Front. Behav. Neurosci., 23 Mars 2016 | http://dx.doi.org/10.3389/fnbeh.2016.00054

 Masroor R. Shariff1, Arnauld Belmer1, Mateu J. Fogarty2, Erica WH Mu2,
Masroor R. Shariff1, Arnauld Belmer1, Mateu J. Fogarty2, Erica WH Mu2, 

- 1Instituti i Kërkimeve të Përkthimit dhe Instituti për Shëndetin dhe Inovacionin Biomjekësor, Universiteti i Teknologjisë Queensland, Brisbane, QLD, Australi
- 2Shkolla e Shkencave Biomedical, Universiteti i Queensland, Brisbane, QLD, Australi
Dieta moderne është bërë shumë e ëmbël, duke rezultuar në nivele të pashembullta të konsumit të sheqerit, veçanërisht në mesin e adoleshentëve. Ndërkohë që futja kronike e sheqerit është e njohur për të kontribuar në zhvillimin e çrregullimeve metabolike duke përfshirë diabetin e obezitetit dhe të tipit II, pak dihet për pasojat e drejtpërdrejta të konsumit afatgjatë të sheqerit në tru. BSheqeri që mund të shkaktojë lirimin e dopaminës në bërthamë accumbens (NAc) në mënyrë të ngjashme me medikamentet e abuzimit, ne kemi hetuar ndryshimet në morfologjinë e neuroneve në këtë rajon të trurit pas javëve të shkurtër (4 javë) dhe afatgjata (12 javë) binge- si konsumi i saharozës duke përdorur një paradigmë të përsëritur me dy shishe. Ne përdorëm Golgi-Cox bojë për të ngopur neuronet mesatare të mprehtë (MSNs) nga bërthama e NAc dhe guaskë e minjve të konsumimit të saharozës afatshkurtër dhe afatgjatë dhe i krahasuam ato me kontrollet e ujit që përputheshin me moshën. Ne tregojmë se konsumimi i zgjatur i sojës me saharozë në mënyrë të konsiderueshme ka zvogëluar gjatësinë totale të dendritit të MSN shell shell në krahasim me minjtë e kontrollit që përputhen me moshën. Ne gjithashtu gjetëm se ristrukturimi i këtyre neuroneve rezultoi kryesisht nga kompleksiteti reduktuar dendriti distal. Anasjelltas, ne kemi vërejtur rritje të densitetit të shtyllës kurrizore në urdhërat e distaleve të DHN-së të NAc shell nga minjtë afatgjatë të konsumimit të saharozës. Kombinuar, këto rezultate nxjerrin në pah efektet neurone të marrjes së zgjatur të sikletit të saharozës në morfologjinë MSN të moralit të NAc.
Prezantimi
Gjatë viteve të fundit të 40, ka pasur një rritje të dokumentuar të konsumit të pijeve të sheqerit të ëmbël dhe ushqimeve që përmbajnë sheqerna të shtuara (Nielsen et al., 2002; Popkin, 2010; Ng et al., 2012), me raportet që vlerësojnë se deri në 75% të të gjitha ushqimeve dhe pijeve përmbajnë shuma të larta të sheqernave të shtuara (Ford dhe Dietz, 2013; Bray dhe Popkin, 2014). Gjatë kësaj periudhe, ka patur një rritje paralele të prevalencës së diabetit të obezitetit dhe tipit II, veçanërisht në mesin e adoleshentëve (Arslanian, 2002; Reinehr, 2013; Dabelea et al., 2014; Fryar et al., 2014). Studimet e fundit kanë treguar se fëmijët mbipeshë dhe të trashë shpesh konsumojnë sasi të larta të sheqerit të shtuar, megjithatë kontributi i dietave që përmbajnë sheqer të lartë në rritjen e incidencës së fëmijëve mbipeshë dhe të trashë mbetet e diskutueshme (Hu, 2013; Bray dhe Popkin, 2014; Bucher Della Torre dhe të tjerët, 2015).
Ndërsa një numër gjithnjë në rritje i provave tregon se konsumi i dietave të sheqerit të lartë mundet pjesërisht të kontribuojë në rritjen e peshës në mesin e fëmijëve dhe adoleshentëve (Malik et al., 2010; Te Morenga dhe të tjerët, 2013; Bray dhe Popkin, 2014), më pak vëmendje i është kushtuar pasojave negative jo-metabolike që vijnë nga marrja e tepërt e sheqerit. Interesante, disa modele të zakonshme të sjelljes dhe psikologjive shpesh shfaqen në mesin e një subjekte të atyre që hanë dhe mbajnë dieta të larta që përmbajnë sheqer. Më të dukshmet janë zhvillimi i çrregullimeve të të ngrënit duke përfshirë ushqimin me kërpudha, të kombinuara me fillimin e njëkohshëm të simptomave psikologjike duke përfshirë mungesën e motivimit dhe depresionit (shqyrtuar në Sheehan dhe Herman, 2015). Për më tepër, për shkak se individët që ushqehen me kërpudha shpesh shfaqin një humbje të kontrollit dhe një paaftësi për të kufizuar vetë marrjen e tyre të sheqerit, ka gjasa që këto sjellje të lindin si pasojë e adaptimeve neurologjike në rajonet e trurit që vlerësojnë vlerën hedonike të ushqimit shumë të shijshëm (Saper et al., 2002; Lutter dhe Nestler, 2009; Kenny, 2011). Kjo arsyetim mbështetet gjithashtu nga dëshmi në njerëz që tregojnë se sheqeri dhe ëmbëlsia mund të shkaktojnë dëshira të ngjashme me ato të shkaktuara nga droga të varur si alkooli dhe nikotina (Volkow et al., 2012).
Megjithëse pronat e varësisë të sheqerit janë akoma spekulative, këto vëzhgime kombinohen me studime që tregojnë kontributin e marrjes së tepërt të sheqerit ndaj ndryshimeve në qarkun e shpërblimeve dhe zhvillimin e sjelljeve të ngjashme me të varur dhe gjendjet emocionale në modelet e kafshëve (Avena et al., 2008; Benton, 2010; Ventura et al., 2014), garanton nevojën për hetime të mëtejshme. Studimet e mëparshme në brejtës kanë treguar se aksesi i përhershëm në sukrozë ndryshon veprimtarinë e disa neurotransmetuesve brenda sistemit mesolimbik duke përfshirë dopamin, opioidet dhe acetilkolinën (Shqyrtuar në Avena et al., 2008). Konsumi i sheqerit i saharozës ka treguar të lehtësojë lirimin e dopamines në bërthamë accumbens (NAc), në mënyrë të ngjashme me medikamentet e abuzimit (Avena et al., 2008). Për më tepër, ne kemi treguar se konsumimi afatgjatë i saharozës duke përdorur një paradhënie të zgjedhjes së dy shisheve të qasjes së përkohshme 24 h (Simms et al., 2008) modulon shprehjen nikotinike të receptorit të acetilkolinës (nAChR) në NAc (Shariff dhe të tjerët, në shtyp). Interesante, ne kemi vërejtur gjithashtu se komponimet nAChR të njohura për të moduluar aktivitetin e dopaminës dhe acetilkolinës në NAc, kanë efekte të ndryshme në konsumin e sukrozës pas konsumimit afatshkurtër dhe afatgjatë (Shariff dhe të tjerët, në shtyp).
Derisa këto studime kanë treguar ngjashmëri në ndryshimet e sjelljes dhe neurokemikale të shkaktuara nga qasja e përhershme në sheqer dhe drogë abuzimi, nuk dihet nëse këto efekte lehtësojnë ndryshimet në morfologjinë neurone në NAc. Te tij është në kontrast me substancat e abuzimit, duke përfshirë kokainën, amfetaminën dhe nikotinën, të cilat prodhojnë ndryshime të karakterizuara mirë në morfologjinë e neuroneve të mesme të mprehtë (MSNs) në NAc, duke përfshirë dendësinë e shtrirjes së shtyllës dhe kompleksitetin e ndryshuar të dendritit (Robinson dhe Kolb, 1999, 2004; Li et al., 2003; Crombag et al., 2005). Sepse ne kemi treguar më parë se ekspozimi afatgjatë (java 12) në alkool dhe sukroze duke përdorur paradigmën e zgjedhjes me dy shishe me ndërprerje prodhon një përgjigje diferenciale ndaj ndërhyrjeve farmakoterapeutike në krahasim me marrjen afatshkurtër (javët e 4; Steensland et al., 2007; Shariff dhe të tjerët, në shtyp), ne vlerësuam efektet e konsumit afatshkurtë dhe afatgjatë të saharozës në morfologjinë MSN në NAc. Kemi lejuar që minjtë e adoleshentëve të konsumojnë saharozë në mënyrë të ngjashme me 4 (afatshkurtra) ose 12 (afatgjatë) javë dhe pastaj të analizojnë morfologjinë e NAc MSNs nga minjtë që konsumojnë saharoze afatshkurtra dhe afatgjata dhe e krahasojnë këtë me kontrolleve që përputheshin me moshën, të cilëve u është dhënë aksesi vetëm në ujë. Rezultatet tona tregojnë se MSNs nga shell NAc janë ndryshuar pas konsumi afatgjatë, por jo afatshkurtë të saharozës, duke reduktuar gjatësinë dendritike, por shton densitetin distal të dendritit të shpinë. Për më tepër, ne kemi gjetur morfologjinë e MSNs nga thelbi NAc mbeti relativisht i paprekur pas konsumit të shkurtër dhe afatgjatë të saharozës. Këto rezultate nxjerrin në pah një pasojë të drejtpërdrejtë neurologjike të konsumit afatgjatë të saharozës në një mënyrë të ngjashme me kërcënimin. Për më tepër, këto të dhëna tregojnë nevojën për studime të mëtejshme që kanë për qëllim sqarimin e ndryshimeve molekulare dhe neurokimike që shoqërojnë ristrukturimin morfologjik të MSh shell-së NAc, të nxitur nga marrja e zgjatur e saharozës së ngjyrosur.
Materialet dhe Metodat
Deklarata e Etikës
Të gjitha procedurat eksperimentale janë kryer në përputhje me Kodin Australian për Kujdesin dhe Përdorimin e Kafshëve për Qëllime Shkencore, Edicioni 8th (Këshilli Kombëtar i Shëndetit dhe Mjekësisë, 2013). Protokollet u miratuan nga Komiteti i Etikës së Kafshëve në Universitetin Queensland dhe Komiteti i Etikës së Kafshëve në Queensland.
Kafshët dhe Strehimi
Rats meshkuj Wistar (Kontrolli: 176.4 ± 4.8 g; Sucrose: 178.3 ± 5.0 g) (ARC, WA, Australi), u vendosën në mënyrë individuale në Pleksiglas të dyfishtë të ventiluar® kafaze. Rats ishin aklimatizuar në kushtet individuale të strehimit, trajtimit dhe ciklit të kundërt të dritës 5 ditë para fillimit të eksperimenteve. Të gjitha minjtë u strehuan në një cikël të ndriçuar të dritës / errët 12-hr (dritat off në 9 am) në dhomën me ushqim dhe ujë të gatshëm ad libitum.
Paradigma e pijshëm me zgjedhje me dy shishe me ndërprerje
Qasja me ndërprerje 5% sukrozë paradigmë me dy shishe zgjedhje (Simms et al., 2008) u adaptua nga I urti (1973). Të gjitha lëngjet u paraqitën në shishe plastike të diplomuar të 300 ml me çelësat e pijshëm të çelikut inoks futur përmes dy grommets në para të kafazit pas fillimit të ciklit të dritës së errët. Peshat e secilës shishe u regjistruan përpara prezantimit të shisheve. Janë paraqitur dy shishe njëkohësisht: një shishe që përmban ujë; shishe e dytë që përmban 5% (w / v) saharoze. Vendosja e xNUMX% (w / v) shishe sucrose u alternuar me çdo ekspozim për të kontrolluar për preferencat anësore. Shishe u peshuan 5 h pas lëngjeve u paraqitën, dhe matjet u morën në 24 g më të afërt. Pesha e çdo miu është matur gjithashtu për të llogaritur gramin e konsumit të saharozës për kilogram të peshës trupore. Në ditën 0.1 të periudhës së pirjes, minjtë (n = 6-9) u është dhënë akses në një shishe të saharozës 5% (w / v) dhe një shishe uji. Pas 24 h, shishe e sucrose u zëvendësua me një shishe të dytë të ujit që ishte në dispozicion për 24 h ardhshëm. Ky model u përsërit të mërkurën dhe të premtën. Rats kishte qasje të pakufizuar në ujë në të gjitha ditët e tjera. Konsumi i ngjyrosur i saharozës rezultoi në një përshkallëzim në marrjen totale të saharozës (ml) me kalimin e kohës (Figura plotësuese 1) dhe u shoqërua me nivele të qëndrueshme bazike të pijshëm të bazuara në peshën e trupit [20 ± 5 g / kg të 5% (w / v)] gjatë periudhave afatshkurtra [~ 4 javë (seancat e pijshëm të 13)] dhe afatgjatë [ ~ 12 javë (seancat e pijshëm të 37)] periudha të pijshëm. Një grup i veçantë i minjve të kontrollit (n = 6-9) u është dhënë aksesi në ujë në të dy shishet (dmth., Nuk ka sukroze) nën të njëjtat kushte të përshkruara më sipër. Pesha mesatare e trupit të kontrollit dhe minjtë konsumues të saharozës në fund të ekspozimit afatshkurtër ishin 405.7 ± 40.8 g dhe 426.4 ± 31.2 g përkatësisht. Në fund të ekspozimit afatgjatë, pesha mesatare e trupit për grupet e kontrollit dhe saharozës ishte 578.8 ± 53.4 g dhe 600.2 ± 45.2 g.
Golgi-Cox Bojë
Pas sesionit të fundit të pirjes, minjtë u transferuan nga objekti i kafshëve për të lejuar përpunimin e mostrave të trurit në institucionin e histologjisë në Shkollën e Shkencave Biomedikale të Universitetit të Queensland (St Lucia, Australi). Të gjitha masat e miratuara janë ndërmarrë për të zvogëluar stresin gjatë transportit, pas së cilës, minjtë janë lejuar të rimarrin brenda natës. Ditën tjetër, minjtë u sakrifikuan nga overdoza pentobarbital natriumi (60-80 mg / kg, ip Vetcare, Brisbane, Australi) dhe perfshire intracardially me ~ 300 ml lëngu artificial cerebro-spinal që përmbante, (në mM): 130 NaCl, 3 KCl, 26 NaHCO3, 1.25 NaH2PO4, 5 MgCl2, 1 CaCl2, dhe 10 D-glukozë. Çdo kafshë u dekapitua më pas dhe truri hiqej dhe inkubohej në errësirë në Golgi-Cox që përmbante dikromate kaliumi 5%, kromat kaliumi 5% dhe klorid merkurik 5% (të gjitha kimikatet nga Sigma-Aldrich) që u bë ditë të freskëta 3 para sakrificës siç është përshkruar më parë (Rutledge et al., 1969). Inkubimi i njollave Golgi-Cox dhe metodat pas përpunimit janë modifikuar nga Ranjan dhe Mallick (2010). Trurit nga kafshët që konsumojnë saharozë afatshkurtër u inkubuan për ditët 6 në 37 ° C, ndërsa trurit nga kafshët që konsumonin saharozë afatgjatë u inkubuan për ditët 10, me një ndryshim në Golgi-Cox të freskët pas 4 ditëve të inkubacionit.
Pas inkubimit, seksionet koronale të 300 μm u prenë duke përdorur një mikrotome vibruese Zeiss Hyrax V50 (Carl Zeiss, Gjermani). Fletët pastaj u vendosën në mënyrë të vazhdueshme në pllakat 24-puro të mbushura me 30% (w / v) sukroze në 0.1 M fosfat të tretshëm me kripëra dhe të përpunuara siç përshkruhet në (Ranjan dhe Mallick, 2010). Shkurtimisht, seksionet u dehidratuan në 50% etanol për 5 min, pastaj vendosen në 0.1 M NH4OH për 30 min, shpëlarë dy herë me ujë të distiluar për 5 min dhe vendosen në Fujihunt film fixer (Fujifilm, Singapore) për 30 min në errësirë. Feta u pastruan pastaj dy herë në ujë të distiluar për 2 min secila dhe dehidratuar në 70, 90, 95 dhe 100% etanol dy herë për 5 min secila. Seksionet u pastruan pastaj në XXM (1: 1: 1 kloroform: ksilen: alkool) për 10 min dhe montuan në DPX (Sigma-Aldrich) në slides Superfrost Plus (Menzel-Glaser, Lomb Scientific, Australi) (Menzel-Glaser, Gjermani). Slides ishin lënë në errësirë për të thatë në temperaturën e dhomës brenda natës.
Përzgjedhja dhe gjurmimi neuronale brenda Nucleus Accumbens
Fetë koronale midis bregma + 2.8 dhe + 1.7 u anketuan për MSNs brenda bërthamës dhe shell e NAc, duke përdorur barkushe anësore dhe komisionin e mëparshëm si pikë referimi me ndihmën e atlasit të trurit të trurit (Paxinos dhe Watson, 2007) (Figura 1). Funksioni kontur në Neurolucida 7 (MBF Bioscience, VT, SHBA) është përdorur për të kufizuar thelbin e NAc dhe shell NAc në çdo fetë (Figura 2). Midis 2 dhe 9 neuronet për rajonin për kafshë u gjurmuan për parametrat e gjatësisë dendritike duke përdorur një objektiv 63x ose për densitetin e shtyllës kurrizore (raportuar si spines per 100 μm) duke përdorur një objektiv 100x në Zeiss Axioskop II (Carl Zeiss, Gjermani) xyz fazë të drejtuar nga Neurolucida® Softueri 7 (MBF Biosciences, VT, SHBA). Të gjitha gjetjet u kryen në mënyrë të verbuar në lidhje me trajtimin. Parametrat morfologjike të neuroneve të ngopura Golgi-Cox u analizuan në një mënyrë të ngjashme me raportet e mëparshme (Klenowski et al., 2015).
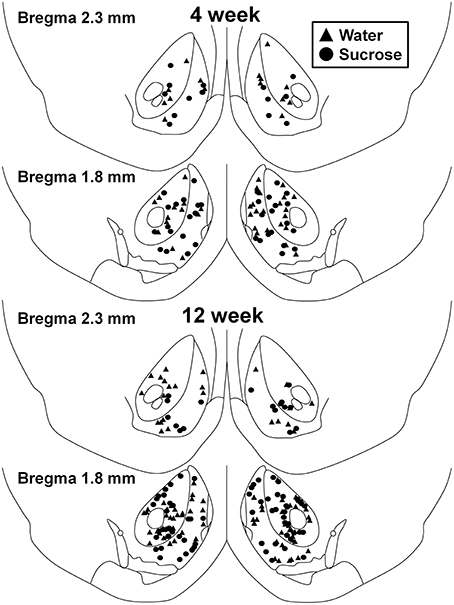
Figura 1. Hartë që tregon vendet e neuroneve të mesme me gjemba të mostruara nga bërthama e bërthamës accumbens dhe guaskë e 4 dhe 12 javë të saharozës që konsumojnë minjtë dhe kontrollet që përputhen me moshën. Dy panelet kryesore tregojnë lokacionet e neuroneve të marrura nga bërthama e nucleus accumbens dhe shell e kontrollit të javës 4 (trekëndëshat) dhe kafshëve të saharozës (qarqeve). Në fund të dy panelet tregojnë pozicionet e neuroneve të kampionuara nga kontrolli i javës 12 (trekëndëshat) dhe kafshët e saharozës (qarqeve).
Analiza statistikore
Mesatarja dhe gabimi standard i mesatares (SEM) janë llogaritur për çdo grup të të dhënave me kafshën si n, duke përdorur të dhënat morfometrike mesatare nga të gjitha NAc MSN kryesore ose shell (n = 7 për shell NAc dhe n = 6 për bërthamën e NP-së 4-javë, n = 9 për grupet 12-javore). Aty ku tregohet, dy studentë të palidhur me bisht tTestet ose ANOVA me dy mënyra me Bonferroni pas testeve u kryen për të gjitha analizat që përfshijnë krahasimin e mjeteve të grupit, duke përdorur versionin GraphPad Prism 6.02 (GraphPad Software, San Diego, CA). Rëndësia statistikore u pranua në P <0.05. Të gjitha të dhënat në seksionin e rezultateve paraqiten si mjete EM SEM. Ndryshimet në përqindje llogariten në lidhje me vlerën e kontrollit.
Rezultatet
Neuronet e mesme me gjemba nga Shell i Nucleus Accumbens kanë zvogëluar gjatësinë e dendritit, zvogëlon kompleksitetin e dendritit, por shton dendësinë mesatare të shtyllës kurrizore në porosi distal të degëve pas konsumit të gjatë, por jo afatshkurtë të saharozës
Pas konsumit afatshkurtër të konsumit të saharozës (4 javë), nuk kishte dallime të konsiderueshme në parametrat morfometrikë MSN të Shellit NAc (Tabela 1). Nuk kishte gjithashtu dallime të konsiderueshme në mes të konsumit afatshkurtë të saharozës dhe kontrollit të ujit NAc shell MSNs në analizat që lidhen me rendin e degës centrifugale. Gjegjësisht, segmentet dendriti sipas rendit të degës (P = 0.4111), do të thotë gjatësia dendritike sipas rendit të degës (P = 0.5581) dhe dendësia mesatare e shpinës sipas rendit të degëve (P = 0.2977, ANOVA me dy drejtime) nuk ishin dukshëm të ndryshme midis grupeve. Një hartë vendore që tregon pozicionet e përafërta të neuroneve të mostruara është treguar në Figura 1.
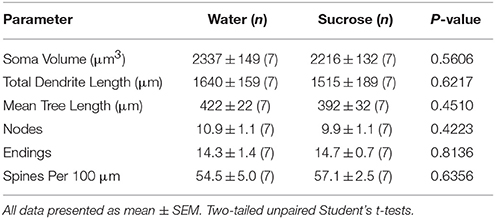
Tabela 1. Parametrat e përgjithshëm morfologjik të neuroneve të mesme të mprehtë nga gërmadha e nucleus accumbens të minjve të konsumimit të saharozës afatshkurtër dhe kontrollet e ujit që përputhen me moshën.
Pas një periudhe afatgjatë të konsumit të saharozës (xNUMX javë), gjatësia totale e dendritit të NSc shell NSc u zvogëlua me 12% krahasuar me kontrollet konsumuese të ujit (Uji: 21 ± 1827 μm, n = 9; Sucrosi 1449 ± 78 μm, n = 9, *P = 0.0384, Student i pa çiftuar me dy bishta t-test, Figura 2, Tabela 2). Krahasimi i numrit mesatar të bifurcacioneve dendrite (nyjeve) dhe mbarimit të dendritit midis grupeve të ujit dhe saharozës zbuloi një nivel të reduktuar (edhe pse jo të rëndësishëm) të kompleksitetit dendritik në MSN shell shell (nyjet: Water 12.9 ± 1.4 n = 9, Sucrose 10.1 ± 0.8 n = 9, P = 0.0879; përfundimet: Uji 17.9 ± 1.4 n = 9, Sucrose 14.8 ± 0.7 n = 9, P = 0.0657, Student i pa çiftuar me dy bishta t-test, Tabela 2). Nuk kishte asnjë ndryshim në vëllimin soma (P = 0.9400), do të thotë gjatësia e pemës dendritike (P = 0.1646) ose dendësia totale e shpinë (P = 0.3662) në NAc shell MSNs nga afatgjatë të saharozës konsumojnë minjtë në krahasim me kontrollet e ujit. Këto parametra morfometrikë janë detajuar në Tabelën 2.
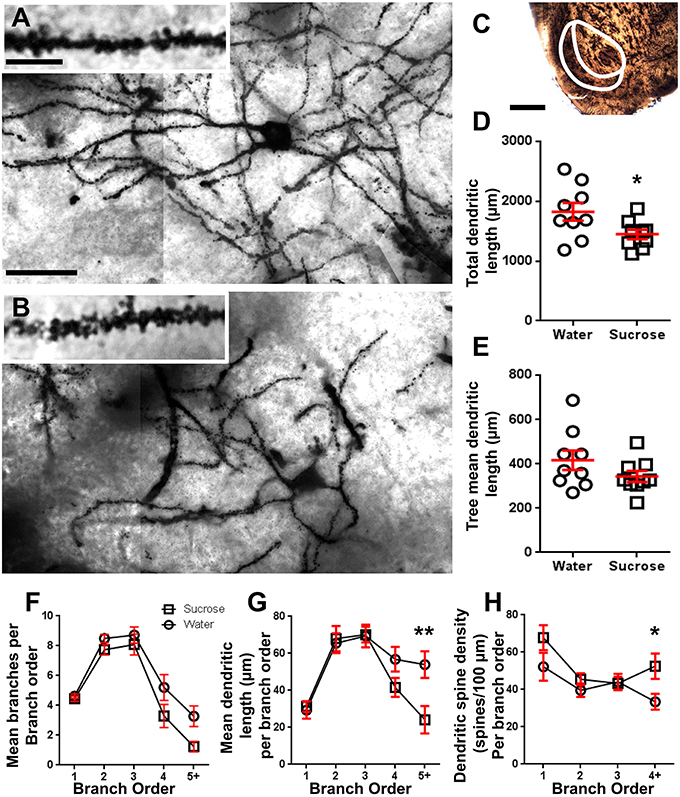 Figura 2. Rritja e dendritit dhe rritja e densitetit të dendritit distal të dendësisë së mesit të mesit (NSs) nga bërthama accumbens (NAc) e minjtë afatgjatë të trajtimit të saharozës në krahasim me minjtë e kontrollit. (A, B) tregojnë përfaqësimet e kontrollit (lartë) dhe afatgjatë (12 javë) saharoze (fund) trajtuar brightfield z-stacionet e mozaikut të MSN-ve të ngopura nga Golgi nga shell NAc (zmadhimi 63x). Shtratin e (A, B) tregon kontrollin dhe imazhet afatgjata të trajtimit të saharozës të ndritshme të dendriteve MSN mbingarkuar nga Golgi dhe shtyllat dendritike nga shell NAc (zmadhimi 100x). (C) tregon rajonet anatomike që MSN ishin përzgjedhur nga ky studim. (D) tregon një grumbull të shpërndarë të drithit total dendritik MSN (mesatarja ± SEM) nga predha NAc në kafshët afatgjata të saharozës (sheshe) në krahasim me kontrollet (qarqet), nxënësit e palëvizur t-test, *P <0.05, n = 9; kontroll dhe n = 9; 12 javë saharoze. (E) tregon një gjerësi të pandryshuar të pemës mesatare të pemës dendriti MSN (mesatarja ± SEM) nga predha NAc në kafshët afatgjata të saharozës (sheshe) në krahasim me kontrollet (qarqet), nxënësit e palëvizur t-Test, P > 0.05, n = 9; kontroll dhe n = 9; 12 javë saharoze. Analiza e rendit të degës (mean ± SEM) e numrit të segmentit dendritik sipas rendit të degës (F), do të thotë gjatësia dendriti sipas rendit të degës (G) dhe dendësia e dendritit të shtyllës kurrizore sipas rendit të degës (H). Konsumi afatgjatë i saharozës ka zvogëluar gjatësinë dendrite në urdhërat e distaleve të degës (5 +) dhe shtimin e densitetit të shtyllës dendriti në urdhërat e distaleve të degëve (4 +) krahasuar me kontrollet (G, H), ANOVA me dy drejtime me Bonferroni pas testeve, *P <0.05, **P <0.01, n = 9; kontroll dhe n = 9; saharoze afatgjate. Bare shkallë: (A, B) = 20 μm; brenda (A, B) = 10 μm; (C) = 1 mm.
Figura 2. Rritja e dendritit dhe rritja e densitetit të dendritit distal të dendësisë së mesit të mesit (NSs) nga bërthama accumbens (NAc) e minjtë afatgjatë të trajtimit të saharozës në krahasim me minjtë e kontrollit. (A, B) tregojnë përfaqësimet e kontrollit (lartë) dhe afatgjatë (12 javë) saharoze (fund) trajtuar brightfield z-stacionet e mozaikut të MSN-ve të ngopura nga Golgi nga shell NAc (zmadhimi 63x). Shtratin e (A, B) tregon kontrollin dhe imazhet afatgjata të trajtimit të saharozës të ndritshme të dendriteve MSN mbingarkuar nga Golgi dhe shtyllat dendritike nga shell NAc (zmadhimi 100x). (C) tregon rajonet anatomike që MSN ishin përzgjedhur nga ky studim. (D) tregon një grumbull të shpërndarë të drithit total dendritik MSN (mesatarja ± SEM) nga predha NAc në kafshët afatgjata të saharozës (sheshe) në krahasim me kontrollet (qarqet), nxënësit e palëvizur t-test, *P <0.05, n = 9; kontroll dhe n = 9; 12 javë saharoze. (E) tregon një gjerësi të pandryshuar të pemës mesatare të pemës dendriti MSN (mesatarja ± SEM) nga predha NAc në kafshët afatgjata të saharozës (sheshe) në krahasim me kontrollet (qarqet), nxënësit e palëvizur t-Test, P > 0.05, n = 9; kontroll dhe n = 9; 12 javë saharoze. Analiza e rendit të degës (mean ± SEM) e numrit të segmentit dendritik sipas rendit të degës (F), do të thotë gjatësia dendriti sipas rendit të degës (G) dhe dendësia e dendritit të shtyllës kurrizore sipas rendit të degës (H). Konsumi afatgjatë i saharozës ka zvogëluar gjatësinë dendrite në urdhërat e distaleve të degës (5 +) dhe shtimin e densitetit të shtyllës dendriti në urdhërat e distaleve të degëve (4 +) krahasuar me kontrollet (G, H), ANOVA me dy drejtime me Bonferroni pas testeve, *P <0.05, **P <0.01, n = 9; kontroll dhe n = 9; saharoze afatgjate. Bare shkallë: (A, B) = 20 μm; brenda (A, B) = 10 μm; (C) = 1 mm.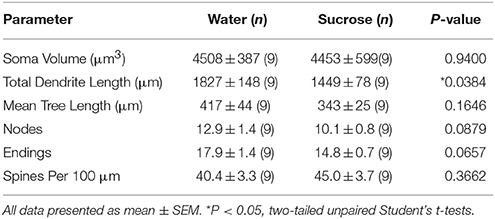
Tabela 2. Parametrat e përgjithshëm morfologjik të neuroneve të mesme të mprehtë nga gërmadha e bërthamës accumbens të minjtë që konsumojnë saharozë afatgjatë dhe kontrollet e ujit që përputhen me moshën.
Pas karakterizimit të morfologjisë së përgjithshme të dendritit të saharozës afatgjatë që konsumojnë MSN shell shell NAc, ne analizuam arborizations dendritic dhe dendësia shpinë në lidhje me karakteristikat e rendit të degës së tyre. Vlerësimi ynë gjithëpërfshirës i pemëve dendriti quantifikon numrin e segmenteve dendriti sipas rendit të degës, gjatësia mesatare e segmenteve dendriti sipas rendit të degës dhe dendësia mesatare e shpinës sipas rendit të degëve të shell-ve të NAc-së të kontrollit të ujit dhe të minjve që konsumojnë saharozë afatgjatë. Një përmbledhje e të dhënave dhe analizës së rendit të degës është paraqitur në Tabelë 3.
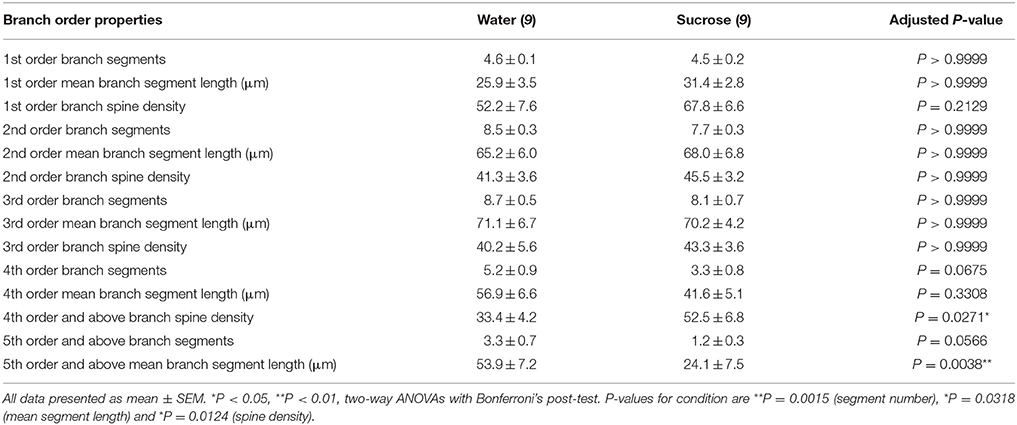 Tabela 3. Karakteristikat e rendit të degëve të neuroneve të mesme të shtyllave nga saharoza afatgjatë dhe minjtë që pijnë ujë.
Tabela 3. Karakteristikat e rendit të degëve të neuroneve të mesme të shtyllave nga saharoza afatgjatë dhe minjtë që pijnë ujë.Numri mesatar i segmentit të degës së degës sipas rendit të degëve të MSN shell shell NAc u reduktua ndjeshëm në minjtë që konsumojnë saharozë afatgjatë në krahasim me kontrollet e ujit (**P = 0.0015, dyanshëm ANOVA). Testet pas testeve të Bonferroni zbuluan një trend drejt një numri të reduktuar të segmenteve të degëve në 4th (Uji: 5.2 ± 0.9, n = 9; Sucrose 3.3 ± 0.8, n = 9, P = 0.0675, Figura 2F, Tabela 3), dhe rendi 5th dhe urdhrat e degëve më lart (Uji: 3.3 ± 0.7, n = 9; Sucrose 1.2 ± 0.3, n = 9, P = 0.0566, Figura 2F, Tabela 3). Gjatësia e segmentit dendritik sipas rendit të degëve të MSN shell shell NAc u reduktua ndjeshëm në minjtë që konsumojnë saharozë afatgjatë në krahasim me kontrollet e ujit (*P = 0.0444, dyanshëm ANOVA). Bonferroni pas testeve tregoi një reduktim të 55% në degët e rendit 5th dhe më gjerë (Uji: 53.9 ± 7.2 μm, n = 9; Sucrosi 24.1 ± 7.5 μm, n = 9, **P = 0.0038, Figura 2G, Tabela 3).
Analiza e rendit të degës tregoi një rritje të konsiderueshme në densitetin e shtyllës së dendritit të shell-ve të NAc-së të minjve afatgjatë të konsumimit të saharozës krahasuar me kontrollet (*P = 0.0124, dyanshëm ANOVA). Testet post-Bonferroni treguan një rritje të densitetit të shtyllës kurrizore të 57% në degët me distancë 4th dhe përtej (Uji: 33.4 ± 4.2, n = 9; Sucrose 52.5 ± 6.8, n = 9, P = 0.0271 *, pjesë e figurave 2A, B, H, Tabela 3). Imazhet përfaqësuese të arkitekturës së përgjithshme të MSN-it dhe të densitetit të shpinës distale (inset) janë përshkruar në Figura 2A, B.
Të marra së bashku, këto rezultate tregojnë se konsumi afatshkurtë i saharozës ka pak efekt në parametrat morfologjike të MSN-ve brenda shell NAc. Megjithatë, pas konsumit të zgjatur, ka një rënie të ndjeshme në gjatësinë dhe kompleksitetin e gjurit neuronal, veçanërisht në degët dendrite distale. Rritja dendëshe e shpeshtë e shpeshtë e njëpasnjëshme është gjithashtu e dukshme në MSN shell shell të afatgjatë të minjtë që konsumojnë saharozë.
Neuronet e Mesme të Mesme nga Core Nucleus Accumbens kanë zvogëluar kompleksitetin e degëzuar pas Konsumit të gjatë, por jo afatshkurtër të saharozës
Pas konsumit të shkurtër të konsumit të saharozës, nuk kishte dallime të rëndësishme në parametrat morfometrikë MSN të Bazës Kombëtare të NAc (Tabela 4). Gjithashtu nuk kishte dallime të rëndësishme midis konsumit të saharozës së javës së 4 dhe NSs kryesore të kontrollit të ujit në analizat që lidhen me rendin e degës së centrifugës. Gjegjësisht, segmentet dendriti sipas rendit të degës (P = 0.7717), do të thotë gjatësia dendritike sipas rendit të degës (P = 0.2096), dhe dendësia mesatare e shpinës sipas rendit të degës (P = 0.3521, ANOVA me dy drejtime) nuk ishin të ndryshme në mes të grupeve.
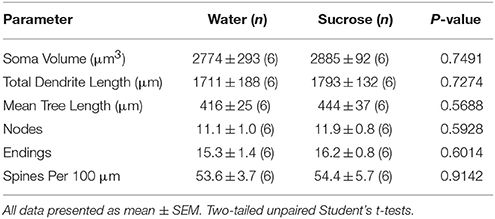 Tabela 4. Parametrat e përgjithshëm morfologjik të neuroneve të mesme me gjemba nga bërthama e bërthamës accumbens të minjve që konsumojnë saharozë afatshkurtër dhe kontrollet e ujit.
Tabela 4. Parametrat e përgjithshëm morfologjik të neuroneve të mesme me gjemba nga bërthama e bërthamës accumbens të minjve që konsumojnë saharozë afatshkurtër dhe kontrollet e ujit.Konsumi i zgjatur i saharozës gjithashtu nuk kishte asnjë rëndësi për parametrat morfometrikë MSN të Bazës Kombëtare të NAc (Tabela 5). Numri mesatar i segmentit të degës së degës sipas rendit të degës së MSN-ve bazë të NAc ishte reduktuar ndjeshëm në minjtë që konsumojnë saharozë afatgjatë në krahasim me kontrollet e ujit (*P = 0.0416, ANOVA me dy drejtime), megjithatë nuk ka pasur dallime domethënëse në gjatësinë mesatare dendrite sipas rendit të degës (P = 0.0995) dhe dendësia mesatare e shpinës sipas rendit të degëve (P = 0.4888, ANOVA me dy drejtime) midis MSNs në bërthamën e NAc të minjtë që konsumojnë saharozë afatgjatë në krahasim me kontrollet e ujit. Të marra së bashku, të dhënat tona tregojnë se bërthama e NAc nuk është aq e përgjegjshme për konsumin afatgjatë të saharozës në krahasim me MSN-të nga rajoni shell NAc.
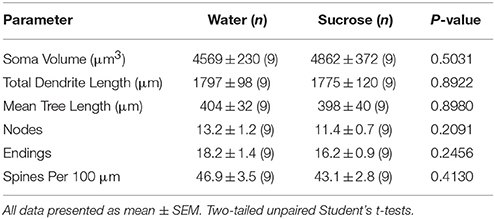 Tabela 5. Parametrat e përgjithshëm morfologjik të neuroneve të mesme me gjemba nga bërthama e bërthamës accumbens të minjve që konsumojnë saharozë afatgjatë dhe kontrollet e ujit që përputhen me moshën.
Tabela 5. Parametrat e përgjithshëm morfologjik të neuroneve të mesme me gjemba nga bërthama e bërthamës accumbens të minjve që konsumojnë saharozë afatgjatë dhe kontrollet e ujit që përputhen me moshën.Diskutim
Shtimi i disponueshmërisë së ushqimeve shumë të ëmbla në dietë perëndimore jo vetëm që ka kontribuar në rritjen e prevalencës dhe barrës ekonomike të obezitetit dhe diabetit të tipit II, por gjithashtu ka çuar në fillimin e çrregullimeve të ngrënies siç janë ngrëniet (Swanson et al., 2011; Kessler et al., 2013; Davis, 2015). Megjithëse pronat e varësisë së sheqernave, përfshirë fruktozën dhe saharozën mbeten spekulative, ekziston një ngjashmëri e dukshme në korrelatet e sjelljes dhe ato nervore që shfaqen si pasojë e konsumimit të tepërt të drogës dhe zgjatjes së përdorimit të drogës (Avena et al., 2008, 2011). Përveç kësaj, sheqeri aktivizon qarkun e shpërblimit të trurit në një mënyrë që është e ngjashme me ilaçet e abuzimit (Volkow et al., 2012) dhe rezultatet nga studimet njerëzore sugjerojnë se sheqeri dhe ëmbëlsia mund të nxisin dëshirat që janë të krahasueshme në madhësi me ato të nxitura nga barna të varur si alkooli dhe nikotina (Volkow et al., 2012). Ne për këtë arsye kemi përdorur një model të konsumimit të kërpudhave të sakuzës në rats për të përcaktuar efektet e konsumit të saharozës së shkurtër (4) dhe afatgjatë (12 javë) të morfologjisë neuronale të MSNs në NAc, një komponent kyç i rrjetit shpërblimor të mbivendosur që është moduluar nga sheqeri dhe droga Addictive. Ne tregojmë se MSNs nga shell NAc e kripës afatgjatë të saharozës që konsumojnë minjtë kanë ulur ndjeshëm gjatësinë dhe kompleksitetin dendritik, por shton distancën dendrite dendrite të shpinës. Konsumi afatgjatë i saharozës nuk kishte efekt mbi morfologjinë e MSNs nga bërthama e NAc, ndërsa konsumi afatshkurtë i saharozës gjithashtu nuk kishte efekt të rëndësishëm në morfologjinë e MSN nga bërthama ose shell e NAc. Këto rezultate jo vetëm që tregojnë një efekt të drejtpërdrejtë të marrjes së zgjatur të saharozës në morfologjinë neuronale të morfologjisë neuronale të MSc shell shell, por ato gjithashtu nxjerrin në pah pasojat e mundshme të dëmshme të konsumit të zgjatur të dieta me sheqer të lartë.
NAc, që është pjesë e striatumit të barkut, përbëhet kryesisht nga MSNs, të cilat karakterizohen në mënyrë morfologjike si neuronet e mesme me arborizime të gjera dendriti dhe densitet të lartë të shtyllës kurrizore (Kemp dhe Powell, 1971; Graveland dhe DiFiglia, 1985; Rafols et al., 1989; Kawaguchi et al., 1990). Neuronet glutamatergic dhe dopaminergic janë dy inputet primare afferent për NAc, kryesisht kontaktuar boshtet dendritic dhe spines e MSNs (Groves, 1980; Kaiya dhe Namba, 1981; Groves et al., 1994). Konkretisht, shell shell dhe core marrjen input glutamatergic nga zona funksionalisht të dallueshme cortical (Brog et al., 1993). Shell NAc është gjithashtu innervated nga afferents excitatory nga rajonet subcortical si hippocampus, thalamus dhe amygdala basolateral (Brog et al., 1993; Wright dhe Groenewegen, 1995). Studimet e mëparshme kanë treguar se këto kontribute glutamatergike luajnë një rol të rëndësishëm në motivimin dhe sjelljet e drejtuara nga qëllimi si ushqimi dhe kërkimi i shpërblimeve (Maldonado-Irizarry et al., 1995; Kelley dhe Swanson, 1997; Reynolds dhe Berridge, 2003; Richard dhe Berridge, 2011). Hyrja tjetër dominuese në NAc MSNs është nga afferent dopaminergjik që projektohet nga zona tegmentale e barkut (Lindvall dhe Björklund, 1978; Veening et al., 1980; Kalivas dhe Miller, 1984). Është interesante se studimet e mëparshme që përdorin modele të ngjashme të qasjes me ndërprerje të sheqerit kanë treguar se konsumimi i ngjashëm me kërcënimin rezulton në një rritje të dopaminës ekstracelulare në NAc ngjashëm (megjithëse në një masë më të vogël) ndaj drogave të abuzimit (Rada et al., 2005; Avena et al., 2006), dhe mund të moduloj shprehjen e receptorit dopamin (Colantuoni et al., 2001, 2002) në thelbin e NAc dhe shell. Është interesant fakti se konsumimi i saharozës, si për shembull, shkakton një përshkallëzim në marrje me kalimin e kohës në mënyrë të ngjashme me vetë-administrimin e drogave të abuzimeve siç janë kokaina dhe heroin (Ahmed dhe Koob, 1998; Ahmed et al., 2000, 2003) e cila është e lidhur me zhvillimin e një shteti "addictive like".
Analiza jonë e morfometrisë së rendit të degës tregon që reduktimi i përgjithshëm i gjatësisë dendrite të MSN shell shell-ve të shkaktuara nga marrja afatgjatë e saharozës, rezulton kryesisht nga reduktimet në kompleksitetin e urdhrave të distaleve të degëve. Kemi vërejtur degëzime të degëzuara të ulëta (rendi 4th dhe 5th dhe urdhrat e degëve të mësipërme) dhe gjatësia mesatare e reduktuar në mënyrë të konsiderueshme në rendin 5th dhe mbi dendritët, kombinuar me shtrirjen e shtrirjes së shpinës në këto radhë dege. Një faktor i përbashkët që ka gjasa të ndikojë në këtë lloj ristrukturimi dendriti përfshin ndryshimet në lidhjen sinaptike dhe / ose funksionin (Russo et al., 2010). Studimet e mëparshme kanë treguar se synapses glutamatergic në MSNs janë formuar kryesisht në spines, veçanërisht në dendritet distale (Groenewegen et al., 1999). Përveç kësaj, bashkë-lokalizimi i dopaminës dhe inputeve glutamatergike nga lëvore paraballore (Sesack dhe Pickel, 1992), hippocampus (Totterdell dhe Smith, 1989; Sesack dhe Pickel, 1990), dhe amygdala (Johnson et al., 1994) janë vërejtur në shtyllat dendrite të MSNs. Këto vëzhgime të kombinuara me rritjen e densitetit të shtyllës kurrizore pas konsumit afatgjatë të saharozës që shihet në studimin tonë, mbështesin formimin e rritjes së inputeve ngacmuese. Prandaj, shfaqet mundësia kur efektet e vazhdueshme të shkaktuara nga marrja e zgjatur e saharozës me kërcitje mund të lehtësojnë rritjen e aktivitetit sinaptik eksitues në dendritet distale të MSNs në shell NAc. Rrjedhimisht, reduktimi dhe / ose tërheqja e dendriteve distale mund të rezultojë nëpërmjet një mekanizmi synetik homeostatik (Reissner dhe Kalivas, 2010), megjithatë kjo mbetet për t'u përcaktuar.
Është interesante të theksohet se Crombag dhe kolegët treguan se nuk kishte rritje të dendësisë së shtyllës kurrizore në shellin e NAc pas konsumit të saharozës së javës 4 nëpërmjet paradigmës së vetadministrimit të hundës pavarësisht nga një blerje më të fuqishme dhe një shkallë më të lartë përgjigjeje ndaj saharozës kur krahasohej me amfetamin (Crombag et al., 2005). Vëzhgimi i tyre për një mungesë ndryshimi në dendësinë e shtyllës kurrizore në javët e 4 pasqyron gjetjet tona. Në të kundërt, megjithatë, studimi ynë tregon se pas një ekspozimi afatgjatë (12 javë) ndaj konsumit kronik të saharozës, ekziston një rritje e ndjeshme e densitetit të shpinës distale në MSN të përvojave të përhapjes së saharozës. Për më tepër, laboratori ynë ka treguar më parë se konsumi afatgjatë i saharozës (12 javë) lehtëson një përgjigje diferenciale farmakologjike ndaj farmakoterapikëve që janë treguar të modulojnë përgjigjet e dopaminës dhe acetilkolinës në nivelin e NAc (Shariff dhe të tjerët, në shtyp). Të marra së bashku, kjo sugjeron që ekspozimi afatgjatë (12 javë dhe më tej) të saharozës, që është më saktësisht reflektues i skenarëve të botës reale, rezulton në përshtatje morfologjike në nivelin e NAc.
Përsa i përket drogës së abuzimit, ekspozimi i përsëritur ndaj barnave të ndryshme prodhon ndryshime afatgjata në strukturën e dendriteve dhe shtyllat dendrite. Për shembull, amfetamina dhe kokaina rrisin dendësinë e shtyllës së shpinës në NAc si në shell ashtu edhe në bazë (Robinson dhe Kolb, 2004). Ekspozimi ndaj nikotinës gjithashtu është treguar të rrisë densitetin e shtyllës kurrizore në shell NAc. Anasjelltas, ekspozimi i morfines çon në një rënie në densitetin e shtyllës kurrizore dhe në kompleksitetin e degës së dendritit (Robinson dhe Kolb, 2004). Në kushtet e konsumit afatgjatë të saharozës, vërehet një rritje e dendësisë së shpinës, e ngjashme me amfetaminën, kokainën dhe nikotinën dhe e kundërta me efektin e morfinës. Sidoqoftë, ndryshe nga amfetamina dhe kokaina, por e ngjashme me nikotinën, rritja e dendësisë së shtyllës kurrizore në ekspozimin afatgjatë të saharozës është e kufizuar në shell NAc. Është gjithashtu interesante që ndryshimet në të dy degëzimet dendrite (Robinson dhe Kolb, 1999) dhe dendësia e shtyllës kurrizore (Li et al., 2003) të prodhuara nga amfetamina ose kokaina janë të kufizuara në dendrite distale të MSNs në NAc, e cila pasqyron gjetjet në studimin tonë. Për më tepër, dhe korroborative për ndryshimet e përshkruara më lart, konsumi i saharozës gjithashtu është treguar më parë për të rritur forcën excitatory synaptic në dopamir neuronet accumbal (Stuber et al., 2008b) si dhe komponentët e tjerë të rrugës së shpërblimit mesolimbik (Stuber et al., 2008a; Chen et al., 2010). Të marra së bashku, kjo paraqet saharozë si një modulator i fuqishëm i morfologjisë së neuroneve pas përdorimit të rëndë të zgjatur, i cili është i ngjashëm me efektet e vërejtura nga drogat e abuzimit.
Megjithëse kërkohen hetime të mëtejshme për të zbuluar mekanizmat celulare dhe sinaptike që kontribuojnë në ndryshimet morfologjike të vërejtura në këtë studim, rezultatet tona tregojnë efekte të rëndësishme neuronale të shkaktuara nga konsumi afatgjatë i saharozës. Në veçanti, një konsideratë që nuk shqyrtohet në studimin tonë është nëse efektet e vëzhguara morfologjike të saharozës gjithashtu mund të nxirren me ëmbëlsuesit jo kalorikë siç është sakarina. Në lidhje me këtë, është e rëndësishme të theksohet se Lenoir dhe kolegët kanë treguar se ëmbëlsia intensive e tejkalon shpërblimin e kokainës, qoftë ajo e prodhuar nga saharin ose saharoza (Lenoir et al., 2007). Për më tepër, një studim i kohëve të fundit i botuar nga laboratori ynë (Shariff dhe të tjerët, në shtyp) tregon se vareniklin, një agonist i pjesshëm i receptorit të acetilkolinës nikotinës redukton sasinë e saharozës dhe saharinë në brejtës duke ndjekur të njëjtin regjim të aksesit të përhershëm afatgjatë të përdorur në këtë studim. Interesante, studimet e mëparshme kanë treguar ngjashmëri në efektet akute të ëmbëlsuesve jo kalorikë siç janë sakarina dhe saharoza në nivelin e NAc (Scheggi et al., 2013; Tukey et al., 2013; Carelli dhe Perëndimi, 2014). Megjithatë, studime të mëtejshme duhet të përcaktojnë nëse jo-kalorike ëmbëlsues mund të nxisin efekte afatgjata të ngjashme me ndryshimet në morfologjinë e MSN shell shell të shkaktuara nga konsumi afatgjatë të saharozës raportuar këtu.
Mungesa e efektit në morfologjinë e NAc MSN pas konsumit afatshkurtë të saharozës, thekson rëndësinë e zbatimit të studimeve afatgjata për të vlerësuar ndikimin e abuzimit të zgjatur të drogës ose shpërblimeve natyrore si saharoza. Përsa i përket varësisë, jo vetëm që janë ciklet e përsëritura të marrjes së hidhur dhe komponentëve kryesorë të abstinencës të ciklit të varësisë, një numër i madh i provave ka zbuluar se kalimi në varësi është një proces progresiv që shpesh ndodh gjatë një periudhe të zgjatur kohore. Megjithëse pronat e varësisë të sheqernave mbeten të pasigurta, besueshmëria e varësisë ndaj shpërblimeve të tjera jo të drogës, si gjinia, bixhozi dhe ushqimi po hetohen gjithnjë e më shumë. Rezultatet nga ky studim shtojnë meritën ndaj hipotezës se sheqernat siç është sukroza potencialisht kanë veti të varur pas konsumit afatgjatë dhe të përtypur. Rezultatet tona kanë gjithashtu implikime për numrin në rritje të fëmijëve dhe adoleshentëve që mbajnë shprehitë e hahetshme të ushqimit (konsumi i lartë i sheqerit dhe ngrënia e sheqerit) në moshë madhore. Në përputhje me rritjen e rrezikut të zhvillimit të efekteve metabolike është gjithashtu e mundur që pasojat neurologjike dhe psikiatrike që ndikojnë në gjendjen shpirtërore dhe motivimin mund të rezultojnë nga këto sjellje.
Kontributet e autorëve
Ka marrë pjesë në projektimin e hulumtimit: PK, SB. Eksperimentet e kryera: PK, MS, AB, MF, EM. Analiza e të dhënave: PK, MF, MS. Interpretuar të dhënat dhe kontribuar në shkrimin e dorëshkrimit: PK, MS, MF, EM, MB, SB. Të gjithë autorët lexuan dhe miratuan dorëshkrimin përfundimtar për dorëzim.
Deklarata e Konfliktit të Interesit
Autorët deklarojnë se hulumtimi është kryer në mungesë të ndonjë marrëdhënie tregtare ose financiare që mund të interpretohet si një konflikt i mundshëm interesi.
Rishikuesit SC, SA dhe Editori i Menaxhimit deklaruan përkatësinë e tyre të përbashkët, dhe Administrimi i Menaxhimit thekson se procesi megjithatë ka përmbushur standardet e një rishikimi të drejtë dhe objektiv.
Mirënjohje
Kjo punë u mbështet nga financimi nga grantet nga Këshilli Australian i Kërkimeve (FT1110884) në SB dhe Këshilli Kombëtar i Shëndetit dhe Mjekësisë (1061979) në SB dhe MB.
Materiale plotësuese
Materiali plotësues për këtë artikull mund të gjendet online në: http://journal.frontiersin.org/article/10.3389/fnbeh.2016.00054
Figura plotësuese 1. Marrja e saharozës dhe preferencat nga 4 dhe 12 javë me sukroze që konsumojnë minjtë. (A, B) tregojnë përshkallëzim në marrjen totale të saharozës (ml) gjatë 4 dhe 12 javëve të ekspozimit. (C, D) tregojnë preferencë të lartë për sukrozën mbi ujë gjatë periudhave të prezantimit të saharozës.
Referencat
Ahmed, SH, dhe Koob, GF (1998). Tranzicioni nga marrja e medikamenteve në drogë: Ndryshimi në pikën hedonike. Shkencë 282, 298-300. doi: 10.1126 / science.282.5387.298
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Ahmed, SH, Lin, D., Koob, GF, dhe Parsons, LH (2003). Shkalla e vetë-administrimit të kokainës nuk varet nga nivelet e dopaminës së ndryshuar të kokainës të shkaktuar nga nucleus accumbens. J. Neurochem. 86, 102-113. doi: 10.1046 / j.1471-4159.2003.01833.x
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Ahmed, SH, Walker, JR, dhe Koob, GF (2000). Rritje e vazhdueshme në motivimin për të marrë heroinë në rats me një histori të eskalimit të drogës. Neuropsychopharmacology 22, 413–421. doi: 10.1016/S0893-133X(99)00133-5
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Arslanian, S. (2002). Lloji i diabetit tip 2 tek fëmijët: aspektet klinike dhe faktorët e rrezikut. Horm Res 57 (Suplement 1), 19-28. doi: 10.1159 / 000053308
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Avena, NM, Bocarsly, ME, Hoebel, BG dhe Gold, MS (2011). Mbivendosjet në nosologjinë e abuzimit të substancave dhe overeating: implikimet translational e "varësisë ushqim". Curr. Abuzimi i drogës Rev. 4, 133-139. doi: 10.2174 / 1874473711104030133
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Avena, NM, Rada, P., dhe Hoebel, BG (2008). Dëshmi për varësinë e sheqerit: efektet e sjelljes dhe neurokimi të marrjes së përzier të sheqerit me ndërprerje. Neurosci. Biobehav. Rev. 32, 20-39. doi: 10.1016 / j.neubiorev.2007.04.019
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Avena, NM, Rada, P., Moise, N., dhe Hoebel, BG (2006). Sucrose sham të ushqyerit në një orar programin lëshon accumbens dopamine në mënyrë të përsëritur dhe eliminon reagimin e ngopjes acetilkoline. Neuroscience 139, 813-820. doi: 10.1016 / j.neuroscience.2005.12.037
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Benton, D. (2010). Besueshmëria e varësisë së sheqerit dhe roli i saj në çrregullimet e obezitetit dhe të ngrënit. Clin. Nutr. 29, 288-303. doi: 10.1016 / j.clnu.2009.12.001
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Bray, GA, dhe Popkin, BM (2014). Sheqeri dietik dhe pesha e trupit: a kemi arritur një krizë në epideminë e obezitetit dhe diabetit ?: shëndetësor të mallkuar! Hidhni në sheqer. Kujdesi për diabet 37, 950-956. doi: 10.2337 / dc13-2085
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Brog, JS, Salyapongse, A., Deutch, AY dhe Zahm, DS (1993). Modelet e innervation afferent të bërthamë dhe shell në pjesën "accumbens" të striatum rat ventral: zbulimin imunohistochemical e floro-ari retrogradely transportohen. J. Comp. Neurol. 338, 255-278. doi: 10.1002 / cne.903380209
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Bucher Della Torre, S., Keller, A., Laure Depeyre, J., dhe Kruseman, M. (2015). Pijet me sheqer të ëmbël dhe rreziku i trashje në fëmijët dhe adoleshentët: një analizë sistematike se si cilësia metodologjike mund të ndikojë në konkluzione. J. Acad. Nutr. Dietë. [Epub para shtypjes]. doi: 10.1016 / j.jand.2015.05.020
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Carelli, RM dhe Perëndimi, EA (2014). Kur një shije e mirë kthehet e keqe: mekanizmat nervore që nënvizojnë shfaqjen e ndikimit negativ dhe zhvlerësimin e shpërblimit natyror që shoqërohet nga kokaina. Neuropharmacology 76 (Pt B), 360-369. doi: 10.1016 / j.neuropharm.2013.04.025
Chen, BT, Hopf, FW dhe Bonci, A. (2010). Plastikiteti synaptik në sistemin mesolimbik: implikime terapeutike për abuzimin e substancave. Ann. NY Acad. Sci. 1187, 129-139. doi: 10.1111 / j.1749-6632.2009.05154.x
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Colantuoni, C., Rada, P., McCarthy, J., Patten, C., Avena, NM, Chadeayne, A., et al. (2002). Dëshmia se futja e përhershme e tepërt e sheqerit shkakton varësi endogjene të opiumit. Obes. Res. 10, 478-488. doi: 10.1038 / oby.2002.66
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Colantuoni, C., Schwenker, J., McCarthy, J., Rada, P., Ladenheim, B., Cadet, JL, et al. (2001). Sasia e tepërt e sheqerit ndryshon në lidhje me receptorët dopamine dhe mu-opioid në tru. Neuroreport 12, 3549–3552. doi: 10.1097/00001756-200111160-00035
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Crombag, HS, Gorny, G., Li, Y., Kolb, B. dhe Robinson, TE (2005). Efektet e kundërta të vetë-administrimit të amfetaminës në spines dendriti në korteksin mesoral dhe orbital paraballor. Cereb. lëvore 15, 341-348. doi: 10.1093 / cercor / bhh136
Dabelea, D., Mayer-Davis, EJ, Saydah, S., Imperatore, G., Linder, B., Divers, J., et al. (2014). Prevalenca e tipit 1 dhe diabetit tip 2 tek fëmijët dhe adoleshentët nga 2001 në 2009. JAMA 311, 1778-1786. doi: 10.1001 / jama.2014.3201
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Davis, C. (2015). Epidemiologjia dhe gjenetika e çrregullimit të hahet (BED). CNS Spectr. 20, 522-529. doi: 10.1017 / s1092852915000462
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Ford, ES dhe Dietz, WH (2013). Trendet në marrjen e energjisë në mesin e të rriturve në Shtetet e Bashkuara: gjetjet nga NHANES. Jam. J. Clin. Nutr. 97, 848-853. doi: 10.3945 / ajcn.112.052662
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Fryar, CD, Carroll, MD, dhe Ogden, CL (2014). Prevalenca e mbipeshës, trashje dhe eshtra ekstreme midis të rriturve: Shtetet e Bashkuara, 1960-1962 përmes 2011-2012. Atlanta, GA: CDC.
Graveland, GA dhe DiFiglia, M. (1985). Frekuenca dhe shpërndarja e neuroneve të mesme me bërthama të pranishme në primat dhe neostriatum brejtës. Brain Res. 327, 307–311. doi: 10.1016/0006-8993(85)91524-0
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Groenewegen, HJ, Wright, CI, Beijer, Av, dhe Voorn, P. (1999). Konvergjenca dhe segregimi i hyrjeve dhe outputeve striatralë të barkut. Ann. NY Acad. Sci. 877, 49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Groves, Kryeministri (1980). Endings synaptic dhe objektivat e tyre postsynaptic në neostriatum: specializimet synaptic zbuluar nga analiza e seksioneve serial. Proc. Natl. Acad. Sci. SHBA 77, 6926-6929. doi: 10.1073 / pnas.77.11.6926
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Groves, PM, Linder, JC, dhe Young, SJ (1994). Axon dopaminergjik me etiketat 5-hidroksidopamine: rikonstruksione tre-dimensionale të aksoneve, synapave dhe objektivave postsinaptikë neostriatum të mijeve. Neuroscience 58, 593–604. doi: 10.1016/0306-4522(94)90084-1
Hu, FB (2013). I zgjidhur: ka prova të mjaftueshme shkencore që zvogëlimi i konsumit të pijeve me sheqer të ëmbël do të reduktojë prevalencën e obezitetit dhe sëmundjeve të lidhura me trashje. Obes. Rev. 14, 606-619. doi: 10.1111 / obr.12040
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Johnson, LR, Aylward, RL, Hussain, Z. dhe Totterdell, S. (1994). Input nga amygdala në nucleus accumbens miu: marrëdhëniet e saj me immunoreactivity tyrosine hydroxylase dhe neurons identifikuar. Neuroscience 61, 851–865. doi: 10.1016/0306-4522(94)90408-1
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Kaiya, H., dhe Namba, M. (1981). Dy lloje të terminaleve nervore dopaminergic në neostriatum miu. Një studim ultrastruktural. Neurosci. Lett. 25, 251–256. doi: 10.1016/0304-3940(81)90400-6
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Kalivas, PW dhe Miller, JS (1984). Neuronet e neurotensineve në zonën tegmentale të barkut projektojnë tek bërthama e mesme qendrore. Brain Res. 300, 157–160. doi: 10.1016/0006-8993(84)91351-9
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Kawaguchi, Y., Wilson, CJ, dhe Emson, PC (1990). Subtipet e projeksioneve të qelizave matrikale neostriatale të miellit zbulohen nga injeksioni intracellular i biocytinës. J. Neurosci. 10, 3421-3438.
Kelley, AE, dhe Swanson, CJ (1997). Ushqimi i shkaktuar nga bllokimi i receptorëve AMPA dhe kainat brenda striatumit të barkut: një studim i hartës së mikroinfuzionit. Behav. Brain Res. 89, 107–113. doi: 10.1016/S0166-4328(97)00054-5
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Kemp, JM dhe Powell, TP (1971). Organizimi synaptik i bërthamës caudate. Philos. Trans. R. Soc. Lond. B Biol. Sci. 262, 403-412. doi: 10.1098 / rstb.1971.0103
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Kenny, PJ (2011). Shpërblimi i mekanizmave në obezitet: njohuri të reja dhe drejtime të ardhshme. Neuron 69, 664-679. doi: 10.1016 / j.neuron.2011.02.016
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Kessler, RC, Berglund, PA, Chiu, WT, Deitz, AC, Hudson, JI, Shahly, V., et al. (2013). Përhapja dhe korrelatet e çrregullimit të hahet me qumësht në Sondazhet Botërore të Shëndetit Mendor të Organizatës Botërore të Shëndetësisë. Biol. psikiatri 73, 904-914. doi: 10.1016 / j.biopsych.2012.11.020
Klenowski, PM, Fogarty, MJ, Belmer, A., Noakes, PG, Bellingham, MC, dhe Bartlett, SE (2015). Karakterizim strukturor dhe funksional i arbors dendritic dhe inputeve GABAergic synaptic në interneurons dhe qelizat kryesore në amygdala basolateral miu. J. Neurophysiol. 114, 942-957. doi: 10.1152 / jn.00824.2014
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Lenoir, M., Serre, F., Cantin, L., dhe Ahmed, SH (2007). Ëmbëlsi e madhe tejkalon shpërblimin e kokainës. PLoS ONE 2: e698. doi: 10.1371 / journal.pone.0000698
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Li, Y., Kolb, B., dhe Robinson, TE (2003). Vendndodhja e ndryshimeve të vazhdueshme të amfetaminës në shkallën e dendësisë së spines dendriti në neuronet e mesme të shtyllës në nucleus accumbens dhe caudate-putamen. Neuropsychopharmacology 28, 1082-1085. doi: 10.1038 / sj.npp.1300115
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Lindvall, O., dhe Björklund, A. (1978). Anatomia e sistemeve neuron ne dopaminergic ne tru e miut. Adv. Biochem. Psychopharmacol. 19, 1-23.
Lutter, M., dhe Nestler, EJ (2009). Sinjalet homeostatike dhe hedonike bashkëveprojnë në rregullimin e marrjes së ushqimit. J. Nutr. 139, 629-632. doi: 10.3945 / jn.108.097618
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Maldonado-Irizarry, CS, Swanson, CJ dhe Kelley, AE (1995). Glutamat receptorët në bërthamën accumbens shell kontrollin e sjelljes së ushqyerit nëpërmjet hypothalamus lateral. J. Neurosci. 15, 6779-6788.
Malik, VS, Popkin, BM, Bray, GA, Després, JP, dhe Hu, FB (2010). Pijet e sheqerit, diabeti i tipit 2 dhe rreziku i sëmundjeve kardiovaskulare. Qarkullim 121, 1356-1364. doi: 10.1161 / CIRCULATIONAHA.109.876185
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Ng, SW, Slining, MM, dhe Popkin, BM (2012). Përdorimi i ëmbëlsirave kalorike dhe joalkalike në ushqimet e paketuara në SHBA, 2005-2009. J. Acad. Nutr. Dietë. 112, 1828-1834 e1821-e1826. doi: 10.1016 / j.jand.2012.07.009
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Nielsen, SJ, Siega-Riz, AM, dhe Popkin, BM (2002). Trendet në marrjen e energjisë në SHBA midis 1977 dhe 1996: ndërrime të ngjashme të shihet në të gjitha grupmoshat. Obes. Res. 10, 370-378. doi: 10.1038 / oby.2002.51
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Paxinos, G., dhe Watson, C. (2007). Truri i Ratit në Koordinatat Stereotoxike. Amsterdam; Boston, MA: Academic Press / Elsevier.
Popkin, BM (2010). Çfarë është e gabuar me qasjen e SHBA ndaj mbipeshes? Virtual Mentor 12, 316–320. doi: 10.1001/virtualmentor.2010.12.4.pfor2-1004
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Rada, P., Avena, NM dhe Hoebel, BG (2005). Bishtimi i përditshëm i sheqerit në mënyrë të përsëritur liron dopamine në guaskën e accumbens. Neuroscience 134, 737-744. doi: 10.1016 / j.neuroscience.2005.04.043
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Rafols, JA, Cheng, HW, dhe McNeill, TH (1989). Studimi Golgi i striatumit të miut: ndryshimet dendritike të moshës në popullatat e ndryshme neuronale. J. Comp. Neurol. 279, 212-227. doi: 10.1002 / cne.902790205
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Ranjan, A., dhe Mallick, BN (2010). Një metodë e modifikuar për ngjyrosje konsistente dhe të besueshme Golgi-cox në kohë të konsiderueshme të reduktuar. Front. Neurol. 1: 157. doi: 10.3389 / fneur.2010.00157
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Reinehr, T. (2013). Lloji 2 diabet mellitus në fëmijët dhe adoleshentët. World J. Diabetes 4, 270-281. doi: 10.4239 / wjd.v4.i6.270
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Reissner, KJ dhe Kalivas, PW (2010). Përdorimi i homeostazës së glutamatit si një objektiv për trajtimin e çrregullimeve të varësisë. Behav. Pharmacol. 21, 514–522. doi: 10.1097/FBP.0b013e32833d41b2
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Reynolds, SM dhe Berridge, KC (2003). Glutamate ansamblet motivues në nucleus accumbens: shkallët rostrocaudal shell e frikës dhe të ushqyerit. Eur. J. Neurosci. 17, 2187-2200. doi: 10.1046 / j.1460-9568.2003.02642.x
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Richard, JM dhe Berridge, KC (2011). Ndërveprimi Nucleus accumbens dopamine / glutamate ndërron mënyra për të gjeneruar dëshirën kundrejt tmerrit: D (1) vetëm për ngrënie të shijshme por D (1) dhe D (2) së bashku për shkak të frikës. J. Neurosci. 31, 12866-12879. doi: 10.1523 / JNEUROSCI.1339-11.2011
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Robinson, TE dhe Kolb, B. (1999). Ndryshime në morfologjinë e dendriteve dhe spines dendritike në bërthamë accumbens dhe korteksit paraballor pas trajtimit të përsëritur me amfetaminën ose kokainën. Eur. J. Neurosci. 11, 1598-1604. doi: 10.1046 / j.1460-9568.1999.00576.x
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Robinson, TE dhe Kolb, B. (2004). Plastikiteti strukturor i lidhur me ekspozimin ndaj drogave të abuzimit. Neuropharmacology 47 (Suplement 1), 33-46. doi: 10.1016 / j.neuropharm.2004.06.025
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Russo, SJ, Dietz, DM, Dumitriu, D., Morrison, JH, Malenka, RC, dhe Nestler, EJ (2010). Synapsi i varur: mekanizmat e plasticitetit synaptik dhe strukturor në nucleus accumbens. Trendet Neurosci. 33, 267-276. doi: 10.1016 / j.tins.2010.02.002
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Rutledge, LT, Duncan, J., dhe Beatty, N. (1969). Një studim i kolaterals aksonale të qelizave piramidale në korteksin cerebral të paprekur dhe pjesërisht të izoluar. Brain Res. 16, 15–22. doi: 10.1016/0006-8993(69)90082-1
Saper, CB, Chou, TC, dhe Elmquist, JK (2002). Nevoja për të ushqyer: kontrollin homeostatik dhe hedonik të ngrënies. Neuron 36, 199–211. doi: 10.1016/S0896-6273(02)00969-8
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Scheggi, S., Secci, ME, Marchese, G., De Montis, MG dhe Gambarana, C. (2013). Ndikimi i palatability në motivimin për të vepruar për ushqim kalorik dhe jo-kalori në jo-ushqimore-privuar dhe minjtë ushqimore-privuar. Neuroscience 236, 320-331. doi: 10.1016 / j.neuroscience.2013.01.027
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Sesack, SR, dhe Pickel, VM (1990). Në nucleus accumbens mediale të miut, terminalet hipokampale dhe katekolaminergjike konvergojnë në neuronet e mprehtë dhe janë në apozicion me njëri-tjetrin. Brain Res. 527, 266–279. doi: 10.1016/0006-8993(90)91146-8
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Sesack, SR, dhe Pickel, VM (1992). Efekte paralele cortical në synapse miu në shënjestrat neonale unlabeled të terminaleve catecholamine në nucleus accumbens septi dhe në neuronet dopamine në zonën tegmentale ventral. J. Comp. Neurol. 320, 145-160. doi: 10.1002 / cne.903200202
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Shariff, M., Quik, M., Holgate, JY, Morgan, M., Patkar, OL, Tam, V., et al. (në shtyp). Nivelet nikotinike të modulatorëve të receptorit të acetilkolinës zvogëlojnë marrjen e sheqerit. PLoS ONE.
Sheehan, DV, dhe Herman, BK (2015). Faktorët psikologjikë dhe mjekësorë të lidhur me çrregullimin e ngrënies që nuk trajtohen. Prim. Sjellja e kujdesit CNS Disord. 17. doi: 10.4088 / PCC.14r01732
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Simms, JA, Steensland, P., Medina, B., Abernathy, KE, Chandler, LJ, Wise, R., et al. (2008). Qasja e përhershme në etanol 20% shkakton konsum të lartë të etanolit në Long-Evans dhe Wistar rats. Alkooli. Clin. Exp. Res. 32, 1816-1823. doi: 10.1111 / j.1530-0277.2008.00753.x
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Steensland, P., Simms, JA, Holgate, J., Richards, JK dhe Bartlett, SE (2007). Vareniklin, një agonist i pjesshëm i receptorit acetilkolinik nikotinik alfa4beta2, zvogëlon selektivisht konsumin dhe kërkimin e etanolit. Proc. Natl. Acad. Sci. SHBA 104, 12518-12523. doi: 10.1073 / pnas.0705368104
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Stuber, GD, Hopf, FW, Hahn, J., Cho, SL, Guillory, A. dhe Bonci, A. (2008a). Marrja vullnetare e etanolit rrit fuqinë sinapsi excitatory në zonën tegmentale ventral. Alkooli. Clin. Exp. Res. 32, 1714-1720. doi: 10.1111 / j.1530-0277.2008.00749.x
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Stuber, GD, Klanker, M., de Ridder, B., Bowers, MS, Joosten, RN, Feenstra, MG, et al. (2008b). Cues shpërblimi-parashikuese rrisin fuqinë excitatory synaptic në neuronet dopamine të midbrain. Shkencë 321, 1690-1692. doi: 10.1126 / science.1160873
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Swanson, SA, Crow, SJ, Le Grange, D., Swendsen, J., dhe Merikangas, KR (2011). Përhapja dhe korrelatet e çrregullimeve të të ngrënit në adoleshentët. Rezultatet nga shtojca e adoleshencës së riprodhimit të studimit kombëtar të komorbiditetit. Arch. Gjenerali psikiatrik 68, 714-723. doi: 10.1001 / archgenpsychiatry.2011.22
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Te Morenga, L., Mallard, S., dhe Mann, J. (2013). Sheqernat dietike dhe pesha e trupit: rishikim sistematik dhe meta-analiza e testeve të kontrolluara me randomizim dhe studime kohorte. BMJ 346: e7492. doi: 10.1136 / bmj.e7492
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Totterdell, S., dhe Smith, AD (1989). Konvergjenca e inputit hippocampal dhe dopaminergjik mbi neuronet e identifikuara në nucleus accumbens të miut. J. Chem. Neuroanat. 2, 285-298.
Tukey, DS, Ferreira, JM, Antoine, SO, D'Amour, JA, Ninan, I., Cabeza de Vaca, S., et al. (2013) Gëlltitja e sakarozës shkakton trafik të shpejtë të receptorëve AMPA. J. Neurosci. 33, 6123-6132. doi: 10.1523 / JNEUROSCI.4806-12.2013
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Veening, JG, Cornelissen, FM, dhe Lieven, PA (1980). Organizimi topik i aferentëve të caudatoputamen të miut. Një studim i peroksidazës së rrikë. Neuroscience 5, 1253–1268. doi: 10.1016/0306-4522(80)90198-0
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Ventura, T., Santander, J., Torres, R., dhe Contreras, AM (2014). Baza neurobiologjike e dëshirës për karbohidratet. Ushqim 30, 252-256. doi: 10.1016 / j.nut.2013.06.010
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Volkow, ND, Wang, GJ, Fowler, JS, Tomasi, D., dhe Baler, R. (2012). Ushqimi dhe shpërblimi i drogës: qarqet mbivendosëse në mbipeshën dhe varësinë njerëzore. Curr. Top. Behav. Neurosci. 11, 1–24. doi: 10.1007/7854_2011_169
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
I mençur, RA (1973). Futje vullnetare e etanolit në minjtë pas ekspozimit ndaj etanolit në orare të ndryshme. Psychopharmacologia 29, 203-210. doi: 10.1007 / BF00414034
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Wright, CI, dhe Groenewegen, HJ (1995). Modele të konvergjencës dhe segregacionit në bërthamën qendrore të akumulimit të miut: marrëdhëniet e aferentëve paralëndorë kortikalë, midal-lineal dhe aferentët amygdaloid bazal. J. Comp. Neurol. 361, 383-403. doi: 10.1002 / cne.903610304
PubMed Abstract | Teksti i plotë i CrossRef | Google Scholar
Fjalë kyçe: konsumi i ngjashëm me kërpudha, termi afatgjatë, neuron i mesëm, nucleus accumbens, saharoze
Citimi: Kryeministri i Klenowski, Shariff MR, Belmer A, Fogarty MJ, Mu EWH, Bellingham MC dhe Bartlett SE (2016) Konsumimi i zgjatur i saharozës në një mënyrë të ngjashme me tym, ndryshon morfologjinë e Neuroneve të Mesme Spiny në Shell Nucleus Accumbens. Front. Behav. Neurosci. 10: 54. doi: 10.3389 / fnbeh.2016.00054
Marrësi: 03 Dhjetor 2015; Pranohet: 07 Mars 2016;
Publikuar: 23 Mars 2016.
E Redaktuar nga:
Djoher Nora Abrous, Institut des Neurosciences de Bordeaux, Francë
Shqyrtuar nga:
Serge H. Ahmed, Qendra Kombëtare de la Recherche Scientifique, Francë
Stéphanie Caille, Qendra Kombëtare de la Recherche Scientifique, Francë
Copyright © 2016 Klenowski, Shariff, Belmer, Fogarty, Mu, Bellingham dhe Bartlett. Ky është një artikull me akses të hapur të shpërndarë nën kushtet e Licenca e Attribution Creative Commons (CC BY). Përdorimi, shpërndarja ose riprodhimi në forume të tjera lejohet, me kusht që autorët ose licencuesit origjinale të kreditohen dhe se publikohet botimi origjinal në këtë ditar, në përputhje me praktikat e pranuara akademike. Nuk lejohet përdorimi, shpërndarja ose riprodhimi i cili nuk përputhet me këto terma.
* Korrespondenca: Selena E. Bartlett, [email mbrojtur]