Komente: Ky rishikim është prodhuar nga kreu i NIDA, Nora Volkow, dhe ekipi i saj. Nuk lë pak dyshime se varësitë kimike dhe varësitë e sjelljes ndajnë mekanizmat e njëjtë ose të ngjashëm dhe qarkun nervor. Kjo ka kuptim të përsosur pasi që varësitë kimike rrëmbejnë qarkun nervor për lidhjen, seksin dhe ngrënien. Që nga seksi lëshon dy herë më shumë dopamine se sa të hahet ushqimi juaj i preferuar dhe një përdorues pornografi mund të mbajë dopamin të ngritur për orë të tëra, është e çmendur të propozojë që varësia nga pornografia të mos ekzistojë.
Curr Top Behav Neurosci. 2011 Tetor 21.
Volkow ND, Wang GJ, Fowler JS, Tomasi D, Baleri R.
Burim
Instituti Kombëtar për Abuzimin e Drogës, 6001 Boulevard Ekzekutiv 6001, Dhoma 5274, Bethesda, MD, 20892, SHBA, [email mbrojtur].
Abstrakt
Si varësia e drogës, ashtu edhe obeziteti mund të përkufizohen si çrregullime në të cilat vlera e shpërhapjes së një lloj shpërblimi (droga dhe ushqimi, respektivisht) rritet në mënyrë jonormale krahasuar me dhe në kurriz të të tjerëve. Ky model është në përputhje me faktin se të dy ilaçet dhe ushqimi kanë efekte të fuqishme përforcuese - pjesërisht të ndërmjetësuara nga rritjet e dopaminës në sistemin limbik - që, në rrethana të caktuara ose tek individë të prekshëm, mund të mbingarkojnë mekanizmat e kontrollit homeostatik të trurit. Paralele të tilla kanë gjeneruar interesa të rëndësishme për të kuptuar dobësitë dhe trajektoret e përbashkëta midis varësisë dhe obezitetit. Tani, zbulimet e imazhit të trurit kanë filluar të zbulojnë karakteristika të përbashkëta midis këtyre dy kushteve dhe të përshkruajnë disa nga qarqet e trurit që mbivendosen, disfunksionet e të cilëve mund të shpjegojnë deficite stereotipike dhe të ngjashme të sjelljes në subjektet njerëzore. TKëto rezultate sugjerojnë se të dy personat e trashë dhe të varur nga droga vuajnë nga dëmtimet në rrugët dopaminergjike që rregullojnë sistemet nervore të lidhura jo vetëm me ndjeshmërinë e shpërblimeve dhe motivimin nxitës, por edhe me kondicionimin (kujtesën / mësimin), kontrollin e impulsit (frenimin e sjelljes), reaktivitetin e stresit , dhe ndërgjegjësim interoceptive. Këtu integrojmë gjetjet që mbizotërojnë kryesisht nga tomografi i emisioneve të pozitronit që hedhin dritë mbi rolin e dopaminës në varësinë e drogës dhe në obezitetin dhe propozojmë një model pune të përditësuar për të ndihmuar në identifikimin e strategjive të trajtimit që mund të përfitojnë të dyja këto kushte.
1 Sfond
2 Roli i dopaminës në shpërblimin akut ndaj drogës dhe ushqimit
3 Imaging DA në përgjigje të drogës dhe në cues të kushtëzuar në varësi
4 Ndikimi i mosfunksionimit në kontrollin inhibitor
5 Përfshirja e Qarqeve të Motivimit
6 Përfshirja e Circuitry Interoceptive
7 Circuitry of Aversion
8 Shpërblimi i drogës patologjike dhe ushqimit: Një model i përditësuar i punës
1 Sfond
Dopamine (DA) konsiderohet si një çelës për efektet shpërblyese të shpërblimeve natyrore dhe të drogës. Megjithatë, roli i saj në humbjen e kontrollit dhe sjelljeve të detyrueshme që lidhen me varësinë dhe obezitetin janë shumë më pak të qarta. Studimet e PET kanë luajtur një rol vendimtar në karakterizimin e rolit të sistemeve DA të trurit në varësi (përveç rolit të saj në shpërblimin e drogës) dhe në obezitetin. Në të vërtetë, medikamentet e abuzimit (duke përfshirë alkoolin) konsumohen nga njerëzit ose vetë-administrohen nga kafshët laboratorike sepse ato janë në thelb shpërblim, një efekt që ndërmjetësohet nëpërmjet pronave të tyre për përmirësimin e DA në sistemin mesolimbik 2009). Hgjithsesi, në rastin e varësisë, studimet e imazhit kanë zbuluar se çrregullimi ndikon jo vetëm në qarkun e shpërblimit të DA, por gjithashtu edhe në rrugët e tjera DA të përfshira në modulimin e kushteve të kondicionimit, motivimit dhe funksioneve ekzekutive (kontrollin frenues, atributin e ndarjes dhe vendimin ), dhe se deficitet DA mund të marrin pjesë gjithashtu në reaktivitetin e zgjerimit të stresit dhe ndërprerjen e ndërgjegjësimit ndëretik të lidhur me varësinë. Studimet preklinike dhe klinike kanë zbuluar gjithashtu neurotransmetues të tjerë (dhe neuropeptide) që luajnë rol të rëndësishëm në shpërblimin e drogës dhe varësisë (dmth. kanabinoidet, opioidet) dhe janë të përfshira në ndryshimet neuroplastike që ndjekin përdorimin e përsëritur të drogës (p.sh., glutamat, opioidet, GABA, faktori që liron kortikotropin). Sistemi glutamatergik është veçanërisht i shquar në këtë drejtim, sepse ai ndërmjetëson ndërprerjet në të dy fuqizimin afatgjatë dhe depresionin afatgjatë që janë vërejtur në modelet e kafshëve të administrimit kronik të drogës (Thomas et al. 2008). Shqyrtimet që u përkasin këtyre sistemeve shtesë mund të gjenden diku tjetër (Kalivas 2009; Koob 1992).
Meqenëse drogat aktivizojnë të njëjtat sisteme shpërblimi që janë bazë shpërblimi ushqimor, nuk është krejtësisht e papritur që, në përgjithësi, studimet e imazhit të trurit kanë mbështetur nocionin se dëmtimet në qarqet moduluar nga DA gjithashtu përfshihen në sjelljet patologjike, të detyrueshme të ngrënies. Cues të ushqimit, si cues të drogës, rritin DA jashtë stronik dhe nxisin motivimin për t'u angazhuar në sjelljet që janë të nevojshme për të blerë dhe ngrënë ushqimin, duke ofruar dëshmi për përfshirjen e DA jo vetëm në shpërblimin e ushqimit, por edhe në motivimin jo-hedonik vetitë e ushqimit (dmth. kërkesat kalorike) dhe zvogëlimi i kontrollit frenues të parë në overeating kompulsive (Avena et al. 2008; Volkow et al. 2008a).
Këtu, ne i rishikojmë të gjeturat nga studimet e imazhit që në mënyrë specifike përqendrohen në mbivendosjet në qarqet e trurit që ndërpriten në obezitetin dhe në varësinë nga droga. Vlen të kujtohet, megjithatë, se rregullimi i sjelljes së marrjes së ushqimit është shumë më kompleks sesa rregullimi i marrjes së drogës. Kjo e fundit është ndërmjetësuar kryesisht nga efektet e dobishme të drogës, ndërsa e para është e moduluar jo vetëm nga efektet e saj të dobishme (faktorët hedonikë), por edhe nga faktorë të shumtë dhe periferikë që ndjejnë kërkesat e lëndëve ushqyese në trupin e nevojshëm për mbijetesë (faktorët homeostatik). Është interesant fakti se faktorët homeostatik (p.sh., insulina, leptina, greliini) e modulojnë marrjen e ushqimit në pjesë, duke rritur ose zvogëluar ndjeshmërinë e qarqeve të shpërblimit të trurit tek stimujt e ushqimit (Volkow et al. 2011a).
2 Roli i dopaminës në shpërblimin akut ndaj drogës dhe ushqimit
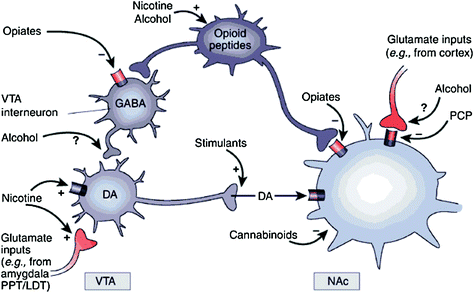
Nëse në mënyrë të drejtpërdrejtë apo të tërthortë, të gjitha barërat e varësisë shfaqin një aftësi për të rritur DA në bërthamë accumbens (NAc) nëpërmjet ndërveprimeve specifike me objektiva të ndryshëm molekulare (Nestler 2004) (Fik. 1). Doza mesolimbike DA [qelizat DA në zonën tegmentale të barkut (VTA) që projektojnë në NAc] duket të jenë vendimtare për shpërblimin e drogës (Wise 2009). Sidoqoftë, siç përshkruhet më poshtë, shtigjet e tjera të DA-së [mezostriatale (qelizat DA në substantia nigra projektuar në striatum dorsal) dhe mesokortik (qelizat DA në VTA projektuar në korteksin frontal)] gjithashtu kontribuojnë në shpërblimin e drogës dhe varësisë 2009). Në përgjithësi, duket se efektet e dobishme dhe të kondicionuar të drogës janë kryesisht të nxitura nga zjarri phasic DA, çka çon në rritje të mëdha dhe të përkohshme DA. Në të kundërt, ndryshimet në rrjedhën e poshtme të funksionit ekzekutiv që ndodhin në varësi janë të lidhura me ndryshimet në qitjen e qelizave tonike DA dhe rezultojnë në nivele më të ulëta, por më të qëndrueshme DA (Grace 2000; Wanat et al. 2009). Kjo, nga ana tjetër, tregon për receptorët D1 (D1R), të cilat janë receptorë DA me afinitet të ulët që stimulojnë sinjalizimin ciklik AMP, si të përfshirë si në shpërblimin akut të drogës, ashtu edhe në kushtëzimin, pasi këto lidhen me përqendrimet e larta DA për të stimuluar D1R. Në të kundërt, D2Rs, të cilat pengojnë sinjalizimin ciklik të AMP, stimulohen nga DA-ja fasike dhe tonike. Vini re se, për shkak të mungesës së radiotracuesve të veçantë për imazhet e PET të receptorëve DA të tipeve D1, D3, D4 dhe D5, shumica e studimeve mbi efektet e drogave të abuzimit dhe të varësisë në trurin e njeriut janë fokusuar në D2R.
Fig 1 Drogat e abuzimit veprojnë në shpërblimin dhe qarqet ndihmëse përmes mekanizmave të ndryshëm, megjithatë, të gjitha ato çojnë në efekte të ngjashme dopaminergjike në VTA dhe NAc. Kështu, stimuluesit nxisin direkt acumbal DA, ndërsa opiatet e bëjnë këtë duke ulur tonin frenues të interneurons GABAergic në DA sinjalizuar si në VTA ashtu edhe në NAc pastaj. Ndërsa mekanizmat e barnave të tjera të abuzimit janë më pak të qarta, ka prova që sugjerojnë që nikotina mund të aktivizojë VTA DA direkt përmes receptorit të acetilkolinës nikotinike (nAChR) në ato neurone dhe në mënyrë indirekte nëpërmjet stimulimit të receptorëve të saj në terminalet e nervave glutamatergike që innervojnë qelizat DA. Alkooli duket se pengon terminalet GABAergic në VTA, duke çuar në disinhibition neuronet DA në VTA. Kanabinoidet veprojnë, ndër të tjera, nëpërmjet aktivizimit të receptorëve CB1 në terminalet nervore glutamatergike dhe GABAergic në NAc, dhe në vetë neuronet NAc. Phencyclidine (PCP) mund të veprojë duke penguar receptorët post-sinaptik NMDA glutamate në NAc. Përveç kësaj, ka disa prova që sugjerojnë që nikotina dhe alkooli gjithashtu mund të ndërveprojnë me rrugët e opiumit endogjen dhe kanabinoid (nuk tregohet). PPT / LDT, pedunculus pontine tegmentum / dengs lateral. Ripublikuar me leje Nestler (2005)
Në njerëzit, studimet PET kanë treguar se disa ilaçe [nxitës (Drevets et al. 2001; Volkow et al. 1999b), nikotina (Brody et al. 2009), alkooli (Boileau et al. 2003) dhe marihuanë (Bossong et al. 2009)] rrit DA në striatum dorsal dhe ventral (ku ndodhet NAc). Këto studime shfrytëzojnë disa radiotracerë, të tillë si [11C] raclopride, që lidhet me D2R, por vetëm kur këto nuk janë DA endogjene (i pabanuar), të cilat në kushte bazë përputhen me 85-90% të D2R striatal (Abi-Dargham et al. 1998). Kështu, një krahasim i [11C] racopridë pas vendimit dhe pas administrimit të drogës mund të na ndihmojë të vlerësojmë zvogëlimin e disponueshmërisë së D2R të shkaktuar nga droga (ose stimuj të tjerë që mund të rrisin DA). Këto ulje në [11C] lidhja racopride janë proporcionale me rritjen e DA (Breier et al. 1997). Këto studime kanë treguar se rritjet e DA-së të drogës në striatum janë proporcionale me intensitetin e përvojës subjektive të euforisë ose "të lartë" [shih rishikimin (Volkow et al. 2009a)] (Fik. 2).
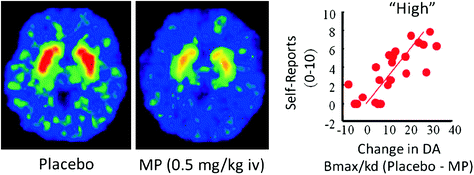
Fig 2 Efektet e metifenidazës intravenoze (MP) në lidhjen e raclopridit dhe marrëdhëniet midis rritjes së striatumeve DA shkaktuar nga MP në striatum dhe vetë raportet e "lartë". Modifikuar nga Volkow et al. (1999b)
Studimet PET gjithashtu kanë zbuluar një marrëdhënie të qartë dhe të drejtpërdrejtë në mes të profilit farmakokinetik të një droge (dmth. Shpejtësia me të cilën hyn dhe e lë trurin) dhe efektet e saj përforcuese. Në mënyrë të veçantë, sa më shpejt një ilaç arrin nivelet e pikut në tru, aq më i fortë është "i lartë" (Volkow et al. 2009a). Për shembull, për një nivel të barabartë të kokainës që arrin trurin (vlerësohet përmes PET), kur kokaina hynte shpejt në tru (smoked ose iv administration), ajo nxiti një "më të lartë" intensive se kur hynte në një shkallë më të ngadaltë (Volkow et al. 2000). Kjo është në përputhje me studimet preklinike që tregojnë një lidhje të ngjashme ndërmjet profilit farmakokinetik të një droge dhe vetive të tij përforcuese (Balster dhe Schuster 1973). unëështë e arsyeshme të supozojmë se një rritje e tillë e papritur dhe e madhe rritet si e shkaktuar nga droga e abuzimit mund të imitojë rritjet e shpejta dhe të mëdha të DA që rezultojnë nga djegie FA fasike që janë shoqëruar në tru me përpunimin e informacionit në lidhje me shpërblimin dhe ngritjen (Schultz 2010). Droga të tilla të shtuara nga droga në NAc mund të jenë të nevojshme për varësinë, por fakti që ato ndodhin edhe në individë jo të varur tregon se ato janë të pamjaftueshme për të shpjeguar përdorimin impulsiv dhe të dobët të drogës karakteristike të varësisë.
Tani ka prova që përgjigjet dopaminergjike të krahasueshme janë të lidhura me shpërblimin e ushqimit dhe se këto mekanizma gjithashtu kanë gjasa të luajnë një rol në konsumimin e tepërt të ushqimit dhe obezitetit. Është e njohur se disa ushqime, veçanërisht ato të pasura me sheqer dhe yndyrë, janë mjaft të dobishme (Lenoir et al. 2007). Ushqimet me kalori të lartë mund të nxisin ushqimin e tepërt (të hahet që nuk ndahet nga nevojat energjitike) dhe të shkaktojë lidhjet e mësuara midis stimulit dhe shpërblimit (kondicionimit). Në terma evolutivë, kjo pronë e ushqimeve të shijshme ishte e dobishme në mjedise ku burimet e ushqimit ishin të pakta dhe / ose jo të besueshme, sepse siguroheshin që ushqimi ishte ngrënë kur ishte në dispozicion, duke mundësuar që energjia të ruhet në trup (si yndyrë) për përdorim në të ardhmen . Për fat të keq, në shoqëritë si tonat, ku ushqimi është i bollshëm dhe vazhdimisht në dispozicion, kjo përshtatje është bërë një detyrim.
Disa neurotransmetues, përfshirë DA, kanabinoidet, opioidet dhe serotonin, si dhe hormonet dhe neuropeptidet e përfshira në rregullimin homeostatik të marrjes së ushqimit, siç janë insulina, oreksina, leptina dhe greliini, janë implikuar në efektet shpërblyese të ushqimit (Atkinson 2008; Cason et al. 2010; Cota et al. 2006). Nga këto, DA është hetuar më së shumti dhe karakterizohet më së miri. Eksperimentet në brejtës kanë treguar se, pas ekspozimit të parë në një shpërblim ushqimi, qitja e DA neuroneve në VTA rritet me një rritje që rezulton në lirimin DA në NAc (Norgren et al. 2006). Në mënyrë të ngjashme, në lëndë të shëndetshme njerëzore me peshë normale, gëlltitja e ushqimit të shijshëm është treguar të lirojë DA në striatumin dorsal në proporcion me vlerësimet e këndshme të vakteve (Small et al. 2003) (Fik. 3). Megjithatë, dhe siç shihet në studimet me abuzuesit e drogës, rritjet e ushqimeve të shkaktuara nga DA vetëm striatum nuk mund të shpjegojnë diferencën midis konsumit normal të ushqimit dhe konsumit të tepërt të ushqimit ushqimor, meqë këto ndodhin edhe në individë të shëndetshëm që nuk hanë tepër. Kështu, siç është rasti për varësinë, përshtatjet në rrjedhën e poshtme ka të ngjarë të përfshihen në humbjen e kontrollit mbi marrjen e ushqimit.
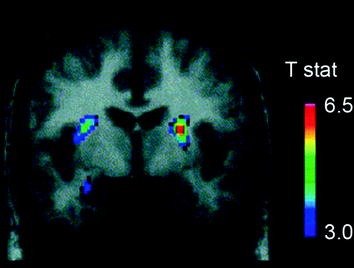
Fig 3 Lirimi i dopamines shkaktohet nga ushqimi. Seksioni coronal nga T-mapi i reduktimeve statistikisht të rëndësishme në [11C] potenciali i lidhjes së raclopridit (BP) pas ushqyerjes. Bar ngjyra përfaqëson vlerat statistikore t. (Ribotohet me leje Small et al. 2003)
3 Imaging DA në përgjigje të drogës dhe në cues të kushtëzuar në varësi
Roli i DA-së në përforcim është më kompleks se vetëm kodimi për shpërblim në vetvete (kënaqësi hedonike); për shembull, stimujt që nxisin rritjen e shpejtë dhe të madhe të DA gjithashtu shkaktojnë reagime të kushtëzuara dhe nxisin motivimin nxitës për t'i blerë ato (Owesson-White et al. 2009). Kjo është e rëndësishme sepse përmes procesit të kondicionimit, stimujt neutralë që janë të lidhura me përforcuesin (nëse një natyror ose një përforcues i drogës) fitojnë vetë aftësinë për të rritur DA në striatum (duke përfshirë NAc) në pritje të shpërblimit, duke krijuar kështu një motivim i fortë për të kërkuar ilaçin (Owesson-White et al. 2009). Sidoqoftë, mekanizmat e shpërblimit dhe të kondicionimit në procesin e varësisë nga droga janë më sfiduese sesa për konsumin e ushqimit, sepse drogat e abuzimit, nëpërmjet efekteve të tyre farmakologjike, aktivizojnë drejtpërdrejt neuronet DA (p.sh., nikotina) ose rritin DA (dmth amfetaminën).
Studimet e imazhit të trurit që krahasuan rritjet e DA-së të nxitura nga medikamentet stimuluese të metilfenidatit (MP) ose amfetaminës (AMPH) në mesin e subjekteve të varur nga kokaina kundrejt kontrolleve treguan një zbutje të dukshme të MP ose AMPH-indukuar të rritjes së DA në striatum (50% më e ulët në abuzuesit e detoksifikuar dhe 80% në abuzuesit aktivë) dhe vetë-raportet e ulëta të efekteve të dobishme të barnave në lidhje me kontrollet që nuk përdorin drogën (Martinez et al. 2007; Volkow et al. 1997) (Fik. 4). Kjo ishte e habitshme që MP dhe AMPH janë farmakologjikisht të ngjashme me kokainën dhe metamfetaminën, respektivisht dhe abuzuesit e drogës nuk mund të bëjnë dallimin mes tyre kur ato administrohen intravenozisht. Meqë reduktimet e dukshme të rritjes së DA-ve të nxitura nga droga janë vërejtur nëse abuzuesit e kokainës ishin detoksifikuar apo jo, kjo tregon që gjendja e tërheqjes nuk është një faktor konfondues (Volkow et al. 2011b). Këto dhe rezultate të ngjashme (Volkow et al. 2009a) janë në përputhje me hipotezën se përgjigja hedonike bëhet e mangët në personat e varur nga droga dhe përforcon më tej nocionin se efektet akut farmakologjike DA për të rritur drogën në NAc nuk mund të shpjegojnë vetë motivimin e rritur për konsumimin e tyre.
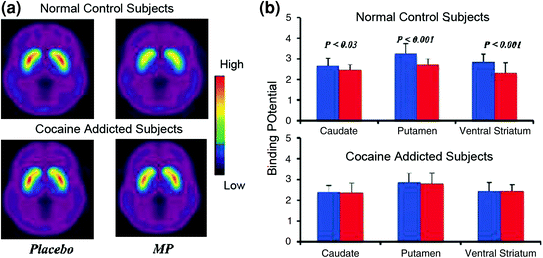
Fig 4 Ndryshimet e DA nxitur nga MP iv në kontrolle dhe në lëndët aktive të kokainës. një imazh mesatar i pandryshueshëm i bidhur (BPND) i [11C] raclopride në lëndët aktive të kokainës (n = 19) dhe në kontrollet (n = 24) testuar pas placebo dhe pas iv MP. b D2R disponueshmëria (BPND) në caudate, putamen dhe striatum ventral pas placebo (blu) dhe pas MP (të kuqe) në kontrolle dhe në lëndët e kokainës. MP reduktoi D2R në kontrolle, por jo në lëndët e kokainës. Vini re se abuzuesit e kokainës tregojnë të dyja zvogëlimet në disponueshmërinë stërhadore D2R bazë (masë placebo) dhe zvogëlohet në lirimin DA kur i jepet MP iv (matur si ulje në disponueshmërinë e D2R nga baseline). Edhe pse mund të pyesim se deri në ç'masë disponueshmëria e ulët striatake D2R në subjektin e kokainës-varur kufizon aftësinë për të zbuluar ulje të mëtejshme nga deputeti, fakti që subjektet e varur nga kokaina tregojnë zvogëlime në disponueshmërinë e D2R kur ekspozohen ndaj cuesit të kokainës, tregon se efektet e zbutura e deputetit në [11C] racloprid të detyrueshme pasqyrojnë uljen e DA lirimit. Ripublikuar me leje (Volkow et al. 1997; Wang et al. 2010)
Përgjigja e neuroneve VTA DA për stimulime të dobishme ndryshon me ekspozimin e përsëritur.
Ndërkohë që qelizat DA ngrenë mbi ekspozimin e parë në një shpërblim të ri, ekspozimi i përsëritur ndaj DA shkakton që neuronet të ndalojnë qitjen me shpërblimin e konsumit dhe zjarrit, në vend kur të ekspozohen ndaj stimujve që parashikojnë shpërblimin (Schultz et al. 1997). Kjo ka gjasa që të mbështetet në rolin e DA-së në mësim dhe kondicionim. Në të vërtetë, sinjalizimi FA fasik i induktuar mund të shkaktojë eventualisht neuroadaptime në qarqet ndihmëse që lidhen me formimin e zakonit dhe kondicionimin e sjelljes. TNdryshimet e tilla janë nxitur kryesisht nga sinjalizimi i D1R dhe ndryshimet sinaptike në receptorët NMDA dhe AMPA të moduluar me glutamat (Luscher dhe Malenka 2011; Zweifel et al. 2009). Rekrutimi i këtyre qarqeve është i rëndësishëm në përparimin e sëmundjes, sepse përgjigjet e kushtëzuara që ndihmojnë ndihmojnë në shpjegimin e dëshirës intensive për drogën (dëshira) dhe përdorimit të dhunshëm që ndodh kur subjektet e varur janë të ekspozuar ndaj cuesve të lidhura me drogën. Kjo hipotezë është në përputhje me vëzhgimet e pavarura (Volkow et al. 2006b; Wong et al. 2006) që tregojnë fuqinë e ekspozimit të sugjerimeve shoqëruese të kokainës për të rritur nivelet e DA në striatumin dorsal dhe shkaktojnë një rritje të njëtrajtshme në përvojën subjektive të dëshirës në abuzuesit e kokainës të detoksifikuar (Fig. 5). Që nga striatum dorsal luan një rol në mësimin e zakonit (Belin et al. 2009; Yin et al. 2004), shoqata ka të ngjarë të pasqyrojë forcimin e zakoneve ashtu si zhvillohet kronika e varësisë. Kjo sugjeron që një ndërprerje bazë në varësi mund të lidhet me përgjigjet e kushtëzuara nga DA-të që rezultojnë në zakone që çojnë në dëshirë intensive dhe në konsum kompulsiv të drogës. Është interesant fakti se, në përdorimin aktiv të subjekteve të varur nga kokaina, rritjet DA të shkaktuara nga cues të kushtëzuar duket të jenë edhe më të mëdha se ato të prodhuara nga vetë droga stimuluese siç vlerësohet në dy grupe të ndara të subjekteve (Volkow et al. 2011b, 2006b), duke sugjeruar që përgjigjet e kushtëzuara mund të nxisin sinjalizimin DA që mban motivimin për të marrë ilaçin edhe kur efektet e saj farmakologjike duket të reduktohen. Kështu, megjithëse barnat fillimisht mund të nxisin ndjenjat e shpërblimit të menjëhershëm nëpërmjet lirimit DA në striatumin e barkut, me përdorim të përsëritur dhe si zhvillohet zakon, duket se ka një ndryshim nga droga në stimul të kushtëzuar. Sipas studimeve në kafshë laboratorike, projeksionet glutamatergike nga lëvore paraballore dhe nga amigdala në VTA / SN dhe NAc ndërmjetësojnë këto përgjigje të kushtëzuara (Kalivas 2009). Në këtë mënyrë, parashikimi i thjeshtë i një shpërblimi përfundimisht mund të bëhet shpërblimi që motivon sjelljen e nevojshme për konsumin e drogës (ose ushqimit).
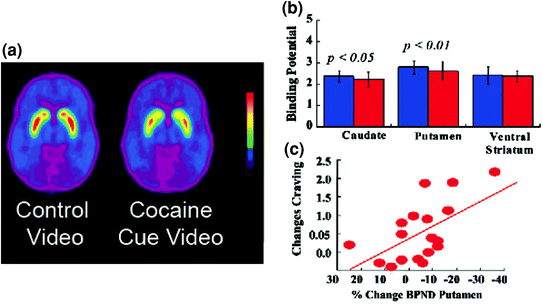
Fig 5 Ndryshimet e DA shkaktuara nga cues të kushtëzuar në lëndët aktive të kokainës. një imazh mesatar i pandryshueshëm i detyrueshëm (BPND) i [11C] raclopride në subjektet e kokainës (n = 17) testuar gjatë shikimit të një videoje neutrale (skenave natyrore) dhe gjatë shikimit të videove të kokainës (subjektet që administrojnë kokainë). b D2R disponueshmëria (BPND) në kauzë, putamen dhe striatumin e barkut për videon neutrale (blu) dhe videokonferencën e kokainës (e kuqe). Kokaine e kokainës ulën D2R në caudate dhe putamen. c Korrelacionet ndërmjet ndryshimeve në D2R (duke reflektuar rritjet e DA) dhe vetë-raportet e dëshirës së kokainës të nxitura nga video kokainë-cues. Modifikuar nga ref. (Volkow et al. 2006b)
INë mënyrë të veçantë, ky lloj i "kaloni" funksional është raportuar edhe për përforcuesit natyrorë, të cilët ka gjasa të nxisin një ndryshim të barazvlefshëm dhe gradual në rritjet e DA, nga barku në rajonet më dorsale të striatumit gjatë tranzicionit nga një stimul novelë që është në thelb duke shpërblyer me atë të cuesave të shoqëruara që e parashikojnë atë. Ky tranzicion është përcjellë përmes sinjalizimit DA, i cili duket të kodojë për një "gabim parashikimi shpërblimi" (Schultz 2010). Aferencat e gjera glutamatergike te DA neuronet nga rajonet e perfshira ne perpunimin e shqetesimit (insula ose korteksit primar gustatory), homeostatic (hypothalamus), shpërblim (NAc), emocional (amygdala dhe hippocampus), dhe multimodal (cortex orbitofrontal për ndarjen e ndarjes) informacioni , modifikojnë aktivitetin e tyre në përgjigje të shpërblimeve dhe cues të kushtëzuar (Geisler and Wise 2008). Më konkretisht, parashikimet nga amygdala dhe korteksit orbitofrontal (OFC) tek DA neuronet dhe NAc janë të përfshira në përgjigjet e kushtëzuara të ushqimit (Petrovich 2010). Në të vërtetë, studimet e imazhit treguan se kur subjektet meshkuj jo të trashë u kërkuan të pengonin dëshirën e tyre për ushqim - ndërkohë që ishin të ekspozuar ndaj cuesve të ushqimit - ata shfaqnin zvogëlim të aktivitetit metabolik në amygdala dhe OFC (si në hippocampus), insula dhe striatum, dhe se zvogëlimet në OFC u shoqëruan me zvogëlimin e dëshirës ushqimore (Wang et al. 2009). Një ndalim i ngjashëm i aktivitetit metabolik në OFC (dhe gjithashtu në NAc) është vërejtur në abuzuesit e kokainës kur u është kërkuar të pengojnë dëshirën e tyre të drogës pas ekspozimit ndaj kokainës (Volkow et al. 2009b).
Gjithsesi, shfaqja e dëshirave të tilla të fuqishme, të cilat për ushqim ndodhin edhe në individë të shëndetshëm që nuk ngopen, nuk do të ishin aq shkatërruese sikur të mos shoqëroheshin me deficite në rritje në aftësinë e trurit për të penguar sjelljet keqdashëse.
4 Ndikimi i mosfunksionimit në kontrollin inhibitor
Aftësia për të penguar përgjigjet e paragjykuara është e detyruar të kontribuojë në aftësinë e një individi për të shmangur përfshirjen në sjellje të papërshtatshme, të tilla si marrja e barnave apo të hahet pas pikës së ngopjes dhe duke rritur kështu cenueshmërinë e tij ndaj varësisë (ose trashje) (Volkow dhe Fowler 2000; Volkow et al. 2008a).
Studimet e PET kanë zbuluar reduktime të konsiderueshme në disponueshmërinë e D2R në striatumin e subjekteve të varur që vazhdojnë për muaj pas detoksifikimit të zgjatur [rishikuar në (Volkow et al. 2009a)]. Në mënyrë të ngjashme, studimet preklinike në primat brejtës dhe jo-njerëzor kanë treguar se ekspozimet e përsëritura të drogës shoqërohen me reduktime në nivelet stigmatike të D2R (Nader et al. 2006; Thanos et al. 2007; Volkow et al. 2001). Në striatum, D2Rs ndërmjetësojnë sinjalizimin në shtegun indirekt striatum që modulon rajonet prefrontale; dhe ulja e tij ka treguar të rrisë sensibilizimin ndaj efekteve të drogës në modelet e kafshëve (Ferguson et al. 2011). Në njerëzit të varur nga droga, reduktimi i D2R striatum është i lidhur me aktivitetin e zvogëluar të rajoneve paraballore siç dëshmohet nga ulja e metabolizmit të glukozës bazë (një marker i funksionit të trurit) në OFC, gyrus anterior cingulate (ACC) dhe korteksit prefrontal dorsolateral (DLPFC ) (Volkow et al. 2001, 1993, 2007) (Fik. 6). Duke qenë se OFC, ACC dhe DLPFC janë të përfshirë me atributet e ndarjes, kontrollin frenues / rregullimin e emocioneve dhe vendimmarrjen, përkatësisht, është vënë në pikëpyetje që rregullimi i tyre i parregullt nga D2R-i ndërmjetësimit të DA në subjekte të varur mund të jetë në themel të rritjes së motivimit të drogës në sjelljen e tyre dhe humbjes së kontrollit mbi marrjen e drogës (Volkow dhe Fowler 2000). Përveç kësaj, për shkak se dëmtimet në OFC dhe ACC janë të lidhura me sjelljet e detyrueshme dhe impulsivitetin (Fineberg et al. 2009), Modulimi i dëmtuar i DA-së i këtyre rajoneve ka të ngjarë të kontribuojë në marrjen e drogës impulsive dhe impulsive të parë në varësi (Goldstein dhe Volkow 2002). Në të vërtetë, në abuzuesit me metamfetaminë, D2R e striatumit të ulët u shoqërua me impulsivitet (Lee et al. 2009), dhe parashikoi gjithashtu administrimin e detyruar të kokainës në brejtës (Everitt et al. 2008). Një skenar i kundërt, në të cilin një dobësi fillestare për përdorim të drogës preexiston në rajonet paraburgore dhe ku përdorimi i përsëritur i drogës shkakton ulje të mëtejshme në D2R striatal, është gjithashtu e mundur. Në të vërtetë, një studim i bërë në subjekte që, megjithëse kishin një rrezik të lartë për alkoolizëm (histori pozitive familjare të alkoolizmit) nuk ishin alkoolikë, zbuluan një disponueshmëri më të lartë se normale stigmatike D2R që ishte e lidhur me metabolizmin normal në OFC, ACC dhe DLPFC (Volkow et al. 2006a). Kjo sugjeron që në këto subjekte në rrezik për alkoolizëm, funksioni normal paraballor ishte i lidhur me sinjalizimin e zgjeruar striatal D2R, i cili mund t'i ketë mbrojtur ata nga abuzimi me alkoolin.
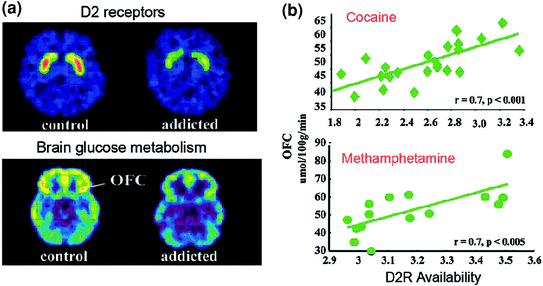
Fig 6 Korrelacionet midis disponueshmërisë stigmatike D2R dhe metabolizmit në rajonet e trurit prefrontal. një imazhe aksiale të trurit për një kontroll dhe për një subjekt kokainë të varur për imazhet bazë e disponueshmërisë D2R në striatum (marrë me [11C] raclopride) dhe të metabolizmit të glukozës në tru në OFC (të marra me [18FDG). b Korrelacionet midis D2R striatal dhe metabolizmit në OFC në subjektet e kokainës dhe të varur nga metamfetamina. Ribotuar nga Volkow et al. (2009a) Copyright (2009), me lejen e Elsevier
Parashikueshme, prova e disregulimit në qarqet e kontrollit është gjetur gjithashtu në mesin e individëve të trashë. Të dy studimet preklinike dhe klinike kanë dhënë dëshmi të zvogëlimit të sinjalizimit striatal D2R, i cili, siç u përmend më lart, lidhet me shpërblimin (NAc), por edhe me krijimin e zakoneve dhe rutinave (striatum dorsal) në obesity (Geiger et al. 2009; Wang et al. 2001). E rëndësishmja, zvogëlimi i disponueshmërisë striatake të D2R ka qenë e lidhur me marrjen e ushqimit ushqimor në bërrylat e trashë (Johnson dhe Kenny 2010) dhe me ulje të aktivitetit metabolik në OFC dhe ACC në njerëz të trashë (Volkow et al. 2008b) (Fik. 7a-c). Duke pasur parasysh se mosfunksionimi në OFC dhe ACC rezulton në kompulsivitet [shih rishikimin (Fineberg et al. 2009)], kjo mund të jetë pjesë e mekanizmit me të cilin sinjalizimi i ulët striatal D2R lehtëson hyperphagia (Davis et al. 2009). Përveç kësaj, pasi sinjalizimi i lidhur me D2R ka gjasa të zvogëlojë ndjeshmërinë ndaj shpërblimeve të tjera natyrore, ky deficit në personat e trashë mund të kontribuojë gjithashtu në mbingarkimin kompensues (Geiger et al. 2008).
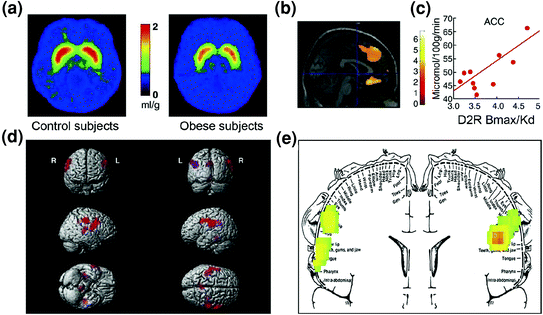
Fig 7 Hyperphagia mund të rezultojë nga një makinë për të kompensuar një qark shpërblimi të dobësuar (të përpunuara përmes qarqeve kortikostriatale të rregulluara dopamine) të kombinuara me një ndjeshmëri të rritur ndaj palatability (vetitë hedonike të ushqimit të përpunuara pjesërisht përmes korteksit somatosensor). a Imazhe mesatare për disponueshmërinë e receptorëve DA D2 (D2R) në kontrollet (n = 10) dhe në subjektet me dhjamë të sëmurë (n = 10). b Rezultatet nga (Hartësimi Parametrik Statistikor) SPM duke identifikuar zonat në tru ku D2R shoqërohej me metabolizmin e glukozës, këto përfshinin OFC mesatare, ACC dhe PFC dorsolaterale (rajoni nuk tregohet). c Pjerrësia e regresionit midis D2R striatal dhe aktivitetit metabolik në ACC në subjektet mbipeshe. d Imazhe SPM të dhëna tre-dimensionale që tregojnë zonat me metabolizëm më të lartë në dhjamë sesa tek subjektet e dobëta (P <0.003, e pa korrigjuar). e Rezultatet e koduara me SPM të ngjyrave shfaqen në një plan koronal me një diagram të mbivendosur të homunculusit somatosensor. Rezultatet (vlera z) paraqiten duke përdorur shkallën e ylberit ku e kuqe> e verdhë> jeshile. Kur krahasohen me subjekte të dobëta, subjektet e dhjamit kishin metabolizëm fillestar më të lartë në zonat somatosensore ku përfaqësohen goja, buzët dhe gjuha dhe të cilat përfshihen në përpunimin e shijes së ushqimit. Modifikuar, me leje, nga Volkow et al. (2008a) (a-c) dhe Wang et al. (2002) (d, e)
Kjo hipotezë është në përputhje me provat preklinike që tregojnë se zvogëlimi i aktivitetit DA në VTA rezulton në një rritje dramatike të konsumit të ushqimeve me yndyrë të lartë (Stoeckel et al. 2008). Në mënyrë të ngjashme, krahasuar me individët me peshë normale, individët e dhjamosur të cilët u paraqitën me fotografi të ushqimeve me kalori të lartë (stimulimet për të cilat ato janë të kushtëzuara) shfaqën një rritje të aktivizimit nervor në rajonet që janë pjesë e qarqeve shpërblimi dhe motivimi (NAc, striatum dorsal, OFC , ACC, amygdala, hippocampus dhe insula) (Killgore dhe Yurgelun-Todd 2005). Në kontrast, në kontrollet normale të peshës, aktivizimi i ACC dhe OFC (rajonet e përfshira në atributet e spitaleve që projektojnë në NAc) gjatë paraqitjes së ushqimit me kalori të lartë u gjetën të lidhen negativisht me indeksin e tyre të trupit (BMI) ( Stice et al. 2008b). Kjo sugjeron një ndërveprim dinamik mes sasisë së ushqimit të ngrënë (që reflektohet pjesërisht në BMI) dhe reaktivitetit të rajoneve shpërblim për ushqim me kalori të lartë (të pasqyruar në aktivizimin e OFC dhe ACC) në individë normalë të peshës, e cila humbet në trashje.
Çuditërisht, individët e trashë shfaqën më pak aktivizim të qarqeve të shpërblimit nga konsumimi aktual i ushqimit (i referuar si shpërblimi ushqimor i konsumueshëm) sesa individët e dobët, ndërsa ata treguan një aktivizim më të madh të rajoneve somatosensorale kortique që përpunonin palatabilitetin kur ata parashikonin konsumimin (Stice et al. 2008b). Vëzhgimi i fundit i korrespondonte rajoneve ku një studim i mëparshëm kishte zbuluar një aktivitet të zgjeruar në subjektin e trashë të testuar në bazë (jo stimulim) (Wang et al. 2002) (Fik. 7d, e). Një aktivitet i zgjeruar i rajoneve që përpunojnë palatabilitetin mund të bëjnë që subjektet e dhjamit të favorizojnë ushqimin mbi përforcuesit e tjerë natyrorë, ndërsa zvogëlimi i aktivizimit të caqeve dopaminergjike nga konsumimi aktual i ushqimit mund të çojë në mbipërdorim si një mjet për të kompensuar sinjalizimin e dobët të D2R-mediuara (Stice et al. 2008a). Kjo përgjigje e reduktuar e rrjetit të shpërblimit për konsum ushqimor në lëndë të trasha kujton rritjen e zvogëluar të DA-së të shkaktuar nga konsumimi i drogës tek individët e varur kur krahasohet me subjektet që nuk janë të varur.
Lëkura paraballore (PFC) luan një rol vendimtar në funksionin ekzekutiv, duke përfshirë kontrollin frenues (Miller dhe Cohen 2001). Këto procese modulohen nga D1R dhe D2R (me sa duket edhe D4R) dhe kështu, aktiviteti i zvogëluar në PFC, si në varësi ashtu edhe në obezitet, ka të ngjarë të kontribuojë në kontroll të dobët dhe në kompulsivitet të lartë. Disponueshmëria më e ulët se normale e D2R në striatum të individëve obezë, e cila është shoqëruar me aktivitet të reduktuar në PFC dhe ACC (Volkow et al. 2008b) prandaj ka të ngjarë të kontribuojë në kontrollin e tyre të mangët mbi marrjen e ushqimit. Në të vërtetë, korrelacioni negativ mes BMI dhe D2R striatal raportohet në obezë (Wang et al. 2001) dhe në mbipeshë (Haltia et al. 2007a) individët e mbështesin këtë. Një kuptim më i mirë i mekanizmave që çojnë në funksionin e dëmtuar të PFC në obezitetin (ose varësinë) mund të lehtësojë zhvillimin e strategjive për të përmirësuar, ose ndoshta edhe për të ndryshuar, dëmtime specifike në domenet kognitive kryesore. Për shembull, skontimi i vonesës, që është tendenca për të zhvlerësuar një shpërblim si një funksion i vonesës kohore të lindjes së saj, është një nga operacionet kognitive më të hulumtuara gjerësisht në lidhje me çrregullimet që lidhen me impulsivitetin dhe kompulsivitetin. Zbritja e vonuar është hetuar më së shumti në abuzuesit e drogës të cilët shfaqin një preferencë të ekzagjeruar për të ardhurat e vogla por të menjëhershme mbi shpërblimet e mëdha por të vonuara (Bickel et al. 2007). Megjithatë, disa studime të kryera me individë të trashë kanë zbuluar gjithashtu dëshmi të një preference për shpërblime të larta, të menjëhershme, pavarësisht nga një shans më i madh për të vuajtur humbje më të larta në të ardhmen (Brogan et al. 2010; Weller et al. 2008). Dhe kohët e fundit, një tjetër studim gjeti një korrelacion pozitiv midis BMI dhe skontimit hiperbolik, ku pagesat negative të ardhshme zbriten më pak se pagesat pozitive të ardhshme (Ikeda et al. 2010). Interesant, vonesa e vonesës duket të varet nga funksioni i striatumit të ventralit (Gregorios-Pippas et al. 2009) dhe të PFC, duke përfshirë OFC anësore (Bjork et al. 2009), dhe është e ndjeshme ndaj manipulimeve DA (Pine et al. 2010). Në mënyrë të veçantë, rritja e sinjalizimit DA (me trajtimin L DOPA) rriti impulsivitetin dhe skontimin kohor.
5 Përfshirja e Qarqeve të Motivimit
Sinjalizimi dopopanergjik gjithashtu modulon motivimin. Tiparet e sjelljes si energjia, këmbëngulja dhe investimi i një përpjekjeje të vazhdueshme drejt arritjes së një qëllimi, janë të gjitha subjekt i modulimit nga DA duke vepruar përmes disa rajoneve të synuara, duke përfshirë NAc, ACC, OFC, DLPFC, amygdala, striatum dorsal dhe pallidum ventral (Salamone et al. 2007). Dysregulimi i sinjalizimit të DA-së shoqërohet me motivim të shtuar për të blerë drogë, një shenjë e varësisë dhe për këtë arsye individët e varur nga droga shpesh angazhohen në sjellje ekstreme për të marrë droga, madje edhe kur ato sjellin pasoja të rënda dhe të padëshiruara (Volkow dhe Li 2005). Për shkak se marrja e drogës bëhet motivi kryesor motivues në varësinë nga droga (Volkow et al. 2003), subjektet e varur janë të ngjallur dhe të motivuar nga procesi i marrjes së drogës, por tentojnë të bëhen të tërhequr dhe apatikë kur ekspozohen ndaj aktiviteteve jo të lidhura me drogën. Ky ndryshim është studiuar duke krahasuar modelet e aktivizimit të trurit që ndodhin me ekspozimin ndaj cues të kushtëzuar me ato që ndodhin në mungesë të këtyre cueseve. Në ndryshim nga zvogëlimi i aktivitetit paraballor të raportuar në abuzuesit e kokainës të detoksifikuar kur nuk stimulohet me ilaçe të drogës ose drogës [shih rishikimin (Volkow et al. 2009a)], këto rajone prefrontale aktivizohen kur abuzuesit e kokainës janë të ekspozuar ndaj stimujve që nxisin nxitjet (ose drogat ose cues) (Grant et al. 1996; Volkow et al. 1999a; Wang et al. 1999). Ky rezultat kujton vëzhgimin që abuzuesit e kokainës, të studiuar menjëherë pas një episodi të bingingut të kokainës, treguan një rritje të aktivitetit metabolik në OFC dhe ACC (gjithashtu striatum dorsal) që shoqërohej me dëshirë (Volkow et al. 1991).
Për më tepër, kur përgjigjet ndaj deputetit iv janë krahasuar në mes të personave të varur nga kokainë dhe jo të varur, të parët janë përgjigjur me rritjen e metabolizmit në ACC dhe ventralin OFC (një efekt i lidhur me dëshirë), ndërsa kjo e fundit ka treguar përgjigjen e kundërt, metabolizmin në këto rajone (Volkow et al. 2005). Kjo sugjeron që aktivizimi i këtyre zonave prefrontale me ekspozimin e drogës mund të jetë specifike për varësinë dhe të shoqëruar me dëshirën e zgjeruar për drogën. Përveç kësaj, një studim që nxiti subjektet e varur nga kokaina që me qëllim të pengonin dëshirën kur u ekspozuan ndaj indikacioneve të drogës treguan se ata subjekte që ishin të suksesshëm në pengimin e shfaqjes së dëshirës zvogëloi metabolizmin në OFC mediale (që përpunon vlerën motivuese të një përforcuesi) dhe NAc (që parashikon shpërblim) (Volkow et al. 2009b). Këto rezultate më tej vërtetojnë përfshirjen e OFC, ACC, dhe striatum në motivimin e zgjeruar për të blerë drogën e parë në varësi.
Parashikueshme, OFC ka qenë gjithashtu i implikuar në atribuimin e vlerës së theksit tek ushqimi (Grabenhorst et al. 2008; Rolls dhe McCabe 2007), duke ndihmuar për të vlerësuar këndshmërinë dhe shijshmërinë e pritshme si një funksion i kontekstit të saj. Studimet me PET me FDG për të matur metabolizmin e glukozës në tru në individë me peshë normale kanë raportuar se ekspozimi ndaj ushqimeve ka rritur aktivitetin metabolik në OFC, i cili ishte një efekt i lidhur me perceptimin e urisë dhe dëshirën për ushqim (Wang et al. 2004). Aktivizimi i zgjerimit të OFC nga stimulimi i ushqimit ka të ngjarë të reflektojë efektet dopaminergjike të rrjedhës së poshtme dhe të marrë pjesë në përfshirjen e DA-së në përzjerjen e konsumit të ushqimit. OFC luan një rol në mësimin e shoqatave të stimulimit-përforcimit dhe kondicionimit (Cox et al. 2005; Gallagher et al. 1999), mbështet ushqimin e kushtëzuar të nxitjes (Weingarten 1983) dhe ndoshta kontribuon në ngrirjen e ushqimit pa marrë parasysh sinjalet e urisë (Ogden dhe Wardle 1990). Në të vërtetë, mosfunksionimi i OFC ka qenë i lidhur me overeating (Machado dhe Bachevalier 2007).
Përkundër disa mospërputhjeve midis studimeve, të dhënat e imazhit të trurit gjithashtu mbështesin nocionin se ndryshimet strukturore dhe funksionale në rajonet e trurit të implikuara në funksionin ekzekutiv (duke përfshirë kontrollin frenues) mund të shoqërohen me BMI të lartë në individë të ndryshëm të shëndetshëm. Për shembull, një studim i MRI i bërë në gratë e moshuara, duke përdorur morfometrinë me bazë voxel, zbuloi një korrelacion negativ midis vëllimeve të BMI dhe vëllimit të ngjyrës gri (duke përfshirë rajonet frontale), të cilat, në OFC, ishin të lidhura me funksionin ekzekutiv të dëmtuar (Walther et al. 2010). Duke përdorur PET për të matur metabolizmin e glukozës në tru në kontrollet e shëndetshme, ne raportuam një korrelacion negativ ndërmjet BMI dhe aktivitetit metabolik në DLPFC, OFC dhe ACC. Në këtë studim, aktiviteti metabolik në rajonet paraballore parashikoi performancën e subjekteve në testet e funksionit ekzekutiv (Volkow et al. 2009c). Në mënyrë të ngjashme, një studim spektroskopik i rezonancës magnetike bërthamore (NMR) në moshën e mesme të shëndetshme dhe kontrollet e moshuara tregoi se BMI ishte lidhur negativisht me nivelet e N-acetil-aspartat (një shënues i integritetit nervor) në korteksin frontal dhe ACC (Gazdzinski et al. 2008; Volkow et al. 2009c).
Studimet e imazhit të trurit që krahasojnë individë të trashë dhe të ligët kanë raportuar gjithashtu dendësi më të ulët të lëndës gri në rajonet ballore (operculum ballor dhe gyrus i mesëm ballor) dhe në gyrus post-qendror dhe putamen (Pannacciulli et al. 2006). Një tjetër studim, nuk gjeti asnjë ndryshim në vëllimet e lëndës gri midis subjekteve të trashë dhe të dobët, megjithatë, ai bëri një korelacion pozitiv midis volumit të bardhë në strukturat e trurit të bazës dhe raportit të belit në hip, një trend që u përmbys pjesërisht nga dieta (Haltia et al. 2007b). Është interesante që zonat kortikale, si DPFC dhe OFC që janë përfshirë në kontrollin inhibitor, janë gjetur të aktivizohen në dietat e suksesshëm në përgjigje të konsumit të ushqimit (DelParigi et al. 2007), duke sugjeruar një objektiv të mundshëm për rikualifikimin e sjelljes në trajtimin e obezitetit (dhe gjithashtu në varësi).
6 Përfshirja e Circuitry Interoceptive
Studimet neuroimaging kanë zbuluar se insula e mesme luan një rol kritik në dëshirat për ushqim, kokainë dhe cigare (Bonson et al. 2002; Pelchat et al. 2004; Wang et al. 2007). Rëndësia e insula është nënvizuar nga një studim që tregoi se duhanpirësit me dëmtim të këtij rajoni (por jo kontrollin e duhanpirësve që kishin pësuar lezione extra-insulare) ishin në gjendje të ndalnin pirjen e duhanit me lehtësi dhe pa përjetuar as dëshirë apo recetë (Naqvi et al . 2007). Insula, veçanërisht në rajonet e saj më të mëparshme, lidhet reciprokisht me disa rajone limbike (p.sh., kortekti parental paralel, amygdala dhe striatum ventral) dhe duket se ka një funksion interoceptiv, duke integruar informacionin autonom dhe visceral me emocionet dhe motivimin, duke siguruar kështu ndërgjegjësimi i ndërgjegjshëm i këtyre nxitjeve (Naqvi dhe Bechara 2009). Në të vërtetë, studimet e lesionit të trurit sugjerojnë se PFC dhe insula ventromediale janë komponentë të domosdoshëm të qarqeve të shpërndara që mbështesin vendimmarrjen emocionale (Clark et al. 2008). Në përputhje me këtë hipotezë, studimet e imazhit vazhdimisht tregojnë aktivizimin diferencial të insulave gjatë dëshirës (Brody et al. 2009; Goudriaan et al. 2010; Naqvi dhe Bechara 2009; Wang et al. 1999). Prandaj, reaktiviteti i këtij rajoni të trurit është sugjeruar për të shërbyer si një biomarker për të ndihmuar parashikimin e rikthimit (Janes et al. 2010).
Insula është gjithashtu një zonë gustatory primare, e cila merr pjesë në shumë aspekte të sjelljeve të hahet, të tilla si shije. Përveç kësaj, insula rostral (e lidhur me korteksin e shijes primare) ofron informacion për OFC që ndikon në përfaqësimin e tij multimodal të kënaqësisë ose vlerës së shpërblimit të ushqimit në hyrje (Rolls 2008). Për shkak të përfshirjes së insula në sensin interoceptive të trupit, në vetëdijen emocionale (Craig 2003) dhe në motivim dhe emocion (Rolls 2008), mund të pritet një kontribut i dëmtimit insular në obezitetin. Në të vërtetë, shtrëngimi gastrik rezulton në aktivizimin e insula të pasmë, e cila ka gjasa të pasqyrojë rolin e saj në ndërgjegjësimin e shteteve të trupit (në këtë rast plotësia) (Wang et al. 2008). Për më tepër, në subjektet e varfër, por jo në obezitet, distention gastrike rezultuar në aktivizimin e amygdala dhe deaktivizimin e insula anterior (Tomasi et al. 2009). Mungesa e reagimit amygdala në subjektet e trashë mund të pasqyrojë një vetëdije ndërhyrëse të shtrënguar të gjendjeve trupore të lidhura me ngopje (stomaku i plotë). Megjithëse modifikimi i aktivitetit insular nga ana e DA është hetuar dobët, dihet se DA është e përfshirë në përgjigjet ndaj provës së ushqimeve të shijshme që ndërmjetësohen përmes insulave (Hajnal dhe Norgren 2005). Studimet e imazhit të njeriut kanë treguar se ushqimi i shijshëm i shijshëm aktivizoi zonat insula dhe midbrain (DelParigi et al. 2005; Frank et al. 2008). Megjithatë, sinjalizimi DA mund të jetë gjithashtu i nevojshëm për të ndjerë përmbajtjen kalori të ushqimit. Për shembull, kur gratë me peshë normale shijonin një ëmbëlsues me kalorive (saharoze), të dyja zonat insula dhe dopaminergic midbrain u aktivizuan, ndërsa provimi i një sheqer diabeti pa kalori (sucralose) aktivizoi vetëm insula (Frank et al. 2008). Subjektet e trashë shfaqin aktivizim më të madh insulare se kontrollet normale kur provojnë një vakt të lëngshëm që përbëhet nga sheqeri dhe yndyra (DelParigi et al. 2005). Në të kundërt, subjektet që janë shëruar nga anorexia nervosa tregojnë më pak aktivizim në insula kur provojnë saharozë dhe asnjë shoqatë të ndjenjave të këndshme me aktivizimin insulare siç vërehet në kontrollet normale (Wagner et al. 2008). Kur kombinohen, këto rezultate e bëjnë të mundur që disregulimi i insulave në përgjigje të stimujve të shijes mund të përfshihet në kontrollin e dëmtuar të sjelljeve të ndryshme joshëse.
7 Circuitry of Aversion
Siç u përmend më parë, trajnimi (kushtëzimi) në një shenjë që parashikon shpërblimin çon në qelizat dopaminergjike që qisin në përgjigje të parashikimit të shpërblimit dhe jo të shpërblimit vetë. Nga ana tjetër, dhe në përputhje me këtë logjikë, është vërejtur se qelizat dopaminergike do të zjarrit më pak se normale nëse shpërblimi i pritshëm nuk arrin të materializohet (Schultz et al. 1997). Dëshmitë kumulative (Christoph et al. 1986; Lisoprawski et al. 1980; Matsumoto dhe Hikosaka 2007; Nishikawa et al. 1986) tregon habenula si një nga rajonet që kontrollon zvogëlimin e qitjes së qelizave dopaminergike në VTA që mund të ndjekin dështimin për të marrë një shpërblim të pritshëm (Kimura et al. 2007). Kështu, një ndjeshmëri e shtuar e habenulës, si rezultat i ekspozimeve kronike të drogës, mund të jetë një reagim më i madh ndaj cuesve të drogës. Në të vërtetë, aktivizimi i habenulës, në subjektet e varur nga kokaina, është shoqëruar me rikthimin e sjelljes ndaj marrjes së medikamenteve pas ekspozimit të sugjerimeve (Brown et al. 2011; Zhang et al. 2005). Në rastin e nikotinës, receptorët nikotinikë të α5 në habenula duket të modifikojnë përgjigjet arancionale në doza të mëdha të nikotinës (Fowler et al. 2011); dhe receptorët α5 dhe α2 në habenula janë të implikuar në tërheqjen e nikotinës (Salas et al. 2009). Për shkak të reagimit të kundërt të habenulës me atë të DA neuroneve për shpërblim (deaktivizim) dhe aktivizimit të tij pas ekspozimit ndaj stimujve ariaktiv, ne i referohemi këtu sinjalizimit habenula si një që përcjell një input "antireward".
Habenula duket se luan një rol të ngjashëm sa i përket shpërblimit ushqimor. Një dietë shumë e shijshme ushqimore mund të nxisë trashje në minjtë, me rritjen e peshës që korrespondon me rritjen e peptidës μ-opioid që lidhet në amygdala bazolaterale dhe basomediale. Interesante, habenula mesatare tregoi një lidhje të dukshme më të lartë të peptideve μ-opioid (me afërsisht 40%) pas ekspozimit ndaj ushqimit të shijshëm në minjtë që fitonin peshë (ato që konsumonin më shumë ushqime), por jo në ato që nuk kishin (Smith et al. 2002). Kjo sugjeron që habenula mund të përfshihet në ngrënie në kushtet e disponueshmërisë së ushqimit të këndshëm. Për më tepër, neuronet në bërthamën tegmentale rostromediale, të cilat marrin një kontribut të madh nga habenula laterale, projektojnë tek neuronet VTA DA dhe aktivizohen pas privimit të ushqimit (Jhou et al. 2009). Këto gjetje janë në përputhje me një rol për habenulën në ndërmjetësimin e përgjigjeve ndaj stimujve aversivë ose shteteve të tilla si ato që ndodhin gjatë dieting ose tërheqjes së drogës.
Përfshirja e habenulës si qendër shpërndarëse brenda rrjeteve emocionale është në përputhje me modelet e mëparshme teorike të varësisë që pasoi reagimet sensibilizuese anti-shpërblim (të ndërmjetësuar përmes ndjeshmërisë së shtuar të amigdalës dhe rritjes së sinjalizimit edhe pse faktori lëshues i kortikotropinës) si futje e drogës në varësi (Koob dhe Le Moal 2008). Përgjigjet e ngjashme antireward mund të kontribuojnë gjithashtu në konsum të tepërt të ushqimit në trashje.
8 Shpërblimi i drogës patologjike dhe ushqimit: Një model i përditësuar i punës
Taftësia për t'i rezistuar thirrjes për të përdorur një drogë ose për të ngrënë në pikën e ngopjes kërkon funksionimin e duhur të qarqeve nervore të përfshira në kontrollin nga lart poshtë në kundërshtim me përgjigjet e kushtëzuara që parashikojnë shpërblimin nga marrja e ushqimit / drogës dhe dëshira për të gëlltitur ushqim / drogës. Këtu, kemi theksuar gjashtë nga këto qarqe: shpërblimi / shpalosja, kondicionimi / zakonet, kontrolli frenues / funksioni ekzekutiv, motivimi / ngasja, ndërhyrja, dhe shmangia e aversionit / reaktiviteti i stresit (Fig. 8). Bazuar në të dhënat e imazhit të paraqitur këtu, ne supozojmë se është mospërputhje midis pritjes për efektet e drogës / ushqimit (përgjigjet e kushtëzuara) dhe efekteve neurofiziologjike të pista që mbajnë marrjen e barnave ose mbiprodhimin e ushqimeve në përpjekje për të arritur shpërblimi i pritshëm. Gjithashtu, nëse testohen gjatë periudhave të hershme ose të zgjatura të abstinencës / dieting, subjektet e varur / obezë tregojnë D2R më të ulët në striatum (përfshirë NAc), të cilat shoqërohen me zvogëlimin e aktivitetit bazë në rajonet e trurit frontal të implikuar në atributet e ndarjes (korteksit orbitofrontal) dhe frenues kontrollit (ACC dhe DLPFC), ndërprerja e të cilave rezulton në kompulsivitet dhe impulsivitet. FNë të vërtetë, janë shfaqur dëshmi mbi rolin e rrjetit interoceptiv dhe ariatik në pabarazitë sistemike që rezultojnë në konsumin kompulsiv të barnave ose ushqimit.
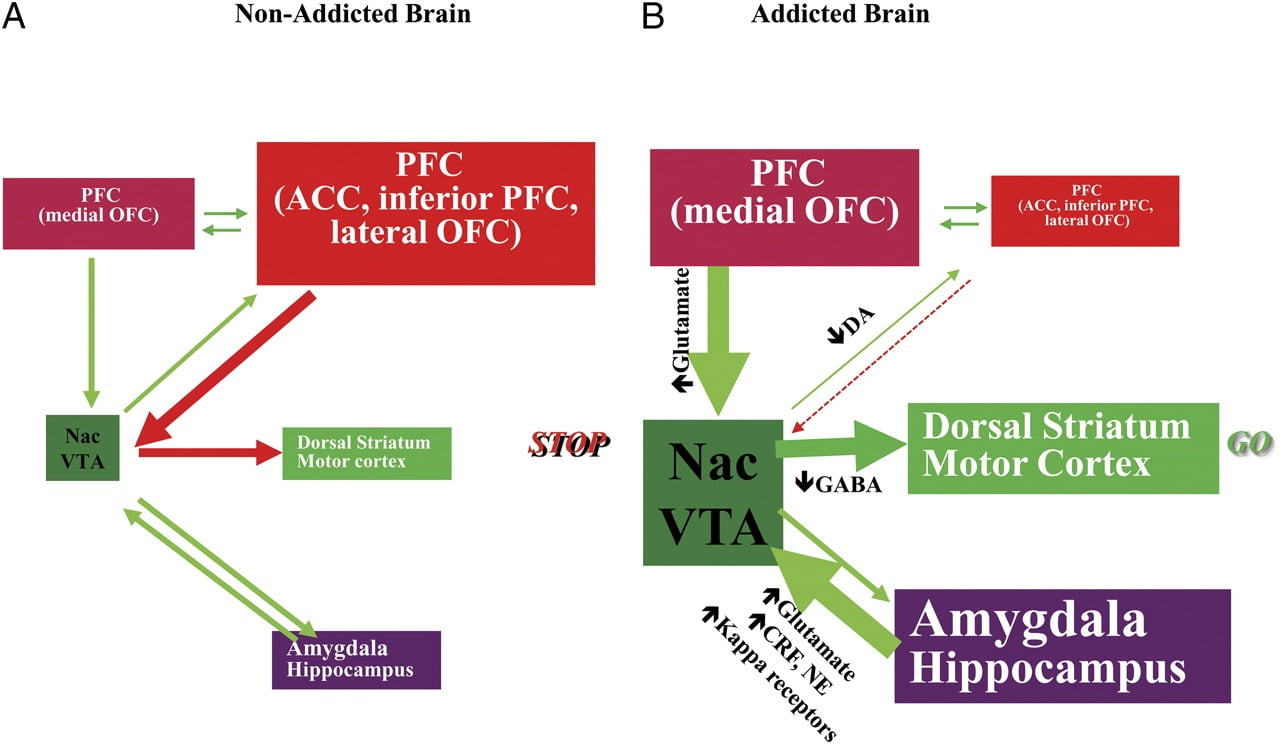
Fig 8 Modeli që propozon një rrjet të qarqeve ndërvepruese, ndërprerje që kontribuojnë në kompleksin e sjelljeve stereotipike që varen nga varësia e drogës dhe overeating kronike: (DLPFC, ACC, korteks inferior i frontalit, dhe lateral OFC), kontrolli ekzekutiv (DLPFC, ACC, cortex inferior i frontalit dhe lateral OFC), kondicionimi / kujtesa (amygdala, OFC mediale për atribuimin e saliency, hippocampus dhe striatum dorsal për zakonet) motivimi / përzgjedhja (OFC mediale për atribuimin e shpeshtësisë, ACC, VTA, SN, striatum dorsal, dhe korteks motorik). Nac, nucleus accumbens, interoception (Insula dhe ACC), dhe aversion / shmangien (Habenula). a Kur këto qarqe janë të balancuara, kjo rezulton në kontrollin e duhur frenues dhe vendimmarrjen. b Gjatë varësisë, kur vlera e shtuar e pritjes së drogës në shpërblimin, motivimin dhe qarqet e kujtesës kapërcen qarkun e kontrollit, duke favorizuar një lak me reagime pozitive të iniciuar nga konsumimi i drogës dhe përjetësuar nëpërmjet aktivizimit të zgjatur të motivimit / dhe qarqeve të memories. Këto qarqe gjithashtu ndërveprojnë me qarqet e përfshira në rregullimin e humorit, duke përfshirë reaktivitetin e stresit (që përfshin amigdalën, hipotalamusin, habenulën) dhe ndërveprimin (që përfshin insula dhe ACC dhe kontribuon në ndërgjegjësimin e dëshirës). Disa neurotransmitters janë të implikuar në këto neuroadaptations, duke përfshirë glutamate, GABA, norepinephrine, faktor lëshues corticotropin, dhe receptorët opioid. CRF, faktor lëshues i kortikotropinës; NE, norepinefrin. Modifikuar me leje nga Volkow et al. (2011b)
Si pasojë e përçarjes sekuenciale në këto qarqe, individët mund të përjetojnë 1) një vlerë të shtuar motivuese të drogës / ushqimit (të mesme për shoqëritë e mësuara përmes kondicionimit dhe zakoneve) në kurriz të armaturave të tjera (dytësore për ndjeshmërinë e zvogëluar të qarkut të shpërblimit ), Një aftësi të dëmtuar për të penguar veprimet e qëllimshme (të drejtuara nga qëllimi) të shkaktuara nga dëshira e fortë për të marrë drogën / ushqimin (dytësor për funksionin ekzekutiv të dëmtuar) që rezultojnë në marrjen e drogës / ushqimit dhe 2) dhe shmangie aversive që rezulton në marrjen impulsive të drogës për të shpëtuar nga gjendja ajetuese.
Ky model sugjeron një qasje terapeutike të shumëfishtë për varësinë e projektuar për të ulur vetitë përforcuese të drogës / ushqimit, rivendosjen / përmirësimin e pronave të dobishme të përforcimeve natyrore, pengimin e shoqatave të mësuara të kushtëzuara, rritjen e motivimit për aktivitetet jo të drogës / ushqimit, zvogëlimin e reaktivitetit të stresit , të përmirësojë gjendjen shpirtërore, dhe të forcojë kontrollin frenues me qëllim të përgjithshëm.
Mirënjohje
Autorët do të donin të falënderonin mbështetjen e programit intramural të Institutit Kombëtar të Shëndetit të NIAAA-s.
Referencat