KOMENTET E YBOP: Efekti Coolidge është pas fuqisë së pornos në internet. Efekti Coolidge është një fenomen që shihet tek speciet e gjitarëve, ku meshkujt (dhe në një masë më të vogël femrat) shfaqin interes të përtërirë seksual nëse prezantohen me partnerë të rinj seksualë pranues, edhe pasi të refuzojnë seksin nga partnerët seksualë të mëparshëm, por akoma në dispozicion. Risia seksuale e tejkalon këtë habitat me eksitim të përtërirë të shkaktuar nga dopamina më e lartë. Një rrymë e vazhdueshme e risive është ajo që e bën pornografinë në internet kaq të ndryshme nga pornot e së kaluarës.
Artikulli origjinal, me grafikë
- Dennis F. Fiorino,
- Ariane Coury, dhe
- Anthony G. Phillips
- Gazeta e Neurosciences, 15 qershor 1997, 17 (12): 4849-4855;
Abstrakt
Efekti Coolidge përshkruan reinitiation e sjelljes seksuale në një kafshë "seksualisht ngopur" në përgjigje të një mik i ri receptive. Duke pasur parasysh rolin e sistemit mesolimbik dopamin (DA) në inicimin dhe mbajtjen e sjelljes së motivuar, mikrodializa është përdorur për të monitoruar transmetimin e DA nga bërthamë accumbens (NAC) gjatë ndërthurjes, ngopjes seksuale dhe reinitiimit të sjelljes seksuale. Në pajtim me raportet e mëparshme, paraqitja e një femre estrous prapa një ekran dhe copulation u shoqëruan me rritje të konsiderueshme në NAC DA efflux. Kthimi i përqendrimeve të NAC DA në vlerat bazë përputhet me një periudhë të ngopjes seksuale, megjithëse përqendrimet e metabolitëve të DA, acidit dihidroksifenilcetilne dhe acid homovanilik, mbetën të larta. Prezantimi i një femre të re të hapur pas një ekrani rezultoi në një rritje të lehtë në NAC DA, e cila u shtua ndjeshëm gjatë ripërsëritjes me femrën e re. Të dhënat e tanishme sugjerojnë se vetitë stimuluese të një femre të re në dispozicion mund të shërbejnë për të rritur transmetimin e NAC DA në një rat mashkull seksualisht të ngopur, dhe kjo, nga ana tjetër, mund të lidhet me reinitiimin e sjelljes seksuale.
Prezantimi
Një mi mashkull që është copuluar në ngopje mund të nxitet për t'u bashkuar përsëri nëse femra fillestare zëvendësohet me një femër të re. Kjo është bërë e njohur si efekti Coolidge dhe është vërejtur në një numër speciesh gjitarësh (Wilson et al., 1963). Faktorë të përgjithshëm si lodhja ose depresioni motorik nuk janë të mjaftueshme për të shpjeguar gjendjen e dukshme të ngopjes seksuale, sepse stimujt nga një femër romane ende mund të nxisin copulimin. Ngopja seksuale gjithashtu mund të "kthehet" në mënyrë farmakologjike, në një shkallë të konsiderueshme, nga administrimi i një sërë barnash që mund të veprojnë në sisteme të ndryshme të neurotransmetuesit. Këto barna përfshijnë yohimbine, 8-OH-DPAT (Rodriguez-Manzo dhe Fernandez-Guasti, 1994, 1995a), nalaxone (Pfaus dhe Gorzalka, 1987; Rodriguez-Manzo dhe Fernandez-Guasti, 1995a,b), dhe apomorfin (Mas et al., 1995c). Megjithëse veprimi periferik i këtyre barnave nuk mund të përjashtohet (p.sh., efektet adrenergike në funksionimin e ngrerë), efektet mbi mekanizmat qendrorë të ngopjes seksuale janë propozuar në bazë të eksperimenteve selektive të qendrimit noradrenergik të lesionit (Rodriguez-Manzo dhe Fernandez-Guasti, 1995a) dhe eksperimentet e mikrodializës që monitoruan metabolizmin dopaminergjik në zonën mediale preoptike (Mas et al., 1995a,b).
Duke pasur parasysh se mekanizmat qendrore mund të ndërmjetësojnë në reinitiation e sjelljes seksuale karakteristike të efektit Coolidge, një kandidat i mundshëm është sistemi mesolimbik dopamin (DA), projektuar nga zona tegmentale ventral në NAC. Mesolimbic DA duket se vepron si një modulator primar në proceset integruese komplekse që përfshijnë vlerësimin e stimujve të mjedisit, të tilla si cues nga një femër seksualisht e hapur dhe organizimi i sjelljeve të drejtuara nga qëllimi, duke përfshirë edhe copulimin (Fibiger dhe Phillips, 1986; Blackburn et al., 1992; Phillips et al., 1992; LeMoal, 1995; Salamone, 1996).
Megjithëse neuronet e midbrain DA përgjigjen ndaj shpërblimeve primare dhe cues që parashikojnë shpërblimin, novela ose stimujt e paparashikueshëm mjedisor nxisin aktivizimin neuronal më të fuqishëm gjatë seancave të përsëritura të trajnimit (Fabre et al., 1983; Schultz, 1992; Mirenowicz dhe Schultz, 1994). Ka shumë prova që mbështesin një rol të rëndësishëm lehtësues për DA mezolimbic në inicimin dhe ruajtjen e sjelljes seksuale të miut (Pfaus dhe Everitt, 1995), dhe një numër i studimeve të mikrodializës raportojnë rritje në NAC DA efflux gjatë fazave të shijshme dhe konsumuese të sjelljes seksuale mashkullore (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995b). Megjithatë, ekzistojnë relativisht pak të dhëna për korrelacionin neurokemikësor të ngopjes seksuale dhe reinitiimin e sjelljes seksuale. Aplikimi i in vivo mikrodializa për të monitoruar neurotransmetimin DA mesolimbik gjatë efektit Coolidge ofron një mundësi unike për të shqyrtuar rolin e NAC DA në copulation, ngopje seksuale, dhe reinitiation e copulation.
U bë një eksperiment mikrodializëm për të përcaktuar si vijon: (1) nëse fillimi i ngopjes seksuale shoqërohet me kthimin e përqëndrimeve ekstrakullulare DA në NAC në vlerat e parablerjes ose më poshtë, dhe (2) nëse rivendosja e sjelljes së copulimit në një " seksualisht i ngopur "miu mashkull me një femër të re në dispozicion është e ndërlidhur me rritjen e NAC DA efflux.
MATERIALE DHE METODA
Subjekteve. Rats Mashkull Sprague Dawley, të marra nga Qendra e Kujdesit të Kafshëve (në Universitetin e British Columbia) dhe rats femra Long-Evans, të marra nga Charles River Canada (St Constant, Quebec, Kanada), ishin vendosur në kafaze rrjetë teli (18 × 25 × 65 cm, pesë për kafaz) në dhoma koloni të veçanta. Dhomat e kolonisë mbaheshin në një temperaturë prej ~20 ° C në një ciklit të kundërt të 12 hr / të errët. Rats kishin qasje të pakufizuar në ushqim (Purina Rat Chow) dhe ujë.
Kirurgjia dhe testimi i sjelljes para mikrodializës së trurit.Rats femra u mbivendosën në mënyrë bilaterale nën anestezi të gazit halothane (Fluothane, Laboratori Ayerst) të paktën 4 javë para testimit. Përgjegjshmëria seksuale në femrat stimuluese u induksionet nga injeksione nënlëkurore të benzoatit estradiol (10 μg) dhe progesteroni (500 μg), 48 dhe 4 hr, përkatësisht, para secilës sesion testimi. Rats meshkuj janë shfaqur për sjellje seksuale në dy raste, 4 d përveç, në dhomat e Plexiglas (35 × 35 × 40 cm) me dysheme me rrjetë teli. Vetëm rats meshkuj që arritën një kriter të performancës, që përfshinte intromizionin brenda 5 min të paraqitjes së femrës dhe ejakulimit brenda 15 min të intromissionit të parë, gjatë dy testeve të shqyrtimit u implantuan me tuba udhëzues të mikrodializës udhëzuese.
Minjtë meshkuj (n = 5) u anestezuan me ketamine hydrochloride (100 mg / kg, ip) dhe ksilazine (10 mg / kg, ip) para operacionit stereotak. Kanalizuesit e udhëzuesit të mikrodializës (matës 19) janë implantuar në mënyrë bilaterale mbi NAC (koordinatat nga bregma: anterior, + 1.7 mm, mesatare, ± 1.1 mm, ventral, -1.0 mm, kafkë e sheshtë) dhe u siguruan në kafkë me akrilik dentar vida e argjendarëve. Implantet e kanulave udhëzuese dypalëshe u përdorën për të maksimizuar mundësinë për një eksperiment të suksesshëm të mikrodializës. Për fat të mirë, në eksperimentin e tanishëm, vetëm një kanulë ishte e nevojshme për çdo miu. Ratsë meshkuj janë vendosur individualisht në kafaze të mëdha plastike me shtratin e kokës për kokë për pjesën e mbetur të eksperimentit. Një javë pas operacionit, minjtë u testuan për sjellje seksuale. Gjatë kësaj pjese të trajnimit, dhoma e testimit ishte e pajisur me një ekran të rrëshqitur të Plexiglas që ndau dhomën në ndarje të mëdha dhe të vogla. Rrotat meshkuj u futën në ndarje të madhe dhe 15 min më vonë, një femër u vendos pas ekranit. Pas një periudhe përgatitore 15 min, ekrani u hoq, dhe minjtë u lejuan të copulonin për 30 min. Janë kryer tre sesione trajnimi, një çdo 4 d. Të gjitha minjtë arritën kriterin e performancës gjatë çdo seance.
Eksperiment Coolidge efekt. Rats u implantuan në mënyrë të njëanshme me hetimet e mikrodializës 12-18 orë përpara eksperimentit të efektit Coolidge dhe vendosën në ndarjen e madhe të dhomës së testimit me qasje të lirë në ushqim dhe ujë. Në mëngjesin e eksperimentit, mostrat e mikrodializës u mblodhën çdo minutë 15. Eksperimenti përbëhej nga shtatë fazat e njëpasnjëshme: (1) bazë (të paktën 60 min); (2) femra 1 pas ekranit (15 min); (3) me 1 femra derisa një periudhë Min 30 kaluar pa një mal; (4) riintrodhimi i femrës 1 prapa ekranit (15 min); (5) në femrën 1 për një periudhë minimale 15, me kusht që të mos ketë montim (në qoftë se ndodh montimi, kjo fazë është trajtuar si fazë 3); (6) futja e femrës 2 prapa ekranit (15 min); 7) me femra 2 për 60 min.
Sjellja u filmua nën ndriçim të ulët duke përdorur një sistem video JVC dhe u vërejt në një monitor video të vendosur jashtë dhomës së testimit. Masat standarde të sjelljes seksuale janë regjistruar duke përdorur një kompjuter dhe softuer të përshtatshëm (Holmes et al., 1987).
Pas eksperimentit të mikrodializës, kafshëve iu dha një mbidozë e hidratit chloral dhe perfundoi intracardially me kripur dhe formalin (4%). Trurit ishin feta dhe të ngrira, dhe më pas, seksionet koronale u njolluan me violin kromi për të përcaktuar vendosjen e sondave të mikrodializës. Vetëm rats me vendosje të hetimeve brenda NAC u përdorën për analiza të sjelljes dhe neurochemical.
Mikrodijalizmi dhe zbulimi i elektrolizës HPLC. Sondat mikrodializuese ishin të përqendruara në dizajn me një membranë fibre boshe gjysmëpërçues (membrana 2 mm e ekspozuar, diametri i jashtëm i 340 μm, ndërprerja e peshës molekulare 65000, Filtral 12, Hospal) në fundin distal. Sondat u perfunduan në 1.0 μl / min me një zgjidhje të ndryshuar të Ringerit (0.01 m buffer fosfat natriumi, pH 7.4, 1.3 mmCaCl2, 3.0 mm KCl, 1.0 mmMgCl2, 147 mm NaCl) duke përdorur një shiringë gastight (Hamilton, Reno, NV) dhe një pompë shiringë (model 22, Harvard Aparati, South Natick, MA). Një jakë udhëzuese për sondën e mikrodializës është përdorur për të siguruar hetimin e mikrodializës brenda kanulës së udhëzuesit. Një spirale çeliku, e lidhur me një xhama të lëngshme (Instech 375s) që ishte montuar në krye të dhomës së testimit, u përdor për të mbrojtur tubin e provës (Fiorino et al., 1993).
Analitat e mikrodializës, të cilat përfshinin DA dhe metabolitëve të saj acid dihyroxyphenylacetic (DOPAC) dhe acid homovanillic (HVA), u ndanë me kromatografi të fazës së kundërt (kolona Ultrasphere, Beckman, Fullerton, CA, ODS 5 μm, 15 cm, 4.6 mm, ) duke përdorur një tampon 0.083m acetat natriumi, pH 3.5 (metanol 5%). Përqendrimet e analizës u quantifikuan nga zbulimi elektrokimik (EC). Aparati përbëhej nga një pompë Bio-Rad (Richmond, CA), një Injector me dy pozicione të Valco Instruments (Houston, TX) EC10W, një EC detektor ECA (Bedford, MA) Coulochem II dhe një regjistrues me dy kanale (Kipp dhe Zonen, Bohemia, NY). Parametrat e detektorit elektrokemik ishin: elektroda 1, + 450 mV; elektrodë 2, -300 mV; dhe qeliza roje, -450 mV. Përpjekjet tipike të hetimeve, të kryera vitro dhe në temperaturën e dhomës, ishin 22% për DA, 18% për DOPAC dhe 18% për HVA.
REZULTATET
Sjellje
Masat e sjelljes nga eksperimenti i efektit Coolidge janë paraqitur në Tabelën 1. Latenzitë për të ngritur, intromituar, dhe ejakuluar, si dhe për intervalin e pozicionimit pas derdhjes së parë ishin të ngjashme me ato të sesionit të mëparshëm të trajnimit (të dhënat nuk janë treguar). Kjo sugjeron që procedura e mikrodializës nuk ka ndryshuar sjelljen normale seksuale. Zhvillimi i ngopjes seksuale, siç tregohet nga numri mesatar i ejakulacioneve para kriterit është përmbushur (7.8 ± 0.5), një rënie progresive në numrin e intromozave që i paraprijnë secilit ejakulim dhe një rritje progresive në intervalin e pozicioneve (të dhënat nuk tregohen) , ishte e ngjashme me atë të raportuar në studimet e mëparshme (Plazh dhe Jordan, 1956; Fowler dhe Whalen, 1961; Fisher, 1962; Bermant et al., 1966; Rodriguez-Manzo dhe Fernandez-Guasti, 1994; Mas et al., 1995d). Ndryshueshmëria individuale është vërejtur në lidhje me numrin e ejakulimeve të arritura me femrën 1, kohën e kaluar duke kopjuar me femrën 1 dhe numrin e paraqitjeve të femrës 1 që kërkohen për të arritur kriterin e ngopjes (Tabela 1, poshtë). Disa rats kërkuan shumëfishim të riintrodukteve të femrave 1 derisa faza 5 ishte e plotë (n = 3). Aktet e vendosjes së femrës 1 prapa ekranit dhe heqja e ndarjes mund të kenë shërbyer si cues parësorë që çojnë në bashkim. Duhet gjithashtu të theksohet se një kriter i ngopjes së 30 min pa një mal, edhe pse përdoret më parë (Plazhi dhe Jordania, 1965; Mas et al., 1995b), është arbitrar dhe nuk garanton që një rat nuk do të kishte qenë i pajisur me më shumë kohë. Megjithatë, vonesat ose procedurat e largimit dhe zëvendësimit nuk rezultuan në mënyrë të besueshme në bashkimin e ripërtërirë me femrën 1 (p.sh., fazat 4 dhe 5).

Sjellje gjatë eksperimentit të efektit Coolidge
Të gjitha minjtë shfaqën efektin Coolidge. Aktiviteti që lidhet me vendosjen e femrës 2 prapa ekranit dhe në veçanti heqja e ndarjes mund të ketë kontribuar në këtë rezultat, por përsëri, këto ngjarje nuk ishin në vetvete të mjaftueshme për të ripërtërirë copulimin më herët në eksperiment. Krahasimet në mes të masave të sjelljes seksuale me femra 1 dhe femra 2 janë bërë duke përdorur t testet me korrigjim Bonferroni. Megjithëse latentet e malit dhe të intromizionit në përgjigje të femrës 2 nuk ishin të ndryshme në mënyrë të konsiderueshme nga ato në periudhën e parë të bashkimit me femrën 1, në përgjithësi, sjellja seksuale me femrën 2 ishte më pak e fuqishme, siç tregohet nga ejaculations dukshëm më pak (të thotë, 0.6 vs 4.2; F = 49.86;p <0.01) dhe intromisimet (mesatarja = 11.2 kundrejt 37.0;F = 20.17; p <0.05) gjatë orës së parë. Numri i montimeve në orën e parë me femrat 1 dhe 2 nuk ishte i ndryshëm në mënyrë të konsiderueshme.
Është e rëndësishme të theksohet se femrat e përdorura gjatë pjesës së ngopjes së eksperimentit (p.sh., femra 1) ende shfaqin sjellje të fuqishme proceptive (dmth. Hopping dhe darting) dhe receptive (dmth. Lordosis) për kohëzgjatjen e plotë të kontaktit të tyre me mashkullin .
neurochemistry
Përqendrimet basal nanomolar të DA dhe metabolitëve të saj në mikrodializate, të paraqitura si mesatare ± SEM, e tre mostrave bazë të parë ishin: DA, 3.0 ± 0.7; DOPAC, 619.1 ± 77.7; dhe HVA, 234.2 ± 49.0 (pa korrigjuar për rikuperimin e sondës;n = 5). Këto vlera përfaqësonin pikët bazë të 100%.
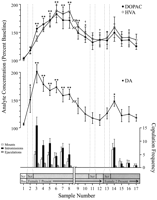
Pikat e të dhënave të përcaktuara në sjellje, që korrespondojnë me çdo fazë të eksperimentit dhe të përbashkëta për çdo rat, janë përdorur për analiza neurochemical. Këto përfshinin në vijim: (1) shtatë mostra pas futjes së parë të femrave 1, (2) katër mostra që shoqërojnë mungesën e sjelljes copulative me femrën 1 dhe (3) pesë mostra pas paraqitjes së femrës 2. figurë 1 ilustron ndryshimet në përqendrimet e DA (grafiku i linjës, mes) dhe metabolitëve DA (grafiku i linjës, më i lartë) paralleling sjellje copulatory (grafik bar, fund) gjatë testit për efektin Coolidge.
Shiko versionin më të madh:
Fig. 1.
Nucleus accumbens correlates neurochemical e sjelljes seksuale gjatë efektit Coolidge. Tetë mostrat e para përfaqësojnë pikat kronologjike të vazhdueshme të të dhënave nga fazat 1 në 3. Shembulli 1 është mostra e katërt dhe e fundit e prepolulimit bazë (Bas). Shembull 2 përfaqëson futjen e femrës 1 prapa ekranit (SCR). Pas 15 min, ekrani u hoq dhe rats lejoheshin të copulonin (mostra 3-8). pushim më x-axi korrespondon me përjashtimin e të dhënave nga tre rats që kombinohen për periudha të gjata me femrën fillestare. Nëntë mostrat e fundit ishin gjithashtu kronologjikisht të vazhdueshme. Mostrat 9 dhe 10 korrespondojnë me periudhën e ngopjes së fazës 3 (dmth, min 30 pa një montim). Femra 1 u riinstalua prapa ekranit (mostër 11) dhe, 15 min më vonë, ekrani u hoq (mostra 12). Pas 15 min pa ndërprerje, femra 2 u vendos pas ekranit (mostër 13). Mostrat 14-17 korrespondojnë me copulimin me femrën 2. Numri i rritjeve, intromissave ose ejakulimeve të lidhura me çdo mostër mikrodializmi 15 min janë treguar në grafiku i poshtëm i barit. Të dhënat neurochemical janë shprehur në përqindje të përqendrimeve bazë. Ndryshimet në NAC DA (sheshet e mbyllura), DOPAC (qarqe të mbyllura), dhe HVA (qarqe të hapura) rrjedh janë paraqitur si line grafikë. U bënë krahasimet e mëposhtme: mostra bazë 1 kundrejt mostrave 2-10; mostër e re bazë 10 kundrejt mostrave 11 dhe 12; mostër e re bazë 12 kundrejt mostrave 13-17 (*p <0.05; ** p <0.01). I pavarur t testet janë bërë midis vlerave bazë (mostrat 1, 10 dhe 12). Për dallime të rëndësishme nga baza e parë (mostër 1), †p <0.05.
I veçuar në një mënyrë, masa të përsëritura ANOVA u kryen në të dhënat neurokemike të shoqëruara me femra 1 (mostrat 1-12) dhe femra 2 (mostra 12-17). A priori krahasimet u bënë duke përdorur testin e shumëfishimit të Dunn-it (Bonferroni t). Dy krahasimet kryesore u bënë: (1) bazë fillestare (mostër 1) kundrejt mostrave 2-10 (ekspozimi i parë ndaj femrës 1), (2) baza e dytë bazë (mostër 10) kundrejt mostrave 11 dhe 12 (riekspozimi ndaj femrës 1) , dhe (3) baza e tretë bazë (mostër 12) kundrejt mostrave 13-17 (ekspozimi ndaj femrës 2).
Ka pasur një ndryshim të rëndësishëm të përgjithshëm në efektin DA në përgjigje të femrës 1 [F (11,44) = 8.48; p <0.001] dhe femra 2 [F (5,20) = 2.83;p <0.05]. Një rritje e konsiderueshme në rrjedhën e DA u gjet kur femra 1 ishte e pranishme pas ekranit (+ 44%,p <0.05; shembulli 2). Gjatë bashkimit, përqendrimet e DA u rritën më tej, duke arritur një vlerë maksimale (+ 95%;p <0.01) gjatë periudhës së parë të përpilimit (shembulli 3). DA mbeti e ngritur gjatë gjithë përpilimit dhe u kthye vetëm në përqendrimet fillestare në periudhën 30 min në të cilën nuk ndodhi asnjë montim (mostrat 9 dhe 10). As rivendosja e femrës 1 pas ekranit (shembulli 11) dhe as mundësia për të bashkëvepruar fizikisht, por pa u montuar (shembulli 12), përqendrimet e ngritura të DA në krahasim me vlerën e dytë fillestare (shembulli 10). Prania e femrës 2 pas ekranit (mostra 13) rezultoi në një rritje të vogël të fluksit DA (12%) nga vlera e tretë fillestare (shembulli 12) që nuk arriti rëndësinë statistikore. Kopulimi i rinovuar me femrën 2 rezultoi në një rritje të konsiderueshme (34%) (p <0.05) në fluksin DA gjatë kampionit të parë të kopulimit (shembulli 14). Megjithëse sjellja e dobët e kopulimit vazhdoi gjatë tre mostrave të ardhshme, përqendrimet e DA u ulën në vlerat fillestare (mostrat 15–17). I pavarur t testet e kryera në mesin e mostrave "bazë" (p.sh., 1, 10 dhe 12) treguan se këto vlera nuk ishin dukshëm të ndryshme.
Në të tre rats që rifilluan copulation kur femra 1 ishte reintroduced, përqendrimet NAC DA rritur kur femra 1 ishte i pranishëm prapa ekranit (varg, 25-47%) dhe gjatë copulation (varg, 13-37%), në krahasim me mostër vetëm para riprodhimit të femrës. Këto rritje, megjithatë, ndodhën vetëm kur sjellja seksuale ishte e fuqishme dhe çoi në derdhje.
Ndryshime të rëndësishme të përgjithshme në DOPAC [F (11,44) = 9.57; p <0.001] dhe HVA [F (11,44) = 12.47; p <0] përqendrimet u gjetën në përgjigje të femrës 001, por jo femrës 1. Përqendrimet e metabolizmit u rritën lehtë (+ 2% në të dy rastet) gjatë prezantimit të femrës 15 pas ekranit (shembulli 1), por kjo nuk ishte e rëndësishme statistikisht. Sidoqoftë, pati rritje të konsiderueshme në përqendrimet e DOPAC dhe HVA gjatë copulimit (mostrat 2-3), duke arritur vlerat maksimale (+8 dhe + 80%, respektivisht; p <0.01) pas 60 min (shembulli 6 në të dy rastet). Megjithëse përqendrimet e metaboliteve u ulën gjatë periudhës së pasivitetit seksual në fund të kontaktit me femrën 1 (mostrat 9 dhe 10), përqendrimet ende mbetën të ngritura në lidhje me fillimin (p <0.05 në të dy rastet). Rivendosja e femrës 1 pas ekranit (shembulli 11), qasja tek femra 1 pas heqjes së ekranit (shembulli 12) dhe futja e femrës 2 (shembulli 13) nuk rezultoi në ndonjë ndryshim në përqendrimet e metabolitit. Rritjet e lehta, por statistikisht të parëndësishme, në përqendrimet e DOPAC dhe HVA (+ 23% në të dy rastet) në krahasim me fillimin (shembulli 12) korrespondonin me periudhën e parë të kompulimit me femrën 2 (shembulli 14). Kjo rritje ishte jetëshkurtër, megjithatë, dhe nuk pranoi vlerat fillestare për tre mostrat e mbetura (15–17). I pavarur ttestet e kryera në mesin e mostrave "bazë" (p.sh., 1, 10 dhe 12) treguan se vlerat e dytë dhe të tretë bazë (mostrat 10 dhe 12 respektivisht), megjithëse jo të ndryshme nga njëri-tjetri, mbetën të larta në krahasim me mostrën e parë bazë për DOPAC dhe HVA (p <0.05 në të dy rastet).
Histologji
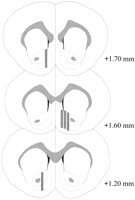
Sondat mikrodializuese janë vendosur në NAC (Fig.2) në një distancë që shtrihet + 1.20 në + 1.70 mm nga bregma (kafka e sheshtë). Ka pasur ndryshueshmëri edhe në planin mediolateral; të dhënat reflektojnë marrjen e mostrave nga nënparetet shell dhe nënligjore të NAC.
Shiko versionin më të madh:
Fig. 2.
Lokacioni i sondave të mikrodializës brenda NAC të minjve meshkuj të përdorur në eksperimentin e efektit Coolidge. Drejtkëndëshe të shaded korrespondojnë me zonën e ekspozuar të membranës të sondave të mikrodializës. Seksionet e trurit koronal të trurit u redrawn ngaPaxinos dhe Watson (1986).
DISKUTIMI
Në pajtim me raportet e mëparshme, rezultatet e tanishme tregojnë rritje të transmetimit mesolimbik DA të shoqëruar me komponente të shijshme dhe të konsumueshme të sjelljes seksuale të meshkujve meshkuj siç vlerësohet ngain vivo microdialysis (Mas et al., 1990; Pfaus et al., 1990;Pleim et al., 1990; Damsma et al., 1992; Wenkstern et al., 1993; Fumero et al., 1994; Mas et al., 1995a,b,d). Përveç kësaj, këto rezultate ofrojnë një korrelacion neurokemik për ngopje seksuale dhe reinitiation pasuese e copulation në përgjigje të një femër të reja receptive (efekti Coolidge). Të dhënat e tanishme sugjerojnë se vetitë stimuluese të një femre të re në dispozicion mund të shërbejnë për të rritur transmetimin e NAC DA në një rat mashkull seksualisht të ngopur që mund, nga ana tjetër, të lidhet me reinitiimin e sjelljes seksuale. Kjo është parë qartë në rritjen e lehtë të NAC DA gjatë prezantimit të një femre të re prapa ekranit dhe ndodh më bindshëm si një rritje më e theksuar gjatë ripërsëritjes së copulimit me femrën 2 (Fig.1).
Prania e femrës së parë të hapur pas ekranit rezultoi në një rritje të fuqishme të NAC DA efflux (44% nga baseline) e ngjashme me madhësinë e asaj që u raportua në eksperimentet e mëparshme duke përdorur një dizajn të ngjashëm (30% Pfaus et al., 1990; 35%,Damsma et al., 1992) Gjithashtu në marrëveshje me këto studime ishte vëzhgimi se rrjedha e NAC DA u rrit më tej gjatë kopulimit (në> 95% mbi nivelin bazë në eksperimentin e tanishëm). Megjithëse ne mund t'i shohim sjelljet konsumuese si të shoqëruara me rritjen e lëshimit të NAC DA (Wenkstern et al., 1993; Wilson et al., 1995), është e rëndësishme që të shqyrtohen termat "oreks" dhe "konsumues" në kontekstin e sjelljes seksuale. Ndërsa faza në të cilën femra është e pranishme pas ekranit është ekskluzivisht apetit ose përgatitor, sjellja gjatë fazës së copulimit nuk mund të konsiderohet thjesht konsumuese. Për shkak se "oreksi" mund të përdoret për të përshkruar të gjitha sjelljet që çojnë në konsumimin e një sjelljeje të motivuar (kopulimi), sjellja primare që ekspozon meshkuj ndërsa është aktive në fazën "konsumatore" përshkruhet më së miri si apetit; burri shpenzon pjesën më të madhe të kohës dhe përpjekjet e tij për të ndjekur femrën për t'u bashkuar. Në këtë drejtim, ne mund të korrelojmë transmetimin maksimal të NAC DA me konsumues si edhe komponentë intensive të oreksit të sjelljes seksuale të meshkujve.
Qasja në femrën e dytë, roman, rezultoi në përtëritje të përsëritura në çdo lëndë. Studimet e mëparshme kanë treguar se pjesa më e madhe e rats lejohet të copulojnë në ngopje, duke përdorur një protokoll të ngjashëm të sjelljes me atë të përdorur në eksperimentin e tanishëm, nuk rifilloi bashkimin kur u testua më vonë 24 (Plazh dhe Jordan, 1956). Është e mundshme që prania e pronave stimuluese romane të femrës 2, e cila mund të ketë përfshirë shenja të nuhatjes, si dhe vizuale dhe auditive, rezultoi në një copulim të përtërirë. Një pyetje interesante, e cila mbetet për t'u përgjigjur, është me çfarë mekanizmi një rat mashkull dallon një femër romane nga një femër me të cilën ai ka bashkuar së fundi. Një vend për këtë mekanizëm mund të qëndrojë në sistemin kryesor të nuhatjes. Është raportuar se integriteti i këtij sistemi është thelbësor për efektin Coolidge në hamsters (Johnston dhe Rasmussen, 1984). Sidoqoftë, sistemi i nuhatjes vomeronasale-accessory, në të cilin një proces memorie pheromonal u përshkrua kohët e fundit në minj (Kaba et al., 1994), është gjithashtu një kandidat kryesor. Në këtë drejtim, vlen të përmendet se rritjet në transmetimin e NAC DA janë matur duke përdorur in vivo voltammetria në minjtë meshkuj të paraqitur me shtrat që ishte ekspozuar ndaj minjve femra në estrus (Louillot et al., 1991; Mitchell dhe Gratton, 1992). Për më tepër, aplikimi i K+ drejtpërdrejt në shtresën nervore vomeronasale të llambës nuhatëse shtesë, si dhe vetë llambës aromatike ndihmëse, ishte e mjaftueshme për të rritur transmetimin e NAC DA (Mitchell dhe Gratton, 1992).
Periudha e parë 15 min e copulation me femra 2 ishte e lidhur me një rritje të konsiderueshme në NAC DA. Në dallim nga femra 1, ndërveprimi me femrën 2 nuk ka prodhuar rritje në NAC DA të të njëjtit magnitudë gjatë fazave të oreksit (12%) ose konsumit (34%). Këto rritje të vogla në NAC DA, megjithatë, korrelojnë mirë me nivelin e reduktuar të sjelljes seksuale të shfaqur me femra 2 krahasuar me femrën 1. Përqendrimet e metaboliteve mbetën të larta gjatë fazës së ngopjes, duke rezultuar në përqëndrime të reja bazë (mostrat 10 dhe 12) që janë ngritur dukshëm nga vlera fillestare bazë (mostër 1).
Vonesa kohore në rritjen e përqendrimeve DOPAC dhe HVA gjatë copulimit është në përputhje me formimin e tyre si metabolitë e përbërësit primar, DA. Është sugjeruar që përqendrimet e metaboliteve të mikrodializës, të paktën gjatë një sjelljeje natyrore që nuk drejtohet në mënyrë farmakologjike, ofrojnë një indeks të dobishëm të aktivitetit nervor (Damsma et al., 1992; Fumero et al., 1994). Fakti që përqendrimet e metaboliteve mbeteshin të larta edhe gjatë periudhave të pasivitetit seksual në këtë eksperiment, kur përqendrimet e DA-ve ishin kthyer për të paracaktuar vlerat bazë, hedh dyshime mbi këtë sugjerim.
Përqendrimi i vazhdueshëm i përqendrimeve të metaboliteve DA shihet në këtë eksperiment pasqyron profilin e mesopozitës preoptike (mPOA) të metabolitëve DA të vërejtur në minjtë ditën e parë pasi ata kishin copuluar në ngopje (Mas et al., 1995a,b). Ngritjet e qëndrueshme të përqendrimeve DOPAC dhe HVA në NAC ose mPOA nuk respektohen gjithmonë kur periudha e çiftëzimit është me kohëzgjatje të caktuar, shumë më e shkurtër se sa koha e nevojshme për të arritur ngopje. Për shembull, shumë studime kanë treguar se përqendrimet e DOPAC u rritën dhe mbetën të larta gjatë bashkimit, por nuk arritën të vlerësonin vlerat bazë menjëherë pas largimit të femrës (Pfaus et al., 1990; Pleim et al., 1990; Damsma et al., 1992;Hull et al., 1993; Wenkstern et al., 1993; Hull et al., 1995). Në studimin e Mas et al. (1995b), përqendrimet bazike ekstracelulare të DOPAC dhe HVA në mPOA mbetën të ngritura gjatë 4 ditëve të njëpasnjëshme që korrespondojnë me një periudhë të pasivitetit seksual. Deri në ditën e katërt, pak para se kafshët rifilluan copulimin, përqendrimet bazale të metabolitëve ishin afër vlerave të precizimit. Autorët e krahasuan modelin e ndryshimeve neurokimike tek ato që shiheshin pas administrimit të bllokuesve të receptorëve DA (Zetterström et al., 1984; Imperato dhe DiChiara, 1985) dhe kanë sugjeruar se gjendja e pasivitetit seksual mund të ndërmjetësohet nëpërmjet lirimit të prolaktinës, i cili mund të veprojë si një "neuroleptik endogjen" (Mas et al., 1995a,b,d). Është e qartë se administrimi neuroleptik shoqërohet me rritje të përqendrimeve të metaboliteve të jashtme dhe DA efflux (Zetterström et al., 1984; Imperato dhe DiChiara, 1985). Për fat të keq, Mas et al. (1995a,b) nuk ishin në gjendje të zbulonin përqendrimet e mPOA DA. Në studimin aktual, përqendrimet e DA në NAC u kthyen në vlerat e parablerjes, ndërsa përqendrimet DOPAC dhe HVA mbetën të larta. Ky model nuk është në përputhje me një rol për një neuroleptik endogjen që vepron në NAC për të nxitur ngopje seksuale.
Duke pasur parasysh përfshirjen e neuroneve mesolimbike DA në sjellje të motivuar (Fibiger dhe Phillips, 1986; Blackburn et al., 1992; Kalivas et al., 1993; LeMoal, 1995) dhe ndjeshmërinë e tyre ndaj stimujve të rinj mjedisorë (Fabre et al., 1983; Schultz, 1992; Mirenowicz dhe Schultz, 1994), rritjet e vërejtura në përqëndrimet ekstracelulare të NAC DA në përgjigje të femrës së re janë në përputhje me hipotezën se aktiviteti në këtë sistem DA është i rëndësishëm për riinitiimin e sjelljes seksuale. Përveç kësaj, raportet e rritjeve të shijshme dhe të përhershme në transmetimin DA (Hull et al., 1993, 1995;Mas et al., 1995b; Sato et al., 1995) dhe aktivitetin neuronal (Shimura et al., 1994) në mPOA të minjve meshkuj gjatë sjelljes seksuale sugjerojnë se kjo strukturë mund të kontribuojë gjithashtu në përtëritjen e copulimit karakteristike të efektit Coolidge.
Në përputhje me një rol të përgjithshëm për sistemin mesolimbik DA në sjellje të motivuar, është vërtetuar se përqendrimet ekstracelulare të DA gjithashtu janë ngritur para, gjatë dhe menjëherë pas konsumimit të një vakt, me një kthim në vlerat bazë ~ 30 min më vonë (Wilson et al., 1995). Dihet mirë se ngopja e shkaktuar nga ushqimi ndikohet nga vetitë e tij sensore. Njerëzit dhe kafshët refuzojnë ushqimin në të cilin ushqehen me ngopje dhe ushqehen me ushqime të tjera që nuk ishin ngrënë (Rolls, 1986). Kjo shtron pyetjen nëse dalja jashtëqelizor DA në NAC do të rritet në mënyrë selektive nga paraqitja e një lloji të ri të ushqimit, por jo nga ushqimi i konsumuar kohët e fundit për ngopje në një mënyrë analoge me atë të raportuar në këtë studim në kontekstin e motivimi seksual. Nëse konfirmohet, kjo marrëdhënie e përgjithshme ndërmjet vetive shqisore të shpërblimeve natyrore, ngopjes dhe transmetimit mesolimbik DA do të nënkuptonte një rol kritik për këtë sistem nervash në rregullimin e proceseve motivuese, çrregullimi i të cilave mund të çojë në çrregullime serioze të ngrënies dhe funksionit seksual .
REFERENCAT
- ↵
- Plazh FA,
- Jordan L
(1956) Shterimi seksual dhe rimëkëmbja në miun meshkuj. QJ Exp Psychol 8: 121-133.
- ↵
- Bermant G,
- Lott DF,
- Anderson L
(1966) Karakteristikat kohore të efektit Coolidge në sjelljen e copulatorit të meshkujve. J Comp Physiol Psychiatry 65: 447-452.
- ↵
- Blackburn JR,
- Pfaus JG,
- Phillips AG
(1992) Funksionet e dopamines në sjelljet joshëse dhe mbrojtëse. Prog Neurobiol 39: 247-279.
- ↵
- Damsma G,
- Pfaus JG,
- Wenkstern D,
- Phillips AG,
- Fibiger HC
(1992) Sjellja seksuale rrit transmetimin e dopamines në bërthamën e akumulimeve dhe striatumit të minjve meshkuj: krahasimi me risi dhe locomotion. Behav Neurosci 106: 181-191.
- ↵
- Fabre M,
- Rolls ET,
- Ashton JP,
- Williams G
(1983) Aktiviteti i neuroneve në rajonin tegmental ventral të majmunit sjellës. Behav Brain Res 9: 213-235.
- ↵
- Fibiger HC,
- Phillips AG
(1986) Shpërblimi, motivimi, njohja: psikobiologjia e sistemeve të dopaminës mesotelencefalike. në Manualin e fiziologjisë: sistemi nervor IV, eds Bloom FE, Geiger SD (American Physiology Society, Bethesda, MD), pp 647-675.
- ↵
- Fiorino DF,
- Coury AG,
- Fibiger HC,
- Phillips AG
(1993) Stimulimi elektrik i vendeve të shpërblimit në zonën tegmentale të barkut rrit transmetimin e dopamines në bërthamën e akumulimeve të miut. Behav Brain Res 55: 131-141.
- ↵
- Fisher A
(1962) Efektet e variacionit të stimujve në ngopje seksuale në miun meshkuj. J Comp Physiol Psychiatry 55: 614-620.
- ↵
- Fowler H,
- Whalen RE
(1961) Variacioni në nxitjen stimuluese dhe sjelljen seksuale në miun meshkuj. J Comp Physiol Psychiatry 54: 68-71.
- ↵
- Fumero B,
- Fernendez-Vera JR,
- Gonzalez-Mora JL,
- Mas M
(1994) Ndryshimet në qarkullimin e monoaminës në zonat paragjykuese të lidhura me sjelljen seksuale mashkullore: një studim i mikrodializës. Brain Res 662: 233-239.
- ↵
- Holmes GM,
- Holmes DG,
- Sachs BD
(1987) Një sistem i mbledhjes së të dhënave të IBM-PC për regjistrimin e sjelljes seksuale të brejtësve dhe për regjistrimin e ngjarjeve të përgjithshme. Physiol Behav 44: 825-828.
- ↵
- Hull EM,
- Eaton RC,
- Moisiu J,
- Lorrain DS
(1993) Copulation rrit aktivitetin dopamin në zonën mediatike preoptic të minjve meshkuj. Life Sci 52: 935-940.
- ↵
- Hull EM,
- Jianfang D,
- Lorrain DS,
- Matuszewich L
(1995) dopamin extracellulare në zonën mediatike preoptike: implikimet për motivimin seksual dhe kontrollin hormonal të kopulimit. J Neurosci 15: 7465-7471.
- ↵
- Imperato A,
- DiChiara G
(1985) Lirimi dhe metabolizmi i dopamines në rats të zgjuar pas neuroleptikëve sistemikë siç është studiuar nga dializa trans-striatale. J Neurosci 5: 297-306.
- ↵
- Johnston RE,
- Rasmussen K
(1984) Njohja individuale e hamsters femra nga meshkujt: roli i cues kimike dhe i sistemeve të nuhatjes dhe vomeronasale. Physiol Behav 33: 95-104.
- ↵
- Kaba H,
- Hayashi Y,
- Higuchi T,
- Nakanishi S
(1994) Induksioni i kujtesës së nuhatjes nëpërmjet aktivizimit të receptorit metabotropik të glutamatit. Shkencë 265: 262-264.
- ↵
- Kalivas PW,
- Sorg BA,
- Hooks MS
(1993) Farmakologjia dhe qarku nervor i sensibilizimit të psikostimulantëve. Behav Pharmacol 4: 315-334.
- ↵
- LeMoal M
(1995) Neuronet dopaminergjike mesokortikolimike. Rolet funksionale dhe rregullatore. në Psychopharmacology: gjenerata e katërt e progresit, eds Bloom FE, Kupfer DJ (Raven, Nju Jork), pp 283-294.
- ↵
- Louillot A,
- Gonzalez-Mora JL,
- Guadalupe T,
- Mas M
(1991) Stimulimet e nuhatjes lidhur me gjininë nxisin një rritje selektive në lirimin e dopamines në bërthamën e akumulimeve të minjve meshkuj. Brain Res 553: 313-317.
- ↵
- Mas M,
- Gonzalez-Mora JL,
- Louillot A,
- Sole C,
- Guadalupe T
(1990) Rritja e lirimit të dopaminës në bërthamën e akumulimeve të përhapjes së minjve të meshkujve, siç dëshmohet nga voltammetria in vivo. Neurosci leton 110: 303-308.
- ↵
- Mas M,
- Fumero B,
- Fernandez-Vera JR,
- Gonzalez-Mora JL
(1995a) Korrelacionet neurokemikale të lodhjes dhe shërimit seksual siç vlerësohet nga mikrodializa in vivo. Brain Res 675: 13-19.
- ↵
- Mas M,
- Fumero B,
- Gonzalez-Mora JL
(1995b) Monitorimi voltammetrik dhe mikrodializmi i lirimit të trurit të monoaminës neurotransmetues gjatë ndërveprimeve socioseksuale. Behav Brain Res 71: 69-79.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I
(1995c) Induksioni i sjelljes mating nga apomorphine në rats seksualisht të ngopura. Eur J Pharmacol 280: 331-334.
- ↵
- Mas M,
- Fumero B,
- Perez-Rodriguez I,
- Gonzalez-Mora JL
(1995d) Neurokimi i ngopjes seksuale. Një model eksperimental i dëshirës së frenuar. në Farmakologji e funksionit seksual dhe mosfunksionim, ed Bancroft J (Raven, Nju Jork), pp 115-126.
- ↵
- Mirenowicz J,
- Schultz W
(1994) Rëndësia e paparashikueshmërisë për përgjigjet e shpërblimit në neuronet primare dopaminergjike. J Neurophysiol 72: 1024-1027.
- ↵
- Mitchell JB,
- Gratton A
(1992) Lirimi Mesolimbic dopamine i nxitur nga aktivizimi i sistemit olfaktor accessory: nje studim krononometrike me shpejtesi te larte. Neurosci leton 140: 81-84.
- ↵
- Paxinos G,
- Watson C
(1986) Truri i miut në koordinatat stereotaaktive (2nd ed). (Akademik, San Diego).
- ↵
- Pfaus JG,
- Damsma G,
- Nomikos GG,
- Wenkstern D,
- Blaha CD,
- Phillips AG,
- Fibiger HC
(1990) Sjellja seksuale rrit transmetimin qendror dopamin në miun meshkuj. Brain Res 530: 345-348.
- ↵
- Pfaus JG,
- Everitt BJ
(1995) Psikofarmacologjia e sjelljes seksuale. në Psychopharmacology: gjenerata e katërt e progresit, eds Bloom FE, Kupfer DJ (Raven, Nju Jork), pp 743-758.
- ↵
- Pfaus JG,
- Gorzalka BB
(1987) Opioidet dhe sjelljet seksuale. Neurosci Biobehav Rev 11: 1-34.
- ↵
- Phillips AG,
- Blaha CD,
- Pfaus JG,
- Blackburn JR
(1992) Korrelacione neurobiologjike të gjendjeve pozitive emocionale: dopamine, pritje dhe shpërblim. në rishikimin ndërkombëtar të studimeve mbi emocionet, Ed Strongman (Wiley, New York), f. 31-50.
- ↵
- Pleim ET,
- Matochik JA,
- Barfield RJ,
- Auerbach SB
(1990) Korrelacioni i lirimit të dopaminës në nucleus accumbens me sjellje seksuale mashkullore në minjtë. Brain Res 524: 160-163.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1994) Anulimi i lodhjes seksuale nga agjentët serotonergik dhe noradrenergik. Behav Brain Res 62: 127-134.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995a) Pjesëmarrja e sistemit qendror noradrenergjik në rivendosjen e sjelljeve copulative të minjve të rraskapitur seksualisht nga yohimbine, naloxone dhe 8-OH-DPAT. Brain Res Bull 38: 399-404.
- ↵
- Rodriguez-Manzo G,
- Fernandez-Guasti A
(1995b) Antagonistët e opioideve dhe fenomenin e ngopjes seksuale. Psychopharmacol 122: 131-136.
- ↵
- Rrotullon BJ
(1986) Ndjeshmëria e ngopjes specifike. Nutr Rev 44: 93-101.
- ↵
- Salamone JD
(1996) Neurokimi i sjelljes së motivimit: çështjet metodologjike dhe konceptuale në studimet e aktivitetit dinamik të dopamines së nucleus accumbens. J Neurosci Metodat 64: 137-149.
- ↵
- Sato Y,
- Wada H,
- Horita H,
- Suzuki N,
- Shibuya A,
- Adachi H,
- Kato R,
- Tsukamoto T,
- Kumamoto Y
(1995) Lirimi i dopamines ne zonen preoptike mesatare gjate sjelljes bashkepunuese te meshkujve ne rats. Brain Res 692: 66-70.
- ↵
- Schultz W
(1992) Aktiviteti i neuroneve dopamine ne primat sjelljen. Semin Neurosci 4: 129-138.
- ↵
- Shimura T,
- Yamamoto T,
- Shimokochi M
(1994) Zona preoptike mesatare përfshihet në eksitimin seksual dhe performancën në minjtë meshkuj: rivlerësimi i aktivitetit të neuroneve në kafshët që lëvizin lirshëm. Brain Res 640: 215-222.
- ↵
- Wenkstern D,
- Pfaus JG,
- Fibiger HC
(1993) Transmetimi i dopamines rritet në bërthamën e akumulatorëve të minjve meshkuj gjatë ekspozimit të tyre të parë në minjtë seksualisht të pranueshëm. Brain Res 618: 41-46.
- ↵
- Wilson C,
- Nomikos GG,
- Collu M,
- Fibiger HC
(1995) Korrelatet dopaminergjike të sjelljes së motivuar: rëndësia e përzjerjes. J Neurosci 15: 5169-5178.
- ↵
- Wilson JR,
- Kahn RE,
- Plazh FA
(1963) Modifikimi në sjelljen seksuale të minjve meshkuj të prodhuar duke ndryshuar femrën stimuluese. J Comp Physiol Psychiatry 56: 636-644.
- ↵
- Zetterström T,
- Sharp T,
- Ungerstedt U
(1984) Efekti i medikamenteve neuroleptike në lirimin striatal dopamin dhe metabolizmin në miun e zgjuar të studiuar nga dializa intracerebrale. Eur J Pharmacol 106: 27-37.
Artikuj që citojnë këtë artikull
- Neuroplasticiteti endogjen i opioidit i Neuroneve dopaminergjike në zonën tegmentale ventrale ndikon në shpërblimin e natyrshëm dhe të kujdesshëm Gazeta e Neuroscience, 25 Qershor 2014, 34 (26): 8825-8836
- Mosbalancim në ndjeshmërinë ndaj llojeve të ndryshme të shpërblimeve në kumar patologjik Truri, 1 gusht 2013, 136 (8): 2527-2538
- Natyrore dhe Shpërblime për Drogën Akti për Mekanizmat e Përbashkët Plastik të Neuralitetit me Delegacionin FosB si Ndërmjetës kryesor Gazeta e Neuroscience, 20 Shkurt 2013, 33 (8): 3434-3442
- Përgjigjet neuronale në Shell Nucleus Accumbens gjatë Sjelljes Seksuale në Rats Mashkull Gazeta e Neuroscience, 1 Shkurt 2012, 32 (5): 1672-1686
- Dashuria romantike: një sistem i trurit gjitar për zgjedhjen e bashkëshortit Transaksionet Filozofike të Shoqërisë Mbretërore B: Shkencat Biologjike, 29 Dhjetor 2006, 361 (1476): 2173-2186
- Hot Spots Hedonic në Brain Neuroscientist, 1 Dhjetor 2006, 12 (6): 500-511
- Modulimi nga Bërthamat Amygdalar Qendrore dhe Basolaterale të Korelateve Dopaminergjike të Ushqyerjes në Saktësi në Bërthamën e Rat Ratacit dhe Cortexin Medial Prefrontal Journal of Neuroscience, 15 Dhjetor 2002, 22 (24): 10958-10965
- Shpeshtësia e Tranzumit të Përqendrimit të Dopamines Rritet Striatum Dorsal dhe Ventral i Rats Mashkull gjatë futjes së personave të zhdukur Journal of Neuroscience, 1 Dhjetor 2002, 22 (23): 10477-10486
- Rishikimi i Librit: Dinamika e dopaminës ekstrakelulare në veprimet akute dhe kronike të kokainës Neuroscientist, 1 gusht 2002, 8 (4): 315-322
- Përfshirja Diferenciale e NMDA, AMPA / Kainate, dhe Dopamine Receptors në Nucleus Accumbens Core në blerjen dhe kryerjen e sjelljes Pavlovian Approach Journal of Neuroscience, 1 Dhjetor 2001, 21 (23): 9471-9477
- Kontrolli i sjelljes që kërkon kokainë nga stimujt e lidhura me drogën në minjtë: Efektet në shërimin e niveleve të qelizave të dendura të reagimit operues dhe extracellulare në amygdala dhe nucleus accumbens PNAS, 11 prill 2000, 97 (8): 4321-4326
- Lehtësimi i Sjelljes Seksuale dhe Shtimi i Efflux-it të Dopamisë në Akumulimin e Bërthamës së Meshkujve pasi D-Amfetamina-Induktuar Sjelljen e Sjelljes Gazeta e Neuroscience, 1 Janar 1999, 19 (1): 456-463
- NEUROSCIENCE: Tërheqja e vëmendjes së trurit Shkenca, 3 tetor 1997, 278 (5335): 35-37
- Korrelacionet Dopaminergjike të Saktësisë Ndjesore-Speciale në Cortezin dhe Bërthamën Prefrontale Mediale të Ratit Gazeta e Neuroscience, 1 tetor 1999, 19 (19): RC29
