种高选择性和有效的非竞争性mGluR5受体拮抗剂,在注射大剂量1后达到完全受体占有率10 h mg / kg(Anderson等,2003)。 表 1 概述了一些用于动物的mGluR5药物以及之前用于初始临床人体试验的药物。
TABLE 1
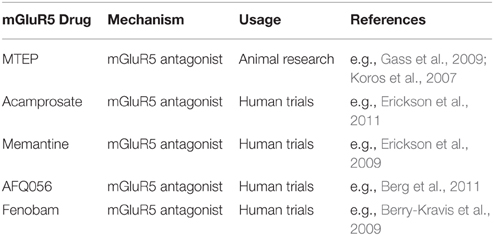
表1。 选择mGluR5作用药物.
精神疾病和基于谷氨酸的人类干预
精神疾病是非常不同的并且共病是常见的。 然而,常见的精神病学药理学治疗基于相对较少的病理生理学机制,例如,增加焦虑和抑郁中的单胺可用性。 因此,迫切需要改进和推进精神病治疗,基于代谢型谷氨酸的药物干预在这方面是一个很有希望的发展(Agid等,2007).
大多数人体试验都是针对脆性X综合征(FXS)的病例进行的。 FXS的mGluR5理论认为缺乏脆弱的X智力迟钝蛋白(FMRP)会通过mGluR5导致过量的谷氨酸能信号传导(Bear等人,2004)。 这导致突触处的局部mRNA翻译增加,因为FMRP不存在以调节该过程。 最后,这会削弱突触并导致更长的未成熟树突棘的数量增加,这可以解释FXS患者中发现的智力残疾。 这种残疾与情绪和焦虑症状有关,并且通常具有自闭症谱系障碍中常见的特征,包括言语和语言发育迟缓,心理理论受损,社交和情感处理受损以及重复行为(Garber等人,2008)。 初步和间接证据表明mGluR5拮抗剂可以改善FXS的社交性(Burket等,2014)希望针对mGluR5的药物可能在与自闭症,精神分裂症和抑郁症等社会过程受损系统相关的普遍精神疾病中临床使用。 此外,FXS中可观察到的重复行为表型可能提示其他精神疾病(如强迫症(OCD)和成瘾)的共同病理生理学。
尽管mGluR5的科学知识和研究有所增加,但药物开发工作相对不成功(Agid等,2007)。 靶向离子型受体的药物通常会产生许多副作用,目前的药物开发策略尚未产生可能减少潜在副作用的离子型受体的选择性靶标。 例如,离子型受体拮抗剂会对人类产生副作用,包括记忆障碍,精神病发作和中风(Swanson等,2005)。 由于离子型谷氨酸受体具有普遍存在的分布,而代谢型受体更不均匀且选择性分布,因此可能出现不利的副作用(Krystal等,2010)。 因此,最近的药物开发集中于靶向代谢型受体的化合物,假设这些药物与那些与快速作用的离子型受体结合的药物相比具有更少的副作用。
在以下部分中,我们将回顾人类PET研究中有关mGluR5参与情绪障碍和成瘾的证据,并将这些发现与动物研究进行比较。 此外,我们将描述人类中mGluR5活性的一些大脑位置,并最终提出一种方法,即直接和间接的mGluR5活性如何参与人类精神病综合征。
mGluR5,病理性焦虑和情绪障碍
病理性焦虑
病理性焦虑发生在焦虑症中,包括广泛性焦虑症,恐慌症(全球最普遍的精神疾病, First和Gibbon,1997),以及其他普遍的精神疾病,如抑郁症和强迫症(OCD)。 这些精神疾病导致社会和职业功能受到严重损害,导致健康成本负担和患者痛苦(First和Gibbon,1997)。 一般来说,焦虑可能与过度的大脑兴奋性有关(哈维和沙希德,2012).
临床前神经生物学研究
来自大量临床前动物试验的结果已经确定了mGluR5拮抗剂治疗在焦虑中的作用。 Swanson等人。 (2005) 回顾了关于针对mGluR5的药物对焦虑样行为的动物研究。 他们得出结论,mGluR5拮抗治疗主要导致实验动物的抗焦虑反应。 特别是,观察到诸如减少恐惧条件冻结,增加的冲击和惩罚接受以及增加的社交互动等效果。 例如,单剂量的2-甲基-6-(苯基乙炔基)吡啶(MPEP)增加了大鼠在实验迷宫的开放臂中所花费的时间,而不影响计划或运动行为(Tatarczyńska等,2001). Krystal等。 (2010) 回顾了在小鼠焦虑模型中检测mGluR5拮抗剂(MTEP,MPEP,fenobam)的临床前动物研究。 这些研究使用不同的结果测量,例如恐惧条件的消退和高架十字迷宫中的反应,以评估药物治疗的有效性。 在所研究的研究中,88.45%报道了mGluR5拮抗剂的抗焦虑作用(Krystal等,2010)。 最近,另一篇关于动物模型焦虑研究的综述,研究了离子型和代谢型谷氨酸受体拮抗剂干预的影响(Riaza Bermudo-Soriano等,2012)。 关于mGluR5,作者列出了43对焦虑的动物研究,除了两种外,其他所有研究均表现出抗焦虑作用。
人类研究
关于焦虑症中谷氨酸功能异常的假设的初步证据来自MRS研究。 例如,使用单体素高场1H磁共振波谱,研究人员发现,与健康对照组相比,患有社交焦虑障碍的患者在前扣带皮层(ACC)中显示出显着更高的谷氨酸水平(Phan等人,2005)。 此外,研究发现社交恐怖症的10患者的全球谷氨酸浓度增加(Pollack等人,2008)。 如前所述,这些研究无法确定哪些谷氨酸受体与谷氨酸过量活性有关。
采用PET研究方法,我们最近首次展示了mGluR5与焦虑症,重度抑郁症(MDD)和强迫症之间的关系。 在一项研究中,我们使用[5C] ABP10 PET研究了OCD和10健康对照的11患者的mGluR688分布容积比(DVR)(Akkus等,2014)。 我们采用耶鲁 - 布朗强迫症量表(Y-BOCS)作为OCD症状严重程度的临床指标。 我们观察到杏仁核,ACC和内侧眶额皮质区域的Y-BOCS观察评分与mGluR5 DVR之间存在显着正相关(Akkus等,2014)。 这些大脑区域以前曾涉及OCD病理生理学。 实际上,杏仁核,ACC和眶额皮质的结构性大脑异常始终与强迫症有关(Rosenberg和Keshavan,1998; Szeszko等人,2008; Van den Heuvel等人,2009)。 鉴于结构成像研究提供了OCD严重程度与灰质体积之间正相关的证据(Zarei等人,2011),OCD中mGluR5结合增加可能反映神经元密度增加。 尽管在DSM-5中,OCD不再被视为焦虑症,但大多数OCD患者会出现焦虑症状。 在一个相对较大的临床样本中,我们之前已经表明,强迫症的强迫症患者焦虑症状和障碍的患病率特别高(Hasler等,2005)。 在本文中,我们认为涉及压力,焦虑或冲突的强迫症可能与杏仁核,ACC和眶额皮质中谷氨酸能神经传递增加有关。
总之,最近使用不同研究方法的研究结果支持了这样一个假设,即在与焦虑相关的情绪障碍中,谷氨酸功能在边缘系统的关键区域是异常的。 异常功能也可能与mGluR5受体有关。 由于临床前以及PET研究显示出一致的结果模式,我们认为拮抗性mGluR5治疗会对患有病理性焦虑症的患者产生显着的抗焦虑作用。
重度抑郁症(MDD)
临床前神经生物学研究
在他们的评论中, Krystal等。 (2010) 描述了在抑郁症动物模型中研究mGluR5拮抗剂MTEP和MPEP的8项研究的结果。 与焦虑症的治疗成功率相比,作者报道只有62.5-75%发现了明显的抗抑郁作用,即使用离子型NMDA拮抗剂氯胺酮治疗也可以导致快速的抗抑郁作用,即使在治疗耐药的患者中也是如此(Pittenger等,2006).
人类研究
最近的一篇综述回顾了心境障碍的13 MRS研究。 作者报告说,这些研究一致发现MDD中谷氨酸减少的证据(Hasler等,2007; Yüksel和Öngür,2010)。 特别是,发现ACC,左背外侧前额叶皮层,背内侧前额叶皮层,腹内侧前额叶皮质,杏仁核和海马中的谷氨酸水平降低。 后来对使用MRS检测MDD中谷氨酸的研究进行了综述,证实ACC中谷氨酸浓度持续降低(Luykx等,2012)。 在枕叶皮质中,一些研究人员发现谷氨酸含量增加,这在MDD患者的忧郁亚组中最高(Sanacora等,2008).
在先前的研究中,我们获得了具有MDD和5健康对照的11未治疗受试者中mGluR11受体结合的PET图像(Deschwanden等,2011)。 我们发现在患有抑郁症的患者的前额皮质,扣带皮层,岛叶,丘脑和海马中区域mGluR5结合减少。 另外,抑郁的严重程度与海马中的mGluR5结合负相关。 我们认为这些发现表明抑郁症中mGluR5神经传递减少,可能是由于谷氨酸系统活性的基础或代偿性变化。 此外,我们检查了5抑郁受试者和15匹配对照的死后脑样本中mGluR15表达的量(Deschwanden等,2011)。 我们观察到从抑郁个体获得的样品中前额皮质中mGluR5表达减少。 据报道,抑郁症患者死后脑中NMDA受体表达降低(Feyissa等,2009)。 实际上,先前已经表明mGluR5拮抗剂的抗抑郁特性可能涉及NMDA受体的抑制。 这可能介导海马神经传递和/或诱导脑源性神经营养因子基因表达(Legutko等人,2006)。 此外,最近的一项研究表明,睡眠剥夺可增加健康人的mGluR5可用性(Hefti等,2013)。 在前扣带回皮层,岛状突,内侧颞叶,海马旁回,纹状体和杏仁核中,这种增加与睡眠剥夺的功效显着相关,如主观嗜睡的增加所反映。 这项研究表明,mGluR5的增加可能是神经生物学机制,可以解释睡眠剥夺具有很高的抗抑郁功效。 临床前研究证实了我们假设的mGluR5与睡眠和抑郁之间的关联。 特别是,一项针对mGluR5基因敲除小鼠的研究提供了重要证据,证明mGluR5参与了NREM睡眠的稳定性-REM睡眠状态转换,NREM慢波活动和对睡眠丧失的稳态反应(Ahnaou等人,2015).
总之,来自动物,死后,MRS和PET研究的趋同证据表明,中央谷氨酸系统重要地参与MDD的病理生理学。 然而,有证据表明mGluR5拮抗作用可能不会直接帮助患有MDD的患者。 这符合我们在MDD患者中减少mGluR5表达的发现。 可以推测,靶向mGluR5系统的药物可能对患有共病焦虑,成瘾障碍和/或昼夜节律受损的抑郁患者特别有用。 此外,社会过程系统中的损伤通常与MDD相关。 不良社交技能似乎是抑郁症的重要危险因素(Segrin,2000)。 这些缺陷包括副语言和语言行为,以及面部表情,凝视,姿势和姿势的损伤,与FXS和自闭症谱系障碍中观察到的相当。 此外,关于社交技能,暴露于慢性社会压力的小鼠实验表明,Homer1 / mGluR5偶联被破坏,表明mGluR5夜间缓解了社交压力的抑郁易感性(Wagner等人,正在报道中)。 此外,在社会缺陷的小鼠模型中,mGluR5抑制导致社会交往正常化(Chung等人,2015)。 总之,这些研究提供了临床前证据,即mGluR5在抑郁症的社会原因和抑郁症患者中经常观察到的社会缺陷中起重要作用。 因此,针对mGluR5的药物可能在预防社会缺陷的青少年抑郁症的发展中发挥重要作用,并可能有助于治疗MDD患者的社会缺陷和低心理社会功能。
mGluR5和成瘾
成瘾的特点是持续的药物摄入,尽管有负面后果,反复尝试停止或减少药物使用,以及耐受和戒断的症状。 虽然多巴胺系统在急性奖励处理中起着关键作用(Kalivas和Volkow,2005),越来越多的证据表明成瘾中的谷氨酸能神经传递(Krystal等,2010).
临床前神经生物学研究
在2001中,发表了关于mGluR5和成瘾的开创性研究(Chiamulera等,2001)。 作者表明,缺乏mGluR5受体的小鼠未能获得静脉内可卡因自我给药,尽管急性注射后伏隔核中的细胞外多巴胺水平增加。 许多后续动物研究表明,mGluR5受体拮抗剂MPEP和MTEP减少了可卡因和尼古丁等成瘾药物的自我管理(Kalivas,2009). 橄榄(2005) 回顾了与药物成瘾有关的动物研究,并提出有证据表明mGluR5可能参与药物使用的发展,奖赏感知和复发,如可卡因,吗啡,尼古丁和乙醇。 作者描述了动物研究中的证据,即mGluR5拮抗剂减少了药物的自我管理,以及随后的药物寻求行为。 例如,最近,在暴食症患者的狒狒模型中发现,MTEP降低糖果消费而不改变寻求糖果的行为(Bisaga等,2008)。 研究表明,与使用mGluR5受体拮抗剂相关的食物摄入减少可能与增强刺激的奖赏价值降低有关(Bisaga等,2008)。 此外,研究报告在实验大鼠中重复使用可卡因后,伏隔核和背外侧纹状体中的mGluR5 mRNA水平显着增加(Bisaga等,2008).
人类研究
在患有酒精性疾病和健康对照的患者的死后脑组织样本中使用放射自显影,研究发现患有酒精性疾病的患者的海马和纹状体中的mNluX-30%mGluR40 / 1结合密度更高(Kupila等,2012)。 这一发现表明,在酒精成瘾患者的某些脑区,mGluR5受体密度可能会增加。 最近,我们使用PET测量尼古丁成瘾患者大脑中mGluR5受体的结合(Akkus等,2013)。 我们发现,与非吸烟者相比,5吸烟者的灰质中mGluR14 DVR全球减少(见图) 2)。 最明显的减少发现在右侧和左侧内侧眶额皮质。 我们认为mGluR5受体结合的这种减少可能是对慢性尼古丁给药诱导的谷氨酸慢性增加的长期适应。 这种适应似乎特定于尼古丁依赖,因为它在慢性非吸烟可卡因使用者中尚未发现(Hulka等,2014).
图2
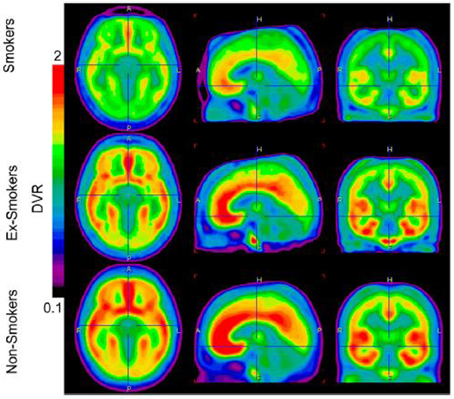
图2。 图像显示三个诊断组中mGluR5 DVR的平均脑摄取。 吸烟者和戒烟者群体的大脑摄取量明显减少。 图像由PMOD软件版本3(PMOD Technologies)计算。 从 Akkus等。 (2013)。 注意:图形及其图例经许可后复制。
实际上,有人提出mGluR5下调代表了代偿性神经适应(Kalivas,2009),药物诱导奖励的增强剂(Rutten等人,2011),或调节条件行为反应中情境线索影响的因素(Tronci等人,2010)。 数字 3 提出了成瘾的mGluR5功能障碍的方法。
图3
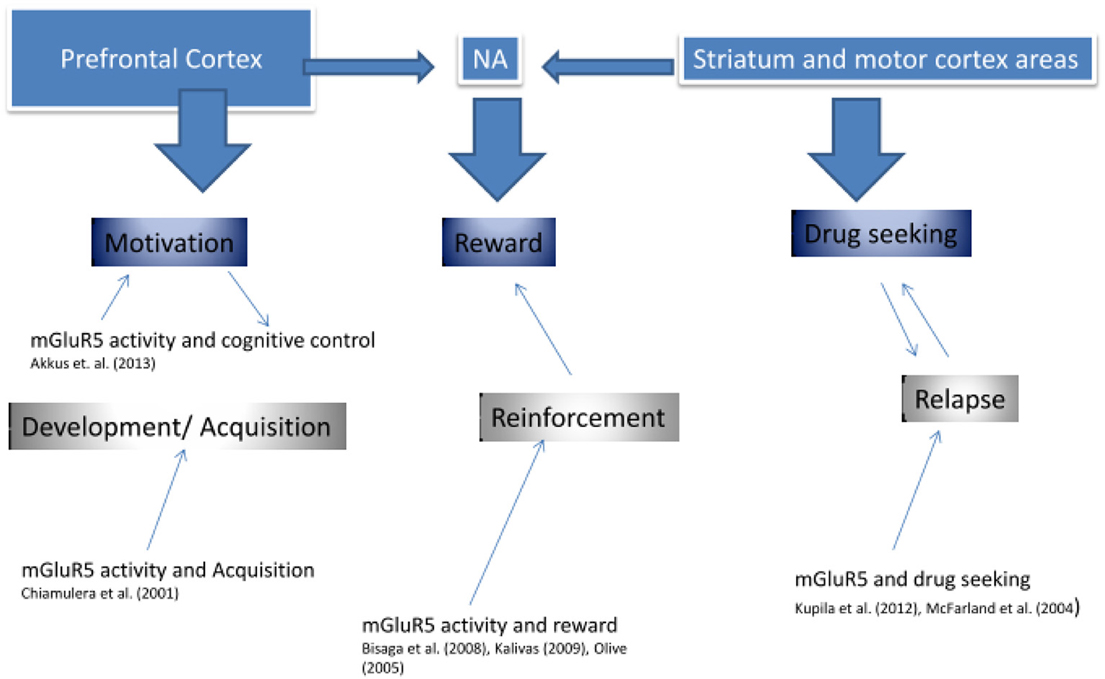
图3。 mGluR5参与成瘾.
如图所示 3以前的研究支持mGluR5涉及成瘾的三个关键阶段,开发和获取,药物强化价值以及成瘾复发的观点。 可能有人提出,每个功能主要由不同的大脑区域描述,这些大脑区域也显示出高的mGluR5受体密度,并且已经显示在我们的PET研究中显示出降低的mGluR5密度(Deschwanden等,2011). Kalivas(2009) 开发了一种广泛的谷氨酸成瘾模型。 作者认为,成瘾与皮质纹状体脑回路的关键区域(包括杏仁核,伏隔核(NA),前额皮质和运动皮层)之间的谷氨酸稳态功能障碍有关。 正如我们在图中所示 3首先(“动机成分”),前额皮质可能在药物成瘾发展的初始阶段调解动机和认知控制。 临床前研究 Chiamulera等。 (2001) 表明mGluR5是通过调解滥用药物的有益特性在成瘾发展的这个阶段所必需的。 其次(“奖励成分”),Nucleus Accumbens(NA)已被证明可以影响药物的奖赏价值,不仅通过多巴胺,还通过mGluR5活性(Bisaga等,2008)。 特别是,提出了前肢皮质和NA之间谷氨酸传递的损害,因此通过更加依赖与NMDA / mGluR5受体系统相关的学习行为来启动寻药(Kalivas,2009)。 第三,(“药物寻求组件”),一些研究支持mGluR5更多地依赖于运动过程 - 减少认知控制 - 通过纹状体(Kupila等,2012)。 我们自己发表的和未发表的研究表明,mGluR5下调与戒烟者复发风险增加有关(Akkus,PNAS)。 这种下调可能是致病性或补偿性变化不足。 目前的研究表明,针对mGluR5的药物可以改善成瘾疾病在其发展的各个阶段的治疗。
讨论
回顾的证据表明mGluR5在焦虑症,强迫症,MDD以及成瘾中的高度参与,并且用mGluR5靶向药物治疗也可能对人类有益。 然而,与强迫症相关的情绪和焦虑症状的发病机制可能与与强迫症无关的情绪和焦虑症状不同。 因此,强迫症状的存在可能是针对mGluR5的药物的抗抑郁药和抗焦虑药反应的重要预测因子。 此外,我们建议针对mGluR5系统的药物可能有助于增加对社会压力的抵抗力并改善抑郁症的社会缺陷。 由于社交压力是迄今为止抑郁症和社会缺陷最重要的非遗传风险因素,与社会功能和生活质量的降低密切相关,动物研究的这些发现具有很高的科学和临床相关性。
最近,国家精神卫生研究所(NIMH)发起了一项新颖的,最先进的项目:研究领域标准(RDoC)。 这反映了NIMH战略1.4的实施“为了研究目的,开发基于可观察行为和神经生物学测量的维度对精神障碍进行分类的新方法。”(http://www.nimh.nih.gov/research-priorities/strategic-objectives/strategic-objective-1.shtml)。 这一举措是重组严格的DSM类别的结果,其中大部分也是在神经科学研究之前开发的(Morris和Cuthbert,2012)。 RDoC描述了五个域或结构; 负价系统,正价系统,认知系统,社会过程系统,唤醒/监管系统。 根据有关mGluR5参与MDD和OCD的审查证据,我们提出mGluR5活动可能与负价系统直接相关,负价系统涉及恐惧,威胁,持续威胁,损失和沮丧的非可观察因素。奖励。 因此,正如所评论的研究表明,用mGluR5进行拮抗治疗可以减少那些可观察到的症状。 此外,我们建议mGluR5治疗也可能有利于“阳性价值障碍”,如成瘾和抑郁,通过脑结构的异常mGluR5活性和与mGluR5功能性相关的谷氨酸能NMDA受体相关的功能,并且重要地参与奖励学习。 例如, Simonyi等人。 (2010) 回顾了许多在敲除小鼠中使用mGluR5受体拮抗剂的动物研究,以确定mGluR5在学习和记忆中的作用。 抑制性学习,例如被动回避学习,是用于研究海马学习过程的动物模型中的一项成熟任务,并且已在许多研究中显示依赖于mGluR5受体(Simonyi等,2010)。 例如,研究表明,CA5中mGluR3蛋白在短期和CA1长期增强过程中过度表达(Riedel等,2000). 海曼(2005) 提出了一种成瘾的生物学模型,它结合了学习和记忆的异常神经过程,形成了成瘾的基本要素。 作者提出长期增强作用,包括谷氨酸受体可用性的改变,以及基因表达的调节作为与药物成瘾相关的异常回路中发现的药物诱导的改变的潜在重要机制。 最后,关于mGluR5和睡眠稳态的研究(Hefti等,2013; Ahnaou等人,2015)表明mGluR5在RDoC的唤醒和调节系统域中起着重要作用。
数字 4 总结了用于调解mGluR5在情绪障碍和成瘾中的作用的过程。 图的顶部 4 描绘了高密度mGluR5大脑区域; 杏仁核,海马,纹状体,NA和前额皮质。 精神病综合症与这些地区相匹配。 杏仁核中建议的mGluR5活动可能介导原发性情绪唤起,例如焦虑和抑郁。 我们已经描述了两种可能的途径; 社会功能和学习可能会调解其他过程。 因此,mGluR5活动已经显示出参与学习,因此,通过海马,NA和纹状体的活动可能参与记忆和奖励,认知控制和动机,也涉及成瘾(参见图 3)。 最后,我们建议mGluR5如何与社会压力反应和社会缺陷相关,这可能与广泛的精神疾病相关。 表 2 概述了RDoC损伤区域,相关的临床表现以及mGluR5的参与情况。
图4
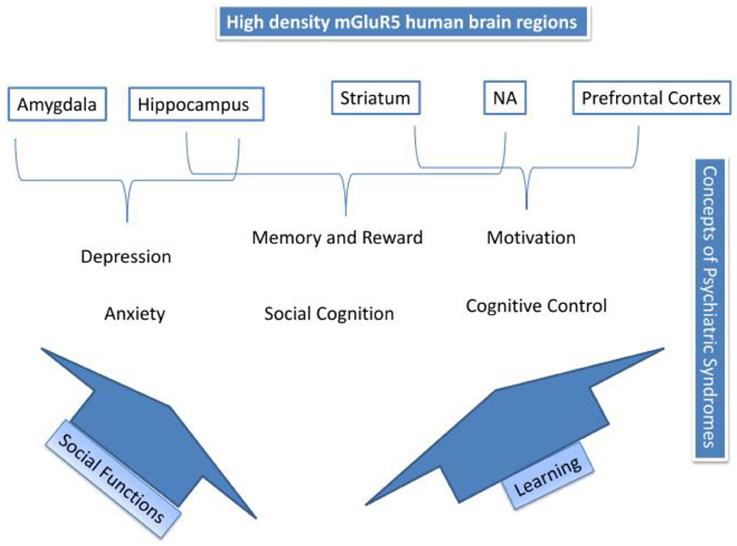
图4。 mGluR5机制摘要.
TABLE 2
![]()
表2。 RDoC和mGluR5.
结束语
该综述描述了mGluR5参与情绪障碍,强迫症和成瘾,并将临床前与人体研究,特别是PET研究进行了比较。 因此,我们比较了不同的方法学方法,如动物研究,MRS和PET研究。 我们建议,如果在人类临床试验(RDoC)中开始mGluR5拮抗治疗,则会出现强烈的直接抗焦虑作用。 在FXS中也报道了mGluR5过度活动,其特征在于重要的社会缺陷。 因此,mGluR5活动可能不仅使负价系统和唤醒系统中的活动正常化,而且还减弱了社会过程系统(RDoC)中的损害。 这具有很高的临床重要性,因为不良的社会功能是诸如强迫症,抑郁症和成瘾等普遍的精神疾病的重要结果,导致巨大的个人痛苦和社会的重要间接成本。 最后,mGluR5也被证明具有显着的药物成瘾作用,主要是由于增加药物的奖励价值。 因此,mGluR5拮抗干预对于治疗病理性焦虑和成瘾以及改善社会压力弹性和社会功能最有效。
利益冲突声明
Terbeck博士,Chesterman博士和Akkus博士没有利益冲突。 Hasler博士已从Novartis获得资助,该公司正在生产和测试针对mGluR5的药物。
参考资料
Abe,T.,Sugihara,H.,Nawa,H.,Shigemoto,R.,Mizuno,N。和Nakanishi,S。(1992)。 新型代谢型谷氨酸受体mGluR5与肌醇磷酸/ Ca2 +信号转导偶联的分子特征。 J.Biol.Chem。 267,13361-13368。
PubMed摘要 | 全文 | Google Scholar
Agid,Y.,Buzsáki,G.,Diamond,DM,Frackowiak,R.,Giedd,J.,Girault,J.-A。,et al。 (2007)。 如何改善精神疾病的药物发现? 纳特。 Rev. Drug Discov。 6,189-201。 doi:10.1038 / nrd2217
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Akkus,F.,Ametamey,SM,Treyer,V.,Burger,C.,Johayem,A.,Umbricht,D.,et al。 (2013)。 通过[5C] ABP11正电子发射断层扫描确定吸烟者和戒烟者中mGluR688受体结合的显着全球减少。 PROC。 国家科。 科学院。 科学。 美国。 110,737-742。 doi:10.1073 / pnas.1210984110
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Akkus,F.,Terbeck,S.,Ametamey,SM,Rufer,MD,Treyer,V.,Burger,C.,et al。 (2014)。 代谢性谷氨酸受体5与强迫症患者的结合。 诠释。 J. Neuropsychopharmacol。 17,1915-1922。 doi:10.1017 / S1461145714000716
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Ametamey,SM,Kessler,LJ,Honer,M.,Wyss,MT,Buck,A.,Hintermann,S.,et al。 (2006)。 11CABP688的放射合成和临床前评估作为成像代谢型谷氨酸受体亚型5的探针。 J. Nucl。 地中海。 47,698-705。
PubMed摘要 | 全文 | Google Scholar
Anderson,JJ,Bradbury,MJ,Giracello,DR,Chapman,DF,Holtz,G.,Roppe,J.,et al。 (2003)。 体内 使用新型放射性配体[5 H] 3-甲氧基-3-(吡啶-5-基乙炔基)吡啶)对mGlu2受体拮抗剂的受体占有率。 欧元。 J. Pharmacol. 473, 35–40. doi: 10.1016/S0014-2999(03)01935-6
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Ahnaou,A.,Raeymaekers,L.,Steckler,T。和Drinkenbrug,WHIM(2015)。 代谢型谷氨酸受体(mGluR5)在调节NREM-REM睡眠周期和体内平衡中的相关性:来自mGluR5( - / - )小鼠的证据。 Behav。 Brain Res。 282,218-226。 doi:10.1016 / j.bbr.2015.01.009
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Bear,MF,Huber,KM和Warren,ST(2004)。 脆性X智力低下的mGluR理论。 趋势神经科学。 27,370-377。 doi:10.1016 / j.tins.2004.04.009
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Berg,D.,Godau,J.,Trenkwalder,C.,Eggert,K.,Csoti,I.,Storch,A.,et al。 (2011)。 AFQ056治疗左旋多巴诱发的运动障碍:2随机对照试验的结果。 乐章。 Disord。 26,1243-1250。 doi:10.1002 / mds.23616
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Berry-Kravis,E.,Hessl,D.,Coffey,S.,Hervey,C.,Schneider,A.,Yuhas,J.,et al。 (2009)。 一项试验性开放标签,单剂量试验的fenobam成人脆性X综合征。 J. Med。 遗传学。 46,266-271。 doi:10.1136 / jmg.2008.063701
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Bisaga,A.,Danysz,W。和Foltin,RW(2008)。 谷氨酸能NMDA和mGluR5受体的拮抗作用降低了酗酒疾病的狒狒模型中的食物消耗。 欧元。 Neuropsychopharmacol。 18,794-802。 doi:10.1016 / j.euroneuro.2008.05.004
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Boer,K.,Encha-Razavi,F.,Sinico,M。和Aronica,E。(2010)。 第I组代谢型谷氨酸受体在发育中的人皮质中的差异分布。 脑;。 1324,24-33。 doi:10.1016 / j.brainres.2010.02.005
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Brennan,BP,Rauch,SL,Jensen,JE和Pope,HG(2012)。 对强迫症的磁共振波谱研究的批评性综述。 生物学。 精神病学 73,31-24。 doi:10.1016 / j.biopsych.2012.06.023
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Burket,JA,Benson,AD,Tang,AH和Deutsch,SI(2014)。 雷帕霉素改善了自闭症谱系障碍的BTBR小鼠模型的社交性。 Brain Res。 公牛。 100,70-75。 doi:10.1016 / j.brainresbull.2013.11.005
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Chiamulera,C.,Epping-Jordan,MP,Zocchi,A.,Marcon,C.,Cottiny,C.,Tacconi,S.,et al。 (2001)。 在mGluR5无效突变小鼠中不存在可卡因的增强和运动刺激作用。 纳特。 神经科学。 4,873-874。 doi:10.1038 / nn0901-873
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Chung,W.,Choi,SY,Lee,E.,Park,H.,Kang,J.,Park,H.,et al。 (2015)。 通过NMDAR和mGluR53抑制改善IRSp5突变小鼠的社交缺陷。 纳特。 神经科学。 18,435-442。 doi:10.1038 / nn.3927
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Cleva,RM和Olive,MF(2011)。 5型代谢型谷氨酸受体(mGluR5)的正变构调节剂及其治疗CNS疾病的治疗潜力。 分子 16,2097-2106。 doi:10.3390 / molecules16032097
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Deschwanden,A.,Karolewicz,B.,Feyissa,AM,Treyer,V.,Ametamey,SM,Johayem,A.,et al。 (2011)。 通过[(5)C] ABP11 PET和死后研究确定的重度抑郁症中代谢型谷氨酸受体688密度降低。 上午。 J.精神病学 168,727-734。 doi:10.1176 / appi.ajp.2011.09111607
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Erickson,CA,Mullett,JE和McDougle,CJ(2009)。 脆性X综合征中的开放标签美金刚。 J.自闭症发展 Disord。 39,1629-1635。 doi:10.1007 / s10803-009-0807-3
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Erickson,CA,Early,M.,Stigler,KA,Wink,LK,Mullett,JE和McDougle,CJ(2011)。 一项关于自闭症青少年阿坎酸的开放标签自然主义试验研究。 J. Child Adolesc。 精神药理学。 21,565-569。 doi:10.1089 / cap.2011.0034
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
首先,MB和Gibbon,M。(1997)。 DSM-IV轴I疾病结构化临床访谈用户指南SCID-I:临床医生版本。 纽约州纽约市:American Psychiatric Pub。
Feyissa,AM,Chandran,A.,Stockmeier,CA和Karolewicz,B。(2009)。 在重度抑郁症的前额叶皮质中,NMDA受体和PSD-2的NR2A和NR95B亚基水平降低。 PROG。 Neuropsychopharmacol。 生物学。 精神病学 33,70-75。 doi:10.1016 / j.pnpbp.2008.10.005
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Garber,KB,Visootsak,J。和Warren,ST(2008)。 脆性X综合症。 欧元。 J. Hum。 遗传学。 16,666-672。 doi:10.1038 / ejhg.2008.61
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Gass,JT,Osborne,MP,Watson,NL,Brown,JL和Olive,MF(2009)。 mGluR5拮抗作用减弱甲基苯丙胺的强化,并阻止大鼠恢复寻求甲基苯丙胺的行为。 神经精神药理学。 34,820-833。 doi:10.1038 / npp.2008.140
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Harvey,BH和Shahid,M。(2012)。 代谢和离子型谷氨酸受体作为焦虑和压力相关疾病的神经生物学目标:专注于药理学和临床前转化模型。 药理学。 生物化学。 Behav。 100,775-800。 doi:10.1016 / j.pbb.2011.06.014
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Hasler,G.,LaSalle-Ricci,VH,Ronquillo,JG,Crawley,SA,Cochran,LW,Kazuba,D.,et al。 (2005)。 强迫症症状维度显示与精神病合并症的特定关系。 精神病;。 135,121-132。 doi:10.1016 / j.psychres.2005.03.003
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Hasler,G.,van der Veen,JW,Tumonis,T.,Meyers,N.,Shen,J.,and Drevets,WC(2007)。 使用质子磁共振波谱法测定的主要抑郁症中的前额谷氨酸/谷氨酰胺和γ-氨基丁酸水平降低。 拱。 精神病学 64,193-200。 doi:10.1001 / archpsyc.64.2.193
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Hasler,G.,van der Veen,JW,Geraci,M.,Shen,J.,Pine,D。和Drevets,WC(2009)。 通过质子磁共振波谱法确定惊恐障碍中的前额皮质γ-氨基丁酸水平。 生物学。 精神病学 65,273-275。 doi:10.1016 / j.biopsych.2008.06.023
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Hefti,K.,Holst,SC,Sovago,J.,Bachmann,V.,Buck,A.,Ametamey,SM,et al。 (2013)。 在一夜无睡眠后,人脑中的代谢型谷氨酸受体亚型5可用性增加。 生物学。 精神病学 73,161-168。 doi:10.1016 / j.biopsych.2012.07.030
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Hyman,SE(2005)。 成瘾:学习和记忆的疾病。 上午。 J.精神病学 162,1414-1422。 doi:10.1176 / appi.ajp.162.8.1414
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Hoffpauir,BK和Gleason,EL(2002)。 mGluR5的激活调节视网膜无长突细胞中的GABAA受体功能。 J.神经生理学。 88,1766-1776。
PubMed摘要 | 全文 | Google Scholar
Hulka,LM,Treyer,V.,Scheidegger,M.,Preller,KH,Vonmoos,M.,Baumgartner,MR,et al。 (2014)。 吸烟但不吸食可卡因与人类中较低的脑代谢型谷氨酸受体5密度相关。 摩尔。 精神病学 19,625-632。 doi:10.1038 / mp.2013.51
Kalivas,PW和Volkow,ND(2005)。 成瘾的神经基础:动机和选择的病理学。 上午。 J.精神病学 162,1403-1413。 doi:10.1176 / appi.ajp.162.8.1403
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Kalivas,PW(2009)。 成瘾的谷氨酸稳态假说。 纳特。 Rev. Neurosci。 10,561-572。 doi:10.1038 / nrn2515
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Krystal,JH,Mathew,SJ,D'Souza,DC,Garakani,A.,Gunduz-Bruce,H.和Charney,DS(2010)。 代谢型谷氨酸受体激动剂和拮抗剂的潜在精神病学应用。 CNS药物 24, 669–693. doi: 10.2165/11533230-000000000-00000
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Kupila,J.,Kärkkäinen,O.,Laukkanen,V.,Tupala,E.,Tiihonen,J。和Storvik,M。(2012)。 酒精受试者大脑中的mGluR1 / 5受体密度:整个半球放射自显影研究。 精神病;。 12,149-155。 doi:10.1016 / j.pscychresns.2012.04.003
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Koros,E.,Rosenbrock,H.,Birk,G.,Weiss,C。和Sams-Dodd,F。(2007)。 选择性mGlu5受体拮抗剂MTEP,类似于NMDA受体拮抗剂,诱导大鼠的社会隔离。 神经精神药理学 32,562-576。 doi:10.1038 / sj.npp.1301133
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Lea,PM和Faden,AI(2006)。 代谢型谷氨酸受体亚型5拮抗剂MPEP和MTEP。 CNS药物Rev。 12,149-166。 doi:10.1111 / j.1527-3458.2006.00149.x
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Legutko,B.,Szewczyk,B.,Pomierny-Chamioło,L.,Nowak,G。和Pilc,A。(2006)。 MPEP处理对脑源性神经营养因子基因表达的影响。 药理学。 众议员。 58,427-430。
PubMed摘要 | 全文 | Google Scholar
Luykx,JJ,Laban,KG,van den Heuvel,MP,Boks,MPM,Mandl,RCW,Kahn,RS,et al。 (2012)。 重度抑郁症的区域和状态特异性谷氨酸下调:(1)H-MRS发现的荟萃分析。 神经科学。 Biobehav。 启。 36,198-205。 doi:10.1016 / j.neubiorev.2011.05.014
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Meldrum,BS(2000)。 谷氨酸作为大脑中的神经递质:生理学和病理学的回顾。 J. Nutr。 130,1007。
PubMed摘要 | 全文 | Google Scholar
Morris,SE和Cuthbert,BN(2012)。 研究领域标准:认知系统,神经回路和行为维度。 对话Clin。 神经科学。 14,29-37。
PubMed摘要 | 全文 | Google Scholar
Olive,MF(2005)。 mGlu5受体:神经解剖学,药理学和药物成瘾中的作用。 CURR。 精神病学牧师。 1,197-214。 doi:10.2174 / 1573400054065578
Phan,KL,Fitzgerald,DA,Cortese,BM,Seraji-Bozorgzad,N.,Tancer,ME和Moore,GJ(2005)。 社交焦虑障碍中的前扣带神经化学:1特斯拉的4H-MRS。 Neuroreport 16, 183–186. doi: 10.1097/00001756-200502080-00024
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Pittenger,C.,Krystal,JH和Coric,V。(2006)。 谷氨酸调节药物作为治疗强迫症的新型药物治疗剂。 NeuroRx 3,69-81。 doi:10.1016 / j.nurx.2005.12.006
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Pollack,MH,Jensen,JE,Simon,NM,Kaufmann,RE和Renshaw,PF(2008)。 社交焦虑症中GABA,谷氨酸和谷氨酰胺的高场MRS研究?:对左乙拉西坦治疗的反应。 PROG。 Neuropsychopharmacol。 生物学。 精神病学 32,739-743。 doi:10.1016 / j.pnpbp.2007.11.023
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Riaza Bermudo-Soriano,C.,Perez-Rodriguez,MM,Vaquero-Lorenzo,C。和Baca-Garcia,E。(2012)。 谷氨酸和焦虑的新视角。 药理学。 生物化学。 Behav。 100,752-774。 doi:10.1016 / j.pbb.2011.04.010
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Riedel,G.,Casabona,G.,Platt,B.,Macphail,EM和Nicoletti,F。(2000)。 恐惧条件诱导的大鼠海马中mGlu5受体蛋白表达的时间和亚区特异性增加。 神经药理学 39, 1943–1951. doi: 10.1016/S0028-3908(00)00037-X
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Rutten,K.,De Vry,J.,Bruckmann,W。和Tzschentke,TM(2011)。 NOP受体的药理学阻断或基因敲除增强了吗啡在大鼠中的奖赏效果。 药物酒精依赖。 114,253-256。 doi:10.1016 / j.drugalcdep.2010.10.004
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Romano,C.,van den Pol,AN和O'Malley,KL(1996)。 增强的代谢型谷氨酸受体mGluR5在大鼠脑中的早期发育表达:蛋白质,mRNA剪接变体和区域分布。 J.比较。 神经病学。 367,403-412。
Rosenberg,DR和Keshavan,MS(1998)。 走向强迫症的神经发育模型。 生物学。 精神病学 43, 623–640. doi: 10.1016/S0006-3223(97)00443-5
Sanacora,G.,Zarate,CA,Krystal,JH和Manji,HK(2008)。 针对谷氨酸能系统开发新的,改进的情绪障碍治疗方法。 纳特。 Rev. Drug Discov。 7,426-437。 doi:10.1038 / nrd2462
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Schoepp,DD(2001)。 揭示中枢神经系统中突触前代谢型谷氨酸受体的功能。 J. Pharmacol。 进出口。 疗法。 299,12-20。
PubMed摘要 | 全文 | Google Scholar
Segrin,C。(2000)。 与抑郁症相关的社交技能缺陷。 临床。 心理学。 启. 20, 379–403. doi: 10.1016/S0272-7358(98)00104-4
Simonyi,A.,Schachtman,TR和Christoffersen,GR(2010)。 代谢型谷氨酸受体亚型5在学习和记忆中的拮抗作用。 欧元。 J. Pharmacol。 639,17-25。 doi:10.1016 / j.ejphar.2009.12.039
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Soares,DP和Law,M。(2009)。 大脑的磁共振波谱:代谢物和临床应用的回顾。 临床。 放射学杂志。 64,12-21。 doi:10.1016 / j.crad.2008.07.002
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Swanson,CJ,Bures,M.,Johnson,MP,Linden,A.-M.,Monn,JA和Schoepp,DD(2005)。 代谢型谷氨酸受体作为焦虑和应激障碍的新靶点。 纳特。 Rev. Drug Discov。 4,131-144。 doi:10.1038 / nrd1630
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Szeszko,PR,Christian,C.,Macmaster,F.,Lencz,T.,Mirza,Y.,Taormina,SP,et al。 (2008)。 精神药物 - 幼稚小儿强迫症的灰质结构改变:优化的基于体素的形态测量研究。 上午。 J.精神病学 165,1299-1307。 doi:10.1176 / appi.ajp.2008.08010033
Tatarczyńska,E.,Klodzińska,A.,Chojnacka-Wójcik,E.,Palucha,A.,Gasparini,F.,Kuhn,R.,et al。 (2001)。 MPEP的潜在抗焦虑和抗抑郁样作用,MPEP是一种有效的,选择性和系统活性的mGlu5受体拮抗剂。 BR。 J. Pharmacol。 132,1423-1430。 doi:10.1038 / sj.bjp.0703923
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Tronci,V.,Vronskaya,S.,Montgomery,N.,Mura,D。和Balfour,DJK(2010)。 mGluR5受体拮抗剂6-甲基-2-(苯基乙炔基) - 吡啶(MPEP)对尼古丁的行为反应的影响。 精神药理学 211, 33–42. doi: 10.1007/s00213-010-1868-x
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Van den Heuvel,OA,Remijnse,PL,Mataix-Cols,D.,Vrenken,H.,Groenewegen,HJ,Uylings,HBM,et al。 (2009)。 强迫症的主要症状维度是由部分不同的神经系统介导的。 大脑 132,853-868。 doi:10.1093 / brain / awn267
Wagner,KV,Hartmann,J.,Labermaier,C.,Häusl,AS,Zhao,G.,Harbich,D.,et al。 (在新闻)。 Homer1 / mGluR5活动缓解了长期社会压力的脆弱性。 神经精神药理学。 doi:10.1038 / npp.2014.308
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Yüksel,C。和Öngür,D。(2010)。 磁共振波谱研究谷氨酸相关的情绪障碍异常。 生物学。 精神病学 68,785-794。 doi:10.1016 / j.biopsych.2010.06.016
Zarei,M.,Mataix-Cols,D.,Heyman,I.,Hough,M.,Doherty,J.,Burge,L.,et al。 (2011)。 强迫症青少年灰质体积和白质微观结构的变化。 生物学。 精神病学 70,1083-1090。 doi:10.1016 / j.biopsych.2011.06.032
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
关键词:mGluR5,PET,情绪障碍,成瘾,焦虑
引用:Terbeck S,Akkus F,Chesterman LP和Hasler G(2015)代谢型谷氨酸受体5在情绪障碍和成瘾的发病机制中的作用:将临床前证据与人体正电子发射断层扫描(PET)研究相结合。 面前。 神经科学. 9:86。 doi:10.3389 / fnins.2015.00086
收到:07 1月2015; 接受:27二月2015;
发布时间:18 March 2015。
编辑:
阿肖克·库马尔,美国佛罗里达大学
点评人:
Karthik Bodhinathan,Sanford Burnham医学研究所,美国
Amber M. Muehlmann,美国佛罗里达大学
版权所有©2015 Terbeck,Akkus,Chesterman和Hasler。 这是一份根据条款分发的开放获取文章 知识共享署名许可(CC BY)。 允许在其他论坛中使用,分发或复制,前提是原始作者或许可人被记入贷方,并且根据公认的学术惯例引用本期刊中的原始出版物。 不允许使用,分发或复制,不符合这些条款。
*通讯:英国普利茅斯PL4 8AA,普利茅斯大学健康与人文科学学院心理学院Sylvia Terbeck [电子邮件保护]
精神疾病和基于谷氨酸的人类干预
精神疾病是非常不同的并且共病是常见的。 然而,常见的精神病学药理学治疗基于相对较少的病理生理学机制,例如,增加焦虑和抑郁中的单胺可用性。 因此,迫切需要改进和推进精神病治疗,基于代谢型谷氨酸的药物干预在这方面是一个很有希望的发展(Agid等,2007).
大多数人体试验都是针对脆性X综合征(FXS)的病例进行的。 FXS的mGluR5理论认为缺乏脆弱的X智力迟钝蛋白(FMRP)会通过mGluR5导致过量的谷氨酸能信号传导(Bear等人,2004)。 这导致突触处的局部mRNA翻译增加,因为FMRP不存在以调节该过程。 最后,这会削弱突触并导致更长的未成熟树突棘的数量增加,这可以解释FXS患者中发现的智力残疾。 这种残疾与情绪和焦虑症状有关,并且通常具有自闭症谱系障碍中常见的特征,包括言语和语言发育迟缓,心理理论受损,社交和情感处理受损以及重复行为(Garber等人,2008)。 初步和间接证据表明mGluR5拮抗剂可以改善FXS的社交性(Burket等,2014)希望针对mGluR5的药物可能在与自闭症,精神分裂症和抑郁症等社会过程受损系统相关的普遍精神疾病中临床使用。 此外,FXS中可观察到的重复行为表型可能提示其他精神疾病(如强迫症(OCD)和成瘾)的共同病理生理学。
尽管mGluR5的科学知识和研究有所增加,但药物开发工作相对不成功(Agid等,2007)。 靶向离子型受体的药物通常会产生许多副作用,目前的药物开发策略尚未产生可能减少潜在副作用的离子型受体的选择性靶标。 例如,离子型受体拮抗剂会对人类产生副作用,包括记忆障碍,精神病发作和中风(Swanson等,2005)。 由于离子型谷氨酸受体具有普遍存在的分布,而代谢型受体更不均匀且选择性分布,因此可能出现不利的副作用(Krystal等,2010)。 因此,最近的药物开发集中于靶向代谢型受体的化合物,假设这些药物与那些与快速作用的离子型受体结合的药物相比具有更少的副作用。
在以下部分中,我们将回顾人类PET研究中有关mGluR5参与情绪障碍和成瘾的证据,并将这些发现与动物研究进行比较。 此外,我们将描述人类中mGluR5活性的一些大脑位置,并最终提出一种方法,即直接和间接的mGluR5活性如何参与人类精神病综合征。
mGluR5,病理性焦虑和情绪障碍
病理性焦虑
病理性焦虑发生在焦虑症中,包括广泛性焦虑症,恐慌症(全球最普遍的精神疾病, First和Gibbon,1997),以及其他普遍的精神疾病,如抑郁症和强迫症(OCD)。 这些精神疾病导致社会和职业功能受到严重损害,导致健康成本负担和患者痛苦(First和Gibbon,1997)。 一般来说,焦虑可能与过度的大脑兴奋性有关(哈维和沙希德,2012).
临床前神经生物学研究
来自大量临床前动物试验的结果已经确定了mGluR5拮抗剂治疗在焦虑中的作用。 Swanson等人。 (2005) 回顾了关于针对mGluR5的药物对焦虑样行为的动物研究。 他们得出结论,mGluR5拮抗治疗主要导致实验动物的抗焦虑反应。 特别是,观察到诸如减少恐惧条件冻结,增加的冲击和惩罚接受以及增加的社交互动等效果。 例如,单剂量的2-甲基-6-(苯基乙炔基)吡啶(MPEP)增加了大鼠在实验迷宫的开放臂中所花费的时间,而不影响计划或运动行为(Tatarczyńska等,2001). Krystal等。 (2010) 回顾了在小鼠焦虑模型中检测mGluR5拮抗剂(MTEP,MPEP,fenobam)的临床前动物研究。 这些研究使用不同的结果测量,例如恐惧条件的消退和高架十字迷宫中的反应,以评估药物治疗的有效性。 在所研究的研究中,88.45%报道了mGluR5拮抗剂的抗焦虑作用(Krystal等,2010)。 最近,另一篇关于动物模型焦虑研究的综述,研究了离子型和代谢型谷氨酸受体拮抗剂干预的影响(Riaza Bermudo-Soriano等,2012)。 关于mGluR5,作者列出了43对焦虑的动物研究,除了两种外,其他所有研究均表现出抗焦虑作用。
人类研究
关于焦虑症中谷氨酸功能异常的假设的初步证据来自MRS研究。 例如,使用单体素高场1H磁共振波谱,研究人员发现,与健康对照组相比,患有社交焦虑障碍的患者在前扣带皮层(ACC)中显示出显着更高的谷氨酸水平(Phan等人,2005)。 此外,研究发现社交恐怖症的10患者的全球谷氨酸浓度增加(Pollack等人,2008)。 如前所述,这些研究无法确定哪些谷氨酸受体与谷氨酸过量活性有关。
采用PET研究方法,我们最近首次展示了mGluR5与焦虑症,重度抑郁症(MDD)和强迫症之间的关系。 在一项研究中,我们使用[5C] ABP10 PET研究了OCD和10健康对照的11患者的mGluR688分布容积比(DVR)(Akkus等,2014)。 我们采用耶鲁 - 布朗强迫症量表(Y-BOCS)作为OCD症状严重程度的临床指标。 我们观察到杏仁核,ACC和内侧眶额皮质区域的Y-BOCS观察评分与mGluR5 DVR之间存在显着正相关(Akkus等,2014)。 这些大脑区域以前曾涉及OCD病理生理学。 实际上,杏仁核,ACC和眶额皮质的结构性大脑异常始终与强迫症有关(Rosenberg和Keshavan,1998; Szeszko等人,2008; Van den Heuvel等人,2009)。 鉴于结构成像研究提供了OCD严重程度与灰质体积之间正相关的证据(Zarei等人,2011),OCD中mGluR5结合增加可能反映神经元密度增加。 尽管在DSM-5中,OCD不再被视为焦虑症,但大多数OCD患者会出现焦虑症状。 在一个相对较大的临床样本中,我们之前已经表明,强迫症的强迫症患者焦虑症状和障碍的患病率特别高(Hasler等,2005)。 在本文中,我们认为涉及压力,焦虑或冲突的强迫症可能与杏仁核,ACC和眶额皮质中谷氨酸能神经传递增加有关。
总之,最近使用不同研究方法的研究结果支持了这样一个假设,即在与焦虑相关的情绪障碍中,谷氨酸功能在边缘系统的关键区域是异常的。 异常功能也可能与mGluR5受体有关。 由于临床前以及PET研究显示出一致的结果模式,我们认为拮抗性mGluR5治疗会对患有病理性焦虑症的患者产生显着的抗焦虑作用。
重度抑郁症(MDD)
临床前神经生物学研究
在他们的评论中, Krystal等。 (2010) 描述了在抑郁症动物模型中研究mGluR5拮抗剂MTEP和MPEP的8项研究的结果。 与焦虑症的治疗成功率相比,作者报道只有62.5-75%发现了明显的抗抑郁作用,即使用离子型NMDA拮抗剂氯胺酮治疗也可以导致快速的抗抑郁作用,即使在治疗耐药的患者中也是如此(Pittenger等,2006).
人类研究
最近的一篇综述回顾了心境障碍的13 MRS研究。 作者报告说,这些研究一致发现MDD中谷氨酸减少的证据(Hasler等,2007; Yüksel和Öngür,2010)。 特别是,发现ACC,左背外侧前额叶皮层,背内侧前额叶皮层,腹内侧前额叶皮质,杏仁核和海马中的谷氨酸水平降低。 后来对使用MRS检测MDD中谷氨酸的研究进行了综述,证实ACC中谷氨酸浓度持续降低(Luykx等,2012)。 在枕叶皮质中,一些研究人员发现谷氨酸含量增加,这在MDD患者的忧郁亚组中最高(Sanacora等,2008).
在先前的研究中,我们获得了具有MDD和5健康对照的11未治疗受试者中mGluR11受体结合的PET图像(Deschwanden等,2011)。 我们发现在患有抑郁症的患者的前额皮质,扣带皮层,岛叶,丘脑和海马中区域mGluR5结合减少。 另外,抑郁的严重程度与海马中的mGluR5结合负相关。 我们认为这些发现表明抑郁症中mGluR5神经传递减少,可能是由于谷氨酸系统活性的基础或代偿性变化。 此外,我们检查了5抑郁受试者和15匹配对照的死后脑样本中mGluR15表达的量(Deschwanden等,2011)。 我们观察到从抑郁个体获得的样品中前额皮质中mGluR5表达减少。 据报道,抑郁症患者死后脑中NMDA受体表达降低(Feyissa等,2009)。 实际上,先前已经表明mGluR5拮抗剂的抗抑郁特性可能涉及NMDA受体的抑制。 这可能介导海马神经传递和/或诱导脑源性神经营养因子基因表达(Legutko等人,2006)。 此外,最近的一项研究表明,睡眠剥夺可增加健康人的mGluR5可用性(Hefti等,2013)。 在前扣带回皮层,岛状突,内侧颞叶,海马旁回,纹状体和杏仁核中,这种增加与睡眠剥夺的功效显着相关,如主观嗜睡的增加所反映。 这项研究表明,mGluR5的增加可能是神经生物学机制,可以解释睡眠剥夺具有很高的抗抑郁功效。 临床前研究证实了我们假设的mGluR5与睡眠和抑郁之间的关联。 特别是,一项针对mGluR5基因敲除小鼠的研究提供了重要证据,证明mGluR5参与了NREM睡眠的稳定性-REM睡眠状态转换,NREM慢波活动和对睡眠丧失的稳态反应(Ahnaou等人,2015).
总之,来自动物,死后,MRS和PET研究的趋同证据表明,中央谷氨酸系统重要地参与MDD的病理生理学。 然而,有证据表明mGluR5拮抗作用可能不会直接帮助患有MDD的患者。 这符合我们在MDD患者中减少mGluR5表达的发现。 可以推测,靶向mGluR5系统的药物可能对患有共病焦虑,成瘾障碍和/或昼夜节律受损的抑郁患者特别有用。 此外,社会过程系统中的损伤通常与MDD相关。 不良社交技能似乎是抑郁症的重要危险因素(Segrin,2000)。 这些缺陷包括副语言和语言行为,以及面部表情,凝视,姿势和姿势的损伤,与FXS和自闭症谱系障碍中观察到的相当。 此外,关于社交技能,暴露于慢性社会压力的小鼠实验表明,Homer1 / mGluR5偶联被破坏,表明mGluR5夜间缓解了社交压力的抑郁易感性(Wagner等人,正在报道中)。 此外,在社会缺陷的小鼠模型中,mGluR5抑制导致社会交往正常化(Chung等人,2015)。 总之,这些研究提供了临床前证据,即mGluR5在抑郁症的社会原因和抑郁症患者中经常观察到的社会缺陷中起重要作用。 因此,针对mGluR5的药物可能在预防社会缺陷的青少年抑郁症的发展中发挥重要作用,并可能有助于治疗MDD患者的社会缺陷和低心理社会功能。
mGluR5和成瘾
成瘾的特点是持续的药物摄入,尽管有负面后果,反复尝试停止或减少药物使用,以及耐受和戒断的症状。 虽然多巴胺系统在急性奖励处理中起着关键作用(Kalivas和Volkow,2005),越来越多的证据表明成瘾中的谷氨酸能神经传递(Krystal等,2010).
临床前神经生物学研究
在2001中,发表了关于mGluR5和成瘾的开创性研究(Chiamulera等,2001)。 作者表明,缺乏mGluR5受体的小鼠未能获得静脉内可卡因自我给药,尽管急性注射后伏隔核中的细胞外多巴胺水平增加。 许多后续动物研究表明,mGluR5受体拮抗剂MPEP和MTEP减少了可卡因和尼古丁等成瘾药物的自我管理(Kalivas,2009). 橄榄(2005) 回顾了与药物成瘾有关的动物研究,并提出有证据表明mGluR5可能参与药物使用的发展,奖赏感知和复发,如可卡因,吗啡,尼古丁和乙醇。 作者描述了动物研究中的证据,即mGluR5拮抗剂减少了药物的自我管理,以及随后的药物寻求行为。 例如,最近,在暴食症患者的狒狒模型中发现,MTEP降低糖果消费而不改变寻求糖果的行为(Bisaga等,2008)。 研究表明,与使用mGluR5受体拮抗剂相关的食物摄入减少可能与增强刺激的奖赏价值降低有关(Bisaga等,2008)。 此外,研究报告在实验大鼠中重复使用可卡因后,伏隔核和背外侧纹状体中的mGluR5 mRNA水平显着增加(Bisaga等,2008).
人类研究
在患有酒精性疾病和健康对照的患者的死后脑组织样本中使用放射自显影,研究发现患有酒精性疾病的患者的海马和纹状体中的mNluX-30%mGluR40 / 1结合密度更高(Kupila等,2012)。 这一发现表明,在酒精成瘾患者的某些脑区,mGluR5受体密度可能会增加。 最近,我们使用PET测量尼古丁成瘾患者大脑中mGluR5受体的结合(Akkus等,2013)。 我们发现,与非吸烟者相比,5吸烟者的灰质中mGluR14 DVR全球减少(见图) 2)。 最明显的减少发现在右侧和左侧内侧眶额皮质。 我们认为mGluR5受体结合的这种减少可能是对慢性尼古丁给药诱导的谷氨酸慢性增加的长期适应。 这种适应似乎特定于尼古丁依赖,因为它在慢性非吸烟可卡因使用者中尚未发现(Hulka等,2014).
图2
图2。 图像显示三个诊断组中mGluR5 DVR的平均脑摄取。 吸烟者和戒烟者群体的大脑摄取量明显减少。 图像由PMOD软件版本3(PMOD Technologies)计算。 从 Akkus等。 (2013)。 注意:图形及其图例经许可后复制。
实际上,有人提出mGluR5下调代表了代偿性神经适应(Kalivas,2009),药物诱导奖励的增强剂(Rutten等人,2011),或调节条件行为反应中情境线索影响的因素(Tronci等人,2010)。 数字 3 提出了成瘾的mGluR5功能障碍的方法。
图3
图3。 mGluR5参与成瘾.
如图所示 3以前的研究支持mGluR5涉及成瘾的三个关键阶段,开发和获取,药物强化价值以及成瘾复发的观点。 可能有人提出,每个功能主要由不同的大脑区域描述,这些大脑区域也显示出高的mGluR5受体密度,并且已经显示在我们的PET研究中显示出降低的mGluR5密度(Deschwanden等,2011). Kalivas(2009) 开发了一种广泛的谷氨酸成瘾模型。 作者认为,成瘾与皮质纹状体脑回路的关键区域(包括杏仁核,伏隔核(NA),前额皮质和运动皮层)之间的谷氨酸稳态功能障碍有关。 正如我们在图中所示 3首先(“动机成分”),前额皮质可能在药物成瘾发展的初始阶段调解动机和认知控制。 临床前研究 Chiamulera等。 (2001) 表明mGluR5是通过调解滥用药物的有益特性在成瘾发展的这个阶段所必需的。 其次(“奖励成分”),Nucleus Accumbens(NA)已被证明可以影响药物的奖赏价值,不仅通过多巴胺,还通过mGluR5活性(Bisaga等,2008)。 特别是,提出了前肢皮质和NA之间谷氨酸传递的损害,因此通过更加依赖与NMDA / mGluR5受体系统相关的学习行为来启动寻药(Kalivas,2009)。 第三,(“药物寻求组件”),一些研究支持mGluR5更多地依赖于运动过程 - 减少认知控制 - 通过纹状体(Kupila等,2012)。 我们自己发表的和未发表的研究表明,mGluR5下调与戒烟者复发风险增加有关(Akkus,PNAS)。 这种下调可能是致病性或补偿性变化不足。 目前的研究表明,针对mGluR5的药物可以改善成瘾疾病在其发展的各个阶段的治疗。
讨论
回顾的证据表明mGluR5在焦虑症,强迫症,MDD以及成瘾中的高度参与,并且用mGluR5靶向药物治疗也可能对人类有益。 然而,与强迫症相关的情绪和焦虑症状的发病机制可能与与强迫症无关的情绪和焦虑症状不同。 因此,强迫症状的存在可能是针对mGluR5的药物的抗抑郁药和抗焦虑药反应的重要预测因子。 此外,我们建议针对mGluR5系统的药物可能有助于增加对社会压力的抵抗力并改善抑郁症的社会缺陷。 由于社交压力是迄今为止抑郁症和社会缺陷最重要的非遗传风险因素,与社会功能和生活质量的降低密切相关,动物研究的这些发现具有很高的科学和临床相关性。
最近,国家精神卫生研究所(NIMH)发起了一项新颖的,最先进的项目:研究领域标准(RDoC)。 这反映了NIMH战略1.4的实施“为了研究目的,开发基于可观察行为和神经生物学测量的维度对精神障碍进行分类的新方法。”(http://www.nimh.nih.gov/research-priorities/strategic-objectives/strategic-objective-1.shtml)。 这一举措是重组严格的DSM类别的结果,其中大部分也是在神经科学研究之前开发的(Morris和Cuthbert,2012)。 RDoC描述了五个域或结构; 负价系统,正价系统,认知系统,社会过程系统,唤醒/监管系统。 根据有关mGluR5参与MDD和OCD的审查证据,我们提出mGluR5活动可能与负价系统直接相关,负价系统涉及恐惧,威胁,持续威胁,损失和沮丧的非可观察因素。奖励。 因此,正如所评论的研究表明,用mGluR5进行拮抗治疗可以减少那些可观察到的症状。 此外,我们建议mGluR5治疗也可能有利于“阳性价值障碍”,如成瘾和抑郁,通过脑结构的异常mGluR5活性和与mGluR5功能性相关的谷氨酸能NMDA受体相关的功能,并且重要地参与奖励学习。 例如, Simonyi等人。 (2010) 回顾了许多在敲除小鼠中使用mGluR5受体拮抗剂的动物研究,以确定mGluR5在学习和记忆中的作用。 抑制性学习,例如被动回避学习,是用于研究海马学习过程的动物模型中的一项成熟任务,并且已在许多研究中显示依赖于mGluR5受体(Simonyi等,2010)。 例如,研究表明,CA5中mGluR3蛋白在短期和CA1长期增强过程中过度表达(Riedel等,2000). 海曼(2005) 提出了一种成瘾的生物学模型,它结合了学习和记忆的异常神经过程,形成了成瘾的基本要素。 作者提出长期增强作用,包括谷氨酸受体可用性的改变,以及基因表达的调节作为与药物成瘾相关的异常回路中发现的药物诱导的改变的潜在重要机制。 最后,关于mGluR5和睡眠稳态的研究(Hefti等,2013; Ahnaou等人,2015)表明mGluR5在RDoC的唤醒和调节系统域中起着重要作用。
数字 4 总结了用于调解mGluR5在情绪障碍和成瘾中的作用的过程。 图的顶部 4 描绘了高密度mGluR5大脑区域; 杏仁核,海马,纹状体,NA和前额皮质。 精神病综合症与这些地区相匹配。 杏仁核中建议的mGluR5活动可能介导原发性情绪唤起,例如焦虑和抑郁。 我们已经描述了两种可能的途径; 社会功能和学习可能会调解其他过程。 因此,mGluR5活动已经显示出参与学习,因此,通过海马,NA和纹状体的活动可能参与记忆和奖励,认知控制和动机,也涉及成瘾(参见图 3)。 最后,我们建议mGluR5如何与社会压力反应和社会缺陷相关,这可能与广泛的精神疾病相关。 表 2 概述了RDoC损伤区域,相关的临床表现以及mGluR5的参与情况。
图4
图4。 mGluR5机制摘要.
TABLE 2
![]()
表2。 RDoC和mGluR5.
结束语
该综述描述了mGluR5参与情绪障碍,强迫症和成瘾,并将临床前与人体研究,特别是PET研究进行了比较。 因此,我们比较了不同的方法学方法,如动物研究,MRS和PET研究。 我们建议,如果在人类临床试验(RDoC)中开始mGluR5拮抗治疗,则会出现强烈的直接抗焦虑作用。 在FXS中也报道了mGluR5过度活动,其特征在于重要的社会缺陷。 因此,mGluR5活动可能不仅使负价系统和唤醒系统中的活动正常化,而且还减弱了社会过程系统(RDoC)中的损害。 这具有很高的临床重要性,因为不良的社会功能是诸如强迫症,抑郁症和成瘾等普遍的精神疾病的重要结果,导致巨大的个人痛苦和社会的重要间接成本。 最后,mGluR5也被证明具有显着的药物成瘾作用,主要是由于增加药物的奖励价值。 因此,mGluR5拮抗干预对于治疗病理性焦虑和成瘾以及改善社会压力弹性和社会功能最有效。
利益冲突声明
Terbeck博士,Chesterman博士和Akkus博士没有利益冲突。 Hasler博士已从Novartis获得资助,该公司正在生产和测试针对mGluR5的药物。
参考资料
Abe,T.,Sugihara,H.,Nawa,H.,Shigemoto,R.,Mizuno,N。和Nakanishi,S。(1992)。 新型代谢型谷氨酸受体mGluR5与肌醇磷酸/ Ca2 +信号转导偶联的分子特征。 J.Biol.Chem。 267,13361-13368。
PubMed摘要 | 全文 | Google Scholar
Agid,Y.,Buzsáki,G.,Diamond,DM,Frackowiak,R.,Giedd,J.,Girault,J.-A。,et al。 (2007)。 如何改善精神疾病的药物发现? 纳特。 Rev. Drug Discov。 6,189-201。 doi:10.1038 / nrd2217
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Akkus,F.,Ametamey,SM,Treyer,V.,Burger,C.,Johayem,A.,Umbricht,D.,et al。 (2013)。 通过[5C] ABP11正电子发射断层扫描确定吸烟者和戒烟者中mGluR688受体结合的显着全球减少。 PROC。 国家科。 科学院。 科学。 美国。 110,737-742。 doi:10.1073 / pnas.1210984110
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Akkus,F.,Terbeck,S.,Ametamey,SM,Rufer,MD,Treyer,V.,Burger,C.,et al。 (2014)。 代谢性谷氨酸受体5与强迫症患者的结合。 诠释。 J. Neuropsychopharmacol。 17,1915-1922。 doi:10.1017 / S1461145714000716
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Ametamey,SM,Kessler,LJ,Honer,M.,Wyss,MT,Buck,A.,Hintermann,S.,et al。 (2006)。 11CABP688的放射合成和临床前评估作为成像代谢型谷氨酸受体亚型5的探针。 J. Nucl。 地中海。 47,698-705。
PubMed摘要 | 全文 | Google Scholar
Anderson,JJ,Bradbury,MJ,Giracello,DR,Chapman,DF,Holtz,G.,Roppe,J.,et al。 (2003)。 体内 使用新型放射性配体[5 H] 3-甲氧基-3-(吡啶-5-基乙炔基)吡啶)对mGlu2受体拮抗剂的受体占有率。 欧元。 J. Pharmacol. 473, 35–40. doi: 10.1016/S0014-2999(03)01935-6
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Ahnaou,A.,Raeymaekers,L.,Steckler,T。和Drinkenbrug,WHIM(2015)。 代谢型谷氨酸受体(mGluR5)在调节NREM-REM睡眠周期和体内平衡中的相关性:来自mGluR5( - / - )小鼠的证据。 Behav。 Brain Res。 282,218-226。 doi:10.1016 / j.bbr.2015.01.009
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Bear,MF,Huber,KM和Warren,ST(2004)。 脆性X智力低下的mGluR理论。 趋势神经科学。 27,370-377。 doi:10.1016 / j.tins.2004.04.009
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Berg,D.,Godau,J.,Trenkwalder,C.,Eggert,K.,Csoti,I.,Storch,A.,et al。 (2011)。 AFQ056治疗左旋多巴诱发的运动障碍:2随机对照试验的结果。 乐章。 Disord。 26,1243-1250。 doi:10.1002 / mds.23616
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Berry-Kravis,E.,Hessl,D.,Coffey,S.,Hervey,C.,Schneider,A.,Yuhas,J.,et al。 (2009)。 一项试验性开放标签,单剂量试验的fenobam成人脆性X综合征。 J. Med。 遗传学。 46,266-271。 doi:10.1136 / jmg.2008.063701
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Bisaga,A.,Danysz,W。和Foltin,RW(2008)。 谷氨酸能NMDA和mGluR5受体的拮抗作用降低了酗酒疾病的狒狒模型中的食物消耗。 欧元。 Neuropsychopharmacol。 18,794-802。 doi:10.1016 / j.euroneuro.2008.05.004
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Boer,K.,Encha-Razavi,F.,Sinico,M。和Aronica,E。(2010)。 第I组代谢型谷氨酸受体在发育中的人皮质中的差异分布。 脑;。 1324,24-33。 doi:10.1016 / j.brainres.2010.02.005
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Brennan,BP,Rauch,SL,Jensen,JE和Pope,HG(2012)。 对强迫症的磁共振波谱研究的批评性综述。 生物学。 精神病学 73,31-24。 doi:10.1016 / j.biopsych.2012.06.023
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Burket,JA,Benson,AD,Tang,AH和Deutsch,SI(2014)。 雷帕霉素改善了自闭症谱系障碍的BTBR小鼠模型的社交性。 Brain Res。 公牛。 100,70-75。 doi:10.1016 / j.brainresbull.2013.11.005
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Chiamulera,C.,Epping-Jordan,MP,Zocchi,A.,Marcon,C.,Cottiny,C.,Tacconi,S.,et al。 (2001)。 在mGluR5无效突变小鼠中不存在可卡因的增强和运动刺激作用。 纳特。 神经科学。 4,873-874。 doi:10.1038 / nn0901-873
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Chung,W.,Choi,SY,Lee,E.,Park,H.,Kang,J.,Park,H.,et al。 (2015)。 通过NMDAR和mGluR53抑制改善IRSp5突变小鼠的社交缺陷。 纳特。 神经科学。 18,435-442。 doi:10.1038 / nn.3927
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Cleva,RM和Olive,MF(2011)。 5型代谢型谷氨酸受体(mGluR5)的正变构调节剂及其治疗CNS疾病的治疗潜力。 分子 16,2097-2106。 doi:10.3390 / molecules16032097
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Deschwanden,A.,Karolewicz,B.,Feyissa,AM,Treyer,V.,Ametamey,SM,Johayem,A.,et al。 (2011)。 通过[(5)C] ABP11 PET和死后研究确定的重度抑郁症中代谢型谷氨酸受体688密度降低。 上午。 J.精神病学 168,727-734。 doi:10.1176 / appi.ajp.2011.09111607
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Erickson,CA,Mullett,JE和McDougle,CJ(2009)。 脆性X综合征中的开放标签美金刚。 J.自闭症发展 Disord。 39,1629-1635。 doi:10.1007 / s10803-009-0807-3
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Erickson,CA,Early,M.,Stigler,KA,Wink,LK,Mullett,JE和McDougle,CJ(2011)。 一项关于自闭症青少年阿坎酸的开放标签自然主义试验研究。 J. Child Adolesc。 精神药理学。 21,565-569。 doi:10.1089 / cap.2011.0034
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
首先,MB和Gibbon,M。(1997)。 DSM-IV轴I疾病结构化临床访谈用户指南SCID-I:临床医生版本。 纽约州纽约市:American Psychiatric Pub。
Feyissa,AM,Chandran,A.,Stockmeier,CA和Karolewicz,B。(2009)。 在重度抑郁症的前额叶皮质中,NMDA受体和PSD-2的NR2A和NR95B亚基水平降低。 PROG。 Neuropsychopharmacol。 生物学。 精神病学 33,70-75。 doi:10.1016 / j.pnpbp.2008.10.005
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Garber,KB,Visootsak,J。和Warren,ST(2008)。 脆性X综合症。 欧元。 J. Hum。 遗传学。 16,666-672。 doi:10.1038 / ejhg.2008.61
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Gass,JT,Osborne,MP,Watson,NL,Brown,JL和Olive,MF(2009)。 mGluR5拮抗作用减弱甲基苯丙胺的强化,并阻止大鼠恢复寻求甲基苯丙胺的行为。 神经精神药理学。 34,820-833。 doi:10.1038 / npp.2008.140
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Harvey,BH和Shahid,M。(2012)。 代谢和离子型谷氨酸受体作为焦虑和压力相关疾病的神经生物学目标:专注于药理学和临床前转化模型。 药理学。 生物化学。 Behav。 100,775-800。 doi:10.1016 / j.pbb.2011.06.014
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Hasler,G.,LaSalle-Ricci,VH,Ronquillo,JG,Crawley,SA,Cochran,LW,Kazuba,D.,et al。 (2005)。 强迫症症状维度显示与精神病合并症的特定关系。 精神病;。 135,121-132。 doi:10.1016 / j.psychres.2005.03.003
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Hasler,G.,van der Veen,JW,Tumonis,T.,Meyers,N.,Shen,J.,and Drevets,WC(2007)。 使用质子磁共振波谱法测定的主要抑郁症中的前额谷氨酸/谷氨酰胺和γ-氨基丁酸水平降低。 拱。 精神病学 64,193-200。 doi:10.1001 / archpsyc.64.2.193
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Hasler,G.,van der Veen,JW,Geraci,M.,Shen,J.,Pine,D。和Drevets,WC(2009)。 通过质子磁共振波谱法确定惊恐障碍中的前额皮质γ-氨基丁酸水平。 生物学。 精神病学 65,273-275。 doi:10.1016 / j.biopsych.2008.06.023
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Hefti,K.,Holst,SC,Sovago,J.,Bachmann,V.,Buck,A.,Ametamey,SM,et al。 (2013)。 在一夜无睡眠后,人脑中的代谢型谷氨酸受体亚型5可用性增加。 生物学。 精神病学 73,161-168。 doi:10.1016 / j.biopsych.2012.07.030
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Hyman,SE(2005)。 成瘾:学习和记忆的疾病。 上午。 J.精神病学 162,1414-1422。 doi:10.1176 / appi.ajp.162.8.1414
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Hoffpauir,BK和Gleason,EL(2002)。 mGluR5的激活调节视网膜无长突细胞中的GABAA受体功能。 J.神经生理学。 88,1766-1776。
PubMed摘要 | 全文 | Google Scholar
Hulka,LM,Treyer,V.,Scheidegger,M.,Preller,KH,Vonmoos,M.,Baumgartner,MR,et al。 (2014)。 吸烟但不吸食可卡因与人类中较低的脑代谢型谷氨酸受体5密度相关。 摩尔。 精神病学 19,625-632。 doi:10.1038 / mp.2013.51
Kalivas,PW和Volkow,ND(2005)。 成瘾的神经基础:动机和选择的病理学。 上午。 J.精神病学 162,1403-1413。 doi:10.1176 / appi.ajp.162.8.1403
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Kalivas,PW(2009)。 成瘾的谷氨酸稳态假说。 纳特。 Rev. Neurosci。 10,561-572。 doi:10.1038 / nrn2515
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Krystal,JH,Mathew,SJ,D'Souza,DC,Garakani,A.,Gunduz-Bruce,H.和Charney,DS(2010)。 代谢型谷氨酸受体激动剂和拮抗剂的潜在精神病学应用。 CNS药物 24, 669–693. doi: 10.2165/11533230-000000000-00000
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Kupila,J.,Kärkkäinen,O.,Laukkanen,V.,Tupala,E.,Tiihonen,J。和Storvik,M。(2012)。 酒精受试者大脑中的mGluR1 / 5受体密度:整个半球放射自显影研究。 精神病;。 12,149-155。 doi:10.1016 / j.pscychresns.2012.04.003
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Koros,E.,Rosenbrock,H.,Birk,G.,Weiss,C。和Sams-Dodd,F。(2007)。 选择性mGlu5受体拮抗剂MTEP,类似于NMDA受体拮抗剂,诱导大鼠的社会隔离。 神经精神药理学 32,562-576。 doi:10.1038 / sj.npp.1301133
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Lea,PM和Faden,AI(2006)。 代谢型谷氨酸受体亚型5拮抗剂MPEP和MTEP。 CNS药物Rev。 12,149-166。 doi:10.1111 / j.1527-3458.2006.00149.x
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Legutko,B.,Szewczyk,B.,Pomierny-Chamioło,L.,Nowak,G。和Pilc,A。(2006)。 MPEP处理对脑源性神经营养因子基因表达的影响。 药理学。 众议员。 58,427-430。
PubMed摘要 | 全文 | Google Scholar
Luykx,JJ,Laban,KG,van den Heuvel,MP,Boks,MPM,Mandl,RCW,Kahn,RS,et al。 (2012)。 重度抑郁症的区域和状态特异性谷氨酸下调:(1)H-MRS发现的荟萃分析。 神经科学。 Biobehav。 启。 36,198-205。 doi:10.1016 / j.neubiorev.2011.05.014
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Meldrum,BS(2000)。 谷氨酸作为大脑中的神经递质:生理学和病理学的回顾。 J. Nutr。 130,1007。
PubMed摘要 | 全文 | Google Scholar
Morris,SE和Cuthbert,BN(2012)。 研究领域标准:认知系统,神经回路和行为维度。 对话Clin。 神经科学。 14,29-37。
PubMed摘要 | 全文 | Google Scholar
Olive,MF(2005)。 mGlu5受体:神经解剖学,药理学和药物成瘾中的作用。 CURR。 精神病学牧师。 1,197-214。 doi:10.2174 / 1573400054065578
Phan,KL,Fitzgerald,DA,Cortese,BM,Seraji-Bozorgzad,N.,Tancer,ME和Moore,GJ(2005)。 社交焦虑障碍中的前扣带神经化学:1特斯拉的4H-MRS。 Neuroreport 16, 183–186. doi: 10.1097/00001756-200502080-00024
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Pittenger,C.,Krystal,JH和Coric,V。(2006)。 谷氨酸调节药物作为治疗强迫症的新型药物治疗剂。 NeuroRx 3,69-81。 doi:10.1016 / j.nurx.2005.12.006
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Pollack,MH,Jensen,JE,Simon,NM,Kaufmann,RE和Renshaw,PF(2008)。 社交焦虑症中GABA,谷氨酸和谷氨酰胺的高场MRS研究?:对左乙拉西坦治疗的反应。 PROG。 Neuropsychopharmacol。 生物学。 精神病学 32,739-743。 doi:10.1016 / j.pnpbp.2007.11.023
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Riaza Bermudo-Soriano,C.,Perez-Rodriguez,MM,Vaquero-Lorenzo,C。和Baca-Garcia,E。(2012)。 谷氨酸和焦虑的新视角。 药理学。 生物化学。 Behav。 100,752-774。 doi:10.1016 / j.pbb.2011.04.010
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Riedel,G.,Casabona,G.,Platt,B.,Macphail,EM和Nicoletti,F。(2000)。 恐惧条件诱导的大鼠海马中mGlu5受体蛋白表达的时间和亚区特异性增加。 神经药理学 39, 1943–1951. doi: 10.1016/S0028-3908(00)00037-X
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Rutten,K.,De Vry,J.,Bruckmann,W。和Tzschentke,TM(2011)。 NOP受体的药理学阻断或基因敲除增强了吗啡在大鼠中的奖赏效果。 药物酒精依赖。 114,253-256。 doi:10.1016 / j.drugalcdep.2010.10.004
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Romano,C.,van den Pol,AN和O'Malley,KL(1996)。 增强的代谢型谷氨酸受体mGluR5在大鼠脑中的早期发育表达:蛋白质,mRNA剪接变体和区域分布。 J.比较。 神经病学。 367,403-412。
Rosenberg,DR和Keshavan,MS(1998)。 走向强迫症的神经发育模型。 生物学。 精神病学 43, 623–640. doi: 10.1016/S0006-3223(97)00443-5
Sanacora,G.,Zarate,CA,Krystal,JH和Manji,HK(2008)。 针对谷氨酸能系统开发新的,改进的情绪障碍治疗方法。 纳特。 Rev. Drug Discov。 7,426-437。 doi:10.1038 / nrd2462
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Schoepp,DD(2001)。 揭示中枢神经系统中突触前代谢型谷氨酸受体的功能。 J. Pharmacol。 进出口。 疗法。 299,12-20。
PubMed摘要 | 全文 | Google Scholar
Segrin,C。(2000)。 与抑郁症相关的社交技能缺陷。 临床。 心理学。 启. 20, 379–403. doi: 10.1016/S0272-7358(98)00104-4
Simonyi,A.,Schachtman,TR和Christoffersen,GR(2010)。 代谢型谷氨酸受体亚型5在学习和记忆中的拮抗作用。 欧元。 J. Pharmacol。 639,17-25。 doi:10.1016 / j.ejphar.2009.12.039
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Soares,DP和Law,M。(2009)。 大脑的磁共振波谱:代谢物和临床应用的回顾。 临床。 放射学杂志。 64,12-21。 doi:10.1016 / j.crad.2008.07.002
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Swanson,CJ,Bures,M.,Johnson,MP,Linden,A.-M.,Monn,JA和Schoepp,DD(2005)。 代谢型谷氨酸受体作为焦虑和应激障碍的新靶点。 纳特。 Rev. Drug Discov。 4,131-144。 doi:10.1038 / nrd1630
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Szeszko,PR,Christian,C.,Macmaster,F.,Lencz,T.,Mirza,Y.,Taormina,SP,et al。 (2008)。 精神药物 - 幼稚小儿强迫症的灰质结构改变:优化的基于体素的形态测量研究。 上午。 J.精神病学 165,1299-1307。 doi:10.1176 / appi.ajp.2008.08010033
Tatarczyńska,E.,Klodzińska,A.,Chojnacka-Wójcik,E.,Palucha,A.,Gasparini,F.,Kuhn,R.,et al。 (2001)。 MPEP的潜在抗焦虑和抗抑郁样作用,MPEP是一种有效的,选择性和系统活性的mGlu5受体拮抗剂。 BR。 J. Pharmacol。 132,1423-1430。 doi:10.1038 / sj.bjp.0703923
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Tronci,V.,Vronskaya,S.,Montgomery,N.,Mura,D。和Balfour,DJK(2010)。 mGluR5受体拮抗剂6-甲基-2-(苯基乙炔基) - 吡啶(MPEP)对尼古丁的行为反应的影响。 精神药理学 211, 33–42. doi: 10.1007/s00213-010-1868-x
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Van den Heuvel,OA,Remijnse,PL,Mataix-Cols,D.,Vrenken,H.,Groenewegen,HJ,Uylings,HBM,et al。 (2009)。 强迫症的主要症状维度是由部分不同的神经系统介导的。 大脑 132,853-868。 doi:10.1093 / brain / awn267
Wagner,KV,Hartmann,J.,Labermaier,C.,Häusl,AS,Zhao,G.,Harbich,D.,et al。 (在新闻)。 Homer1 / mGluR5活动缓解了长期社会压力的脆弱性。 神经精神药理学。 doi:10.1038 / npp.2014.308
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
Yüksel,C。和Öngür,D。(2010)。 磁共振波谱研究谷氨酸相关的情绪障碍异常。 生物学。 精神病学 68,785-794。 doi:10.1016 / j.biopsych.2010.06.016
Zarei,M.,Mataix-Cols,D.,Heyman,I.,Hough,M.,Doherty,J.,Burge,L.,et al。 (2011)。 强迫症青少年灰质体积和白质微观结构的变化。 生物学。 精神病学 70,1083-1090。 doi:10.1016 / j.biopsych.2011.06.032
PubMed摘要 | 全文 | CrossRef全文 | Google Scholar
关键词:mGluR5,PET,情绪障碍,成瘾,焦虑
引用:Terbeck S,Akkus F,Chesterman LP和Hasler G(2015)代谢型谷氨酸受体5在情绪障碍和成瘾的发病机制中的作用:将临床前证据与人体正电子发射断层扫描(PET)研究相结合。 面前。 神经科学. 9:86。 doi:10.3389 / fnins.2015.00086
收到:07 1月2015; 接受:27二月2015;
发布时间:18 March 2015。
编辑:
阿肖克·库马尔,美国佛罗里达大学
点评人:
Karthik Bodhinathan,Sanford Burnham医学研究所,美国
Amber M. Muehlmann,美国佛罗里达大学
版权所有©2015 Terbeck,Akkus,Chesterman和Hasler。 这是一份根据条款分发的开放获取文章 知识共享署名许可(CC BY)。 允许在其他论坛中使用,分发或复制,前提是原始作者或许可人被记入贷方,并且根据公认的学术惯例引用本期刊中的原始出版物。 不允许使用,分发或复制,不符合这些条款。
*通讯:英国普利茅斯PL4 8AA,普利茅斯大学健康与人文科学学院心理学院Sylvia Terbeck [电子邮件保护]